
Microbial Influence on the Mobility of +3 Actinides from a Salt-Based Nuclear Waste Repository
Abstract
:1. Introduction
2. Materials and Methods
2.1. Solution Preparation
2.2. Organism Preparation
2.3. Experimental Batch Setup
2.4. Sampling and Analysis
2.5. SEM-EDS Analysis
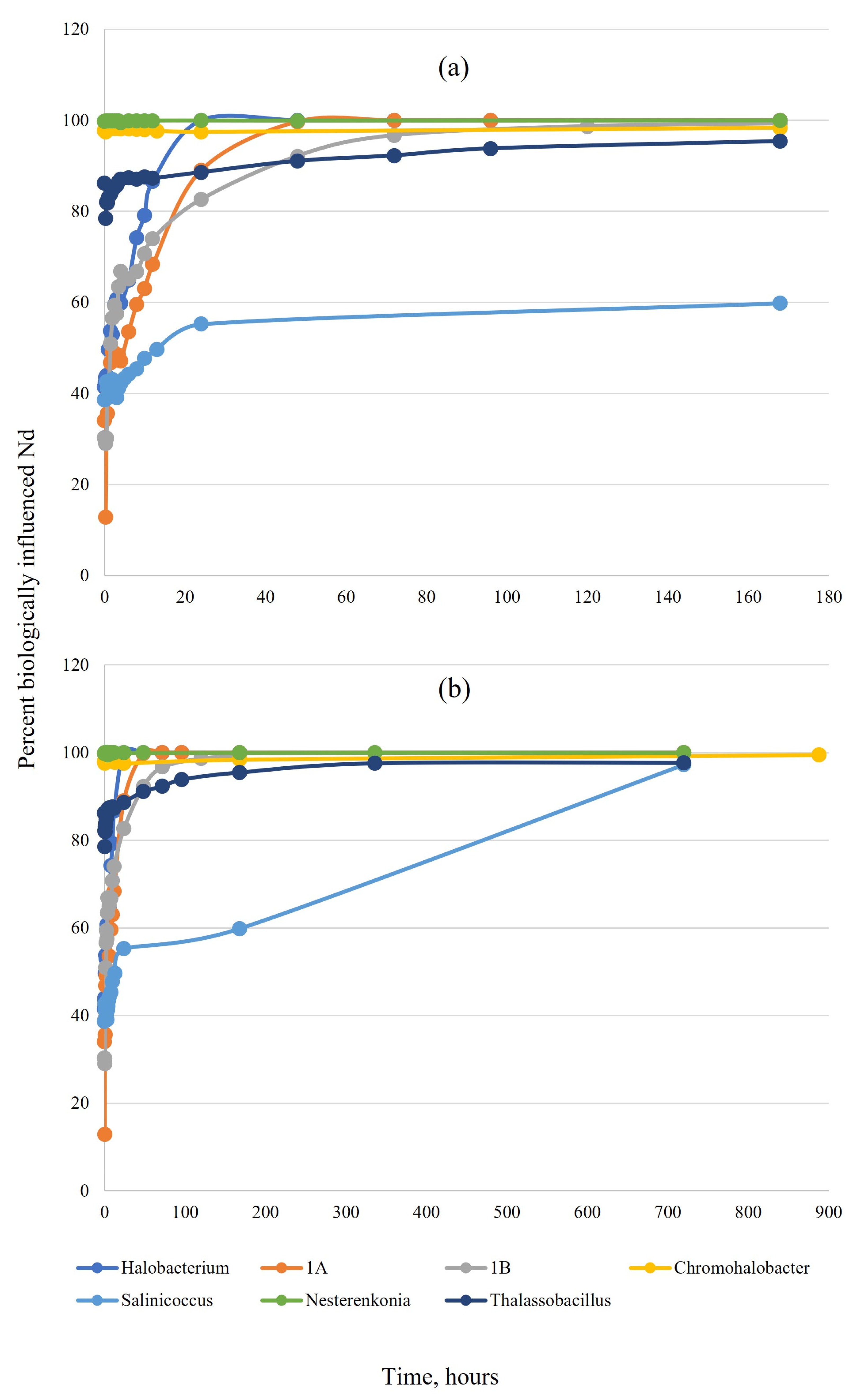
3. Results
3.1. Tests in Simplified NaCl Solution
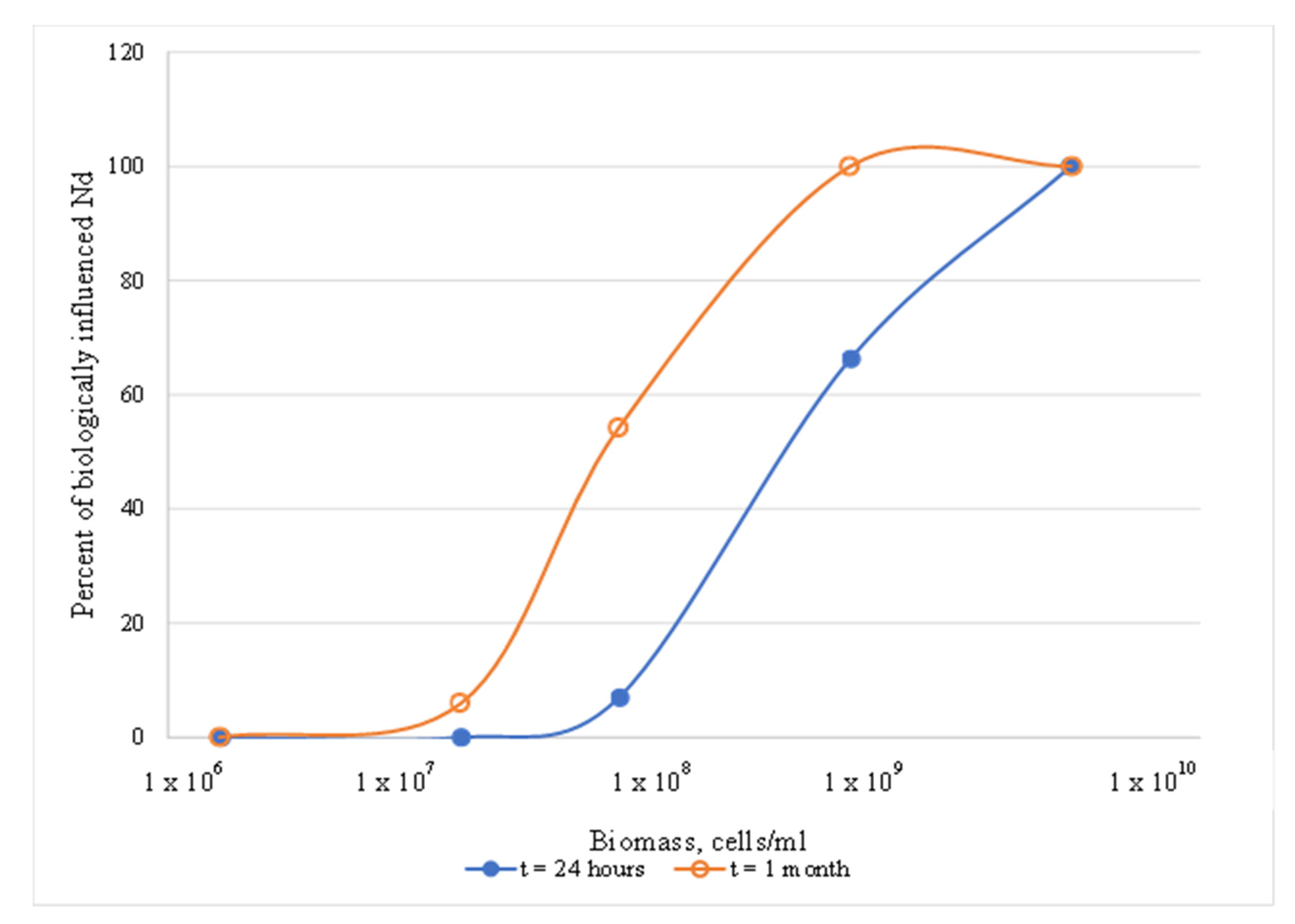
3.1.1. Effects of Biomass Concentration
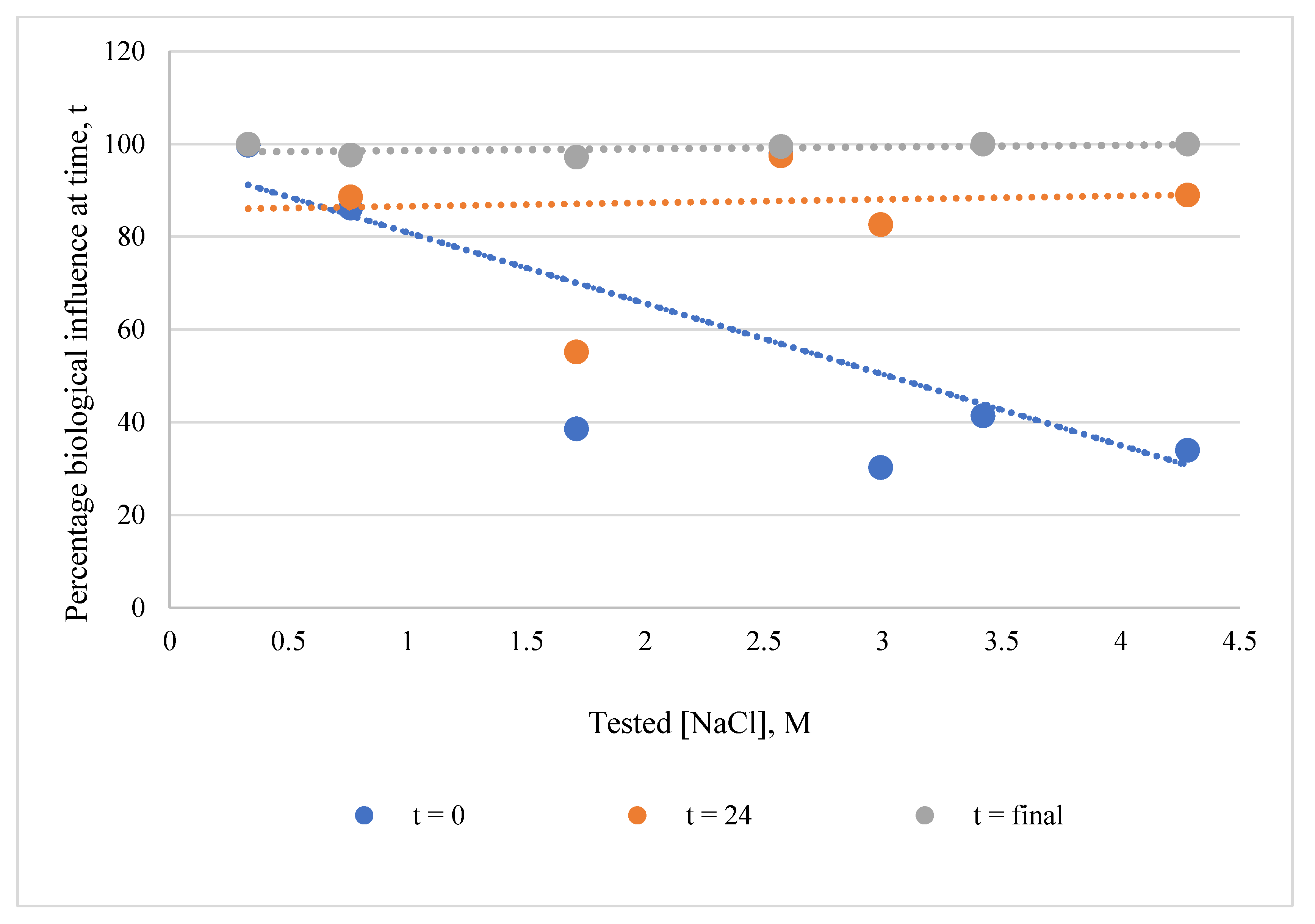
3.1.2. Effects of NaCl Concentration
3.2. pH Effects
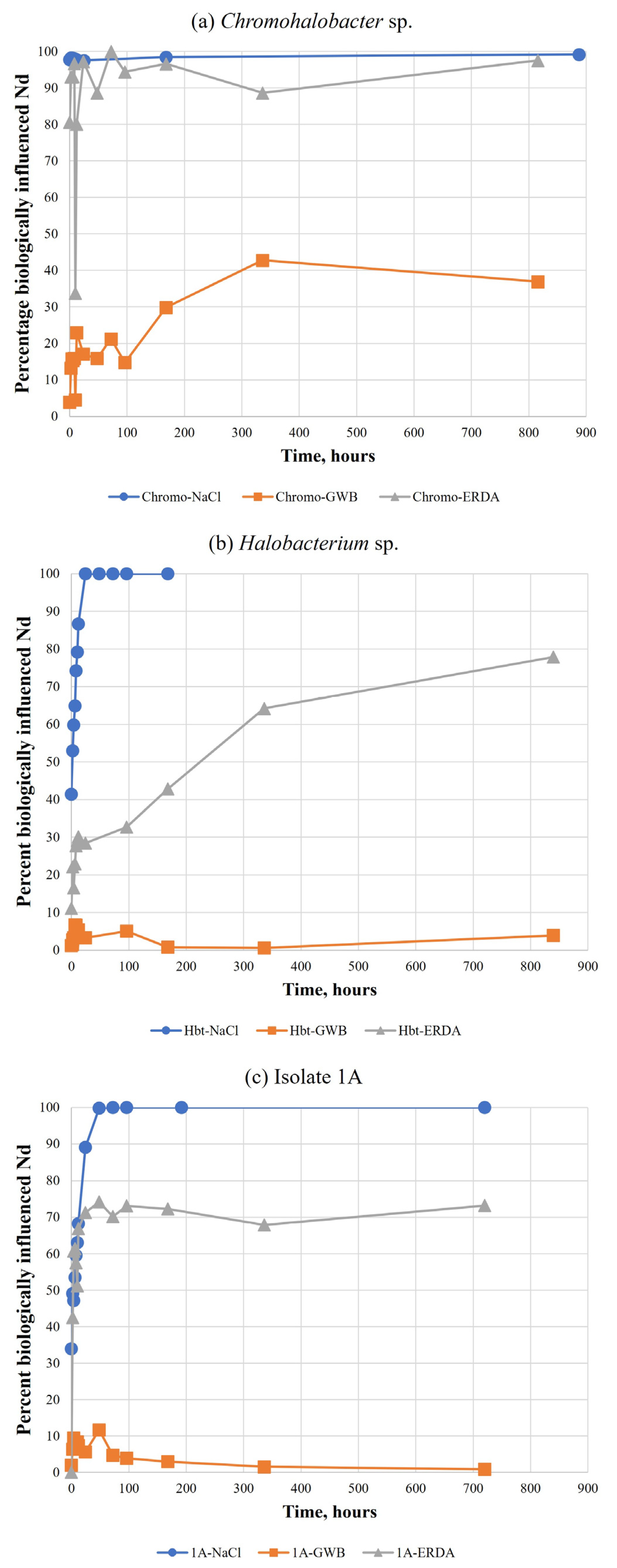
3.3. Tests in WIPP Brines
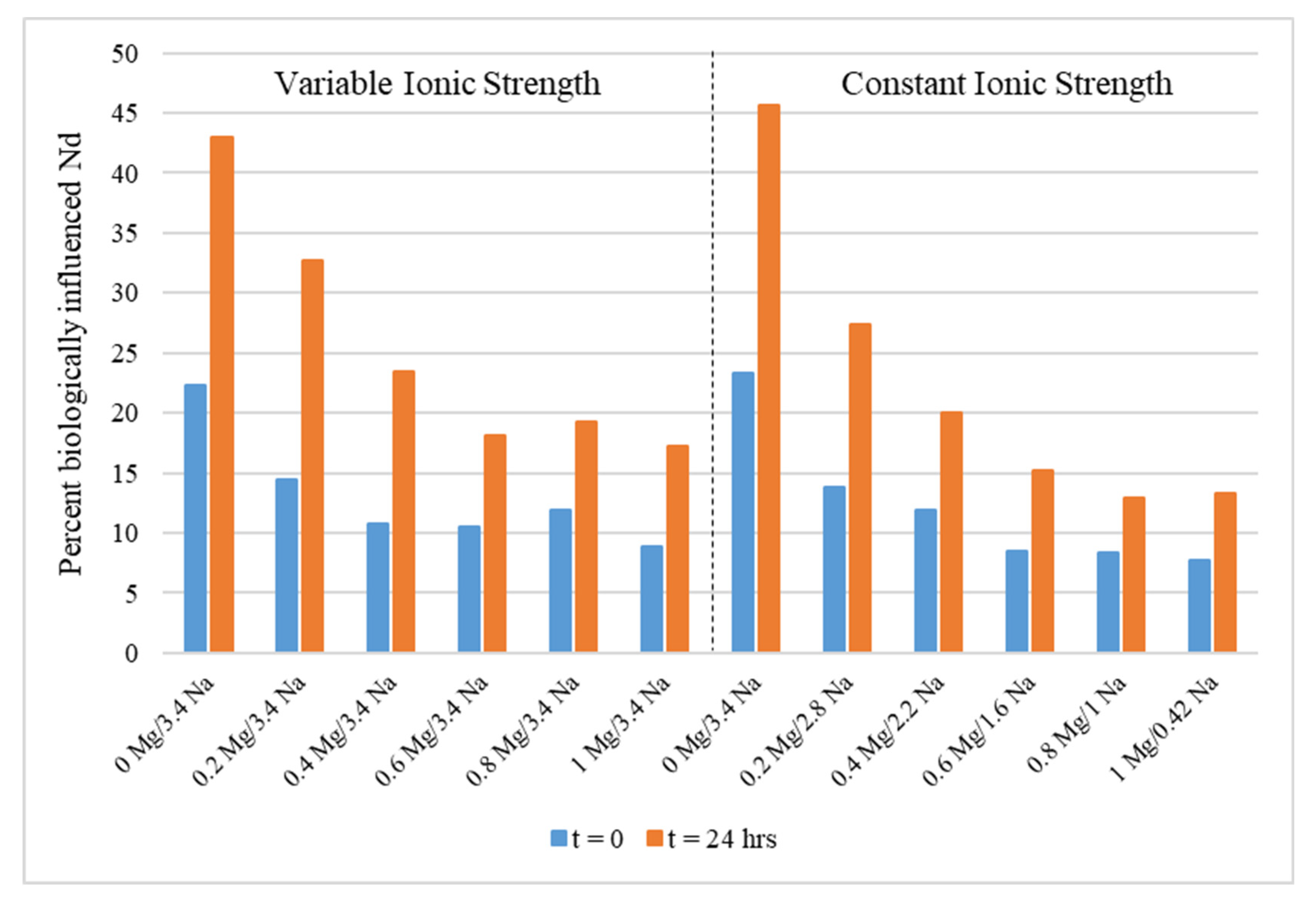
3.4. Tests in MgCl2-NaCl Solutions
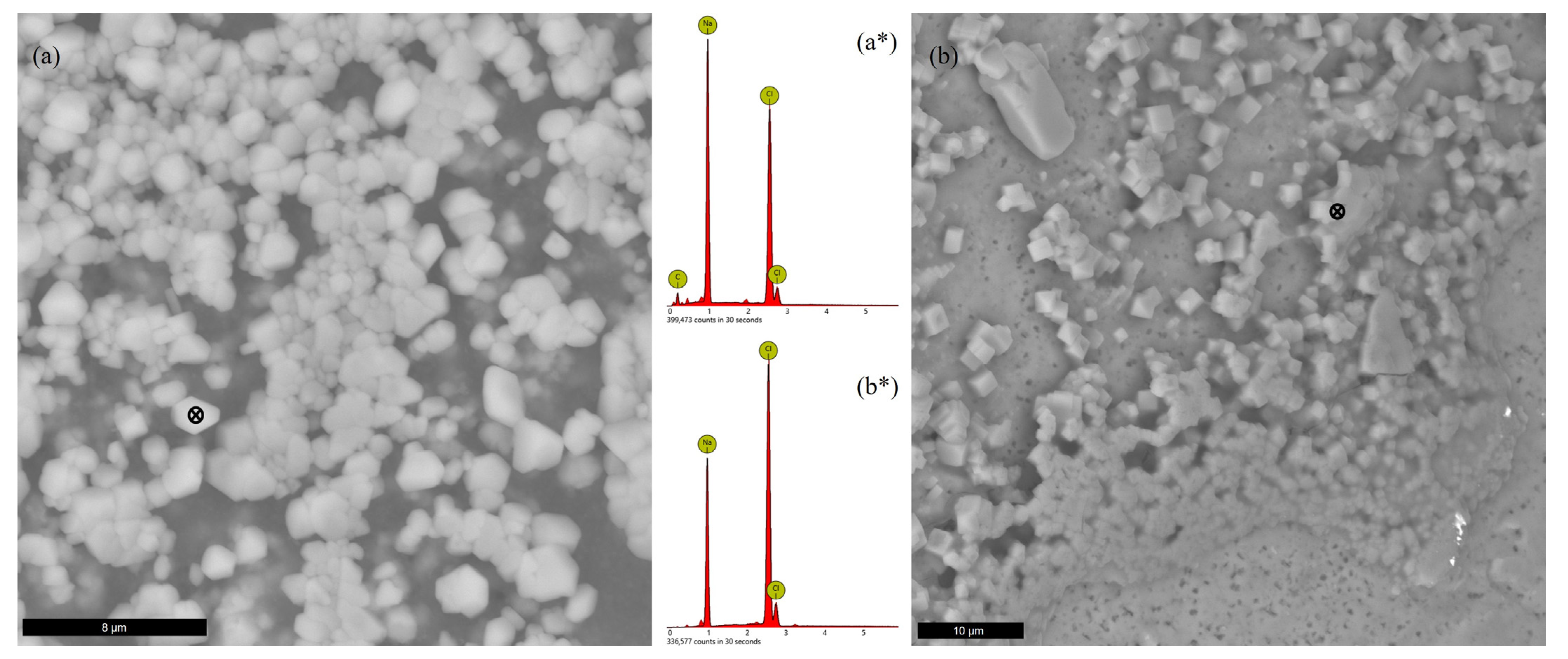
3.5. SEM-EDS Analyses
3.5.1. Precipitated Solids
3.5.2. Sedimented Biological Material
4. Discussion
4.1. Comparison between Organisms
4.2. Comparison between Simple NaCl Solutions and WIPP Brines
4.2.1. Neodymium Speciation
4.2.2. Loss of S-Layers
4.2.3. Magnesium Concentrations in Test Matrices
4.2.4. Biologically Induced Formation of Nd-Containing Precipitates
