

Microbiome of Ceca from Broiler Chicken Vaccinated or Not against Coccidiosis and Fed Berry Pomaces
 , ,
, ,
Abstract
:1. Introduction
2. Methodology
2.1. Berry Pomaces Preparation
2.2. Chicken Trial Design and Sample Collections
2.3. Genomic DNA Extraction and Metagenomics Data Analysis
2.4. Targeted-Resistome Sequencing
2.5. Data Analysis
3. Results
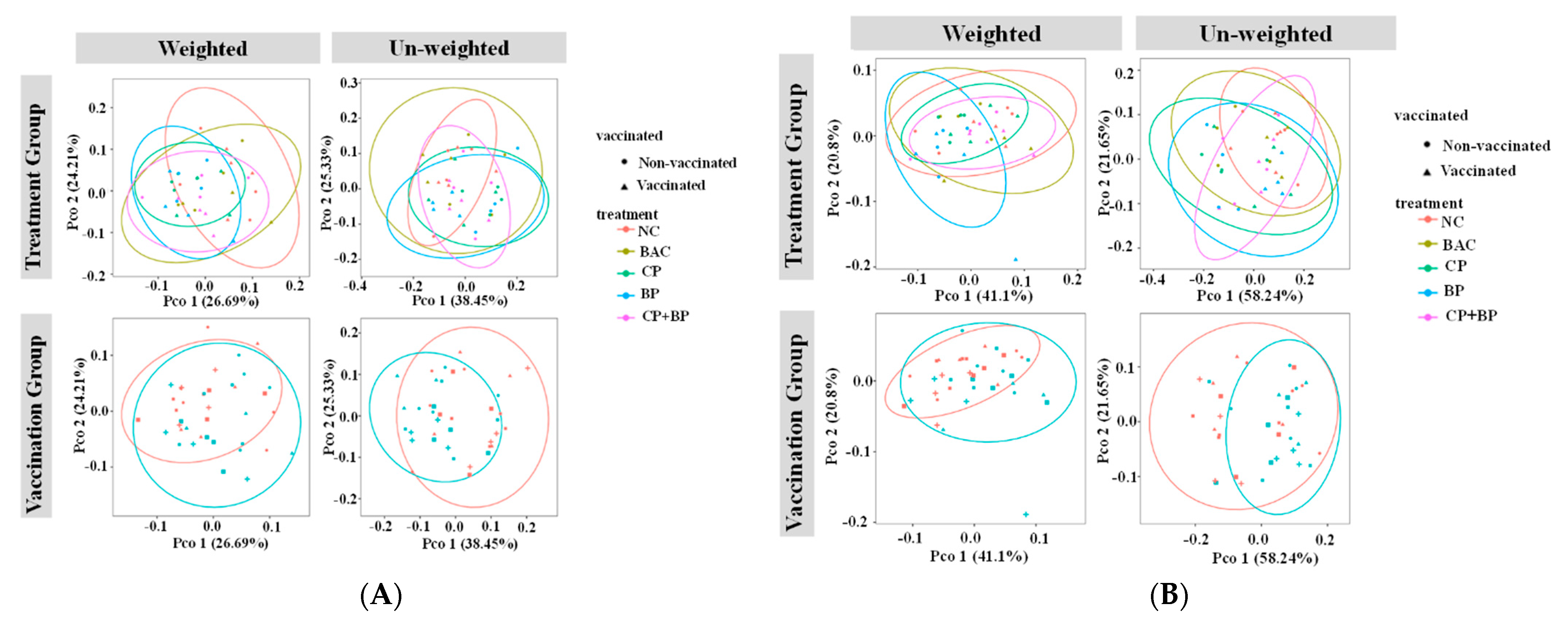
3.1. Relative Abundance of Cecal Microbiota by MetaPhlAn
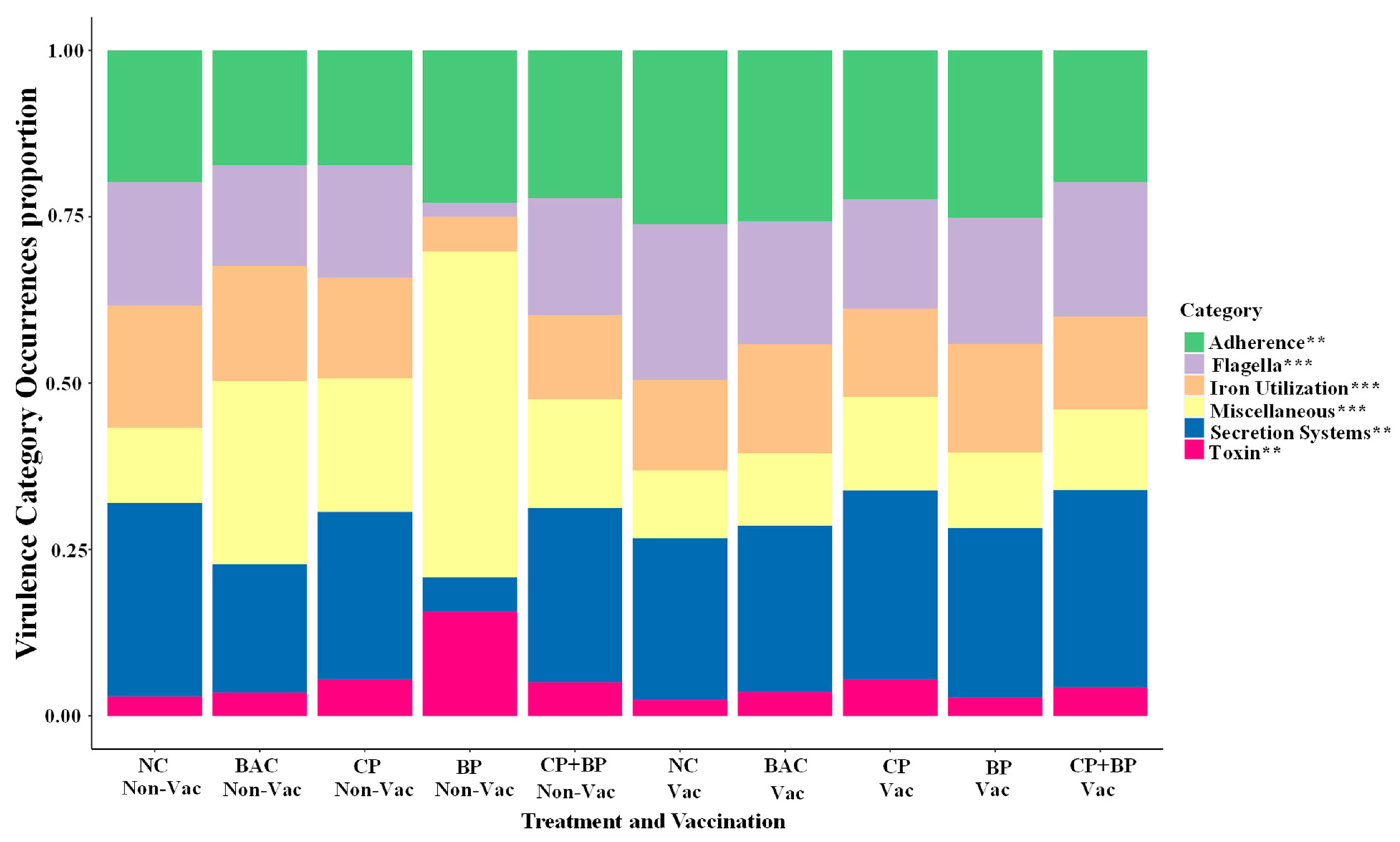
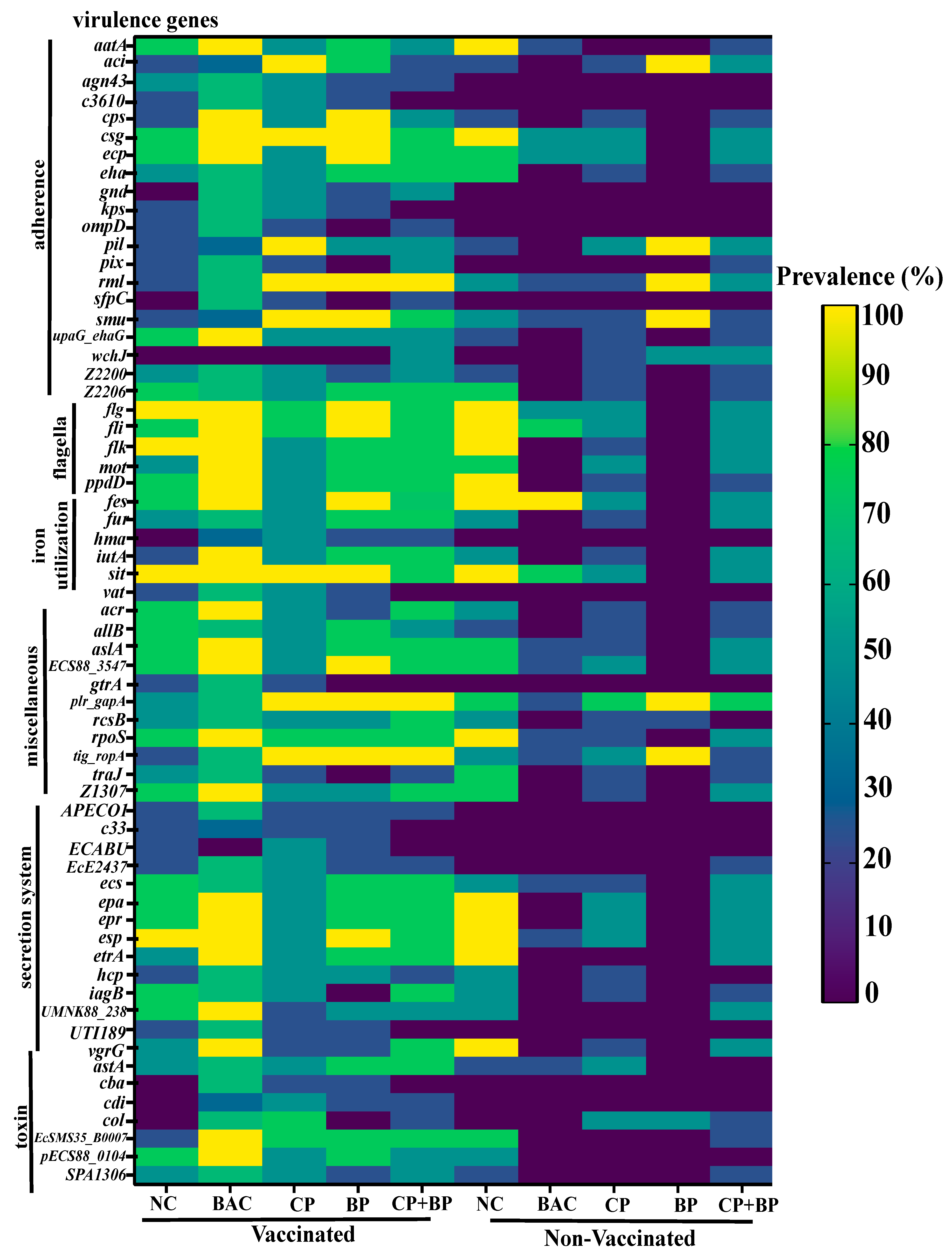
3.2. Virulence Genes (VGs) by SRST2
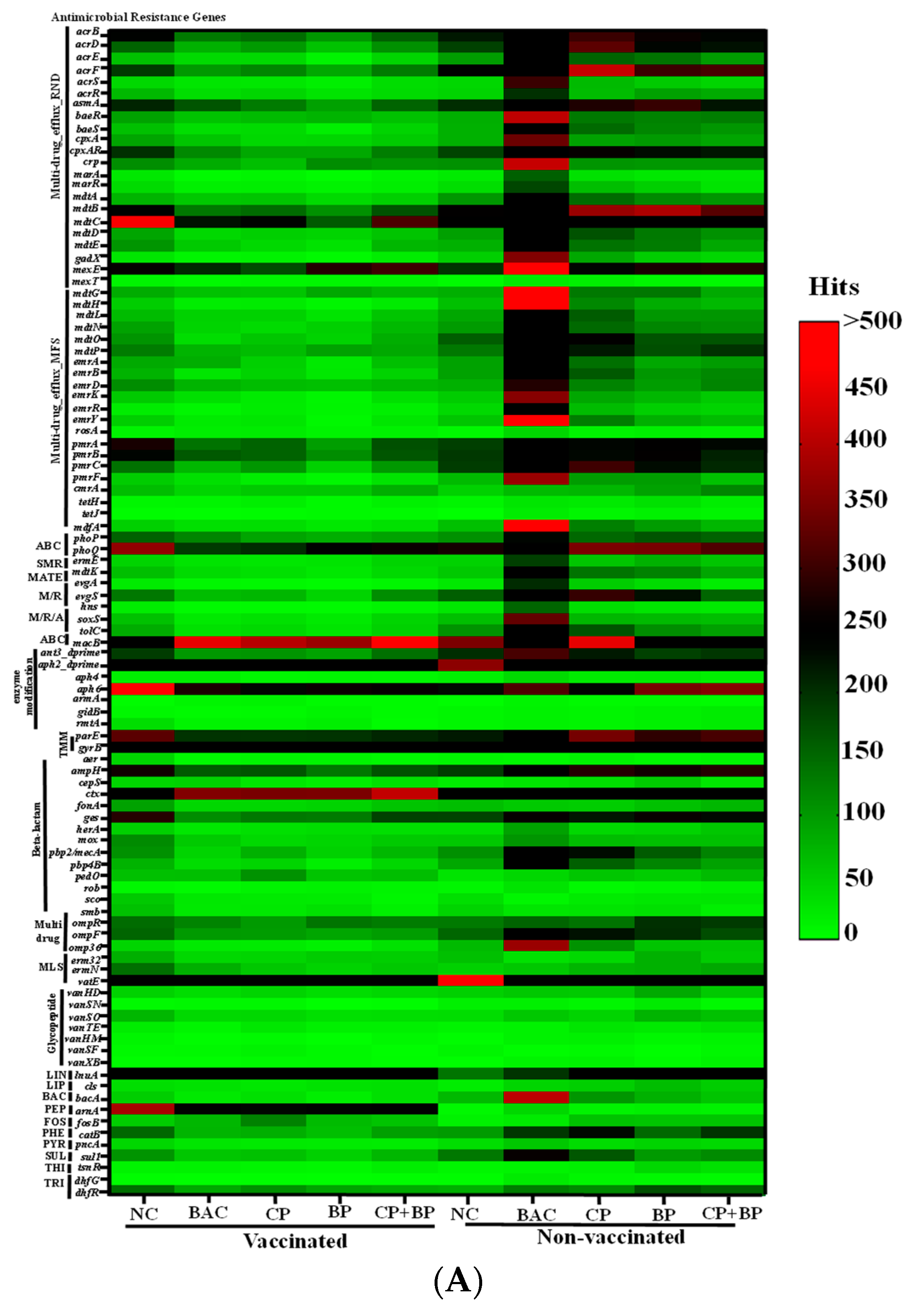
3.3. Antimicrobial Resistant Genes from Shotgun Metagenomics
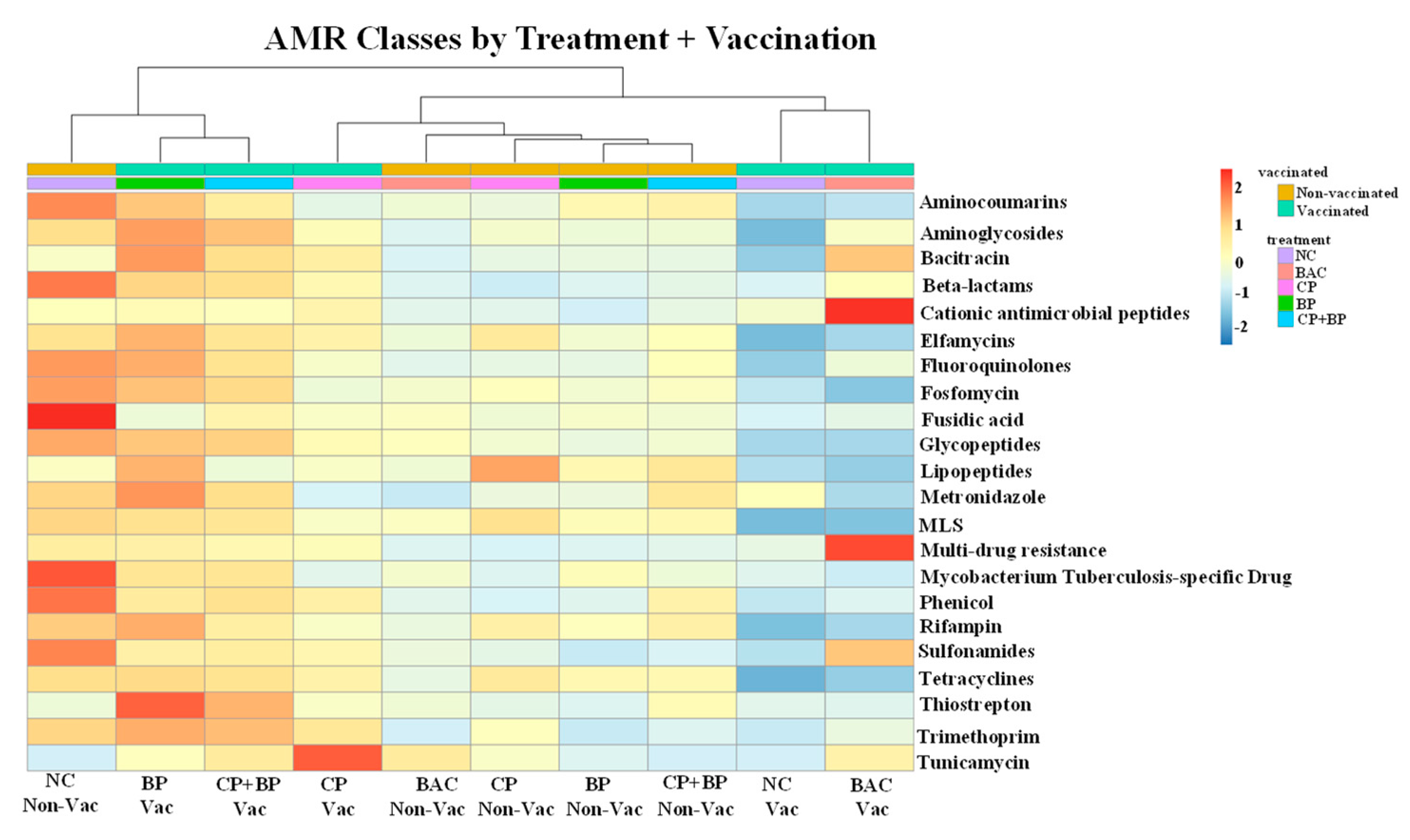
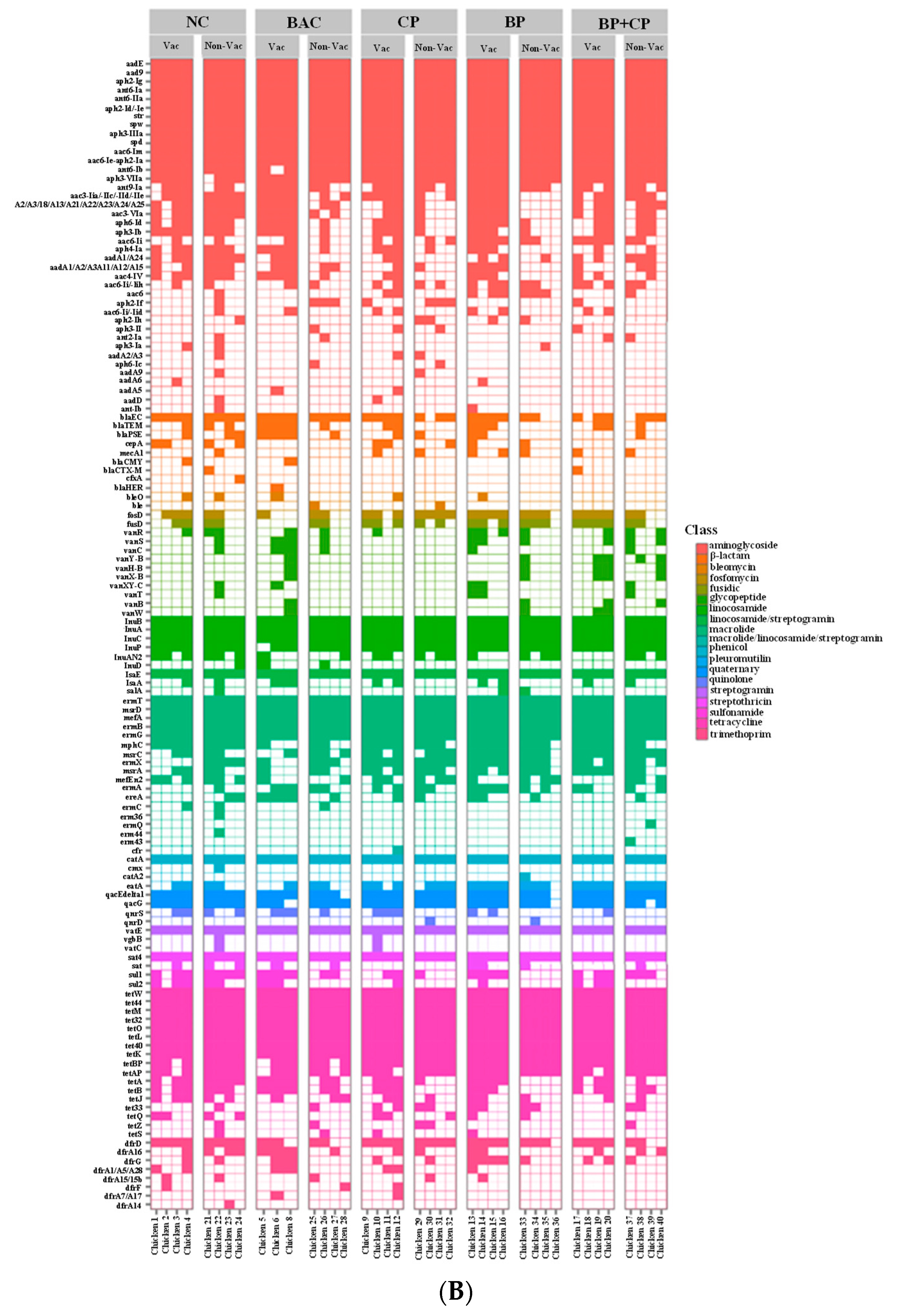
3.4. Antimicrobial Resistance Genes by Targeted-Resistome Sequencing
3.5. Correlations between Microbial Genera and ARGs
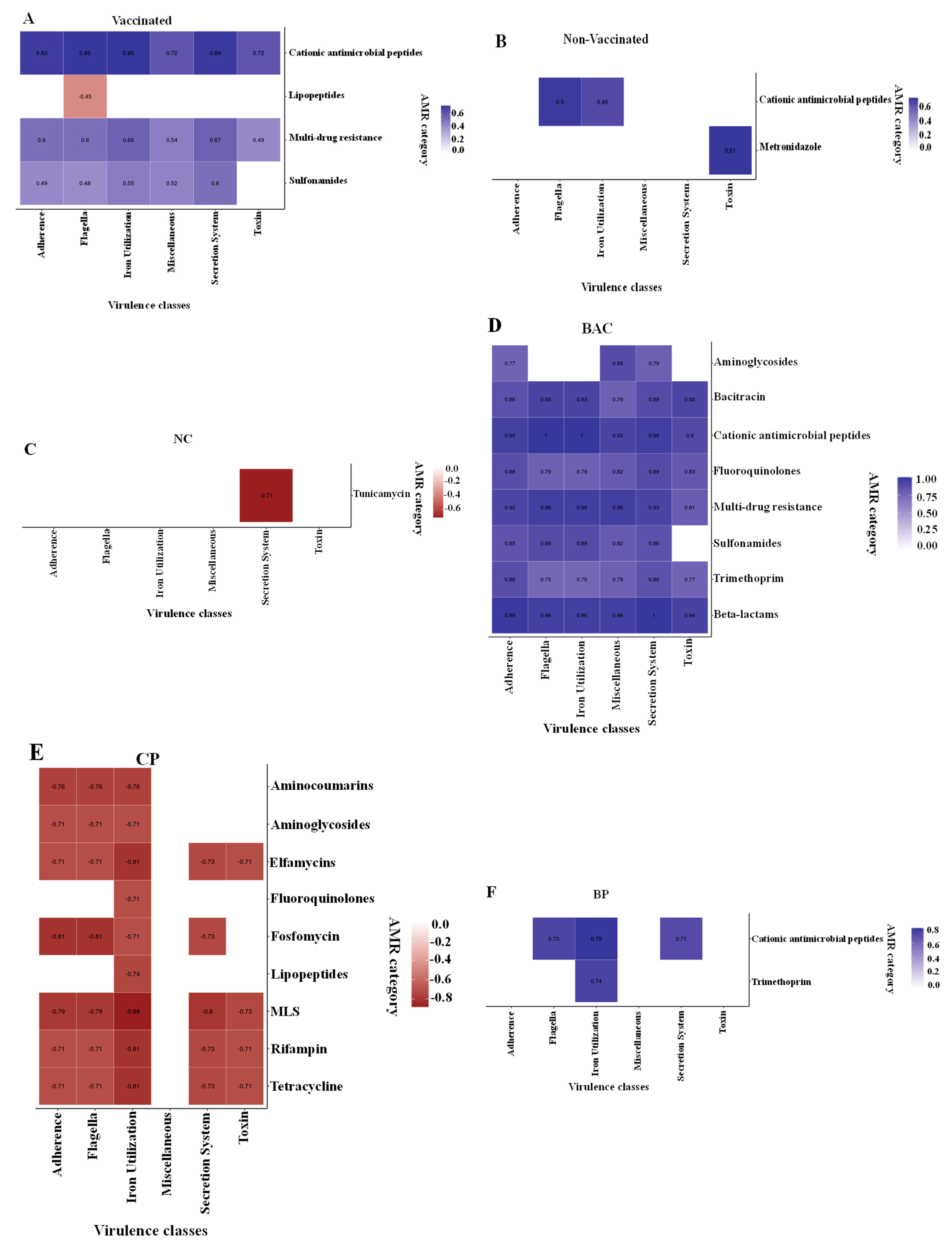
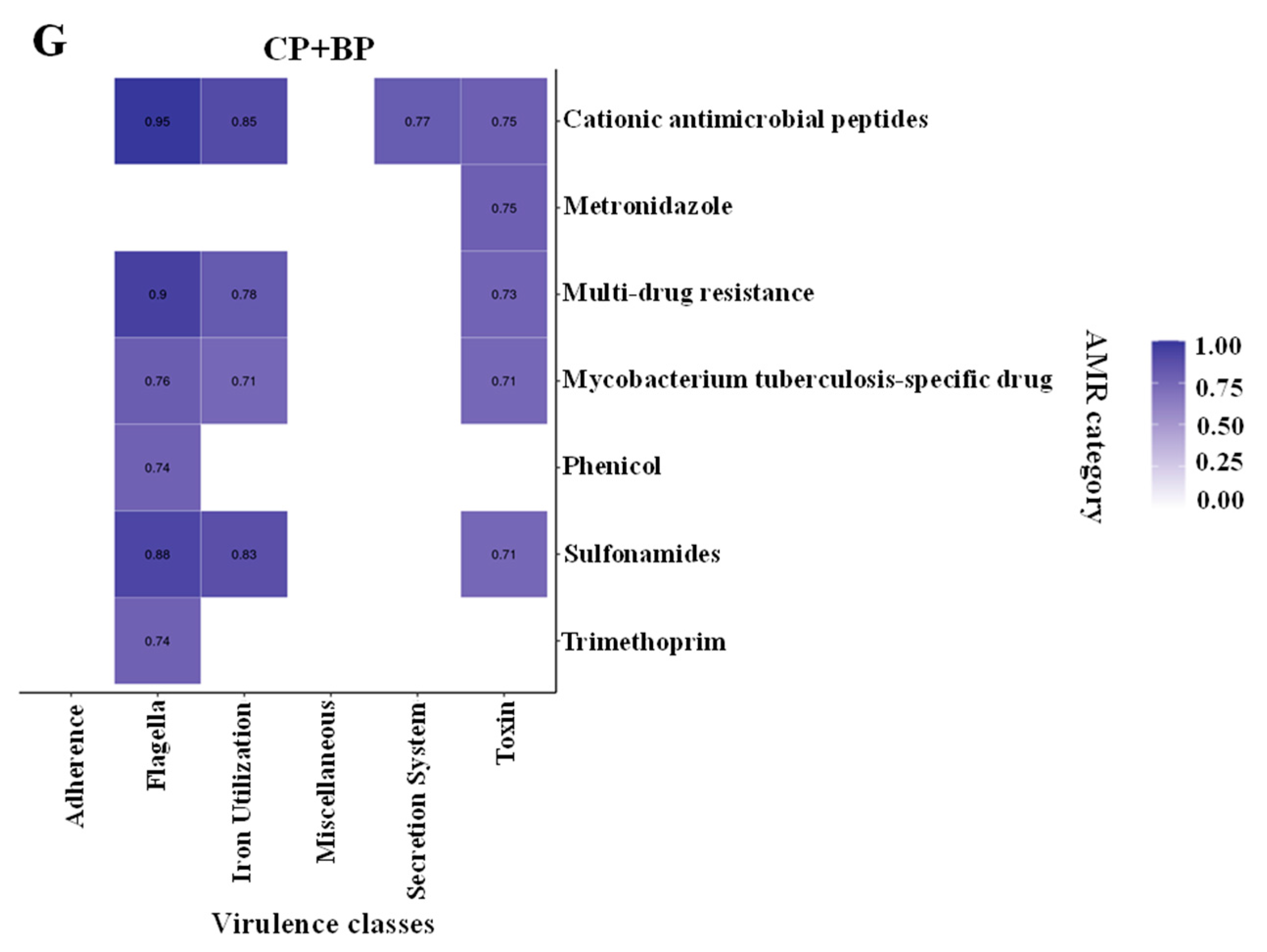
3.6. Correlations between Virulence Categories and ARGs
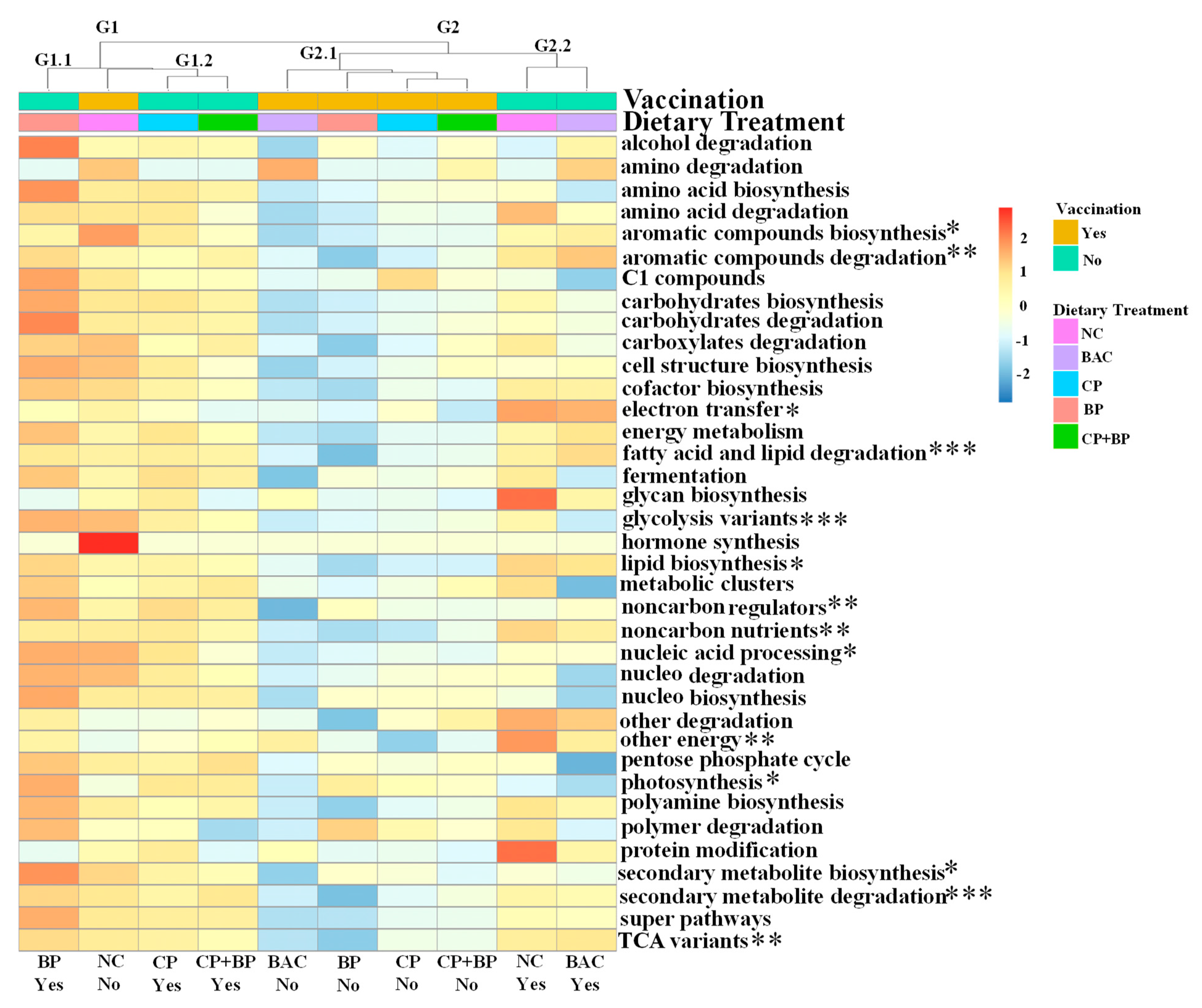
3.7. Metabolic Pathways
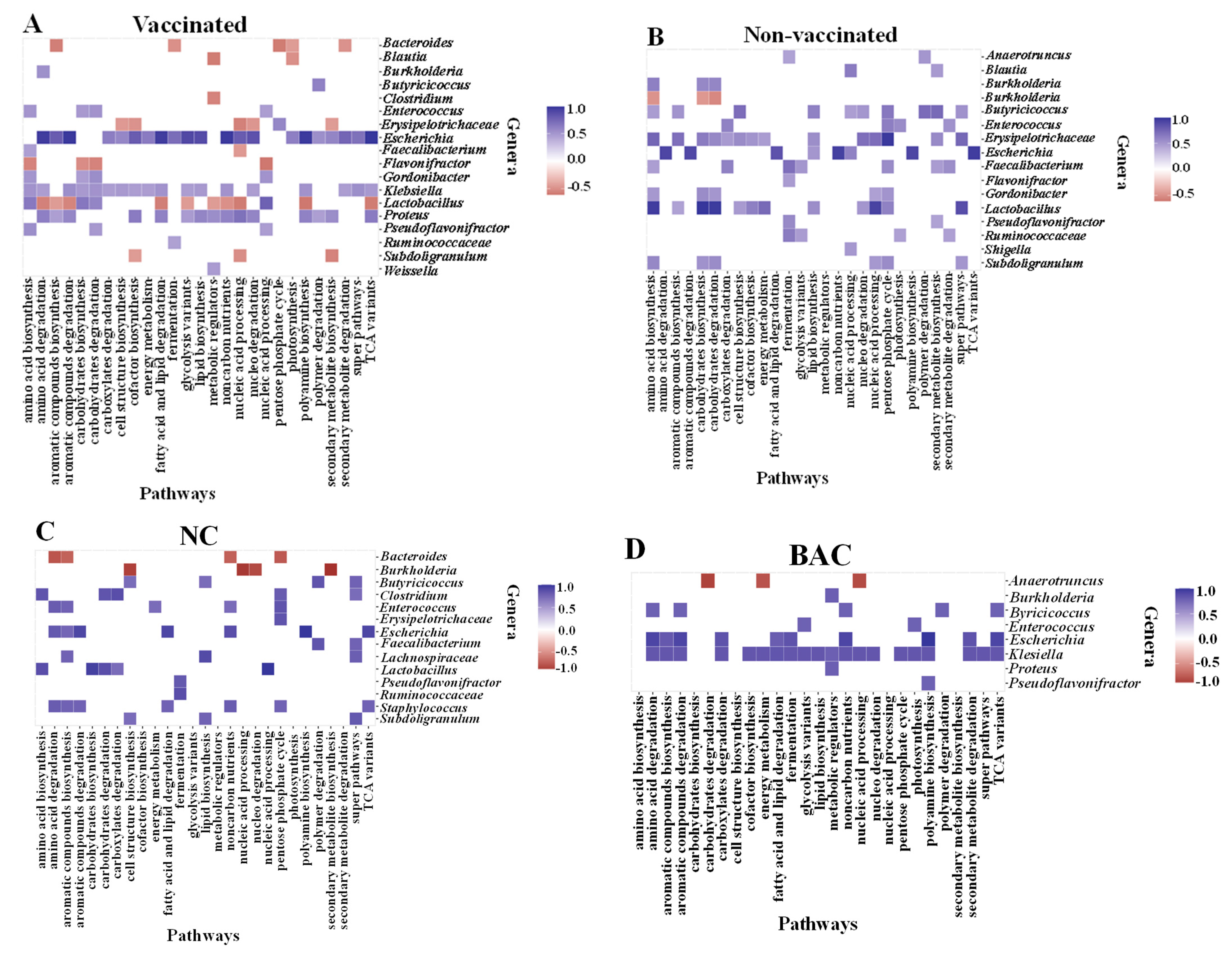
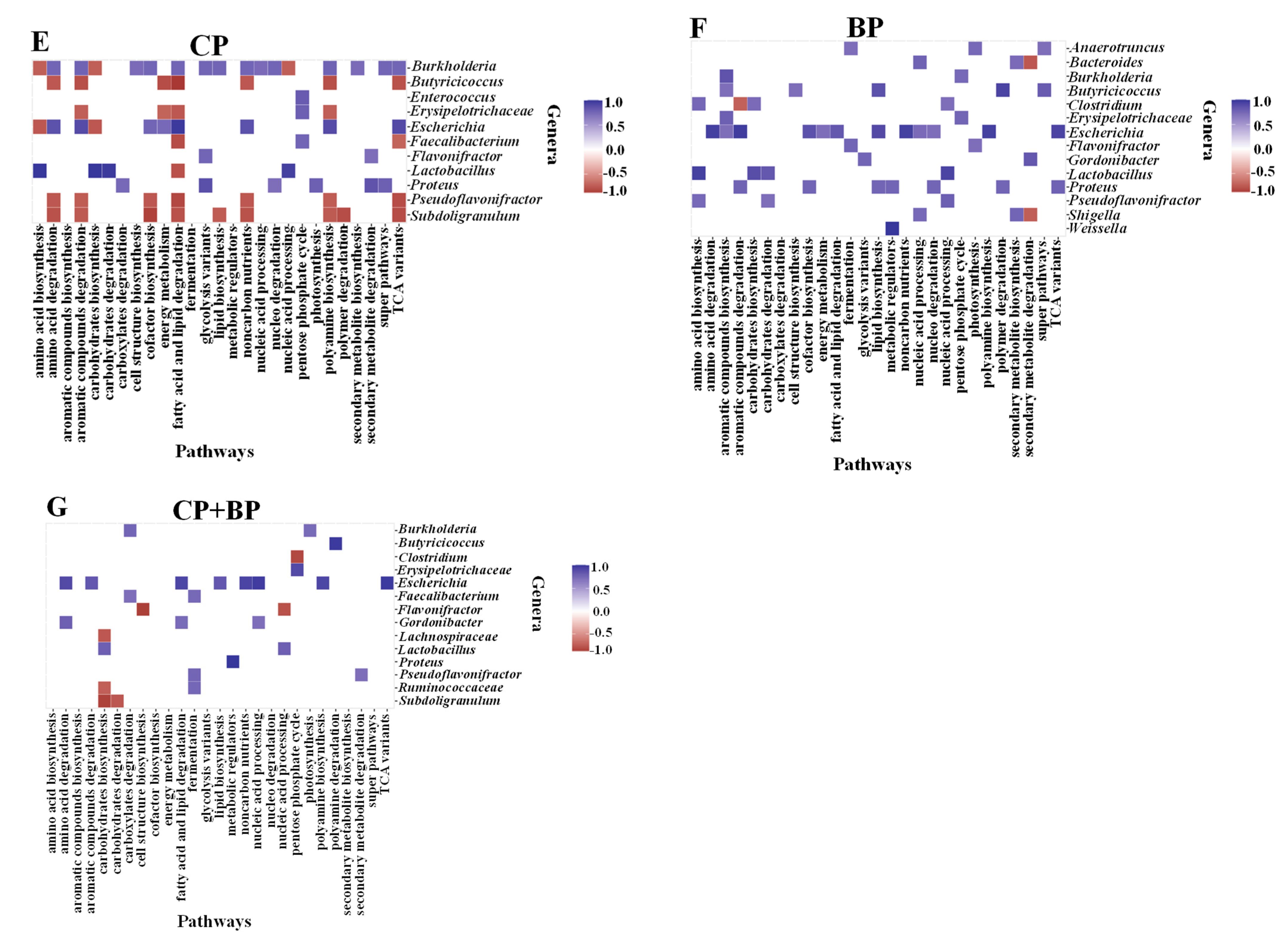
3.8. Correlations between Bacterial Genera and Metabolic Pathways
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abudabos, A.M.; Samara, E.M.; Hussein, E.O.; Al-Ghadi, M.A.Q.; Al-Atiyat, R.M. Impacts of stocking density on the performance and welfare of broiler chickens. Ital. J. Anim. Sci. 2013, 12, e11. [Google Scholar] [CrossRef]
- Chapman, H.D.; Barta, J.R.; Blake, D.; Gruber, A.; Jenkins, M.; Smith, N.C.; Suo, X.; Tomley, F.M. A selective review of advances in coccidiosis research. Adv. Parasitol. 2013, 83, 93–171. [Google Scholar]
- Estévez, M. Oxidative damage to poultry: From farm to fork. Poult. Sci. 2015, 94, 1368–1378. [Google Scholar] [CrossRef]
- Shojadoost, B.; Vince, A.R.; Prescott, J.F. The successful experimental induction of necrotic enteritis in chickens by Clostridium perfringens: A critical review. Vet. Res. 2012, 43, 74. [Google Scholar] [CrossRef]
- Soucy, S.; Huang, J.; Gogarten, J. Horizontal gene transfer: Building the web of life. Nat. Rev. Genet. 2015, 16, 472–482. [Google Scholar] [CrossRef]
