
Using Genomics to Decipher the Enigmatic Properties and Survival Adaptation of Candidate Phyla Radiation

1
Aix-Marseille Université, IRD, APHM, MEPHI, 13005 Marseille, France
2
IHU Méditerranée Infection, 13005 Marseille, France
*
Author to whom correspondence should be addressed.
Microorganisms 2023, 11(5), 1231; https://doi.org/10.3390/microorganisms11051231
Submission received: 10 April 2023
/
Revised: 28 April 2023
/
Accepted: 5 May 2023
/
Published: 7 May 2023
(This article belongs to the Special Issue New Methods in Microbial Research 3.0)
Abstract
:Microbial ecology is a critical field for understanding the composition, diversity, and functions of microorganisms in various environmental and health-related processes. The discovery of Candidate Phyla Radiation (CPR) through culture-independent methods has introduced a new division of microbes characterized by a symbiotic/parasitic lifestyle, small cell size, and small genome. Despite being poorly understood, CPRs have garnered significant attention in recent years due to their widespread detection in a variety of environmental and clinical samples. These microorganisms have been found to exhibit a high degree of genetic diversity compared to other microbes. Several studies have shed light on their potential importance in global biogeochemical cycles and their impact on various human activities. In this review, we provide a systematic overview of the discovery of CPRs. We then focus on describing how the genomic characteristics of CPRs have helped them interact with and adapt to other microbes in different ecological niches. Future works should focus on discovering the metabolic capacities of CPRs and, if possible, isolating them to obtain a better understanding of these microorganisms.
1. Introduction: Progress in High-Throughput Technologies Enables the Identification of New Prokaryotic Taxa
Greater in-depth knowledge of the microbial diversity of any community is crucial in order to identify its roles in various processes that affect human health and the environment. Our knowledge was previously limited to the detectable bacterial domains, of which approximately 2% are cultivable taxa [1]. Equally crucial is the hunt for unknown microbes, so-called “microbial dark matter”, for their importance in maintaining the different ecosystems [2,3]. Now, as a result of high-throughput technologies, a tsunami of new genome sequences has reshaped our understanding of the diversity of life [4]. However, these methods have revealed that the proportion of unknowns may be even higher than expected. Due to insufficient sequencing depth and the complexity of performing genome assemblies, it was initially challenging to sequence populations with less than 1% relative abundance in complex metagenomes [5]. Single-cell genomics was the solution, but the technique was hampered by an amplification bias [2,6]. However, rapid improvements in the quality of sequencing enabled the discovery of Candidate Phyla Radiation (CPR) (Figure 1) [7]. This group of prokaryotic organisms is a large evolutionary radiation whose members have not yet been fully characterized. As the name indicates, a “candidate phylum” (analogous to a “candidate division”) has no cultured representative to date, but its existence has been established through culture-independent molecular methods [8]. Notably, some CPR groups could not be detected in rRNA research due to primer mismatch, introns, or both [9,10]. However, microfluidic single-cell dispenser systems have made it possible to analyze non-culturable microbial cells, including CPRs (Figure 1) [11].
CPRs, also referred to as “ultramicrobacteria” or “nanobacteria”, have a reduced genome (around one megabase pair) and are small in size (100 nm to 300 nm), with a cell volume of 0.01 μm3 (Figure 1 and Table 1) [5,7,12]. This volume was revealed by cryogenic transmission electron microscope images that targeted post-0.2 mm filtrates collected from environmental samples [13]. Moreover, metagenomic and/or metabarcoding analyses have revealed that CPRs are ubiquitous in different ecosystems around the world. They have been detected in samples of drinking water, deep subsurface sediments, soil, and even in anaerobic subsurfaces where organisms from extremely ancient lineages that diverged from primitive life forms reside [4,9,14,15,16,17,18]. CPR superphyla have also been reported in clinical samples of the human microbiome (Figure 1) [19,20,21,22,23].
The CPR group is diverse, representing more than 26% of known microbial species [24]. This abundance is uncertain and depends on the type of analysis performed and the time it is carried out [7,12,25]. A dramatic increase in biological data has significantly expanded the scale of the microbial community, and new species are regularly being identified. CPRs include more than 70 phyla, with around 11,000 genomes, most of which are incomplete and considered as metagenome-assembled genomes (MAGs) in the National Center for Biotechnology Information (NCBI) database [26]. CPRs are also sometimes referred to as “Patescibacteria”, since more than 74% of their genomes belong to this superphylum or phylum [27,28]. Nevertheless, the distinction between superphylum and phylum has not yet been clearly described for this radiation. Moreover, as there is no proposed formal name for this division, the misuse of “Patescibacteria” as a synonym for CPR is confusing when addressing a superphylum/phylum particularity without generalizing it to the whole CPR group.
2. CPR Genomes: Discovery of a New Division Is Challenging for Evolutionists
The first sequence of CPR was detected in 1996 [29]. It was discovered by detecting a 350 bp fragment of a 16S ribosomal subunit (rRNA) of a TM7 (Torf, mittlere Schicht, clone 7, or “peat, middle layer, clone 7”) [29]. It was then inserted in a phylogenetic tree analysis. TM7 is now referred to as Candidatus Saccharibacteria superphylum, due to its sugar metabolism, after the first complete genome was revealed in 2007 [30].
Two years after a CPR sequence was first detected, in 1998, the 16S rRNA phylogenetic tree revealed CPR sequences as a distinct phylum independent of archaea and eukaryotes but related to bacteria (Figure 1) [31]. Due to significant divergent genetic sequences, some studies have shown the inadequacy of using 16S rRNA for CPR classification [7,32]. However, the use of other conserved coding genes, such as ribonucleotide reductase (RNR), topoisomerase IIA (Topo IIA), elongation factor 1 (EF1), DNA-dependent RNA polymerase subunit II (RNAP II), thymidylate synthase (ThyA), and DNA polymerase, has ultimately resulted in a very similar classification of CPR genomes [32,33]. Thus, a recent genomic analysis conducted by Megrian et al. showed the conserved presence of division and cell wall (dcw) genes with the necessary order (synteny) in over 1000 bacterial genomes, including CPRs [34]. Based on the concatenation of this gene cluster, phylogenetic trees have shown consistency with previous phylogenies in which dcw genes can be used as alternative markers to produce a robust phylogenetic tree (Figure 1) [34].
CPRs may have grouped together, as they share the protein profile (presence and absence) of certain protein groups in comparison to other domains of life, although they were directly branched as a deep ramification of the bacterial kingdom (Figure 1) [12,35,36,37]. Therefore, CPR cannot be regarded as a novel domain but rather as a superphylum of bacteria, since their contribution of only 1.22% to the repertoire of bacterial proteins was insufficiently significant [37]. Studies using phylogenomic trees based on single or multiple genes have shown that CPR and bacteria are two paraphyletic groups and not two distinct divisions (Figure 1 and Table 1) [36,37]. This model is currently a matter of debate. In contrast, rhizome-based classification, based on the total genome sequences and taking into consideration sequences generated by processes such as transfers, recombination, fusion, degradation, and de novo synthesis, has shown that CPR is a genetically independent division [38]. This classification also takes ORFans into account, which are genes that are specific to a certain species and have an unknown evolutionary identity. The rhizomes of CPR were mosaic and composed of sequences from heterogeneous origins, unlike the homogeneous profile of those of bacteria [38]. These differences in rhizomal profiles confirm the particularity of the historical evolution of CPR genomes.
One study in which CPR was distinguished based on protein family content suggested two possibilities regarding how CPRs have evolved [36]. The first possibility is that CPR and non-CPR bacteria could have co-evolved after emerging from the protogenote community, and CPR genomes subsequently diversified over time. The second possibility is that CPRs could have evolved from non-CPR bacteria after having undergone a severe genome reduction [36]. The latter scenario has not been supported by numerous findings, mainly since CPRs do not share genomic features with recent genome reduction organisms (Figure 1 and Table 1) [39,40]. CPRs have thus demonstrated the capacity to maintain the integrity of their genomes by having genes involved in DNA mismatch repair and homologous recombination pathways (such as Holliday junction resolvases) [9]. Moreover, CPRs exhibit active metabolic patterns that aggregate independently of another metabolically reduced symbiont (Figure 1) [7,12,24,41,42]. Therefore, the uniformly small cell size and genome of CPRs were found to be ancestral characteristics of their group rather than the result of starvation.
