
Ectothiorhodospira lacustris sp. nov., a New Purple Sulfur Bacterium from Low-Mineralized Soda Lakes That Contains a Unique Pathway for Nitric Oxide Reduction

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation
2.2. Morphological, Physiological, and Biochemical Characteristics
2.3. Genome Sequencing, 16S rRNA Gene Sequencing, Phylogenetic Analyses
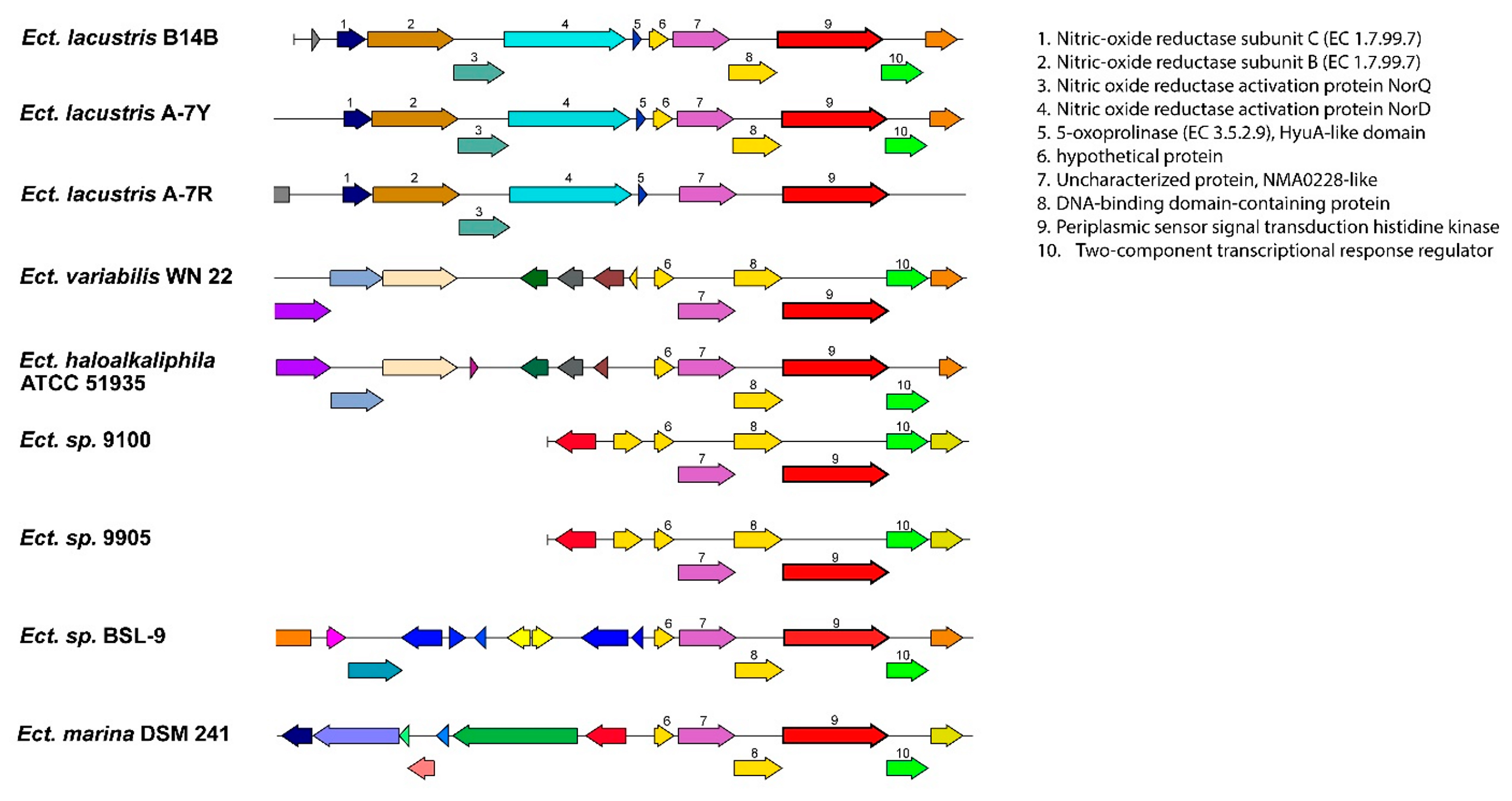
3. Results and Discussion
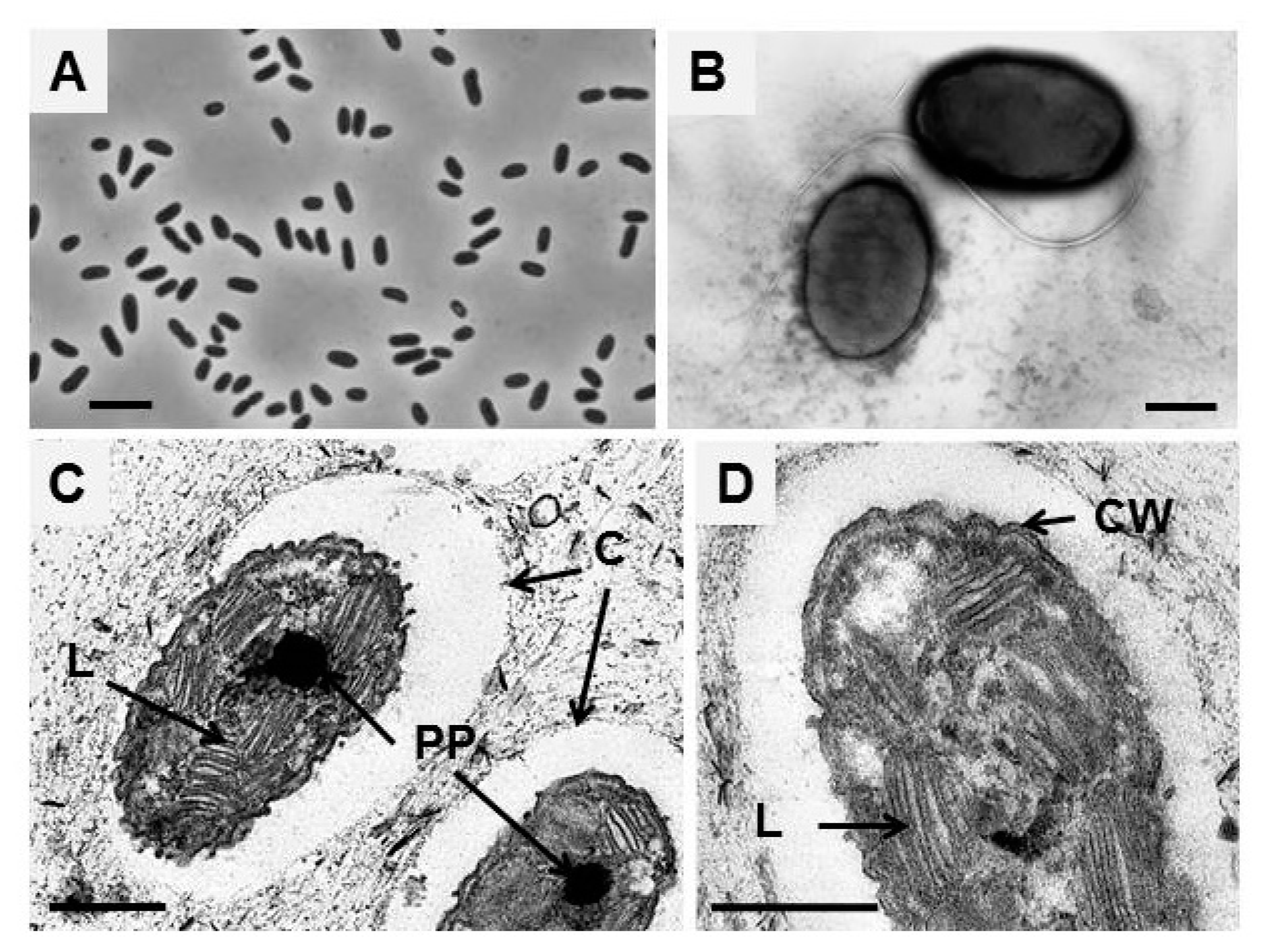
3.1. Morphology and Ultrastructure
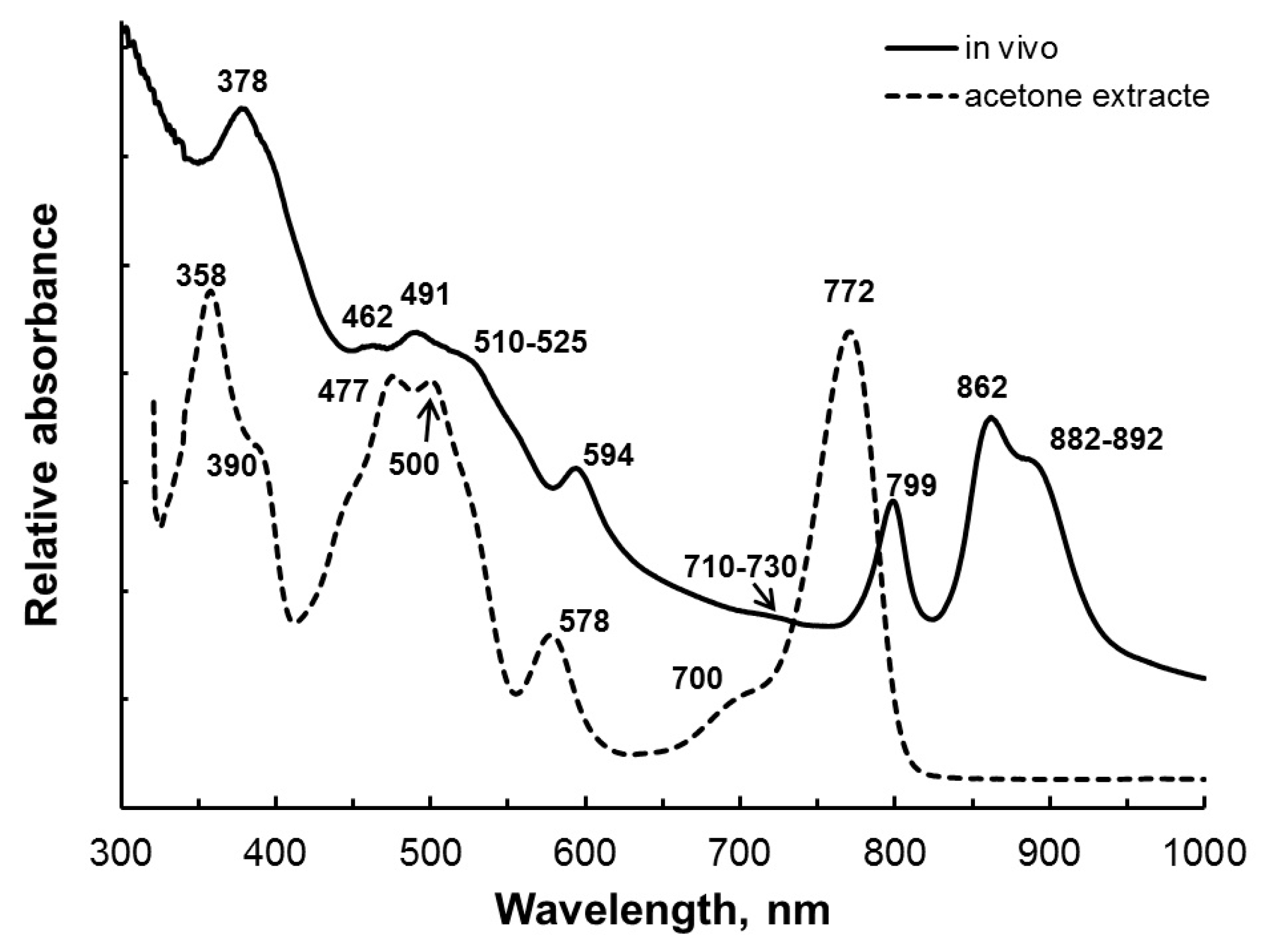
3.2. Pigments
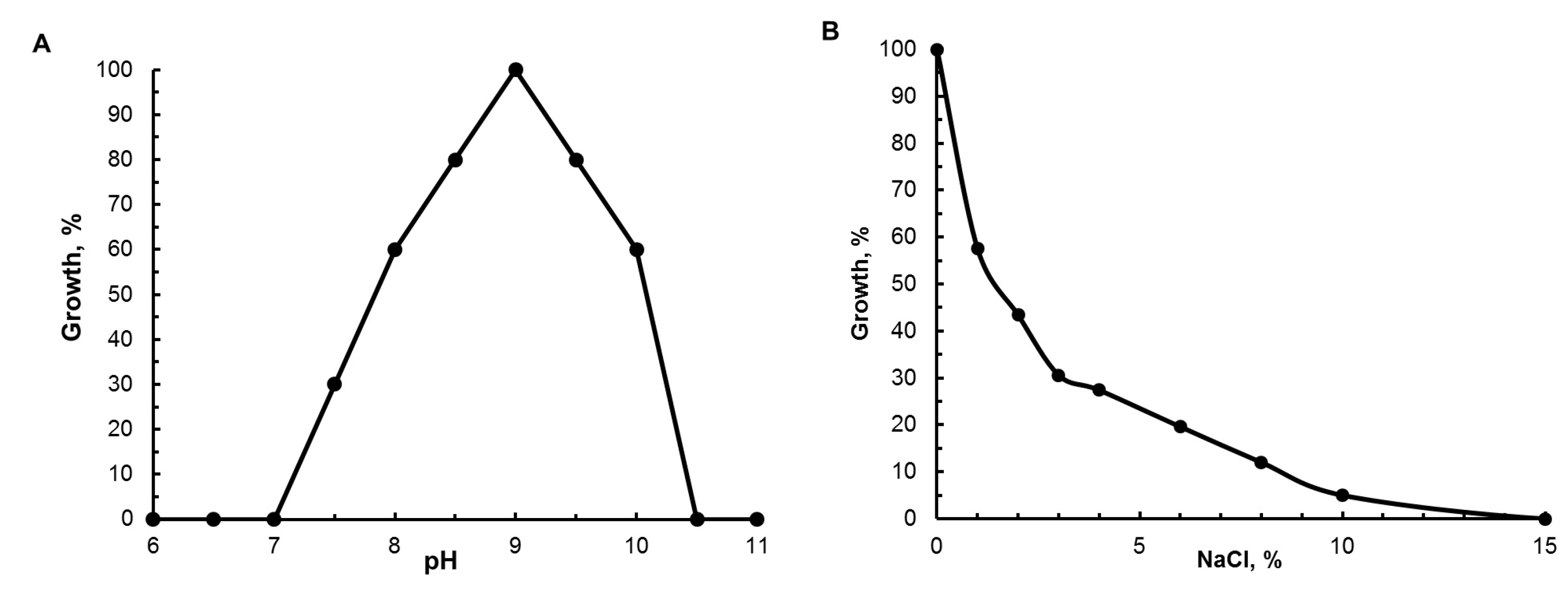
3.3. Physiological Characteristics
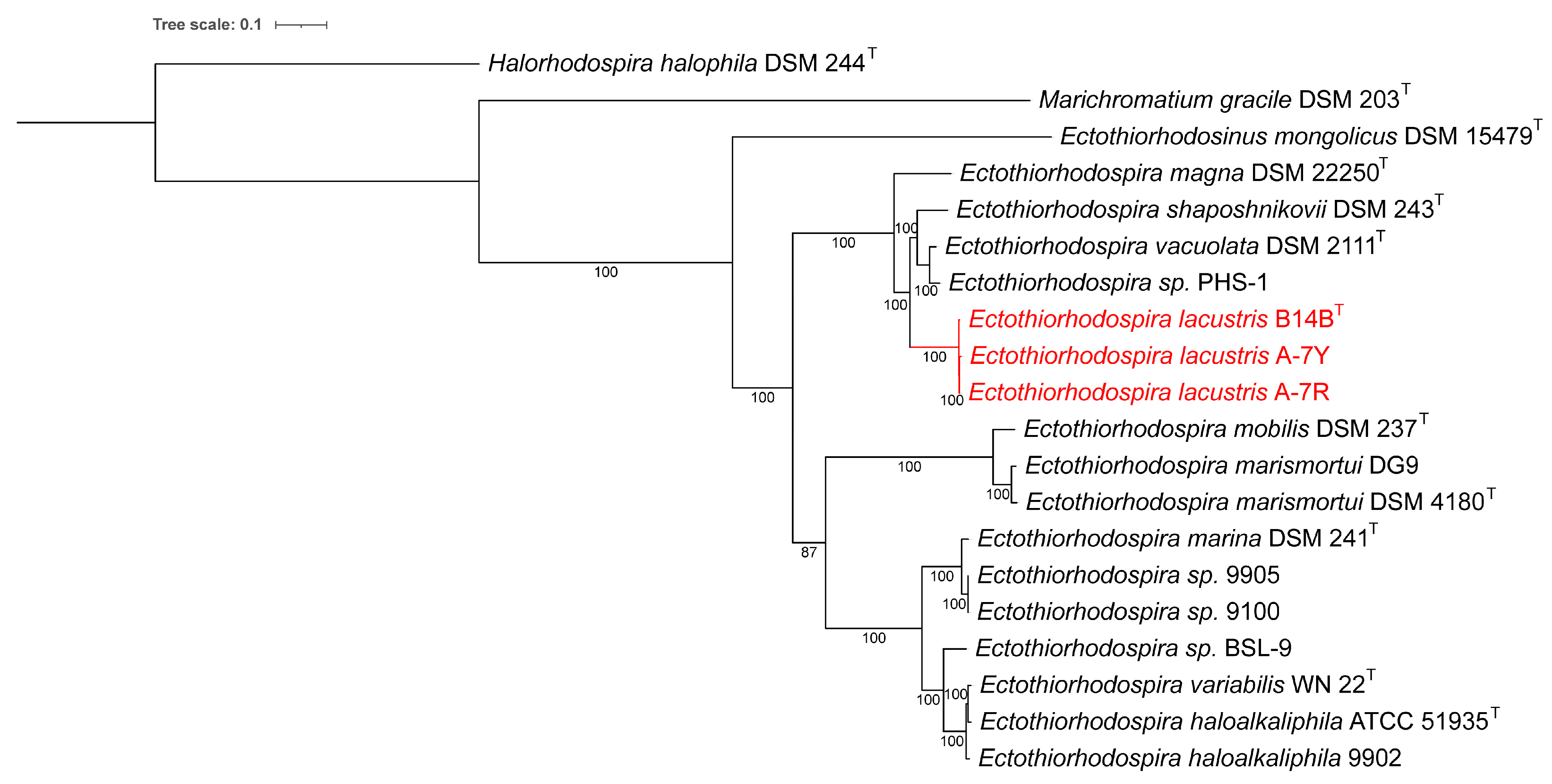
3.4. Taxonomic Position
4. Description of Ectothiorhodospira lacustris sp. nov.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imhoff, J.F.; Kyndt, J.A.; Meyer, T.E. Genomic comparison, phylogeny and taxonomic reevaluation of the Ectothiorhodospiraceae and description of Halorhodospiraceae fam. nov. and Halochlorospira gen. nov. Microorganisms 2022, 10, 295. [Google Scholar] [CrossRef]
- Imhoff, J.F. Genus ectothiorhodospira. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; Volume 2, pp. 43–48. [Google Scholar]
- Frigaard, N.U.; Dahl, C. Sulfur metabolism in phototrophic sulfur bacteria. Adv. Microb. Physiol. 2009, 54, 103–200. [Google Scholar] [CrossRef] [PubMed]
