Antimicrobial and Fermentation Potential of Himanthalia elongata in Food Applications


 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Standards and Reagents
2.3. Bacterial Strains Used for Fermentation
2.4. Set-Up of Fermentation Conditions and Monitoring
2.5. High-Pressure Processing (HPP)
2.6. Extraction Process
2.7. Pathogenic Strains
2.8. Evaluation of Antimicrobial Activity In Vitro
2.9. Total Phenolic Content
2.10. UHPLC-ESI-MS2 Analysis
2.11. Statistical Analyses
3. Results and Discussion
3.1. Himanthalia elongata Fermentation
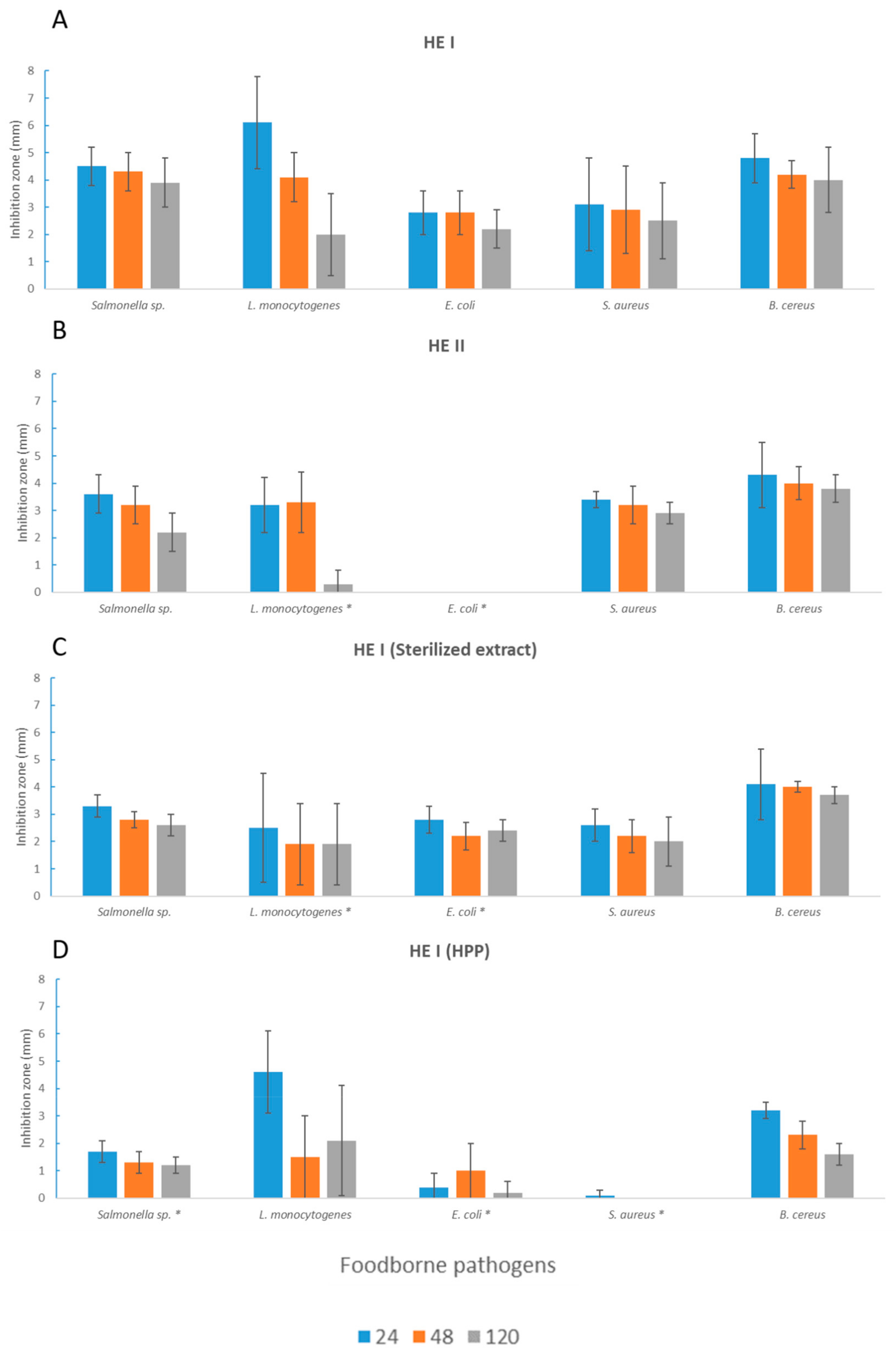
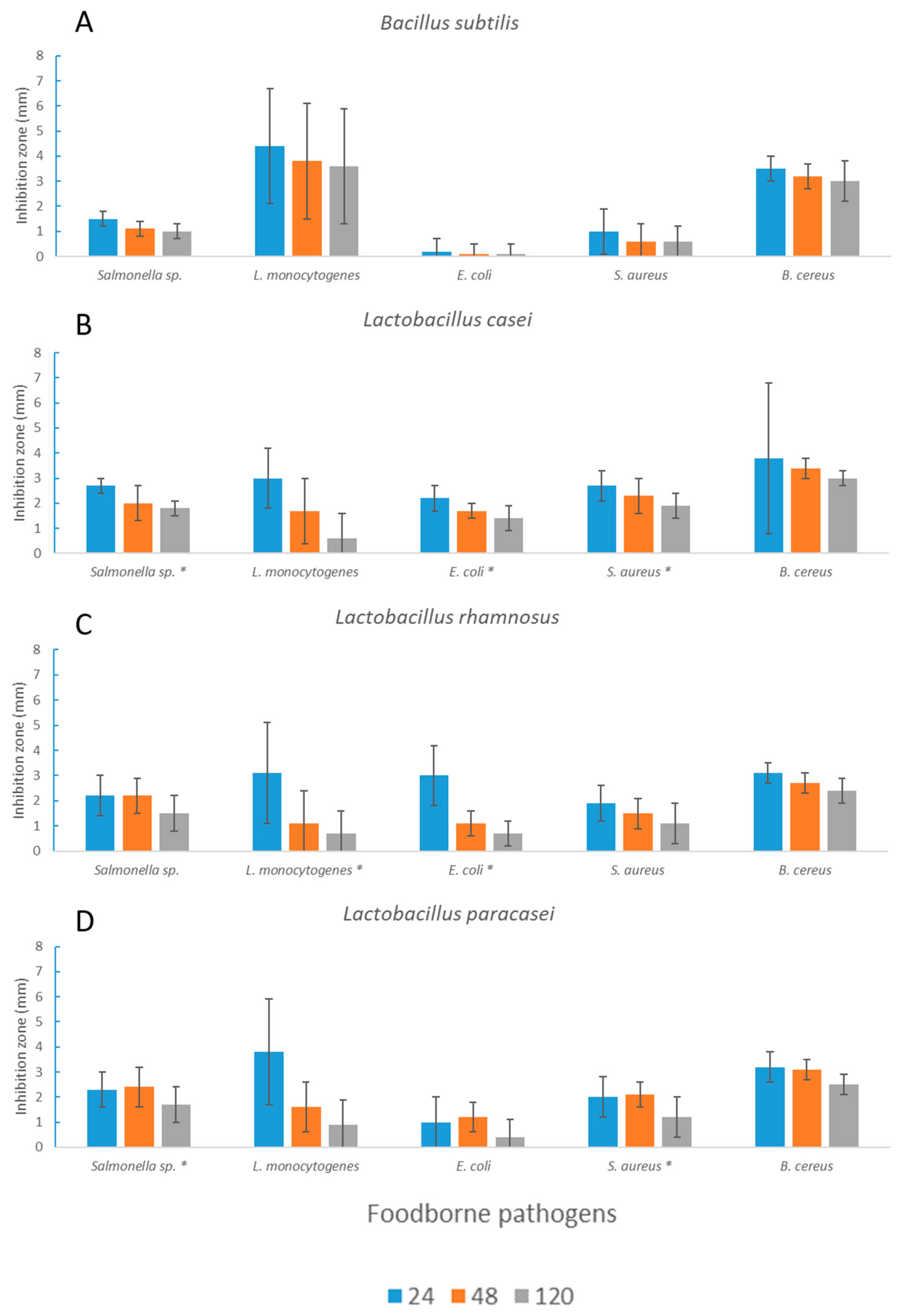
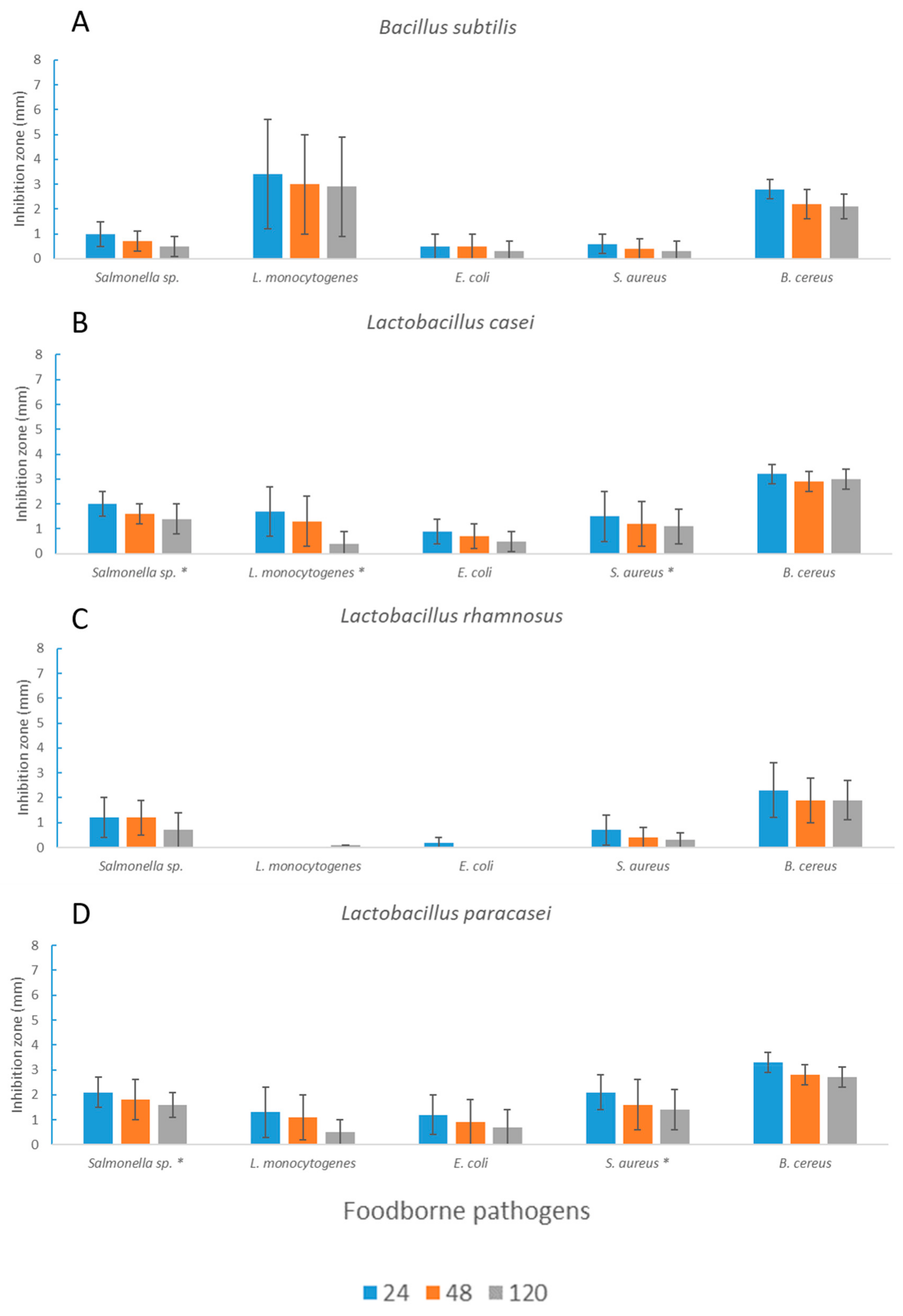
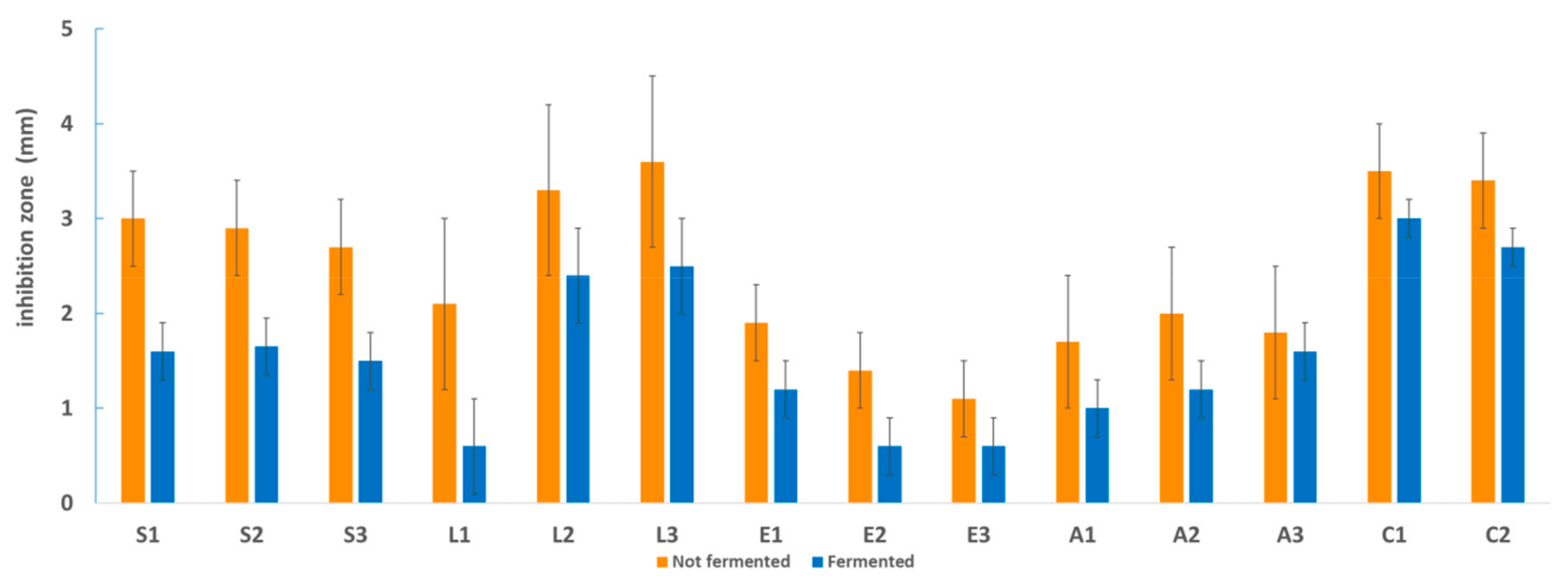
3.2. Antimicrobial Activity toward Foodborne Pathogens
3.3. Total Phenolic Content
3.4. Phlorotannin Identification by UHPLC-MS2 Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Control 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Zbakh, H.; Chiheb, H.; Bouziane, H.; Sánchez, V.M.; Riadi, H. Antibacterial activity of benthic marine algae extracts from the Mediterranean coast of Morocco. J. Microbiol. Biotechnol. Food Sci. 2012, 2, 219–228. [Google Scholar]
- Hongayo, M.C.; Larino, R.C.; Malingin, D.L. Antibacterial and Antioxidant Effects of Brown Alga Padina australis Hauck Crude Extract. IAMURE Int. J. Sci. Clin. Lab. 2012, 2, 13. [Google Scholar] [CrossRef]
- Alghazeer, R.; Whida, F.; Abduelrhman, E.; Gammoudi, F.; Azwai, S. Screening of antibacterial activity in marine green, red and brown macroalgae from the western coast of Libya. Nat. Sci. 2013, 5, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Saleh, B.; Al-Mariri, A. Antimicrobial Activity of the Marine Algal Extracts against Selected Pathogens. J. Agr. Sci. Tech. 2017, 19, 1067–1077. [Google Scholar]