Through the detection of CPR sequences by metagenomic analysis, these microorganisms were identified as a new microbial discovery, the definition of which changes from one analysis to another. However, CPRs are distinct and divergent from other groups of life (Figure 1). Their taxonomy and classification represent a continuous challenge for microbiologists. Consequently, their placement within the tree of life is not yet well-established, and their evolution remains a mystery to be elucidated.
3. Distinct Biology of CPR Cells
3.1. Common and Unusual Genomic Features of CPRs
CPRs are characterized by the presence of a single copy of 16S rRNA with self-splicing introns and protein insertions that are rarely reported in classical bacteria (Figure 1 and Table 1) [7]. This has also been shown in sequences of 23S rRNA and tRNA [9,43]. Due to these intron sequences, CPRs have larger rRNA than known bacteria, but are expected to be similar in length after transcription (Table 1) [44,45]. Moreover, genomic analysis of CPR ribosomes revealed missing protein regions on their surface structure. This was also the case for CPR rRNA (23S rRNA, 5S rRNA, and 16S rRNA) (Figure 1 and Table 1) [44]. Thus, the absence of a few proteins in CPR ribosomes was revealed by genomic comparisons such as rpL30, uL1, uL30, and bL9, which are usually found in bacteria [44]. This suggests that CPR ribosomal proteins have distinctive compositions compared to those of classical bacteria. Furthermore, in some CPR genomes of different phyla (such as Parcubacteria, Saccharibacteria, Microgenomates, and Katanobacteria), a lack of rpL1, rpL9, rpL25, uL29, bL28, bL32, and bL33 has been identified [44]. Some of these ribosomal proteins are universal in bacteria and are involved in essential processes of the formation, growth, and regulation of ribosome subunits [7]. All these heterogeneities of proteins in ribosome structures point to diverse trajectories of ribosome evolution between bacteria and CPRs and across CPR lineages [44]. It is thought, therefore, that CPRs probably have analogous proteins and/or alternatives to fulfil the functions of those that are missing.
Genomes of CPR have shown fewer open reading frames (ORFs) than other bacteria, which indicates a correlation between bacterial genome size and the number of encoded proteins (Table 1) [45]. Furthermore, based on an analysis of a massive metagenomic dataset, CPR genomes were found to harbor diversity-generating retroelements (DGRs) [46]. DGRs are mobile elements that diversify DNA sequences by introducing nucleotide substitutions at specific locations, thereby creating different protein-encoding genes [47]. They were found to be abundant in microorganisms with small genomes and episymbiotic lifestyles, such as CPRs [48]. Their function expands ligand specificity for different cellular binding proteins involved in attachment with host cells [28]. This process could accelerate the evolution of CPRs to exploit potential hosts in different environments. They may be a result of a myriad of selective pressures to impose genetic hyper-variability, which is particularly advantageous for adaptation (Figure 1) [47,49,50].
A high ribosome copy number is considered to be a driver of rapid growth, suggesting that multiple rRNA gene-carrying microbes are generally more efficient in responding to resource availability (Figure 1 and Table 1) [51]. However, CPRs with a single copy of the 16S rRNA call upon multiple mechanisms in life-and-death struggles, providing prodigious levels of diversity. From splicing introns in their rRNA to DGRs in their genomes, these different genetic diversification mechanisms enable CPR cells to adapt to various environmental conditions (such as limitations on energy or nutrients).
3.2. Metabolic Pathways: CPRs Need to Do More with Less
Little is currently understood about the metabolic activity of CPRs, since most of their genomes are incomplete, which prevents a total/complete in silico analysis. Thus, CPRs with no pure culture to date lack isolated representatives, hence in vitro analysis of their metabolic pathways is not possible (Figure 1 and Table 1). However, CPRs are characterized by their obligate exo-symbiotic/parasitic lifestyle in which their survival depends on a host (most often anaerobic bacteria) (Table 1). This dependency is variable between CPR lineages, as they have different biosynthetic capacities. Almost all CPRs have shown the inability to autonomously synthesize nucleotides (de novo), amino acids, and membrane components (Figure 1) [7,13,15,52]. However, the analysis of five genomes of the Peregrinibacteria phylum revealed a semi-independent lifestyle toward its host [53]. They showed fuller metabolic pathways than other CPRs, with the ability to significantly synthesize cell wall components, including peptidoglycan, isoprenoids through a near-complete mevalonate pathway, and other elements [53]. However, this absence of recognized/essential processes in all CPR cells has made them polylithic in terms of metabolism and auxotrophic microorganisms (Figure 1 and Table 1).
CPRs lack the necessary pathways to synthesize lipopolysaccharide (LPS) and produce fatty acids [9,54]. They appear to lack the ability to synthesize lipid A, which is necessary for a Gram-stain-negative cell envelope [55]. However, multiple studies anticipate cell-wall-containing peptidoglycan, in which all required genes for its biosynthesis were identified (Figure 1 and Table 1) [9,53]. Hence, microscopic observations were consistent, in that CPR cells have a Gram-stain-positive-like membrane [13]. They were thus found to encode large extracellular proteins rich in cysteine, which may function in cell attachment [53]. Moreover, genomic analysis of the Saccharibacteria and Absconditabacteria genomes has revealed an interesting pattern for several metabolites, with high PM values (quantifying metric for biosynthetic capabilities) [56]. These metabolites were mostly components of the CPR cell wall [56].
Based on genome analysis, CPRs have shown an anaerobic respiratory lifestyle with metabolic patterns that match the Earth’s early anaerobic environment [57]. Most CPRs have the glycolysis Embden-Meyerhof-Parnas (EMP) and the pentose phosphate pathway (PPP), whereas downstream biosynthetic pathways were absent for the PPP [57]. Thus, they have neither an electron transport chain, nor tricarboxylic acid (TCA) cycle, nor subunits of NADH dehydrogenase (Figure 1) [57]. The absence of these essential compounds suggests a fermentation-based metabolism, especially given that CPRs have shown the synthesis of acetate, lactate, ethanol, and hydrogen as fermentation end products with missing crucial elements of respiratory pathways [57]. However, a particular CPR genome known as “Candidatus Parcunitrobacter nitroensis” has revealed a complete electron transfer chain. This genome contained all genes involved in the nitrogen-respiration process (hydroxylamine redox enzyme, nitrate reductase, and nitric oxide reductase) [58]. Moreover, CPR genomes have shown the presence of nitrite reductase genes (nirK), key to the denitrification process (Figure 1) [16,59]. These are responsible for the nitrogen cycle, as they reduce nitrite to nitric oxide.
Furthermore, the analysis of CPR genomes has shown different glycoside hydrolase (GH) profiles/families [60]. This diversity allows carbon processing to a wide range of carbon substrates and complex sugars present in the environment (Figure 1) [61]. This dynamic behavior of different CPR phyla has also been shown through the presence of genes such as NiFe hydrogenases and iron-only hydrogenases involved in sulfur cycling [57]. Thus, despite the absence of sulfur respiration genes, CPRs can effectively participate in the sulfur cycle via hydrogenase-mediated sulfur reductase activity (Figure 1) [62]. Some other genomes have shown sulfur dioxygenase (sdo) and sulfate reduction genes (sat, cysC, and cysN) [63]. Through these processes, CPR organisms can play a vital role in nutrient cycling/secondary metabolism by breaking down complex organic molecules that other microorganisms are not capable of doing.
It is important to note that, as is the case with symbiotic cells, CPRs can vary their metabolic capabilities depending on the content of the niche and on the availability of what the host offers/provides. These distinctions have been shown by the content of enzyme-encoding genes with antibiotic resistance activity in CPR genomes detected in the human microbiomes being higher than those detected in environmental samples (Figure 1) [64]. It has also been shown through the detection of an arginine deiminase system (ADS) in Saccharibacteria genomes during the niche transition from the environment to mammalian microbiomes [65].
The diversity of metabolic capacities in widespread microorganisms suggests that CPRs are key members in driving critical elements via biogeochemical cycles for the microbe community [66], especially as CPRs have been found in various environments, including soil, freshwater, and marine systems (Figure 1). However, the different genes of metabolic pathways in CPR cells may function to obtain necessary nutrients and/or amino acids that are not encoded in their genomes, to capture energy, or to protect against a particular substrate in the niche they occupy. They may also play a regulatory role in the selection and attachment to the bacterial host membrane.
