Microbial Community and Fermentation Dynamics of Corn Silage Prepared with Heat-Resistant Lactic Acid Bacteria in a Hot Environment

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Forage and Ensiling
2.2. Chemical, Fermentation and Microbial Counts Analysis
2.3. Bacterial Community Analysis
2.3.1. DNA Extraction
2.3.2. Sequencing
2.3.3. Sequences Analyses
2.4. Statistical Analyses
3. Results
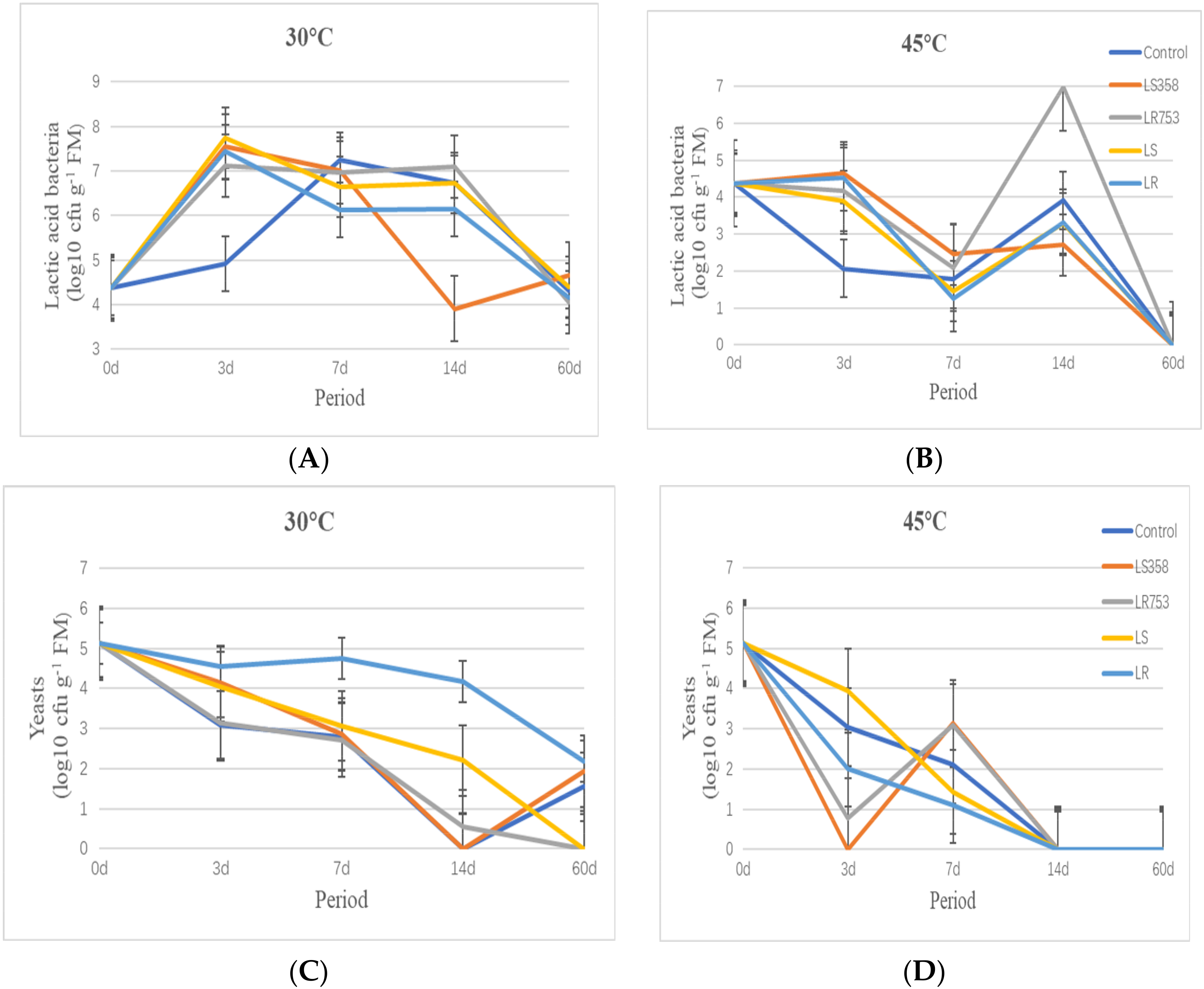
3.1. Fermentation Characteristics and Microbial Counts of Silage at Different Period
3.2. Chemical Composition of Corn Silages Ensiled for 60 d
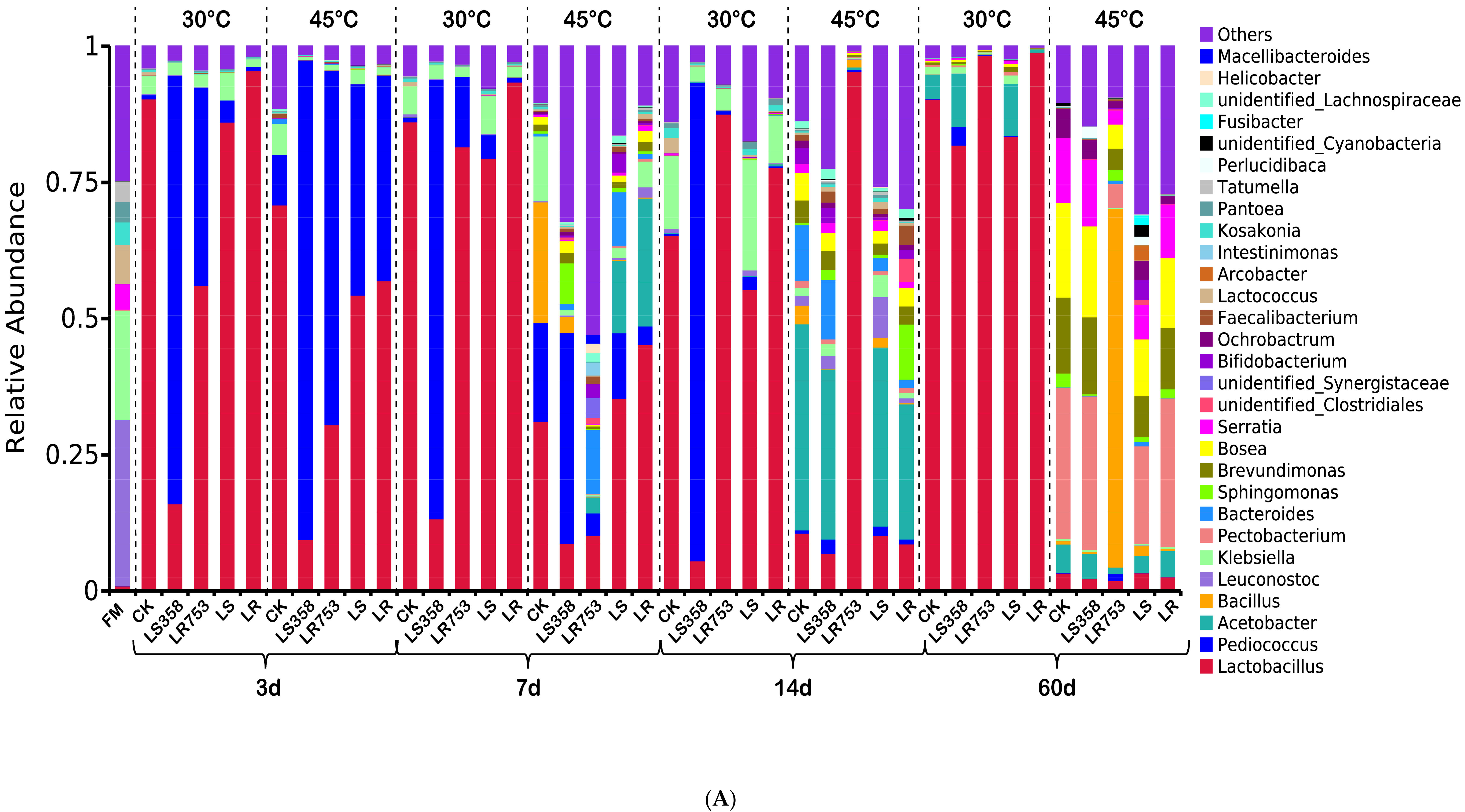
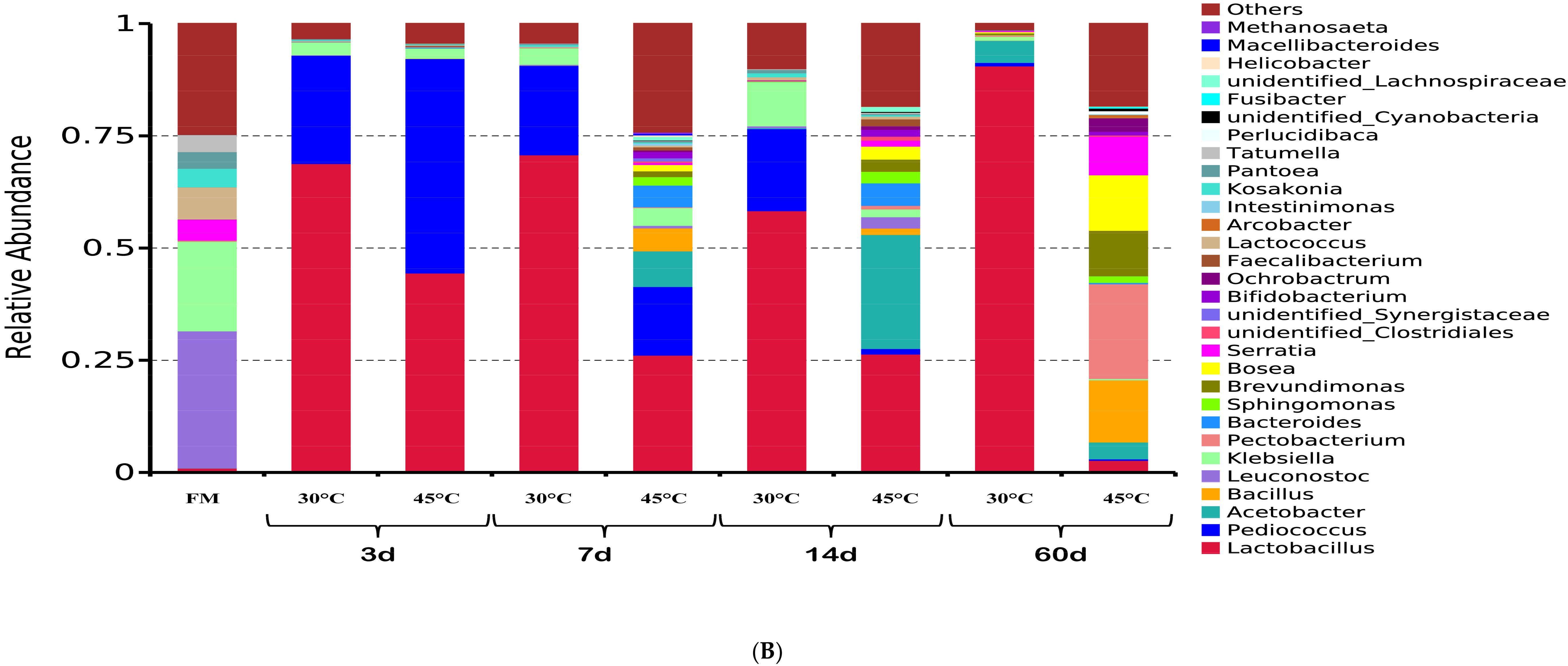
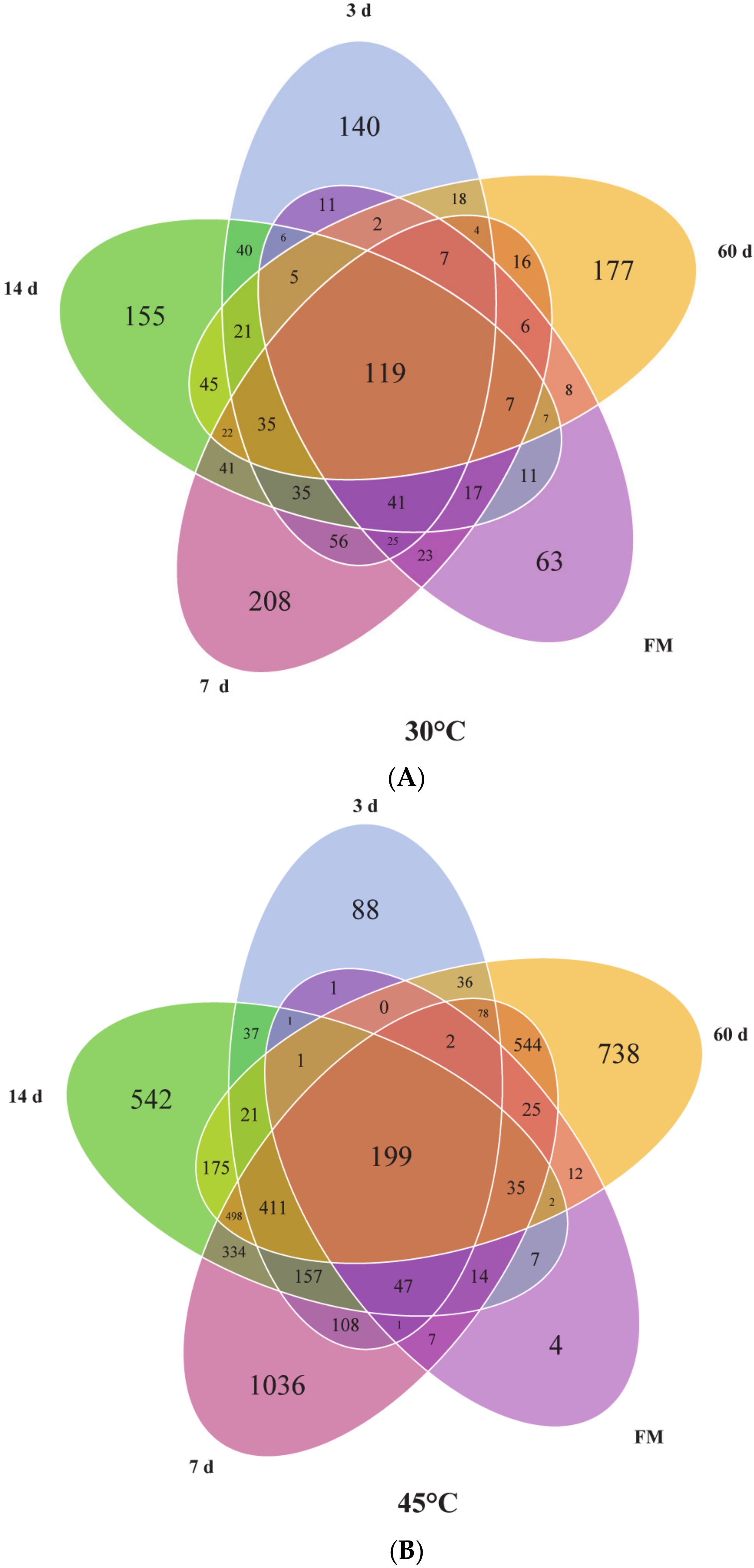
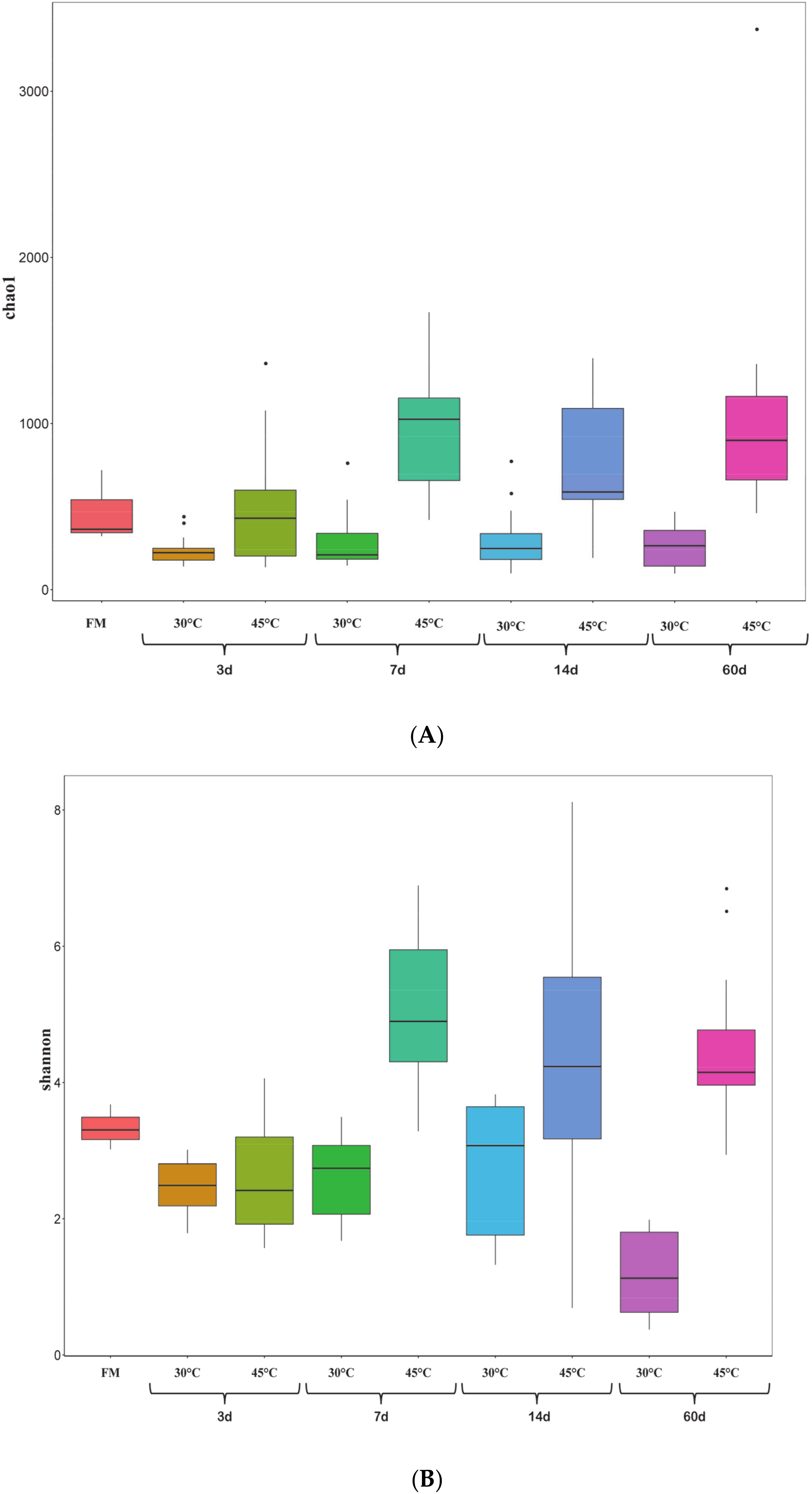
3.3. Dynamic Changes of Microbial Communities
4. Discussion
4.1. Dynamic Changes of Silage Fermentation Characteristics and Microbial Composition
4.2. Dynamic Changes of Microbial Communities
4.3. Chemical Composition of Corn Silages Ensiled for 60 d