- Bailey, M.; Taylor, R.; Brar, J.; Corkran, S.; Velasquez, C.; Novoa-Rama, E.; Oliver, H.F.; Singh, M. Prevalence and antimicrobial resistance of Salmonella from antibiotic-free broilers during organic and conventional processing. J. Food Prot. 2020, 83, 491–496. [Google Scholar] [CrossRef]
- Millman, J.M.; Waits, K.; Grande, H.; Marks, A.R.; Marks, J.C.; Price, L.B.; Hungate, B.A. Prevalence of antibiotic-resistant E. coli in retail chicken: Comparing conventional, organic, kosher, and raised without antibiotics. F1000Research. 2013, 2, 155. [Google Scholar] [CrossRef]
- Haque, M.; Sarker, S.; Islam, M.; Karim, M.; Kayesh, M.E.H.; Shiddiky, M.J.; Anwer, M.S. Sustainable antibiotic-free broiler meat production: Current trends, challenges, and possibilities in a developing country perspective. Biology 2020, 9, 411. [Google Scholar] [CrossRef]
- Fancher, C.A.; Zhang, L.; Kiess, A.S.; Adhikari, P.A.; Dinh, T.T.; Sukumaran, A.T. Avian pathogenic Escherichia coli and Clostridium perfringens: Challenges in no antibiotics ever broiler production and potential solutions. Microorganisms 2020, 8, 1533. [Google Scholar] [CrossRef]
- Dalloul, R.A.; Lillehoj, H.S. Poultry coccidiosis: Recent advancements in control measures and vaccine development. Expert Rev. Vaccines. 2006, 5, 143–163. [Google Scholar] [CrossRef]
- Chapman, H.D.; Jeffers, T.K. Vaccination of chickens against coccidiosis ameliorates drug resistance in commercial poultry production. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 214–217. [Google Scholar] [CrossRef]
- Williams, R.B. Anticoccidial vaccines for broiler chickens: Pathways to success. Avian Pathol. 2002, 31, 317–353. [Google Scholar] [CrossRef]
- Soutter, F.; Werling, D.; Tomley, F.M.; Blake, D.P. Poultry coccidiosis: Design and interpretation of vaccine studies. Front. Vet. Sci. 2020, 7, 101. [Google Scholar] [CrossRef]
- Das, Q.; Islam, M.; Lepp, D.; Tang, J.; Yin, X.; Mats, L.; Liu, H.; Ross, K.; Kennes, Y.M.; Yacini, H.; et al. Gut microbiota, blood metabolites, and spleen immunity in broiler chickens fed berry pomaces and phenolic-enriched extractives. Front. Vet. Sci. 2020, 7, 150. [Google Scholar] [CrossRef] [PubMed]
- Das, Q.; Shaym, J.; Gauthier, M.; Yin, X.; Hasted, T.L.; Ross, K.; Julien, C.; Yacini, H.; Kennes, Y.M.; Warriner, K.; et al. Effects of vaccination against coccidiosis on gut microbiota and immunity in broiler fed bacitracin and berry pomace. Front. Immunol. 2021, 12, 621803. [Google Scholar] [CrossRef]