- Dahl, C. Sulfur Metabolism in Phototrophic Bacteria. In Modern Topics in the Phototrophic Prokaryotes; Hallenbeck, P.C., Ed.; Springer: Cham, Switzerland, 2017; pp. 27–66. [Google Scholar] [CrossRef]
- Oren, A. The Family Ectothiorhodospiraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 199–222. [Google Scholar] [CrossRef]
- Bryantseva, I.; Gorlenko, V.M.; Kompantseva, E.I.; Imhoff, J.F.; Süling, J.; Mityushina, L. Thiorhodospira sibirica gen. nov., sp. nov., a new alkaliphilic purple sulfur bacterium from a Siberian soda lake. Int. J. Syst. Bacteriol. 1999, 49, 697–703. [Google Scholar] [CrossRef]
- Gorlenko, V.M.; Bryantseva, I.A.; Panteleeva, E.E.; Tourova, T.P.; Kolganova, T.V.; Makhneva, Z.K.; Moskalenko, A. Ectothiorhodosinus mongolicum gen. nov., sp. nov., a new purple bacterium from a soda lake in Mongolia. Microbiology 2004, 73, 66–73. [Google Scholar] [CrossRef]
- Euzeby, J.P. Validation list no. 116. List of new names and new combinations previously effectively, but not validly, published. Int. J. Syst. Evol. Microbiol. 2007, 57, 1371–1373. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Keller, A.; Neulinger, S.C. Osmotic adaptation and compatible solute biosynthesis of phototrophic bacteria as revealed from genome analyses. Microorganisms 2019, 9, 46. [Google Scholar] [CrossRef]