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wilkinson, J.M.; Bolsen, K.K.; Lin, C.J. History of Silage. Silage Sci. Technol. 2003. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, T.F.; Daniel, J.L.P.; Adesogan, A.T.; McAllister, T.A.; Drouin, P.; Nussio, L.G.; Huhtanen, P.; Tremblay, G.F.; Belanger, G.; Cai, Y. Silage review: Unique challenges of silages made in hot and cold regions. J. Dairy Sci. 2018, 101, 4001–4019. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, Z.G.; Muck, R. New trends and opportunities in the development and use of inoculants for silage. FEMS Microbiol. Rev. 1996, 19, 53–68. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biotechnol. 2018, 102, 4025–4037. [Google Scholar] [CrossRef] [PubMed]
- Elferink, S.O.; Driehuis, F.; Krooneman, J.; Gottschal, J.; Spoelstra, S. Lactobacillus buchneri can improve the aerobic stability of silage via a novel fermentation pathway: The anaerobic degradation of lactic acid to acetic acid and 1, 2-propanediol. In Proceedings of the 12th International Silage Conference (ISC 1999), Uppsala, Sweden, 5–7 July 1999; pp. 266–267. [Google Scholar]
- Reich, L.J.; Kung, L., Jr. Effects of combining Lactobacillus buchneri 40788 with various lactic acid bacteria on the fermentation and aerobic stability of corn silage. Anim. Feed Sci. Technol. 2010, 159, 105–109. [Google Scholar] [CrossRef]
- Whiter, A.G.; Kung, L., Jr. The effect of a dry or liquid application of Lactobacillus plantarum MTD1 on the fermentation of alfalfa silage. J. Dairy Sci. 2001, 84, 2195–2202. [Google Scholar] [CrossRef]
- Chen, M.M.; Liu, Q.H.; Xin, G.R.; Zhang, J.G. Characteristics of lactic acid bacteria isolates and their inoculating effects on the silage fermentation at high temperature. Lett. Appl. Microbiol. 2013, 56, 71–78. [Google Scholar] [CrossRef]