- Xu, Q.; Si, W.; Mba, O.I.; Sienkiewicz, O.; Ngadi, M.; Ross, K.; Kithama, M.; Kiarie, E.G.; Kennes, Y.M.; Diarra, M.S.; et al. Research Note: Effects of supplementing cranberry and blueberry pomaces on meat quality and antioxidative capacity in broilers. Poult. Sci. 2021, 100, 100900. [Google Scholar] [CrossRef]
- Truong, D.T.; Franzosa, E.A.; Tickle, T.L.; Scholz, M.; Weingart, G.; Pasolli, E.; Tett, A.; Huttenhower, C.; Segata, N. MetaPhlAn2 for enhanced metagenomic taxonomic profiling. Nat. Methods 2015, 12, 902–903. [Google Scholar] [CrossRef]
- Inouye, M.; Dashnow, H.; Raven, L.-A.; Schultz, M.B.; Pope, B.J.; Tomita, T.; Zobel, J.; Holt, K.E. SRST2: Rapid genomic surveillance for public health and hospital microbiology labs. Genome Med. 2014, 6, 90. [Google Scholar] [CrossRef]
- Doster, E.; Lakin, S.M.; Dean, C.J.; Wolfe, C.; Young, J.G.; Boucher, C.; Belk, K.E.; Noyes, N.R.; Morley, P.S. MEGARes 2.0: A database for classification of antimicrobial drug, biocide and metal resistance determinants in metagenomic sequence data. Nucleic Acids Res. 2019, 48, D561–D569. [Google Scholar] [CrossRef]
- Franzosa, E.A.; McIver, L.J.; Rahnavard, G.; Thompson, L.R.; Schirmer, M.; Weingart, G.; Lipson, K.S.; Knight, R.; Caporaso, J.G.; Segata, N.; et al. Species-level functional profiling of metagenomes and metatranscriptomes. Nat. Methods 2018, 15, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Breitwieser, F.P.; Lu, J.; Salzberg, S.L. A review of methods and databases for metagenomic classification and assembly. Brief. Bioinform. 2019, 20, 1125–1136. [Google Scholar] [CrossRef]
- Ross, K.A.; Ehret, D.; Godfrey, D.; Fukumoto, L.; Diarra, M. Characterization of pilot scale processed Canadian organic cranberry (Vaccinium macrocarpon) and blueberry (V. angustifolium) juice pressing residues and phenolic-enriched extractives. Int. J. Fruit Sci. 2017, 17, 202–232. [Google Scholar] [CrossRef]
- Cobb-Vantress Inc. Cobb 500 Broiler Performance and Nutrition Supplement; Cobb-Vantress Inc.: Siloam Springs, AR, USA, 2016. [Google Scholar]