4.3. Possible Internal Accumulation of Neodymium
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Francis, A.J.; Gillow, J.B.; Dodge, C.J.; Dunn, M.; Mantione, K.; Strietelmeier, B.A.; Pansoy-Hjelvik, M.E.; Papenguth, H.W. Role of bacteria as biocolloids in the transport of actinides from a deep underground radioactive waste repository. Radiochim. Acta 1998, 82, 347–354. [Google Scholar] [CrossRef]
- Takenaka, Y.; Ozaki, T.; Ohnuki, T. Influence of ionic strength on curium(III) and europium(III) sorption on Halomonas elongata. J. Nucl. Sci. Technol. 2004, 41, 1125–1127. [Google Scholar] [CrossRef]
- Ozaki, T.; Gillow, J.B.; Kimura, T.; Ohnuki, T.; Yoshida, Z.; Francis, A.J. Sorption behavior of europium(III) and curium(III) on the cell surfaces of microorganisms. Radiochim. Acta 2004, 92, 741–748. [Google Scholar] [CrossRef]
- Bader, M.; Moll, H.; Steudtner, R.; Lösch, H.; Drobot, B.; Stumpf, T.; Cherkouk, A. Association of Eu(III) and Cm(III) onto an extremely halophilic archaeon. Environ. Sci. Pollut. R 2019, 26, 9352–9364. [Google Scholar] [CrossRef] [PubMed]