- Watson, S.B.; Cruz-Rivera, E. Algal chemical ecology: An introduction to the special issue. Phycologia 2003, 42, 319–323. [Google Scholar] [CrossRef] [Green Version]
- Heo, S.; Park, E.; Lee, K.; Jeon, Y. Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour. Technol. 2005, 96, 1613–1623. [Google Scholar] [CrossRef]
- Chandini, S.K.; Ganesan, P.; Bhaskar, N. In vitro antioxidant activities of three selected brown seaweeds of India. Food Chem. 2008, 107, 707–713. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Hintz, T.; Matthews, K.K.; Di, R. The Use of Plant Antimicrobial Compounds for Food Preservation. BioMed Res. Int. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chai, T.-T.; Tan, Y.-N.; Ee, K.-Y.; Xiao, J.; Wong, F.-C. Seeds, fermented foods, and agricultural by-products as sources of plant-derived antibacterial peptides. Crit. Rev. Food Sci. Nutr. 2019, 59, 162–177. [Google Scholar] [CrossRef] [PubMed]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as biopreservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- Ennouali, M.; Ouhssine, M.; Ouhssine, K.; Elyachioui, M. Biotransformation of algal waste by biological fermentation. Afr. J. Biotechnol. 2006, 5, 1233–1237. [Google Scholar]
- Uchida, M.; Murata, M. Fermentative preparation of single cell detritus from seaweed, Undaria pinnatifida, suitable as a replacement hatchery diet for unicellular algae. Aquaculture 2002, 207, 345–357. [Google Scholar] [CrossRef]
- Prachyakij, P.; Charernjiratrakul, W.; Kantachote, D. Improvement in the quality of a fermented seaweed beverage using an antiyeast starter of Lactobacillus plantarum DW3 and partial sterilization. World J. Microbiol. Biotechnol. 2008, 24, 1713–1720. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N.; Scannell, A.G.M. Growth and kinetics of Lactobacillus plantarum in the fermentation of edible Irish brown seaweeds. Food Bioprod. Process. 2011, 89, 346–355. [Google Scholar] [CrossRef] [Green Version]