- Shay, J.A.; Haniford, L.S.E.; Cooper, A.; Carrillo, C.D.; Blais, B.W.; Lau, C.H.-F. Exploiting a targeted resistome sequencing approach in assessing antimicrobial resistance in retail foods. Environ. Microbiome 2023, 18, 25. [Google Scholar] [CrossRef]
- Meyer, M.; Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5448. [Google Scholar] [CrossRef]
- Yang, C.; Kennes, Y.M.; Lepp, D.; Yin, X.; Wang, Q.; Yu, H.; Yang, C.; Gong, J.; Diarra, M.S. Effects of encapsulated cinnamaldehyde and citral on the performance and cecal microbiota of broilers vaccinated or not vaccinated against coccidiosis. Poult. Sci. 2020, 99, 936–948. [Google Scholar] [CrossRef]
- Segata, N.; Waldron, L.; Ballarini, A.; Narasimhan, V.; Jousson, O.; Huttenhower, C. Metagenomic microbial community profiling using unique clade-specific marker genes. Nat. Methods 2012, 9, 811–814. [Google Scholar] [CrossRef]
- Wang, L.; Fang, M.; Hu, Y.; Yang, Y.; Yang, M.; Chen, Y. Characterization of the most abundant Lactobacillus species in chicken gastrointestinal tract and potential use as probiotics for genetic engineering. Acta Biochim. Biophys. Sin. 2014, 46, 612–619. [Google Scholar] [CrossRef]
- Mason, A.S.; Lund, A.R.; Hocking, P.M.; Fulton, J.E.; Burt, D.W. Identification and characterisation of endogenous Avian Leukosis Virus subgroup E (ALVE) insertions in chicken whole genome sequencing data. Mobile DNA 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Laurent, F.; Mancassola, R.; Lacroix, S.; Menezes, R.; Naciri, M. Analysis of chicken mucosal immune response to Eimeria tenella and Eimeria maxima infection by quantitative reverse transcription-PCR. Infect. Immun. 2001, 69, 2527–2534. [Google Scholar] [CrossRef]
- Frank, D.N.; St Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2017, 104, 13780–13785. [Google Scholar] [CrossRef]
- Kathayat, D.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Avian Pathogenic Escherichia coli (APEC): An overview of virulence and pathogenesis factors, zoonotic potential, and control strategies. Pathogens 2021, 10, 467. [Google Scholar] [CrossRef]
- Yang, C.; Rehman, M.A.; Yin, X.; Carrillo, C.D.; Wang, Q.; Yang, C.; Gong, J.; Diarra, M.S. Antimicrobial resistance phenotypes and genotypes of Escherichia coli isolates from broiler chickens fed encapsulated cinnamaldehyde and citral. J. Food Prot. 2021, 84, 1385–1399. [Google Scholar] [CrossRef]