4. Beyond the Surface: Revealing the Diversity of Interactions and Signaling Mechanisms of CPRs in Microbial Ecosystems
The study of CPR is still in its early stages, and much remains to be learned about their interactions with other microbes. These microorganisms are small but well-equipped with genes that play a role in the functioning of ecosystems including the formation of biofilm, the production of virulence factors, and antibiotic resistance.
4.1. Chemical Signalling
CPR cells are involved in three modes of communication to maintain their interactions with other species in the environment: quorum sensing (QS), signal presence, and attentiveness to external signals. CPR organisms have been found to use quorum sensing to coordinate the expression of their genes in influencing their hosts/neighbors (Figure 1) [67]. Chemical signaling via QS is mediated by emitting small molecules known as autoinducers. When these autoinducers reach a threshold concentration, CPR can detect them and responds by altering its gene expression, depending on the type of autoinducer.
The rich repertoire of quorum-sensing proteins in CPRs was detected using a computational in silico analysis. Charles et al. showed that these protein-coding sequences are divergent and under strong selective pressure in CPR genomes [67]. The diversity of QS signals allows a wide range of interactions/communications between CPRs and other microorganisms. Thus, CPRs were involved in the biofilm formation of their bacterial host by emitting the AI-2 QS signal [68,69,70]. This surface-associated community allows resistance to a stressful stimulus due to the creation of a dense community of CPR/host in their niche [71,72]. Moreover, the abundance of some CPR cells has been correlated with the increase of virulence factors controlled by the QS signals/receptors system [15]. Some virulence factors, such as oligopeptide transport and iron acquisition, have been detected and correlated with the presence of the CPR phylum “Candidatus Saccharibacteria” in periodontal mucosal disease [73]. Several studies have shown that CPR phyla are associated with a wide range of pathologies including inflammatory, mucosal, and infectious diseases [74,75,76]. These microorganisms therefore have an impact on human health, as their abundance was recently highlighted in the human microbiome [77]. Different CPR phyla have been detected in the oral microbiota and include Candidatus Absconditabacteria (SR1), Candidatus Parcubacteria (OD1), Candidatus Microgenomates (OP11 and OP10), Candidatus Gracilibacteria (GN02) and, mainly, Candidatus Saccharibacteria [78,79]. These and others have also been detected in the gastrointestinal tract, blood, eyes, skin, and even in the genital tracts of both genders (Figure 1) [19,20,21,22,23]. However, it remains unclear how CPR cells affect the physiology of the human microbiome. Thus, a recent study was conducted on diapause larvae of Clanis bilineata tsingtauica to investigate the mechanisms by which gut microbiota regulate nutrient synthesis and metabolism in edible insects [80]. The study revealed that TM7 plays a crucial role in modulating intestinal metabolites by modulating metabolic pathways, leading to an increase in linolelaidic acid (LA) and a decrease in tricosanoic acid (TA) levels via intestinal enzymes [80].
Finally, these processes of production, detection and response allow CPR cells to contribute towards the functioning of the ecosystem in which they live. Furthermore, the combination of these different communication systems may allow for a more sophisticated form of interaction to convey different types of information. It helps CPR cells to coordinate their existence with their hosts/neighbors in order to adapt to various environments.
4.2. Defense Mechanisms
CPRs harbor enzyme-encoding genes with antibiotic resistance (AR) activity [64]. In 2021, an exhaustive in silico analysis using a new adapted strategy of AR gene research was applied to CPR genomes (n = 4062; all CPR genomes available on 12 September 2020 in the NCBI database) (Figure 1). According to the NCBI Conserved Domain Database (CDD) search, these AR genes have the functional domain necessary to inactivate the antibiotic and to protect or alter its target. This analysis has revealed a rich reservoir of 30,545 AR genes, with 89 enzyme activities in all CPR superphyla (Candidatus Parcubacteria, Candidatus Microgenomates, Candidatus Saccharibacteria, unclassified Patescibacteria group, Candidate division WWE3 (Katanobacteria), Candidatus Peregrinibacteria, Candidatus Berkelbacteria, Candidatus Dojkabacteria, Candidatus Doudnabacteria, Candidatus Gracilibacteria, Candidatus Absconditabacteria, Candidate division Kazan-3B-28, and Candidatus Wirthbacteria, with an average of 7.5 AR/genome [0 to 41]) [64]. They were associated with 14 antibiotic families, the most abundant of which are glycopeptides, beta-lactams, macrolide–lincosamide–streptogramin (MLS) resistance families, tetracycline, and aminoglycoside. Moreover, the activity of five beta-lactamase sequences from CPR have been confirmed in vitro using liquid chromatography-mass spectrometry [81]. These CPR proteins have shown the ability to hydrolyze different substrates of beta-lactam (amoxicillin, ampicillin, and penicillin G), even after the addition of clavulanic acid (beta-lactamase inhibitor) [81]. Thus, some of these proteins have shown an additional RNase activity in the presence and absence of EDTA chelator and sulbactam (beta-lactamase inhibitors) [81].
However, all these enzyme-encoding genes detected in CPR genomes may have not existed for AR activity as a main function, but rather to degrade/hydrolyze different compounds in their ecosystems. Hence, a functional in silico analysis of all uncharacterized CPR MBL sequences (n = 3349) revealed highly divergent sequences with many protein domain functions. This recent study revealed a multifunctional activity for CPR MBL when compared to bacterial sequences, which opens possibilities for CPR cells to adapt to different environments.
Furthermore, a comparative genomic analysis of CPR in activated sludge systems has provided insights into the adaptive response of CPRs to high oxygen conditions in aeration tanks. This was revealed by significantly enriched genes for oxygen stress resistance [66]. Additionally, numerous CPR genomes were found to harbor multidrug resistance genes, including multidrug efflux pump (LfrA) and methylenomycin A resistance protein (mmr) [66].
Most CPR genomes belonging to the Patescibacteria superphylum have shown an absence of phage receptors on their cell membranes, which could help protect them against phage invasion [82]. Thus, a meta-analysis conducted by Burstein et al. showed that some Parcubacteria and Microgenomates genomes can avoid these invasive genetic elements by having viral defense mechanisms such as the CRISPR-Cas system (clustered regularly interspaced short palindromic repeats, CRISPR-associated) (Figure 1) [83]. It should be noted that these systems have rarely been found in CPR cells, but when detected they showed an unexpected diversity in CPR genomes and were significantly distinct from those of bacteria [84,85,86]. However, few CPR genomes have been reported with phage infection [87]. The infection process has also been observed via cryogenic electron microscopy [13]. Moreover, analysis of CPR genomes has shown other phage defense systems such as the DISARM (defense island system associated with restriction–modification), and the BREX (Bacteriophage Exclusion) system [88]. A genomic analysis for Saccharibacteria genomes has shown the presence of restriction–modification systems to defend against foreign DNA [89]. In addition, it was shown that CPR cells can be resistant to heavy metals (cobalt–zinc–cadmium, copper, and arsenic) present in their habitat as well as resistant to ultraviolet light (Figure 1) [88].
4.3. Contact-Dependent Signaling Involves Direct Cell-to-Cell Interactions
One essential feature of CPRs is their obligate attachment to a bacterial host [77,90]. To date, this episymbiotic lifestyle of CPRs continues to be underexploited, although a few studies have shown a key role of type IV pili (T4P) (Figure 1) [13]. Type IV pili enable CPR cells to perform twitch-like movements that are essential for host recognition and cell adhesion in parasitic association [77]. Generally, T4P are ubiquitous in CPRs with different functions involved in processes such as protein secretion, and they exchange supplies (metabolites) with their host (Figure 1) [13,91]. This physical association provides easier DNA uptake (competence) and sequence transfer (lateral transfer) between the host and CPR cells, which have been shown to harbor genes from organisms living in the same habitat [92,93]. T4P can even act as nanowires carrying an electric current between the two cells [94]. Moreover, the pilin proteins of T4P are important components of the extracellular matrix which is crucial for maximal biofilm formation. Thus, it is considered a valuable factor in virulence mechanisms when pili biosynthesis via QS is correlated with an increase in TM7 in the oral microbiota of patients with periodontitis disease [73].