- Borkowski, M. Actinide(III) Solubility in WIPP Brine: Data Summary and Recommendations; Report LCO-ACP-08; Los Alamos National Laboratory: Carlsbad, NM, USA, 2009. [Google Scholar]
- Swanson, J.; Simmons, K.; Norden, D.; Ams, D.; Reed, D. Status Report on the Microbial Characterization of Halite and Groundwater Samples from the WIPP; Los Alamos Report LCO-ACP-21, LA-UR-12-22824; Los Alamos National Laboratory: Carlsbad, NM, USA, 2012. [Google Scholar]
- Swanson, J.; Reed, D. Update on Microbiology Research for the Waste Isolation Pilot Plant; Los Alamos Report LA-UR-18-31313; Los Alamos National Laboratory: Carlsbad, NM, USA, 2018. [Google Scholar]
- Biswas, J.; Paul, A.K. Production of extracellular polymeric substances by halophilic bacteria of solar salterns. Chin. J. Biol. 2014, 2014, 205731. [Google Scholar] [CrossRef]
- Rodrigues-Oliveira, T.; Belmok, A.; Vasconcellos, D.; Schuster, B.; Kyaw, C.M. Archaeal S-layers: Overview and current state of the art. Front. Microbiol. 2017, 8, 2597. [Google Scholar] [CrossRef] [PubMed]