- Das, Q.; Hasted, T.L.; Lepp, D.; Yin, X.; Tang, J.; Chalmers, G.; Ross, K.; Boerlin, P.; Diarra, M.S. Transcriptional profiling of extraintestinal Escherichia coli exposed to cranberry pomace extracts alone or in combination with ceftriaxone. Front. Sustain. Food Syst. 2023, 6, 957099. [Google Scholar] [CrossRef]
- Ranfaing, J.; Dunyach-Remy, C.; Louis, L.; Lavigne, J.P.; Sotto, A. Propolis potentiates the effect of cranberry (Vaccinium macrocarpon) against the virulence of uropathogenic Escherichia coli. Sci. Rep. 2018, 8, 10706. [Google Scholar] [CrossRef]
- Huang, P.; Zhang, Y.; Xiao, K.; Jiang, F.; Wang, H.; Tang, D.; Liu, D.; Liu, B.; Liu, Y.; He, X.; et al. The chicken gut metagenome and the modulatory effects of plant-derived benzylisoquinoline alkaloids. Microbiome 2018, 6, 211. [Google Scholar] [CrossRef]
- Orso, C.; Stefanello, T.B.; Franceschi, C.H.; Mann, M.B.; Varela, A.P.M.; Castro, I.M.S.; Frazzon, J.; Frazzon, A.P.G.; Andretta, I.; Ribeiro, A.M.L. Changes in the ceca microbiota of broilers vaccinated for coccidiosis or supplemented with salinomycin. Poult. Sci. 2021, 100, 100969. [Google Scholar] [CrossRef]
- Forstner, J.F.; Oliver, M.G.; Sylvester, F.A. Production, structure, and biologic relevance of gastrointestinal mucins. In Infections of the Gastrointestinal Tract; Guerrant, L.R., Ed.; Raven Press: New York, NY, USA, 1995; pp. 71–88. [Google Scholar]
- Biasato, I.; Ferrocino, I.; Grego, E.; Dabbou, S.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Gut microbiota and mucin composition in female broiler chickens fed diets including yellow mealworm (Tenebrio molitor, L.). Animals 2019, 9, 213. [Google Scholar] [CrossRef]
- Robertson, A.M.; Wright, D.P. Bacterial glycosulphatases and sulphomucin degradation. Can. J. Gastroenterol. 1997, 11, 361–366. [Google Scholar] [CrossRef]
- Maisuria, V.B.; Lopez-de Los Santos, Y.; Tufenkji, N.; Déziel, E. Cranberry-derived proanthocyanidins impair virulence and inhibit quorum sensing of Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 30169. [Google Scholar] [CrossRef]
- Samarasinghe, S.; Reid, R.; Al-Bayati, M. The anti-virulence effect of cranberry active compound proanthocyanins (PACs) on expression of genes in the third-generation cephalosporin-resistant Escherichia coli CTX-M-15 associated with urinary tract infection. Antimicrob. Resist. Infect. Control. 2019, 8, 181. [Google Scholar] [CrossRef]