5. Conclusions
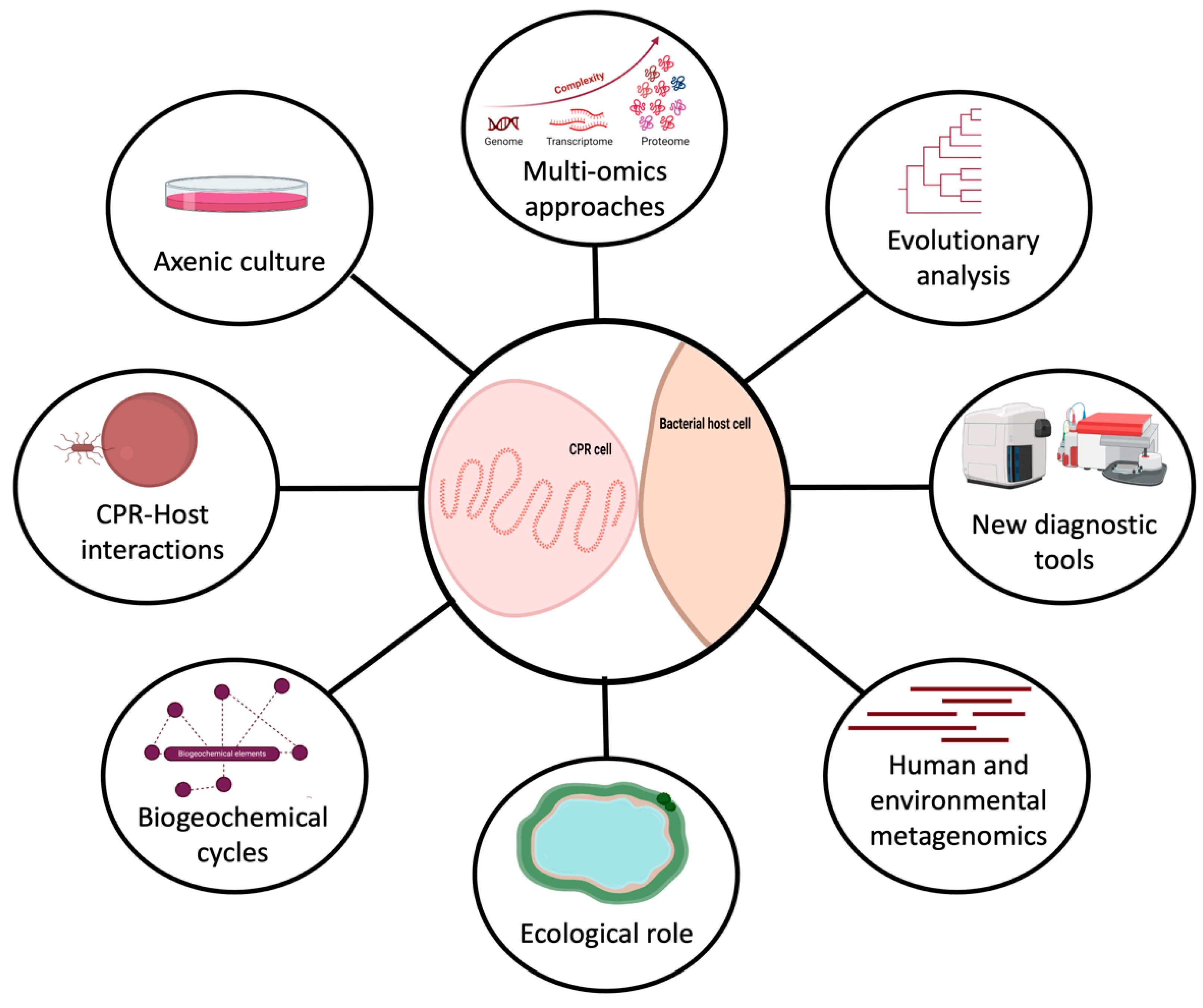
One interesting aspect of CPR organisms is that they have been found to have a high degree of genetic diversity and limited, but divergent, metabolic capacities that make them able to colonize a wide range of environments. These minimalist microbes have the potential to serve as useful model organisms for studying adaptation in various ecological niches. They are small but powerful microorganisms that can do more than expected with minimum genetic content. There is a critical need to improve techniques to enable fast and comprehensive analysis of their metabolic pathways, especially in order to access the biologically useful context that can be extracted from this information (Figure 2). CPRs hold enormous potential for biological and medical discoveries, providing valuable insights into the diversity and complexity of life on Earth.
CPRs should be an essential target for future research to address microbial ecology problems, such as nutrient cycling competition, cross-feeding, community dynamics resulting from microbe predation, and antagonistic interactions between microorganisms (Figure 2). Their interactions with other organisms significantly impact various human activities, including agriculture, water treatment, etc. (Figure 2) [95]. The study of these ecological phenomena requires isolated strains, as direct observation of natural ecosystems is much more challenging due to the higher complexity and lack of experimental control. For instance, around half of the genes in most CPR genomes have no known function. This suggests that studying these microorganisms could yield significant opportunities for scientific discovery. As technology continues to improve, we can therefore expect to isolate these fastidious microbes and discover even more about the roles they play in their environments (Figure 2).
Author Contributions
M.M. wrote the manuscript. J.-M.R. and F.B. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the French Government under the “Investissements d’avenir” (Investments for the Future) programme managed by the Agence Nationale de la Recherche (ANR, fr: National Agency for Research), (reference: Méditerranée Infection 10-IAHU-03). This work was supported by Région Provence Alpes Côte d’Azur and European funding (FEDER (Fonds européen de développement régional) PRIMMI (Plateformes de Recherche et d’Innovation Mutualisées Méditerranée Infection)).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ms, R.; Sj, G. The Uncultured Microbial Majority. Annu. Rev. Microbiol. 2003, 57, 369–394. [Google Scholar] [CrossRef]
- Chen, S.-C.; Budhraja, R.; Adrian, L.; Calabrese, F.; Stryhanyuk, H.; Musat, N.; Richnow, H.-H.; Duan, G.-L.; Zhu, Y.-G.; Musat, F. Novel Clades of Soil Biphenyl Degraders Revealed by Integrating Isotope Probing, Multi-Omics, and Single-Cell Analyses. ISME J. 2021, 15, 3508–3521. [Google Scholar] [CrossRef]
- Costa, O.Y.A.; de Hollander, M.; Pijl, A.; Liu, B.; Kuramae, E.E. Cultivation-Independent and Cultivation-Dependent Metagenomes Reveal Genetic and Enzymatic Potential of Microbial Community Involved in the Degradation of a Complex Microbial Polymer. Microbiome 2020, 8, 76. [Google Scholar] [CrossRef]
- Anantharaman, K.; Brown, C.T.; Hug, L.A.; Sharon, I.; Castelle, C.J.; Probst, A.J.; Thomas, B.C.; Singh, A.; Wilkins, M.J.; Karaoz, U.; et al. Thousands of Microbial Genomes Shed Light on Interconnected Biogeochemical Processes in an Aquifer System. Nat. Commun. 2016, 7, 13219. [Google Scholar] [CrossRef]
- Nakai, R. Size Matters: Ultra-Small and Filterable Microorganisms in the Environment. Microbes Environ. 2020, 35, ME20025. [Google Scholar] [CrossRef] [PubMed]
- Rinke, C.; Schwientek, P.; Sczyrba, A.; Ivanova, N.N.; Anderson, I.J.; Cheng, J.-F.; Darling, A.; Malfatti, S.; Swan, B.K.; Gies, E.A.; et al. Insights into the Phylogeny and Coding Potential of Microbial Dark Matter. Nature 2013, 499, 431–437. [Google Scholar] [CrossRef]
- Brown, C.T.; Hug, L.A.; Thomas, B.C.; Sharon, I.; Castelle, C.J.; Singh, A.; Wilkins, M.J.; Wrighton, K.C.; Williams, K.H.; Banfield, J.F. Unusual Biology across a Group Comprising More than 15% of Domain Bacteria. Nature 2015, 523, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Achtman, M.; Wagner, M. Microbial Diversity and the Genetic Nature of Microbial Species. Nat. Rev. Microbiol. 2008, 6, 431–440. [Google Scholar] [CrossRef]
- Castelle, C.J.; Brown, C.T.; Anantharaman, K.; Probst, A.J.; Huang, R.H.; Banfield, J.F. Biosynthetic Capacity, Metabolic Variety and Unusual Biology in the CPR and DPANN Radiations. Nat. Rev. Microbiol. 2018, 16, 629–645. [Google Scholar] [CrossRef]
- Eloe-Fadrosh, E.A.; Ivanova, N.N.; Woyke, T.; Kyrpides, N.C. Metagenomics Uncovers Gaps in Amplicon-Based Detection of Microbial Diversity. Nat. Microbiol. 2016, 1, 15032. [Google Scholar] [CrossRef]
- Wiegand, S.; Dam, H.T.; Riba, J.; Vollmers, J.; Kaster, A.-K. Printing Microbial Dark Matter: Using Single Cell Dispensing and Genomics to Investigate the Patescibacteria/Candidate Phyla Radiation. Front. Microbiol. 2021, 12, 635506. [Google Scholar] [CrossRef]
- Hug, L.A.; Baker, B.J.; Anantharaman, K.; Brown, C.T.; Probst, A.J.; Castelle, C.J.; Butterfield, C.N.; Hernsdorf, A.W.; Amano, Y.; Ise, K.; et al. A New View of the Tree of Life. Nat. Microbiol. 2016, 1, 16048. [Google Scholar] [CrossRef]