- Plaza, M.; Santoyo, S.; Jaime, L.; García-Blairsy Reina, G.; Herrero, M.; Señoráns, F.J.; Ibáñez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef]
- Eom, S.-H.; Kim, Y.-M.; Kim, S.-K. Antimicrobial effect of phlorotannins from marine brown algae. Food Chem. Toxicol. 2012, 50, 3251–3255. [Google Scholar] [CrossRef]
- Michell, C.; Lahaye, M.; Bonnet, C.; Mabeau1, S.; Barry, J.-L. In vitro fermentation by human faecal bacteria of total and purified dietary fibres from brown seaweeds. Br. J. Nutr. 1996, 75, 263–280. [Google Scholar] [CrossRef] [Green Version]
- Jun, X. Micromechanism of ultrahigh pressure extraction of active ingredients from green tea leaves. Food Control. 2011, 22, 1473–1476. [Google Scholar] [CrossRef]
- Jun, X. High-Pressure Processing as Emergent Technology for the Extraction of Bioactive Ingredients From Plant Materials. Crit. Rev. Food Sci. Nutr. 2013, 53, 837–852. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Hamilton Turley, G.; Rajauria, G.; Abu-Ghannam, N.; Jaiswal, A.K. Antioxidant potential and antimicrobial efficacy of seaweed (Himanthalia elongata) extract in model food systems. J. Appl. Phycol. 2014, 26, 1823–1831. [Google Scholar] [CrossRef]
- Kim, S.-J.; Cho, A.R.; Han, J. Antioxidant and antimicrobial activities of leafy green vegetable extracts and their applications to meat product preservation. Food Control. 2013, 29, 112–120. [Google Scholar] [CrossRef]