- Das, Q.; Lepp, D.; Yin, X.; Ross, K.; McCallum, J.L.; Warriner, K.; Marcone, M.F.; Diarra, M.S. Transcriptional profiling of Salmonella enterica serovar Enteritidis exposed to ethanolic extract of organic cranberry pomace. PLoS ONE 2019, 14, e0219163. [Google Scholar] [CrossRef]
- Beaudry, M.S.; Thomas, J.C.; Baptista, R.P.; Sullivan, A.H.; Norfolk, W.; Devault, A.; Enk, J.; Kieran, T.J.; Rhodes, O.E., Jr.; Perry-Dow, K.A.; et al. Escaping the fate of Sisyphus: Assessing resistome hybridization baits for antimicrobial resistance gene capture. Env. Microbiol. 2021, 23, 7523–7537. [Google Scholar] [CrossRef]
- Diarra, M.S.; Block, G.; Rempel, H.; Oomah, B.D.; Harrison, J.; McCallum, J.; Boulanger, S.; Brouillette, É.; Gattuso, M.; Malouin, F. In vitro and in vivo antibacterial activities of cranberry press cake extracts alone or in combination with β-lactams against Staphylococcus aureus. BMC Complement. Altern. Med. 2013, 13, 90. [Google Scholar] [CrossRef]
- Lacombe, A.; Wu, V.C. The potential of berries to serve as selective inhibitors of pathogens and promoters of beneficial microorganisms. Food Qual. Saf. 2017, 1, 3–12. [Google Scholar] [CrossRef]
- Omardien, S.; Brul, S.; Zaat, S.A. Antimicrobial activity of cationic antimicrobial peptides against gram-positives: Current progress made in understanding the mode of action and the response of bacteria. Front. Cell Dev. Biol. 2016, 4, 111. [Google Scholar] [CrossRef]
- Maisuria, V.B.; Okshevsky, M.; Déziel, E.; Tufenkji, N. Proanthocyanidin interferes with intrinsic antibiotic resistance mechanisms of Gram-negative bacteria. Adv. Sci. 2019, 6, 1802333. [Google Scholar] [CrossRef]
- Amerah, A.M.; Ravindran, V. Effect of coccidia challenge and natural betaine supplementation on performance, nutrient utilization, and intestinal lesion scores of broiler chickens fed suboptimal level of dietary methionine. Poult. Sci. 2015, 94, 673–680. [Google Scholar] [CrossRef]
- Gautier, A.E.; Rochell, S.J. Influence of coccidiosis vaccination on nutrient utilization of corn, soybean meal, and distillers dried grains with soluble in broilers. Poult. Sci. 2020, 99, 3540–3549. [Google Scholar] [CrossRef]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken gut microbiota: Importance and detection technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef]