- Luef, B.; Frischkorn, K.R.; Wrighton, K.C.; Holman, H.-Y.N.; Birarda, G.; Thomas, B.C.; Singh, A.; Williams, K.H.; Siegerist, C.E.; Tringe, S.G.; et al. Diverse Uncultivated Ultra-Small Bacterial Cells in Groundwater. Nat. Commun. 2015, 6, 6372. [Google Scholar] [CrossRef] [PubMed]
- Dudek, N.K.; Sun, C.L.; Burstein, D.; Kantor, R.S.; Aliaga Goltsman, D.S.; Bik, E.M.; Thomas, B.C.; Banfield, J.F.; Relman, D.A. Novel Microbial Diversity and Functional Potential in the Marine Mammal Oral Microbiome. Curr. Biol. 2017, 27, 3752–3762.e6. [Google Scholar] [CrossRef] [PubMed]
- He, X.; McLean, J.S.; Edlund, A.; Yooseph, S.; Hall, A.P.; Liu, S.-Y.; Dorrestein, P.C.; Esquenazi, E.; Hunter, R.C.; Cheng, G.; et al. Cultivation of a Human-Associated TM7 Phylotype Reveals a Reduced Genome and Epibiotic Parasitic Lifestyle. Proc. Natl. Acad. Sci. USA 2015, 112, 244–249. [Google Scholar] [CrossRef]
- Danczak, R.E.; Johnston, M.D.; Kenah, C.; Slattery, M.; Wrighton, K.C.; Wilkins, M.J. Members of the Candidate Phyla Radiation Are Functionally Differentiated by Carbon- and Nitrogen-Cycling Capabilities. Microbiome 2017, 5, 112. [Google Scholar] [CrossRef] [PubMed]
- Orsi, W.D.; Richards, T.A.; Francis, W.R. Predicted Microbial Secretomes and Their Target Substrates in Marine Sediment. Nat. Microbiol. 2018, 3, 32–37. [Google Scholar] [CrossRef]
- Starr, E.P.; Shi, S.; Blazewicz, S.J.; Probst, A.J.; Herman, D.J.; Firestone, M.K.; Banfield, J.F. Stable Isotope Informed Genome-Resolved Metagenomics Reveals That Saccharibacteria Utilize Microbially-Processed Plant-Derived Carbon. Microbiome 2018, 6, 122. [Google Scholar] [CrossRef]
- Abou Chacra, L.; Fenollar, F. Exploring the Global Vaginal Microbiome and Its Impact on Human Health. Microb. Pathog. 2021, 160, 105172. [Google Scholar] [CrossRef]
- Villmones, H.C.; Haug, E.S.; Ulvestad, E.; Grude, N.; Stenstad, T.; Halland, A.; Kommedal, Ø. Species Level Description of the Human Ileal Bacterial Microbiota. Sci. Rep. 2018, 8, 4736. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The Skin Microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Fredricks, D.N.; Fiedler, T.L.; Marrazzo, J.M. Molecular Identification of Bacteria Associated with Bacterial Vaginosis. N. Engl. J. Med. 2005, 353, 1899–1911. [Google Scholar] [CrossRef] [PubMed]
- Kuehbacher, T.; Rehman, A.; Lepage, P.; Hellmig, S.; Fölsch, U.R.; Schreiber, S.; Ott, S.J. Intestinal TM7 Bacterial Phylogenies in Active Inflammatory Bowel Disease. J. Med. Microbiol. 2008, 57, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H. Recovery of Nearly 8,000 Metagenome-Assembled Genomes Substantially Expands the Tree of Life. Nat. Microbiol. 2017, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Schulz, F.; Eloe-Fadrosh, E.A.; Bowers, R.M.; Jarett, J.; Nielsen, T.; Ivanova, N.N.; Kyrpides, N.C.; Woyke, T. Towards a Balanced View of the Bacterial Tree of Life. Microbiome 2017, 5, 140. [Google Scholar] [CrossRef]
- Bacteria Candidate Phyla—Assembly—NCBI. Available online: https://www.ncbi.nlm.nih.gov/assembly/?term=bacteria+candidate+phyla (accessed on 2 March 2023).
- Gios, E.; Mosley, O.E.; Weaver, L.; Close, M.; Daughney, C.; Handley, K.M. Ultra-Small Bacteria and Archaea Exhibit Genetic Flexibility towards Groundwater Oxygen Content, and Adaptations for Attached or Planktonic Lifestyles. ISME Commun. 2023, 3, 13. [Google Scholar] [CrossRef]
- Castelle, C.J. Major New Microbial Groups Expand Diversity and Alter Our Understanding of the Tree of Life. Cell 2018, 172, 1181–1197. [Google Scholar] [CrossRef]
- Rheims, H.; Rainey, F.A.; Stackebrandt, E. A Molecular Approach to Search for Diversity among Bacteria in the Environment. J. Ind. Microbiol. Biotechnol. 1996, 17, 159–169. [Google Scholar] [CrossRef]
- Albertsen, M.; Hugenholtz, P.; Skarshewski, A.; Nielsen, K.L.; Tyson, G.W.; Nielsen, P.H. Genome Sequences of Rare, Uncultured Bacteria Obtained by Differential Coverage Binning of Multiple Metagenomes. Nat. Biotechnol. 2013, 31, 533–538. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of Culture-Independent Studies on the Emerging Phylogenetic View of Bacterial Diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar] [CrossRef]
- Meng, P.; Lu, C.; Lou, X.; Zhang, Q.; Jia, P.; Yan, Z.; Lin, J.; Chen, F. A Computational Protocol to Characterize Elusive Candidate Phyla Radiation Bacteria in Oral Environments Using Metagenomic Data. bioRxiv 2018, 358812. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.-A.; Hugenholtz, P. A Standardized Bacterial Taxonomy Based on Genome Phylogeny Substantially Revises the Tree of Life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Megrian, D.; Taib, N.; Jaffe, A.L.; Banfield, J.F.; Gribaldo, S. Ancient Origin and Constrained Evolution of the Division and Cell Wall Gene Cluster in Bacteria. Nat. Microbiol. 2022, 7, 2114–2127. [Google Scholar] [CrossRef]
- Jaffe, A.L.; Bardot, C.; Le Jeune, A.-H.; Liu, J.; Colombet, J.; Perrière, F.; Billard, H.; Castelle, C.J.; Lehours, A.-C.; Banfield, J.F. Variable Impact of Geochemical Gradients on the Functional Potential of Bacteria, Archaea, and Phages from the Permanently Stratified Lac Pavin. Microbiome 2023, 11, 14. [Google Scholar] [CrossRef]
- Méheust, R.; Burstein, D.; Castelle, C.J.; Banfield, J.F. The Distinction of CPR Bacteria from Other Bacteria Based on Protein Family Content. Nat. Commun. 2019, 10, 4173. [Google Scholar] [CrossRef] [PubMed]