- Gulfam, A.; Guo, G.; Tajebe, S.; Chen, L.; Liu, Q.; Yuan, X.; Bai, Y.; Saho, T. Characteristics of lactic acid bacteria isolates and their effect on the fermentation quality of Napier grass silage at three high temperatures. J. Sci. Food Agric. 2017, 97, 1931–1938. [Google Scholar] [CrossRef]
- Guan, H.; Ke, W.; Yan, Y.; Shuai, Y.; Li, X.; Ran, Q.; Yang, Z.; Wang, X.; Cai, Y.; Zhang, X. Screening of natural lactic acid bacteria with potential effect on silage fermentation, aerobic stability and aflatoxin B1 in hot and humid area. J. Appl. Microbiol. 2020, 128, 1301–1311. [Google Scholar] [CrossRef]
- Muck, R. Recent advances in silage microbiology. Agric. Food Sci. 2013, 22, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Duniere, L.; Xu, S.; Long, J.; Elekwachi, C.; Wang, Y.; Turkington, K.; Forster, R.; McAllister, T.A. Bacterial and fungal core microbiomes associated with small grain silages during ensiling and aerobic spoilage. BMC Microbiol. 2017, 17, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Li, X.; Guan, H.; Huang, L.; Ma, X.; Peng, Y.; Li, Z.; Nie, G.; Zhou, J.; Yang, W.; et al. Microbial community and fermentation characteristic of Italian ryegrass silage prepared with corn stover and lactic acid bacteria. Bioresour. Technol. 2019, 279, 166–173. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 2002, 39, 971–974. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, J.M.; Muck, R.E. Ensiling in 2050: Some challenges and opportunities. Grass Forage Sci. 2019, 74, 178–187. [Google Scholar] [CrossRef]
- Wieringa, G. Some factors affecting silage fermentation. In Proceedings of the 8th International Grassland Congress (IGC 1960), Wageningen, The Netherlands, 11–21 July 1960; p. 6. [Google Scholar]
- Kung Jr, L.; Shaver, R.; Grant, R.; Schmidt, R. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Muck, R. A lactic acid bacteria strain to improve aerobic stability of silages. Res. Summ. 1996, 42–43. Available online: https://www.ars.usda.gov/ARSUserFiles/50901500/research_summaries/RS96_pdfs/RS96-20.pdf (accessed on 9 May 2020).