- Migdisov, A.A.; Williams-Jones, A.E. A spectrophotometric study of neodymium(III) complexation in chloride solutions. Geochim. Cosmochim. Acta 2002, 66, 4311–4323. [Google Scholar] [CrossRef]
- Neck, V.; Altmaier, M.; Rabung, T.; Lützenkirchen, J.; Fanghänel, T. Thermodynamics of trivalent actinides and neodymium in NaCl, MgCl2 and CaCl2 solutions: Solubility, hydrolysis, and ternary Ca-M(III)-OH complexes. Pure Appl. Chem. 2009, 81, 1555–1568. [Google Scholar] [CrossRef]
- Hinz, K.; Altmaier, M.; Gaona, X.; Rabung, T.; Schild, D.; Richmann, M.; Reed, D.T.; Alekseev, E.V.; Geckeis, H. Interaction of Nd(III) and Cm(III) with borate in dilute to concentrated alkaline NaCl, MgCl2 and CaCl2 solutions: Solubility and TRLFS studies. New J. Chem. 2015, 39, 849–859. [Google Scholar] [CrossRef]
- Horikoshi, K.; Aono, R.; Nakamura, S. The triangular halophilic archaebacterium Haloarcula japonica strain TR-1. Experientia 1993, 49, 497–502. [Google Scholar] [CrossRef]
- Mobili, P.; de los Angeles Serradell, M.; Mayer, C.; Arluison, V.; Gomez-Zavaglia, A. Biophysical methods for the elucidation of the S-layer proteins/metal interaction. Int. J. Biochem. Res. 2013, 3, 39–62. [Google Scholar] [CrossRef] [PubMed]