- Bokhari, R.H.; Amirjan, N.; Jeong, H.; Kim, K.M.; Caetano-Anollés, G.; Nasir, A. Bacterial Origin and Reductive Evolution of the CPR Group. Genome Biol. Evol. 2020, 12, 103–121. [Google Scholar] [CrossRef]
- Ibrahim, A.; Colson, P.; Merhej, V.; Zgheib, R.; Maatouk, M.; Naud, S.; Bittar, F.; Raoult, D. Rhizomal Reclassification of Living Organisms. Int. J. Mol. Sci. 2021, 22, 5643. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A.; Wernegreen, J.J.; Moran, N.A.; Wernegreen, J.J. Lifestyle Evolution in Symbiotic Bacteria: Insights from Genomics. Trends Ecol. Evol. 2000, 15, 321–326. [Google Scholar] [CrossRef]
- Moran, N.A.; Bennett, G.M. The Tiniest Tiny Genomes. Annu. Rev. Microbiol. 2014, 68, 195–215. [Google Scholar] [CrossRef]
- Comolli, L.R.; Banfield, J.F. Inter-Species Interconnections in Acid Mine Drainage Microbial Communities. Front. Microbiol. 2014, 5, 367. [Google Scholar] [CrossRef]
- Brown, C.T.; Olm, M.R.; Thomas, B.C.; Banfield, J.F. Measurement of Bacterial Replication Rates in Microbial Communities. Nat. Biotechnol. 2016, 34, 1256–1263. [Google Scholar] [CrossRef]
- Kadnikov, V.V.; Mardanov, A.V.; Beletsky, A.V.; Karnachuk, O.V.; Ravin, N.V. Complete Genome of a Member of a New Bacterial Lineage in the Microgenomates Group Reveals an Unusual Nucleotide Composition Disparity Between Two Strands of DNA and Limited Metabolic Potential. Microorganisms 2020, 8, 320. [Google Scholar] [CrossRef]
- Tsurumaki, M.; Saito, M.; Tomita, M.; Kanai, A. Features of Smaller Ribosomes in Candidate Phyla Radiation (CPR) Bacteria Revealed with a Molecular Evolutionary Analysis. RNA 2022, 28, 1041–1057. [Google Scholar] [CrossRef]
- Tsurumaki, M.; Saito, M.; Maruyama, S.; Kanai, A. Importance of Candidate Phyla Radiation (CPR) Bacteria for the Origin of Life. J. Geogr. 2020, 129, 881–898. [Google Scholar] [CrossRef]
- Paul, B.G.; Burstein, D.; Castelle, C.J.; Handa, S.; Arambula, D.; Czornyj, E.; Thomas, B.C.; Ghosh, P.; Miller, J.F.; Banfield, J.F.; et al. Retroelement-Guided Protein Diversification Abounds in Vast Lineages of Bacteria and Archaea. Nat. Microbiol. 2017, 2, 17045. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Arambula, D.; Ghosh, P.; Miller, J.F. Diversity-Generating Retroelements in Phage and Bacterial Genomes. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.G.; Bagby, S.C.; Czornyj, E.; Arambula, D.; Handa, S.; Sczyrba, A.; Ghosh, P.; Miller, J.F.; Valentine, D.L. Targeted Diversity Generation by Intraterrestrial Archaea and Archaeal Viruses. Nat. Commun. 2015, 6, 6585. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.L.; Le Coq, J.; Hodes, A.; Barbalat, R.; Miller, J.F.; Ghosh, P. Selective Ligand Recognition by a Diversity-Generating Retroelement Variable Protein. PLoS Biol. 2008, 6, e131. [Google Scholar] [CrossRef]
- Handa, S.; Paul, B.G.; Miller, J.F.; Valentine, D.L.; Ghosh, P. Conservation of the C-Type Lectin Fold for Accommodating Massive Sequence Variation in Archaeal Diversity-Generating Retroelements. BMC Struct. Biol. 2016, 16, 13. [Google Scholar] [CrossRef]
- Bremer, H.; Dennis, P.P. Modulation of Chemical Composition and Other Parameters of the Cell at Different Exponential Growth Rates. EcoSal Plus 2008, 3, 1–49. [Google Scholar] [CrossRef]
- Kantor, R.S.; Wrighton, K.C.; Handley, K.M.; Sharon, I.; Hug, L.A.; Castelle, C.J.; Thomas, B.C.; Banfield, J.F. Small Genomes and Sparse Metabolisms of Sediment-Associated Bacteria from Four Candidate Phyla. mBio 2013, 4, e00708-13. [Google Scholar] [CrossRef]
- Anantharaman, K.; Brown, C.T.; Burstein, D.; Castelle, C.J.; Probst, A.J.; Thomas, B.C.; Williams, K.H.; Banfield, J.F. Analysis of Five Complete Genome Sequences for Members of the Class Peribacteria in the Recently Recognized Peregrinibacteria Bacterial Phylum. PeerJ 2016, 4, e1607. [Google Scholar] [CrossRef] [PubMed]
- Kowarsky, M.; Camunas-Soler, J.; Kertesz, M.; De Vlaminck, I.; Koh, W.; Pan, W.; Martin, L.; Neff, N.F.; Okamoto, J.; Wong, R.J.; et al. Numerous Uncharacterized and Highly Divergent Microbes Which Colonize Humans Are Revealed by Circulating Cell-Free DNA. Proc. Natl. Acad. Sci. USA 2017, 114, 9623–9628. [Google Scholar] [CrossRef]
- Sieber, C.M.K.; Paul, B.G.; Castelle, C.J.; Hu, P.; Tringe, S.G.; Valentine, D.L.; Andersen, G.L.; Banfield, J.F. Unusual Metabolism and Hypervariation in the Genome of a Gracilibacterium (BD1-5) from an Oil-Degrading Community. mBio 2019, 10, e02128-19. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.B.; Dewhirst, F.E.; Segre, D. Metabolic Network Percolation Quantifies Biosynthetic Capabilities across the Human Oral Microbiome. eLife 2019, 8, e39733. [Google Scholar] [CrossRef]
- Wrighton, K.C.; Thomas, B.C.; Sharon, I.; Miller, C.S.; Castelle, C.J.; VerBerkmoes, N.C.; Wilkins, M.J.; Hettich, R.L.; Lipton, M.S.; Williams, K.H.; et al. Fermentation, Hydrogen, and Sulfur Metabolism in Multiple Uncultivated Bacterial Phyla. Science 2012, 337, 1661–1665. [Google Scholar] [CrossRef] [PubMed]
- Castelle, C.J.; Brown, C.T.; Thomas, B.C.; Williams, K.H.; Banfield, J.F. Unusual Respiratory Capacity and Nitrogen Metabolism in a Parcubacterium (OD1) of the Candidate Phyla Radiation. Sci. Rep. 2017, 7, 40101. [Google Scholar] [CrossRef]
- León-Zayas, R.; Peoples, L.; Biddle, J.F.; Podell, S.; Novotny, M.; Cameron, J.; Lasken, R.S.; Bartlett, D.H. The Metabolic Potential of the Single Cell Genomes Obtained from the Challenger Deep, Mariana Trench within the Candidate Superphylum Parcubacteria (OD1). Environ. Microbiol. 2017, 19, 2769–2784. [Google Scholar] [CrossRef]