- Gorlenko, V.M.; Imhoff, J.F. Biodiversity of alkaliphilic purple sulphur bacteria of Ectothiorhodospiraceae family. In Proceedings of the Program and Abstracts of the 12th International Symposium on Phototrophic Prokaryotes, Pau, France, 27 August–1 September 2006; p. 74. [Google Scholar]
- Pfennig, N.; Lippert, K.D. Über das Vitamin B12-Bedürfnis phototropher Schwefelbakterien. Arch. Mikrobiol. 1966, 55, 245–256. [Google Scholar] [CrossRef]
- Ryter, A.; Kellenberger, E.; Birch-Andersen, A.; Maaløe, O. Etude au microscope électronique des plasmes contenant de l’acide déoxyribonucléique 1 Les nucléoides des bactéries en croissance active. Z. Naturforsch. 1958, 13, 597–605. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–218. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R.; Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J.J.S. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bryantseva, I.A.; Tourova, T.P.; Kovaleva, O.L.; Kostrikina, N.A.; Gorlenko, V.M. Ectothiorhodospira magna sp. nov., a new large alkaliphilic purple sulfur bacterium. Microbiology 2010, 79, 780–790. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Henrik, C.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Braker, G.; Tiedje, J.M. Nitric oxide reductase (norB) genes from pure cultures and environmental samples. Appl. Environ. Microbiol. 