- Medina-Remón, A.; Barrionuevo-González, A.; Zamora-Ros, R.; Andres-Lacueva, C.; Estruch, R.; Martínez-González, M.-Á.; Diez-Espino, J.; Lamuela-Raventos, R.M. Rapid Folin–Ciocalteu method using microtiter 96-well plate cartridges for solid phase extraction to assess urinary total phenolic compounds, as a biomarker of total polyphenols intake. Anal. Chim. Acta 2009, 634, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Mena, P.; Martí, N.; Saura, D.; Valero, M.; García-Viguera, C. Combinatory Effect of Thermal Treatment and Blending on the Quality of Pomegranate Juices. Food Bioprocess. Technol. 2013, 6, 3186–3199. [Google Scholar] [CrossRef]
- R. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 16 October 2019).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Lee, S.-J.; Lee, D.-G.; Park, S.-H.; Kim, M.; Kong, C.-S.; Kim, Y.-Y.; Lee, S.-H. Comparison of biological activities in Sargassum siliquanstrum fermented by isolated lactic acid bacteria. Biotechnol. Bioprocess. Eng. 2015, 20, 341–348. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N.; Rajauria, G. Effect of heating and probiotic fermentation on the phytochemical content and antioxidant potential of edible Irish brown seaweeds. Bot. Mar. 2012, 55, 527–537. [Google Scholar] [CrossRef]
- Uchida, M.; Miyoshi, T. Algal Fermentation—The Seed for a New Fermentation Industry of Foods and Related Products. JARQ 2013, 47, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Pina-Pérez, M.C.; Rivas, A.; Martínez, A.; Rodrigo, D. Antimicrobial potential of macro and microalgae against pathogenic and spoilage microorganisms in food. Food Chem. 2017, 235, 34–44. [Google Scholar] [CrossRef]
- Sridhar, K.R.; Vidyavathi, N. Antimicrobial Activity of Seaweeds. Acta Hydrochim. Hydrobiol. 1991, 19, 455–496. [Google Scholar] [CrossRef]
- Mishra, A.K. Sargassum, Gracilaria and Ulva Exhibit Positive Antimicrobial Activity against Human Pathogens. OALib 2018, 5, 1–12. [Google Scholar] [CrossRef]