- Chiriac, M.-C.; Bulzu, P.-A.; Andrei, A.-S.; Okazaki, Y.; Nakano, S.; Haber, M.; Kavagutti, V.S.; Layoun, P.; Ghai, R.; Salcher, M.M. Ecogenomics Sheds Light on Diverse Lifestyle Strategies in Freshwater CPR. Microbiome 2022, 10, 84. [Google Scholar] [CrossRef]
- Vigneron, A.; Cruaud, P.; Langlois, V.; Lovejoy, C.; Culley, A.I.; Vincent, W.F. Ultra-Small and Abundant: Candidate Phyla Radiation Bacteria Are Potential Catalysts of Carbon Transformation in a Thermokarst Lake Ecosystem. Limnol. Oceanogr. Lett. 2020, 5, 212–220. [Google Scholar] [CrossRef]
- He, C.; Keren, R.; Whittaker, M.L.; Farag, I.F.; Doudna, J.A.; Cate, J.H.D.; Banfield, J.F. Genome-Resolved Metagenomics Reveals Site-Specific Diversity of Episymbiotic CPR Bacteria and DPANN Archaea in Groundwater Ecosystems. Nat. Microbiol. 2021, 6, 354–365. [Google Scholar] [CrossRef]
- Lavy, A.; McGrath, D.G.; Matheus Carnevali, P.B.; Wan, J.; Dong, W.; Tokunaga, T.K.; Thomas, B.C.; Williams, K.H.; Hubbard, S.S.; Banfield, J.F. Microbial Communities across a Hillslope-Riparian Transect Shaped by Proximity to the Stream, Groundwater Table, and Weathered Bedrock. Ecol. Evol. 2019, 9, 6869–6900. [Google Scholar] [CrossRef]
- Maatouk, M.; Ibrahim, A.; Rolain, J.-M.; Merhej, V.; Bittar, F. Small and Equipped: The Rich Repertoire of Antibiotic Resistance Genes in Candidate Phyla Radiation Genomes. mSystems 2021, 6, e00898-21. [Google Scholar] [CrossRef]
- Tian, J.; Utter, D.R.; Cen, L.; Dong, P.-T.; Shi, W.; Bor, B.; Qin, M.; McLean, J.S.; He, X. Acquisition of the Arginine Deiminase System Benefits Epiparasitic Saccharibacteria and Their Host Bacteria in a Mammalian Niche Environment. Proc. Natl. Acad. Sci. USA 2022, 119, e2114909119. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Hu, Y.; Liu, L.; Liu, S.-J.; Zhang, T. Genome-Centric Metagenomics Reveals the Host-Driven Dynamics and Ecological Role of CPR Bacteria in an Activated Sludge System. Microbiome 2023, 11, 56. [Google Scholar] [CrossRef]
- Bernard, C.; Lannes, R.; Li, Y.; Bapteste, É.; Lopez, P. Rich Repertoire of Quorum Sensing Protein Coding Sequences in CPR and DPANN Associated with Interspecies and Interkingdom Communication. mSystems 2020, 5, e00414-20. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Domenech, M.; García, E.; Moscoso, M. Biofilm Formation in Streptococcus Pneumoniae. Microb. Biotechnol. 2012, 5, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Domenech, M.; Ramos-Sevillano, E.; García, E.; Moscoso, M.; Yuste, J. Biofilm Formation Avoids Complement Immunity and Phagocytosis of Streptococcus Pneumoniae. Infect. Immun. 2013, 81, 2606–2615. [Google Scholar] [CrossRef]
- Bedree, J.K.; Bor, B.; Cen, L.; Edlund, A.; Lux, R.; McLean, J.S.; Shi, W.; He, X. Quorum Sensing Modulates the Epibiotic-Parasitic Relationship Between Actinomyces Odontolyticus and Its Saccharibacteria Epibiont, a Nanosynbacter Lyticus Strain, TM7x. Front. Microbiol. 2018, 9, 2049. [Google Scholar] [CrossRef] [PubMed]
- Soro, V.; Dutton, L.C.; Sprague, S.V.; Nobbs, A.H.; Ireland, A.J.; Sandy, J.R.; Jepson, M.A.; Micaroni, M.; Splatt, P.R.; Dymock, D.; et al. Axenic Culture of a Candidate Division TM7 Bacterium from the Human Oral Cavity and Biofilm Interactions with Other Oral Bacteria. Appl. Environ. Microbiol. 2014, 80, 6480–6489. [Google Scholar] [CrossRef] [PubMed]
- Duran-Pinedo, A.E.; Chen, T.; Teles, R.; Starr, J.R.; Wang, X.; Krishnan, K.; Frias-Lopez, J. Community-Wide Transcriptome of the Oral Microbiome in Subjects with and without Periodontitis. ISME J. 2014, 8, 1659–1672. [Google Scholar] [CrossRef]
- Huang, S.; Yang, F.; Zeng, X.; Chen, J.; Li, R.; Wen, T.; Li, C.; Wei, W.; Liu, J.; Chen, L.; et al. Preliminary Characterization of the Oral Microbiota of Chinese Adults with and without Gingivitis. BMC Oral Health 2011, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Kharitonova, M.; Vankov, P.; Abdrakhmanov, A.; Mamaeva, E.; Yakovleva, G.; Ilinskaya, O. The Composition of Microbial Communities in Inflammatory Periodontal Diseases in Young Adults Tatars. AIMS Microbiol. 2021, 7, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Bor, B.; Bedree, J.K.; Shi, W.; McLean, J.S.; He, X. Saccharibacteria (TM7) in the Human Oral Microbiome. J. Dent. Res. 2019, 98, 500–509. [Google Scholar] [CrossRef]
- Naud, S.; Ibrahim, A.; Valles, C.; Maatouk, M.; Bittar, F.; Tidjani Alou, M.; Raoult, D. Candidate Phyla Radiation, an Underappreciated Division of the Human Microbiome, and Its Impact on Health and Disease. Clin. Microbiol. Rev. 2022, 35, e0014021. [Google Scholar] [CrossRef]
- Ribeiro, A.A.; Paster, B.J. Dental Caries and Their Microbiomes in Children: What Do We Do Now? J. Oral Microbiol. 2023, 15, 2198433. [Google Scholar] [CrossRef]
- McLean, J.S.; Bor, B.; Kerns, K.A.; Liu, Q.; To, T.T.; Solden, L.; Hendrickson, E.L.; Wrighton, K.; Shi, W.; He, X. Acquisition and Adaptation of Ultra-Small Parasitic Reduced Genome Bacteria to Mammalian Hosts. Cell Rep. 2020, 32, 107939. [Google Scholar] [CrossRef]
- Qian, L.; Chen, B.-J.; Deng, P.; Gui, F.-R.; Cao, Y.; Qin, Y.; Liao, H.-J. TM7 (Saccharibacteria) Regulates the Synthesis of Linolelaidic Acid and Tricosanoic Acid, and Alters the Key Metabolites in Diapause Clanis Bilineata Tsingtauica. Front. Physiol. 2023, 14, 1093713. [Google Scholar] [CrossRef]