- Chandramohan, A.; Duprat, E.; Remusat, L.; Zirah, S.; Lombard, C.; Kish, A. Novel mechanism for surface layer shedding and regenerating in bacteria exposed to metal-contaminated conditions. Front. Microbiol. 2019, 9, 3210. [Google Scholar] [CrossRef]
- Schultze-Lam, S.; Fortin, D.; Davis, B.S.; Beveridge, T.J. Mineralization of bacterial surfaces. Chem. Geol. 1996, 132, 171–181. [Google Scholar] [CrossRef]
- Kish, A.; Miot, J.; Lombard, C.; Guigner, J.-M.; Bernard, S.; Zirah, S.; Guyot, F. Preservation of archaeal surface layer structure during mineralization. Nat. Sci. Rep. 2016, 6, 26512. [Google Scholar] [CrossRef]
- Merroun, M.L.; Raff, J.; Rossberg, A.; Hennig, C.; Reich, T.; Selenska-Pobell, S. Complexation of uranium by cells and S-layer sheets of Bacillus spaericus JG-A12. Appl. Environ. Microb. 2005, 71, 5532–5543. [Google Scholar] [CrossRef] [PubMed]
- Francis, A.J.; Gillow, J.B.; Dodge, C.J.; Harris, R.; Beveridge, T.J.; Papenguth, H.W. Uranium association with halophilic and nn-halophilic bacteria and archaea. Radiochim. Acta 2004, 92, 481–488. [Google Scholar] [CrossRef]
- Frankel, R.B.; Bazylinski, D.A. Biologically induced mineralization by bacteria. Rev. Miner. Geochem. 2003, 54, 95–114. [Google Scholar] [CrossRef]
- Hoffmann, T.D.; Reeksting, B.J.; Gebhard, S. Bacteria-induced mineral precipitation: A mechanistic review. Microbiology 2021, 167, 001049. [Google Scholar] [CrossRef]
- Obst, M.; Dynes, J.J.; Lawrence, J.R.; Swerhone, G.D.W.; Benzerara, K.; Karunakaran, C.; Kaznatcheev, K.; Tyliszczak, T.; Hitchcock, A.P. Precipitation of amorphous CaCO3 (aragonite-like) by cyanobacteria: A STXM study of the influence of EPS on the nucleation process. Geochim. Cosmochim. Acta 2009, 73, 4180–4198. [Google Scholar] [CrossRef]
- Görgen, S.; Benzerara, K.; Skouri-Panet, F.; Gugger, M.; Chauvat, F.; Cassier-Chauvat, C. The diversity of molecular mechanisms of carbonate biomineralization by bacteria. Discov. Mater. 2021, 1, 2. [Google Scholar] [CrossRef]