- Porcheron, G.; Chanteloup, N.K.; Trotereau, A.; Brée, A.; Schouler, C. Effect of fructooligosaccharide metabolism on chicken colonization by an extra-intestinal pathogenic Escherichia coli strain. PLoS ONE 2012, 7, e35475. [Google Scholar] [CrossRef]
- Wu, Z.; Chen, C.; Zhang, Q.; Bao, J.; Fan, Q.; Li, R.; Ishfaq, M.; Li, J. Arachidonic acid metabolism is elevated in Mycoplasma gallisepticum and Escherichia coli co-infection and induces LTC4 in serum as the biomarker for detecting poultry respiratory disease. Virulence 2020, 11, 730–738. [Google Scholar] [CrossRef]
- De Cesare, A.; Sirri, F.; Manfreda, G.; Moniaci, P.; Giardini, A.; Zampiga, M.; Meluzzi, A. Effect of dietary supplementation with Lactobacillus acidophilus D2/CSL (CECT 4529) on caecum microbiome and productive performance in broiler chickens. PLoS ONE 2017, 12, e0176309. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial resistance and virulence: A successful or deleterious association in the bacterial world? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef]
- Mak, P.H.W.; Rehman, M.A.; Kiarie, E.G.; Topp, E.; Diarra, M.S. Production systems and important antimicrobial resistant-pathogenic bacteria in poultry: A review. J. Anim. Sci. Biotechnol. 2022, 13, 148. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.; Gu, J.; Hu, T.; Wang, X.; Sun, Y.; Li, H.; Sun, W.; Qian, X.; Song, Z.; et al. Metagenomic insights into dietary remodeling of gut microbiota and antibiotic resistome in meat rabbits. Sci. Total Environ. 2023, 784, 162006. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomy | Non-Vaccinated 1 | Vaccinated | SEM 2 | Effects 3 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NC | BAC | CP | BP | CP + BP | NC | BAC | CP | BP | CP + BP | Vac | Trt | Vac × Trt | ||
| k_Bacteria | 81.60 | 82.71 | 93.95 | 94.30 | 89.76 | 63.25 | 86.34 | 85.71 | 89.03 | 92.79 | 2.301 | ns | ** | ns |
| k_Eukaryotes | 0 | 0 | 0 | 0 | 0 | 0.1 | 0.02 | 0.07 | 0.01 | 0.01 | 0.011 | * | ns | ns |
| k_Virus | 18.40 | 17.29 | 6.05 | 5.70 | 10.24 | 36.65 | 13.64 | 14.22 | 10.96 | 7.21 | 2.297 | ns | ** | ns |