- Nshimiyumukiza, O.; Kang, S.-K.; Kim, H.-J.; Lee, E.-H.; Han, H.-N.; Kim, Y.; Kim, D.-H.; Kim, J.-H.; Eom, S.-H.; Kim, Y.-M. Synergistic Antibacterial Activity of Ecklonia cava (Phaeophyceae: Laminariales) against Listeria monocytogenes (Bacillales: Listeriaceae). Fish. Aquat. Sci. 2015, 18, 1–6. [Google Scholar]
- Rajauria, G.; Jaiswal, A.K.; Abu-Gannam, N.; Gupta, S. Antimicrobial, antioxidant and free radical-scavenging capacity of brown seaweed Himanthalia elongata from western coast of Ireland: Antimicrobial and antioxidant properties of irish seaweed. J. Food Biochem. 2013, 37, 322–335. [Google Scholar] [CrossRef]
- Hollants, J.; Leliaert, F.; De Clerck, O.; Willems, A. What we can learn from sushi: A review on seaweed-bacterial associations. FEMS Microbiol. Ecol. 2013, 83, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Pérez, M.; Falqué, E.; Domínguez, H. Antimicrobial Action of Compounds from Marine Seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Moayedi, A.; Hashemi, M.; Safari, M. Valorization of tomato waste proteins through production of antioxidant and antibacterial hydrolysates by proteolytic Bacillus subtilis: Optimization of fermentation conditions. J. Food Sci. Technol. 2016, 53, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Kantachote, D.; Charernjiratrakul, W.; Umsakul, K. Antibacterial Activities of Fermented Plant Beverages Collected in Southern Thailand. Antibact. Act. Fermented Plant. Beverages Collect. South. Thail. 2008, 8, 1280–1288. [Google Scholar] [CrossRef] [Green Version]
- Marinho, G.S.; Sørensen, A.-D.M.; Safafar, H.; Pedersen, A.H.; Holdt, S.L. Antioxidant content and activity of the seaweed Saccharina latissima: A seasonal perspective. J. Appl. Phycol. 2019, 31, 1343–1354. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Bernaldo de Quirós, A.; Frecha-Ferreiro, S.; Vidal-Pérez, A.M.; López-Hernández, J. Antioxidant compounds in edible brown seaweeds. Eur. Food Res. Technol. 2010, 231, 495–498. [Google Scholar] [CrossRef]