- Kenward, P.A.; Fowle, D.A.; Goldstein, R.H.; Ueshima, M.; González, L.A.; Roberts, J.A. Ordered low-temperature dolomite mediated by carboxyl-group density of microbial cell walls. AAPG Bull. 2013, 97, 2113–2125. [Google Scholar] [CrossRef]
- Rivadeneyra, M.A.; Gelgado, G.; Soriano, M.; Ramos-Cormenzana, A.; Delgado, R. Precipitation of carbonates by Nesterenkonia halobia in liquid media. Chemosphere 2000, 41, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Rivadeneyra, M.A.; Martín-Algarra, A.; Sánchez-Navas, A.; Martín-Ramos, D. Carbonate and phosphate precipitation by Chromohalobacter marismortui. Geomicrobiol. J. 2006, 23, 89–101. [Google Scholar] [CrossRef]
- Sánchez-Román, M.; McKenzie, J.A.; de Luca Rebello Wagener, A.; Romanek, C.S.; Sánchez-Navas, A.; Vasconcelos, C. Experimentally determined biomediated Sr partition coefficient for dolomite: Significance and implication for natural dolomite. Geochim. Cosmochim. Acta 2011, 75, 887–904. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Concentration of Constituent, M | |
|---|---|---|
| Generic Weep Brine (GWB) | ERDA-6 Brine | |