- Maatouk, M.; Ibrahim, A.; Pinault, L.; Armstrong, N.; Azza, S.; Rolain, J.-M.; Bittar, F.; Raoult, D. New Beta-Lactamases in Candidate Phyla Radiation: Owning Pleiotropic Enzymes Is a Smart Paradigm for Microorganisms with a Reduced Genome. Int. J. Mol. Sci. 2022, 23, 5446. [Google Scholar] [CrossRef]
- Tian, R.; Ning, D.; He, Z.; Zhang, P.; Spencer, S.J.; Gao, S.; Shi, W.; Wu, L.; Zhang, Y.; Yang, Y.; et al. Small and Mighty: Adaptation of Superphylum Patescibacteria to Groundwater Environment Drives Their Genome Simplicity. Microbiome 2020, 8, 51. [Google Scholar] [CrossRef]
- Burstein, D.; Sun, C.L.; Brown, C.T.; Sharon, I.; Anantharaman, K.; Probst, A.J.; Thomas, B.C.; Banfield, J.F. Major Bacterial Lineages Are Essentially Devoid of CRISPR-Cas Viral Defence Systems. Nat. Commun. 2016, 7, 10613. [Google Scholar] [CrossRef]
- Chen, L.-X.; Al-Shayeb, B.; Méheust, R.; Li, W.-J.; Doudna, J.A.; Banfield, J.F. Candidate Phyla Radiation Roizmanbacteria from Hot Springs Have Novel and Unexpectedly Abundant CRISPR-Cas Systems. Front. Microbiol. 2019, 10, 928. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Burstein, D.; Harrington, L.B.; Strutt, S.C.; Probst, A.J.; Anantharaman, K.; Thomas, B.C.; Doudna, J.A.; Banfield, J.F. New CRISPR–Cas Systems from Uncultivated Microbes. Nature 2017, 542, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Paez-Espino, D.; Eloe-Fadrosh, E.A.; Pavlopoulos, G.A.; Thomas, A.D.; Huntemann, M.; Mikhailova, N.; Rubin, E.; Ivanova, N.N.; Kyrpides, N.C. Uncovering Earth’s Virome. Nature 2016, 536, 425–430. [Google Scholar] [CrossRef]
- Kindler, G.S.; Wong, H.L.; Larkum, A.W.D.; Johnson, M.; MacLeod, F.I.; Burns, B.P. Metagenomic Insights into Ecosystem Function in the Microbial Mats of Blue Holes, Shark Bay. bioRxiv 2020. [Google Scholar] [CrossRef]
- Bor, B.; Collins, A.J.; Murugkar, P.P.; Balasubramanian, S.; To, T.T.; Hendrickson, E.L.; Bedree, J.K.; Bidlack, F.B.; Johnston, C.D.; Shi, W.; et al. Insights Obtained by Culturing Saccharibacteria with Their Bacterial Hosts. J. Dent. Res. 2020, 99, 685–694. [Google Scholar] [CrossRef]
- Ji, Y.; Zhang, P.; Zhou, S.; Gao, P.; Wang, B.; Jiang, J. Widespread but Poorly Understood Bacteria: Candidate Phyla Radiation. Microorganisms 2022, 10, 2232. [Google Scholar] [CrossRef]
- Méheust, R.; Burstein, D.; Castelle, C.J.; Banfield, J.F. Biological Capacities Clearly Define a Major Subdivision in Domain Bacteria. bioRxiv 2018. [Google Scholar] [CrossRef]
- Maier, B.; Wong, G.C.L. How Bacteria Use Type IV Pili Machinery on Surfaces. Trends Microbiol. 2015, 23, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.; Dubnau, D. DNA Uptake during Bacterial Transformation. Nat. Rev. Microbiol. 2004, 2, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Melville, S.; Craig, L. Type IV Pili in Gram-Positive Bacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 323–341. [Google Scholar] [CrossRef]
- Li, D.; Li, W.; Zhang, D.; Zhang, K.; Lv, L.; Zhang, G. Performance and Mechanism of Modified Biological Nutrient Removal Process in Treating Low Carbon-to-Nitrogen Ratio Wastewater. Bioresour. Technol. 2023, 367, 128254. [Google Scholar] [CrossRef] [PubMed]
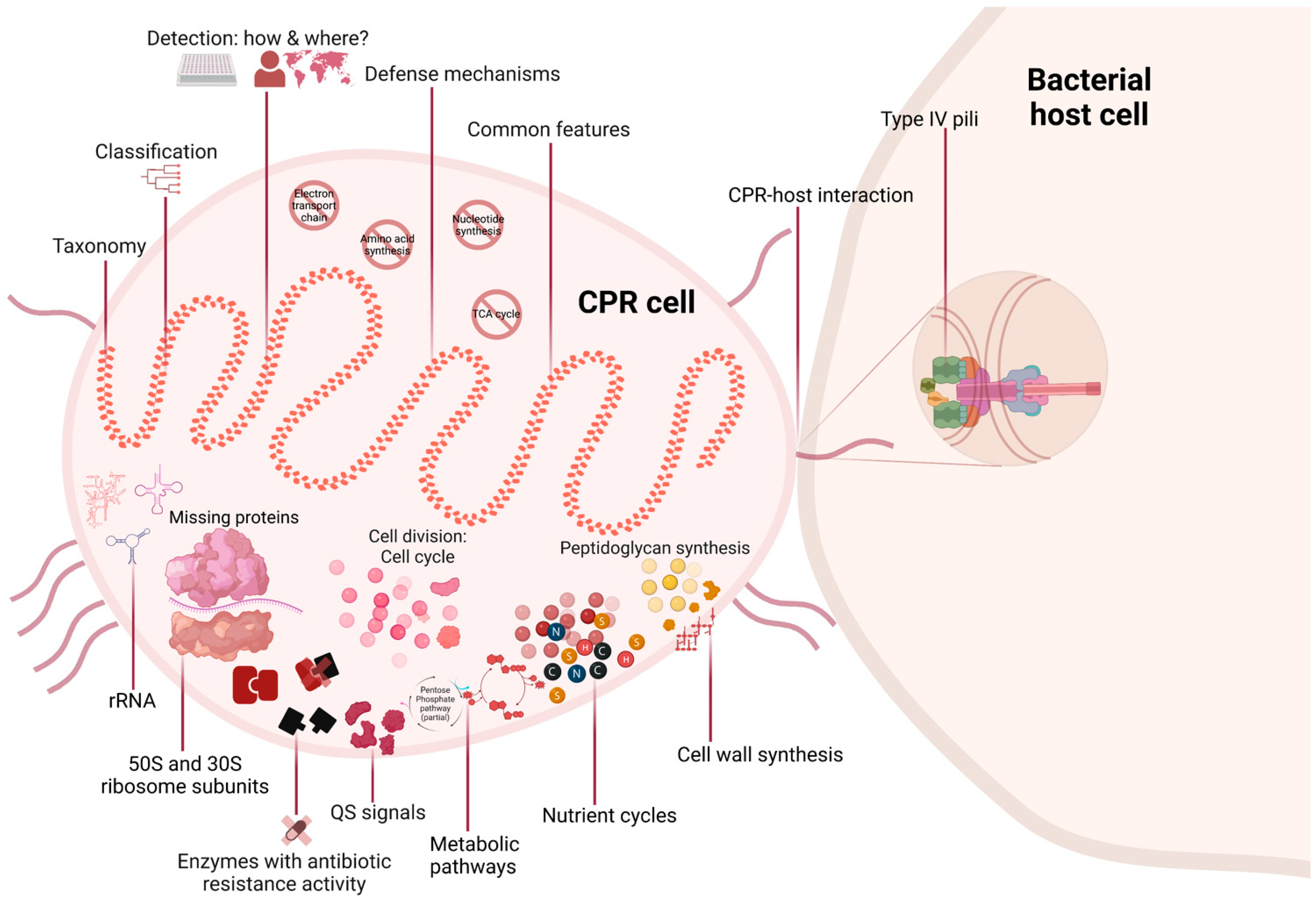
Figure 1.
Overview of all studied axes in this literature review (created with biorender.com).

Figure 2.
Representation of the future directions to study CPR microorganisms (created with biorender.com).
Figure 2.
Representation of the future directions to study CPR microorganisms (created with biorender.com).


{kind=link}
{kind=link}
Table 1.
Comparison of the most common features of CPRs and classical bacteria.
| General Characteristics | Candidate Phyla Radiation | Classical Bacteria |
|---|---|---|
| Genome size | Between 400 and 1500 kilobases | 4.6 Megabase |
| Cell size | Between 100 and 300 nm | Between 0.5 and 3 μm |
| Cell shape | Coccoid | Different cell shapes |
| Lifestyle | Exo-symbiosis or parasitism | Free living |
| Multiplication strategy | Cell division | Cell division |
| Cultured microorganism | Not yet | Yes |
| Genes number | 400 to 1500 | 4500 |
| Cell membrane | Gram+ | Gram+/Gram− |
| Copy of 16S rRNA | One | One or more |
| 16S rRNA gene length | Between 1400 and 4400 bases | 1500 bases |
| 23S rRNA gene length | 5000 bases | 3000 bases |
| 5S rRNA gene length | Between 107 and 117 bases | 120 bases |
| Presence of insertion sequences in RNA | Yes | No |
| Average protein length | 100 amino acids | 300 amino acids |
| Hypothetical protein number | Hight | Low |
| Protein coding genes number | 1000 | 5000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Maatouk, M.; Rolain, J.-M.; Bittar, F. Using Genomics to Decipher the Enigmatic Properties and Survival Adaptation of Candidate Phyla Radiation. Microorganisms 2023, 11, 1231. https://doi.org/10.3390/microorganisms11051231
AMA Style
Maatouk M, Rolain J-M, Bittar F. Using Genomics to Decipher the Enigmatic Properties and Survival Adaptation of Candidate Phyla Radiation. Microorganisms. 2023; 11(5):1231. https://doi.org/10.3390/microorganisms11051231
Chicago/Turabian StyleMaatouk, Mohamad, Jean-Marc Rolain, and Fadi Bittar. 2023. "Using Genomics to Decipher the Enigmatic Properties and Survival Adaptation of Candidate Phyla Radiation" Microorganisms 11, no. 5: 1231. https://doi.org/10.3390/microorganisms11051231
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.