- Driehuis, F.; Oude Elferink, S.; Van Wikselaar, P. Fermentation characteristics and aerobic stability of grass silage inoculated with Lactobacillus buchneri, with or without homofermentative lactic acid bacteria. Grass Forage Sci. 2001, 56, 330–343. [Google Scholar] [CrossRef]
- Filya, I. The effect of Lactobacillus buchneri and Lactobacillus plantarum on the fermentation, aerobic stability, and ruminal degradability of low dry matter corn and sorghum silages. J. Dairy Sci. 2003, 86, 3575–3581. [Google Scholar] [CrossRef] [Green Version]
- Kleinschmit, D.H.; Kung, L., Jr. A meta-analysis of the effects of Lactobacillus buchneri on the fermentation and aerobic stability of corn and grass and small-grain silages. J. Dairy Sci. 2006, 89, 4005–4013. [Google Scholar] [CrossRef]
- Muck, R.E. Effects of corn silage inoculants on aerobic stability. In Proceedings of the 2002 ASAE Annual Meeting, St. Joseph, MI, USA, 28–31 July 2002; p. 1. Available online: https://elibrary.asabe.org/abstract.asp?aid=9144 (accessed on 9 May 2020).
- Kleinschmit, D.; Schmidt, R.; Kung, L., Jr. The effects of various antifungal additives on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2005, 88, 2130–2139. [Google Scholar] [CrossRef] [Green Version]
- Filya, I.; Sucu, E.; Karabulut, A. The effect of Lactobacillus buchneri on the fermentation, aerobic stability and ruminal degradability of maize silage. J. Appl. Microbiol. 2006, 101, 1216–1223. [Google Scholar] [CrossRef]
- Ranjit, N.; Taylor, C.; Kung Jr, L. Effect of Lactobacillus buchneri 40788 on the fermentation, aerobic stability and nutritive value of maize silage. Grass Forage Sci. 2002, 57, 73–81. [Google Scholar] [CrossRef]
- McAllister, T.; Duniere, L.; Drouin, P.; Xu, S.; Wang, Y.; Munns, K.; Zaheer, R. Silage review: Using molecular approaches to define the microbial ecology of silage. J. Dairy Sci. 2018, 101, 4060–4074. [Google Scholar] [CrossRef] [Green Version]
- Drouin, P.; Tremblay, J.; Chaucheyras-Durand, F. Dynamic Succession of Microbiota during Ensiling of Whole Plant Corn Following Inoculation with Lactobacillus buchneri and Lactobacillus hilgardii Alone or in Combination. Microorganisms 2019, 7, 595. [Google Scholar] [CrossRef] [Green Version]
- Aakko, J.; Sánchez, B.; Gueimonde, M.; Salminen, S. Assessment of stress tolerance acquisition in the heat-tolerant derivative strains of Bifidobacterium animalis subsp. lactis BB-12 and Lactobacillus rhamnosus GG. J. Appl. Microbiol. 2014, 117, 239–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, D.V.H. The aerobic deterioration of grass silage and its effect on the water-soluble carbohydrate and the associated heat production. J. Sci. Food Agric. 1982, 33, 499–508. [Google Scholar] [CrossRef]
- Kim, S.C.; Adesogan, A.T. Influence of ensiling temperature, simulated rainfall, and delayed sealing on fermentation characteristics and aerobic stability of corn silage. J. Dairy Sci 2006, 89, 3122–3132. [Google Scholar] [CrossRef]
- Colombatto, D.; Mould, F.L.; Bhat, M.K.; Phipps, R.H.; Owen, E. In vitro evaluation of fibrolytic enzymes as additives for maize (Zea mays L.) silage: I. Effects of ensiling temperature, enzyme source and addition level. Anim. Feed Sci. Technol. 2004, 111, 111–128. [Google Scholar] [CrossRef]