- Lopes, G.; Barbosa, M.; Vallejo, F.; Gil-Izquierdo, Á.; Andrade, P.B.; Valentão, P.; Pereira, D.M.; Ferreres, F. Profiling phlorotannins from Fucus spp. of the Northern Portuguese coastline: Chemical approach by HPLC-DAD-ESI/MS and UPLC-ESI-QTOF/MS. Algal Res. 2018, 29, 113–120. [Google Scholar] [CrossRef]
- Vissers, A.M.; Caligiani, A.; Sforza, S.; Vincken, J.-P.; Gruppen, H. Phlorotannin Composition of Laminaria digitata. Phytochem. Anal. 2017, 28, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Hermund, D.B.; Plaza, M.; Turner, C.; Jónsdóttir, R.; Kristinsson, H.G.; Jacobsen, C.; Nielsen, K.F. Structure dependent antioxidant capacity of phlorotannins from Icelandic Fucus vesiculosus by UHPLC-DAD-ECD-QTOFMS. Food Chem. 2018, 240, 904–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, T.M.; Targett, N.M. To grow and defend: Lack of tradeoffs for brown algal phlorotannins. Oikos 2003, 100, 406–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HE I | 24 h | 72 h | ||
|---|---|---|---|---|
| TPC (mg GAEs/g DW) | Lactobacillus casei | 2.94 ± 0.28 a | 0.27 ± 0.01 b,A | 0.21 ± 0.02 b,B |
| Lactobacillus paracasei | 2.94 ± 0.28 a | 0.20 ± 0.00 b,B | 0.20 ± 0.02 b,B | |
| Lactobacillus rhamnosus | 2.94 ± 0.28 a | 0.22 ± 0.01 b,B | 0.32 ± 0.08 b,A | |
| Bacillus subtilis | 2.94 ± 0.28 a | 0.20 ± 0.02 b,B | 0.36 ± 0.00 b,A |
| Compound | RT (min) | [M–H]– (m/z) | MS2 Ion Fragments(m/z) | Ref. |
|---|---|---|---|---|
| Trimer | 4.56 | 369 | 351 | a |
| Trimer | 5.16 | 369 | 279, 351, 325, 307 | a |
| Trimer | 5.34 | 369 | 351, 295, 325, 307 | a |