| Na+ | 3.13 | 4.37 |
| K+ | 4.14 × 10−1 | 8.73 × 10−2 |
| Mg2+ | 9.01 × 10−1 | 1.71 × 10−2 |
| Ca2+ | 1.22 × 10−2 | 1.08 × 10−2 |
| Li+ | 3.89 × 10−3 | NA |
| B4O72− | 3.49 × 10−2 | 1.42 × 10−2 |
| Cl− | 4.96 | 4.17 |
| SO42− | 1.57 × 10−1 | 1.50 × 10−1 |
| Br− | 2.36 × 10−2 | 9.89 × 10−3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swanson, J.; Navarrette, A.; Knox, J.; Kim, H.; Stanley, F. Microbial Influence on the Mobility of +3 Actinides from a Salt-Based Nuclear Waste Repository. Microorganisms 2023, 11, 1370. https://doi.org/10.3390/microorganisms11061370
Swanson J, Navarrette A, Knox J, Kim H, Stanley F. Microbial Influence on the Mobility of +3 Actinides from a Salt-Based Nuclear Waste Repository. Microorganisms. 2023; 11(6):1370. https://doi.org/10.3390/microorganisms11061370
Chicago/Turabian StyleSwanson, Julie, Adrianne Navarrette, Jandi Knox, Hannah Kim, and Floyd Stanley. 2023. "Microbial Influence on the Mobility of +3 Actinides from a Salt-Based Nuclear Waste Repository" Microorganisms 11, no. 6: 1370. https://doi.org/10.3390/microorganisms11061370