- Wilson, J.; Deinum, B.; Engels, F. Temperature effects on anatomy and digestibility of leaf and stem of tropical and temperate forage species. Neth. J. Agric. Sci. 1991, 39, 31–48. [Google Scholar] [CrossRef]
- Buxton, D.R. Quality-related characteristics of forages as influenced by plant environment and agronomic factors. Anim. Feed Sci. Technol. 1996, 59, 37–49. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Item | 30 °C | 45 °C | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | LS358 | LR753 | LS | LR | Control | LS358 | LR753 | LS | LR | SEM | Temperature | Inoculants | Interaction | |
| 3 d | ||||||||||||||
| pH | 3.91 d | 3.88 de | 3.87 e | 3.86 e | 3.81 f | 4.02 b | 4.02 bc | 3.98 c | 4.10 a | 4.00 bc | 0.018 | ** | ** | ** |
| NH3-N (g kg−1 TN) | 23.82 ab | 15.08 de | 13.88 e | 17.94 cde | 17.89 cde | 25.04 a | 19.47 bcd | 15.15 de | 15.38 de | 20.64 bc | 1.458 | NS | ** | NS |
| LA (g kg−1 DM) | 60.94 abc | 47.43 cd | 41.46 d | 74.12 a | 73.14 a | 50.86 bcd | 54.61 bcd | 60.93 abc | 65.07 ab | 64.88 ab | 6.357 | ** | NS | NS |
| AA (g kg−1 DM) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - | - | - | - |
| LA/AA | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - | - | - | - |
| 7 d | ||||||||||||||
| pH | 3.94 e | 3.94 e | 3.92 e | 3.93 e | 3.93 e | 4.19 a | 4.07 cd | 4.10 bc | 4.15 b | 4.02 d | 0.022 | ** | ** | ** |
| NH3-N (g kg−1 TN) | 25.80 a | 25.57 a | 24.40 abc | 24.84 abc | 22.50 bcde | 25.38 ab | 22.05 cde | 20.50 e | 21.06 de | 20.24 e | 1.073 | ** | * | NS |
| LA (g kg−1 DM) | 63.78 cde | 69.37 cde | 75.07 cd | 75.01 cd | 94.18 a | 88.35 ab | 78.83 bc | 63.13 e | 67.23 cde | 71.94 cde | 3.609 | ** | NS | ** |
| AA (g kg−1 DM) | 32.18 cd | 40.15 b | 55.87 a | 33.49 bcd | 37.93 bc | 0 | 30.26 d | 28.76 d | 0 | 0 | 2.289 | ** | ** | ** |
| LA/AA | 1.99 b | 1.73 bc | 1.34 c | 1.82 bc | 1.77 bc | 0 | 2.76 a | 2.21 b | 0 | 0 | 0.168 | ** | ** | ** |
| 14 d | ||||||||||||||
| pH | 3.92 c | 3.92 c | 3.90 c | 3.90 c | 3.86 c | 4.28 a | 3.91 c | 3.82 c | 4.04 b | 4.02 b | 0.028 | ** | ** | ** |
| NH3-N (g kg−1 TN) | 32.01 ab | 29.60 bcd | 24.87 d | 27.53 bcd | 25.98 d | 36.02 a | 26.87 cd | 25.47 d | 31.41 bc | 27.95 bcd | 0.451 | NS | ** | NS |
| LA (g kg−1 DM) | 61.16 d | 70.53 bcd | 70.82 bcd | 58.93 d | 102.06 a | 71.73 bcd | 86.43 abc | 87.03 ab | 85.57 abc | 65.07 cd | 6.478 | NS | NS | ** |
| AA (g kg−1 DM) | 31.83 d | 47.52 b | 61.22 a | 38.97 c | 31.09 d | 25.85 d | 24.85 d | 28.43 d | 0 | 25.77 d | 2.379 | ** | ** | ** |
| LA/AA | 1.95 bc | 1.48 c | 1.15 c | 1.51 c | 3.38 a | 3.38 a | 3.47 a | 2.54 ab | 0 | 2.63 ab | 0.299 | * | ** | ** |
| 60 d | ||||||||||||||
| pH | 3.84 de | 3.85 de | 3.85 de | 3.81 e | 3.87 cd | 4.06 a | 3.96 b | 3.91 c | 4.00 b | 3.91 c | 0.015 | ** | ** | ** |
| NH3-N (g kg−1 TN) | 42.58 bc | 36.56 d | 25.81 e | 36.78 d | 30.03 e | 47.75 a | 46.88 ab | 37.77 cd | 40.62 cd | 39.29 cd | 1.632 | ** | ** | ** |
| LA (g kg−1 DM) | 68.68 bcd | 66.72 cd | 63.01 de | 51.50 e | 72.23 abcd | 83.39 a | 84.63 a | 80.06 ab | 77.77 abc | 66.45 cd | 4.00 | ** | NS | ** |