| Trimer | 5.65 | 369 | 351, 279, 325, 307 | a |
| Trimer | 7.00 | 369 | 238 | a |
| Trimer | 7.14 | 369 | 238 | a |
| Trimer | 1.50 | 373 | 355, 207, 329, 165 | b |
| Trimer (Phlorethol) | 5.39 | 373 | 231, 355 | c |
| Trimer (Fucophlorethol) | 5.75 | 373 | 233, 247, 229, 355, 125 | c |
| Tetramer | 5.22 | 497 | 479, 407, 371 | |
| Tetramer | 5.40 | 497 | 479, 353, 371, 335 | b |
| Tetramer | 5.85 | 497 | 235 | a |
| Tetramer | 6.36 | 497 | 355, 371, 479 | a |
| Tetramer | 6.69 | 497 | 373, 371, 233, 353, 238, 479 | b |
| Pentamer | 5.95 | 621 | 603, 339, 337 | a,b |
| Pentamer | 6.22 | 621 | 603, 339, 357, 337, 229 | a,b |
| Pentamer (Fucol) | 7.16 | 621 | 495, 371, 497, 477, 229, 603 | c |
| Pentamer (Fuhalol) | 8.06 | 651 | 509, 465, 413, 607, 339, 582 | c |
| Hexamer | 6.51 | 745 | 727, 601 | a |
| Heptamer | 7.12 | 869 | 851, 842, 833 | b,c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martelli, F.; Favari, C.; Mena, P.; Guazzetti, S.; Ricci, A.; Del Rio, D.; Lazzi, C.; Neviani, E.; Bernini, V. Antimicrobial and Fermentation Potential of Himanthalia elongata in Food Applications. Microorganisms 2020, 8, 248. https://doi.org/10.3390/microorganisms8020248
Martelli F, Favari C, Mena P, Guazzetti S, Ricci A, Del Rio D, Lazzi C, Neviani E, Bernini V. Antimicrobial and Fermentation Potential of Himanthalia elongata in Food Applications. Microorganisms. 2020; 8(2):248. https://doi.org/10.3390/microorganisms8020248
Chicago/Turabian StyleMartelli, Francesco, Claudia Favari, Pedro Mena, Stefano Guazzetti, Annalisa Ricci, Daniele Del Rio, Camilla Lazzi, Erasmo Neviani, and Valentina Bernini. 2020. "Antimicrobial and Fermentation Potential of Himanthalia elongata in Food Applications" Microorganisms 8, no. 2: 248. https://doi.org/10.3390/microorganisms8020248