2003, 69, 3476–3483. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.Y.; Dunlap, C.A.; Rooney, A.P.; Chen, G.J.; Du, Z.J. Wenzhouxiangella sediminis sp. nov., isolated from coastal sediment. Int. J. Syst. Evol. Microbiol. 2016, 66, 4575–4579. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Zhang, L.P.; Chen, S.F. Halomonas korlensis sp. nov., a moderately halophilic, denitrifying bacterium isolated from saline and alkaline soil. Int. J. Syst. Evol. Microbiol. 2008, 58, 2582–2588. [Google Scholar] [CrossRef] [PubMed]
- González-Domenech, C.M.; Béjar, V.; Martínez-Checa, F.; Quesada, E. Halomonas nitroreducens sp. nov., a novel nitrate-and nitrite-reducing species. Int. J. Syst. Evol. Microbiol. 2008, 58, 872–876. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Ectothiorhodospira lacustris | Ectothiorhodospira vacuolata | Ectothiorhodospira shaposhnikovii | Ectothiorhodospira magna |
|---|---|---|---|---|
| Cell shape | Oval- to rod-shaped, sometimes slightly bent | Rod-shaped, sometimes slightly bent | Rod-shaped, usually slightly bent vibrios or short spirilla | Vibrioid or spirilloid |
| Size, µm | 0.5–1.3 × 0.7–2.7 | 1.5 × 2–4 | 0.8–0.9 × 1.5–2.5 | 2–3.2 × 9.6–20 |
| Motility | ±, Polar tuft of flagella | +, Polar tuft of flagella | +, Polar tuft of flagella | +, Polar tuft of flagella |