| AA (g kg−1 DM) | 30.98 bc | 35.88 ab | 47.01 a | 21.73 cd | 35.56 ab | 18.43 d | 17.46 d | 33.40 bc | 21.28 cd | 31.17 bc | 3.81 | ** | ** | * |
| LA/AA | 2.21 bc | 1.88 bc | 1.34 bc | 2.43 bc | 2.73 b | 4.53 a | 4.85 a | 2.40 bc | 4.06 a | 1.93 bc | 0.403 | ** | ** | ** |
Item | 30 °C | 45 °C | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | LS358 | LR753 | LS | LR | Control | LS358 | LR753 | LS | LR | SEM | Temperature | Inoculants | Interaction | |
| DMR% | 97.03 a | 96.24 a | 96.53 a | 96.61 a | 96.19 a | 82.22 b | 82.11 b | 84.13 b | 82.85 b | 84.51 b | 1.649 | ** | NS | NS |
| CP g kg−1 DM | 69.16 c | 73.54 b | 64.37 e | 66.66 d | 75.62 a | 69.37 c | 67.08 d | 63.64 e | 56.70 f | 64.58 e | 0.922 | ** | ** | ** |
| WSC g kg−1 DM | 15.47 e | 18.64 e | 14.85 e | 20.65 e | 55.89 d | 140.47 a | 119.40 bc | 108.48 c | 119.73 bc | 123.75 b | 5.767 | ** | ** | ** |
| NDF g kg−1 DM | 446.14 ab | 450.77 ab | 509.23 a | 428.86 b | 484.98 ab | 443.22 ab | 434.32 b | 436.83 b | 454.70 ab | 437.04 b | 1.857 | ** | NS | * |
| ADF g kg−1 DM | 201.29 ab | 208.39 abc | 231.09 a | 184.66 c | 220.71 ab | 211.35 abc | 198.67 bc | 183.05 c | 204.17 abc | 223.21 ab | 1.303 | NS | NS | * |
| Period | Factor | Anosim | Adonis | ||||||
|---|---|---|---|---|---|---|---|---|---|
| R-Value | p-Value | Df | SumsOfSqs | MeanSqs | F.Model | R2 | Pr(>F) | ||
| 3 d | Temperature | 0.5163 | 0.001 | 1(28) | 2.2129(5.4017) | 2.21285(0.19292) | 11.47 | 0.29061(0.70939) | 0.001 |
| Inoculants | 0.3225 | 0.007 | 1(28) | 1.1866(6.4279) | 1.18665(0.22957) | 5.1691 | 0.15584(0.84416) | 0.004 | |
| 7 d | Temperature | 0.3763 | 0.002 | 1(28) | 1.7292(7.2746) | 1.72916(0.25981) | 6.6556 | 0.19205(0.80795) | 0.001 |
| Inoculants | −0.1115 | 0.871 | 1(28) | 0.5382(8.4662) | 0.53816(0.30236) | 1.7799 | 0.05977(0.94023) | 0.11 | |
| 14 d | Temperature | 0.4865 | 0.001 | 1(28) | 2.3373(7.7235) | 2.33728(0.27584) | 8.4733 | 0.23231(0.76769) | 0.001 |
| Inoculants | −0.1529 | 0.916 | 1(28) | 0.2902(9.7711) | 0.29017(0.34897) | 0.8315 | 0.02884(0.97116) | 0.575 | |
| 60 d | Temperature | 0.999 | 0.001 | 1(28) | 5.3947(2.2199) | 5.3947(0.0793) | 68.045 | 0.70847(0.29153) | 0.001 |
| Inoculants | −0.1173 | 0.974 | 1(28) | 0.0829(7.5319) | 0.082906(0.268995) | 0.30821 | 0.01089(0.98911) | 0.752 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, H.; Shuai, Y.; Yan, Y.; Ran, Q.; Wang, X.; Li, D.; Cai, Y.; Zhang, X. Microbial Community and Fermentation Dynamics of Corn Silage Prepared with Heat-Resistant Lactic Acid Bacteria in a Hot Environment. Microorganisms 2020, 8, 719. https://doi.org/10.3390/microorganisms8050719
Guan H, Shuai Y, Yan Y, Ran Q, Wang X, Li D, Cai Y, Zhang X. Microbial Community and Fermentation Dynamics of Corn Silage Prepared with Heat-Resistant Lactic Acid Bacteria in a Hot Environment. Microorganisms. 2020; 8(5):719. https://doi.org/10.3390/microorganisms8050719
Chicago/Turabian StyleGuan, Hao, Yang Shuai, Yanhong Yan, Qifan Ran, Xia Wang, Dandan Li, Yimin Cai, and Xinquan Zhang. 2020. "Microbial Community and Fermentation Dynamics of Corn Silage Prepared with Heat-Resistant Lactic Acid Bacteria in a Hot Environment" Microorganisms 8, no. 5: 719. https://doi.org/10.3390/microorganisms8050719