| Gas vesicles | – | + | – | – |
| Internal membranes | Lamellar stacks | Lamellar stacks | Lamellar stacks | Long strands of lamellae |
| Color of the cell suspensions | Dark red | Pink to red | Red | Red to brownish red |
| Pigments | Bchl a, carotenoids of the spirilloxanthin series | Bchl a, carotenoids of the spirilloxanthin series | Bchl a, carotenoids of the spirilloxanthin series | Bchl a, carotenoids of the spirilloxanthin series |
| Absorption spectra of the whole cells | 378, 462, 491, (510–525), 594, (710–730), 799, 862, (882–892) | 378, 486, 513, 555, 593, 798, 855, (893) | 378, 481, 513, 556, 593, 798, 854, (892) | 373, 485–514, (545), 593, 797, 858, (885) |
| NaCl optimum (range), % | 0 (0–8) | 1–6 (0.5–10) | 3 (0–7) | 0.5–1.5 (0–8) |
| pH optimum (range) | 9.0 (7.5–10.2) | 7.5–9.5 | 8.0–8.5 | 9–10 (8–11) |
| Nitrogen sources | Ammonia, nitrate, urea, dinitrogen (genomic data) | Ammonia, dinitrogen | Ammonia, dinitrogen, nitrate, some amino acids | Ammonia |
| Growth factors | Not required | Not required | Not required | Yeast extract satisfies all requirements |
| Photoautotrophic growth | + * | + | + | + * |
| Photoheterotrophic growth | + | + | u | u |
| Chemolithotrophic growth | – | u | + | – |
| Sulfate assimilation | – | – | + | – |
| Substrates used: | ||||
| Hydrogen | ND | + | + | + * |
| Sulfide | + | + | + | + |
| Thiosulfate | – | + | + | – |
| Sulfur | + | + | + | + |
| Sulfite | – | u | + | – |
| Organic substances | Acetate, butyrate, yeast extract, lactate, malate, pyruvate, succinate, fumarate | Acetate, malate, propionate, pyruvate, succinate, (fructose), fumarate | Acetate, butyrate, lactate, malate, propionate, pyruvate, succinate, fructose, fumarate | Acetate, malate, yeast extract *, propionate *, pyruvate *, succinate, fumarate |
| DNA G + C content of the species, mol% | 62.9–63.0 (genomic DNA data) | 61.4–63.6 (Tm) | 62.0–64.0 (Tm) | 59.2 (Tm) |
| 61.5–62.1 (Tm) | ||||
| DNA G+C content of the type strain, mol% | 62.9 (genomic DNA data) 61.5 (Tm) | 63.4 (genomic DNA data) 63.6 (Tm) | 62.4 (genomic DNA data) 62.0 (Tm) | 60.9 (genomic DNA data) 59.2 (Tm) |
| Subject Strain | dDDH (d4, in %) | C.I. (d4, in %) | G + C% Difference |
|---|---|---|---|
| Ectothiorhodospira vacuolata DSM 2111T | 38.8 | [36.4–41.3] | 0.52 |
| Ectothiorhodospira shaposhnikovii DSM 243T | 36.6 | [34.1–39.1] | 0.53 |
| Ectothiorhodospira magna B7-7T = DSM 22250T | 30 | [27.7–32.6] | 1.97 |
| Thiorhodospira sibirica A12T = ATCC 700588T | 22.3 | [20.1–24.8] | 6.2 |
| Ectothiorhodospira marina DSM 241T | 21.5 | [19.2–23.9] | 0.63 |
| Ectothiorhodospira variabilis WN22T = DSM 21381T | 21.4 | [19.1–23.8] | 0.08 |
| Ectothiorhodospira marismortui DSM 4180T | 20.8 | [18.6–23.2] | 5.33 |
| Thioalkalivibrio sulfidiphilus HL-EbGr7T | 20.7 | [18.5–23.1] | 2.16 |
| Ectothiorhodospira mobilis DSM 237T | 20.7 | [18.5–23.1] | 5.42 |
| Thioalkalivibrio denitrificans ALJDT = DSM 13742T | 20.2 | [18.0–22.7] | 1.67 |
| Ectothiorhodosinus mongolicus M9T = DSM 15479T | 19.7 | [17.5–22.1] | 7.29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryantseva, I.A.; Kyndt, J.A.; Gorlenko, V.M.; Imhoff, J.F. Ectothiorhodospira lacustris sp. nov., a New Purple Sulfur Bacterium from Low-Mineralized Soda Lakes That Contains a Unique Pathway for Nitric Oxide Reduction. Microorganisms 2023, 11, 1336. https://doi.org/10.3390/microorganisms11051336
Bryantseva IA, Kyndt JA, Gorlenko VM, Imhoff JF. Ectothiorhodospira lacustris sp. nov., a New Purple Sulfur Bacterium from Low-Mineralized Soda Lakes That Contains a Unique Pathway for Nitric Oxide Reduction. Microorganisms. 2023; 11(5):1336. https://doi.org/10.3390/microorganisms11051336
Chicago/Turabian StyleBryantseva, Irina A., John A. Kyndt, Vladimir M. Gorlenko, and Johannes F. Imhoff. 2023. "Ectothiorhodospira lacustris sp. nov., a New Purple Sulfur Bacterium from Low-Mineralized Soda Lakes That Contains a Unique Pathway for Nitric Oxide Reduction" Microorganisms 11, no. 5: 1336. https://doi.org/10.3390/microorganisms11051336