| p_Firmicutes | 78.03 | 86.82 | 83.53 | 91.06 | 91.19 | 53.94 | 41.1 | 81.21 | 76.49 | 92.51 | 3.616 | *** | ** | ns |
| p_Proteobacteria | 1.82 | 0.65 | 0.39 | 0.15 | 0.56 | 3.71 | 14.15 | 4.68 | 6.30 | 1.57 | 1.106 | ** | ns | ns |
| p_Viruses | 18.40 | 17.29 | 6.05 | 5.70 | 10.24 | 36.65 | 13.64 | 14.22 | 10.96 | 7.21 | 3.156 | * | ** | ns |
| f_Lactobacillaceae | 43.91 | 63.80 | 81.29 | 68.29 | 62.14 | 31.67 | 37.37 | 52.67 | 58.38 | 68.04 | 3.646 | ns | ** | ns |
| f_Oscillospiraceae | 13.08 | 6.01 | 3.94 | 5.07 | 6.52 | 7.67 | 8.42 | 6.48 | 8.69 | 6.48 | 0.621 | ns | * | ns |
| f_Enterobacteriaceae | 1.60 | 0.31 | 0.35 | 0.09 | 0.58 | 3.63 | 17.82 | 4.85 | 3.47 | 1.86 | 1.105 | *** | ns | * |
| f_Eimeriidae | 0 | 0 | 0 | 0 | 0 | 0.1 | 0.02 | 0.07 | 0.01 | 0.01 | 0.011 | * | ns | ns |
| f_Retroviridae | 18.40 | 17.29 | 6.05 | 5.70 | 9.01 | 36.65 | 13.56 | 14.22 | 10.81 | 7.21 | 2.300 | ns | ** | ns |
| g_Lactobacillus | 43.91 | 63.8 | 81.29 | 68.29 | 62.14 | 31.67 | 37.37 | 52.67 | 58.38 | 68.04 | 3.646 | ** | ** | ns |
| g_Butyricicoccus | 6.59 | 1.13 | 0.47 | 0.46 | 0.91 | 0.45 | 7.42 | 0.55 | 4.4 | 1.43 | 0.692 | ns | ns | * |
| g_Escherichia | 1.58 | 0.3 | 0.35 | 0.09 | 0.58 | 3.6 | 17.14 | 4.82 | 3.41 | 1.84 | 1.059 | *** | ns | * |
| g_Eimeria | 0 | 0 | 0 | 0 | 0 | 0.1 | 0.02 | 0.07 | 0.01 | 0.01 | 0.011 | * | ns | ns |
| g_Retroviridae | 18.40 | 17.29 | 6.05 | 5.70 | 9.01 | 36.65 | 13.56 | 14.22 | 10.81 | 7.21 | 2.300 | ns | ** | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Das, Q.; Rehman, M.A.; Yin, X.; Shay, J.; Gauthier, M.; Lau, C.H.-F.; Ross, K.; Diarra, M.S. Microbiome of Ceca from Broiler Chicken Vaccinated or Not against Coccidiosis and Fed Berry Pomaces. Microorganisms 2023, 11, 1184. https://doi.org/10.3390/microorganisms11051184
Yang C, Das Q, Rehman MA, Yin X, Shay J, Gauthier M, Lau CH-F, Ross K, Diarra MS. Microbiome of Ceca from Broiler Chicken Vaccinated or Not against Coccidiosis and Fed Berry Pomaces. Microorganisms. 2023; 11(5):1184. https://doi.org/10.3390/microorganisms11051184
Chicago/Turabian StyleYang, Chongwu, Quail Das, Muhammad A. Rehman, Xianhua Yin, Julie Shay, Martin Gauthier, Calvin Ho-Fung Lau, Kelly Ross, and Moussa S. Diarra. 2023. "Microbiome of Ceca from Broiler Chicken Vaccinated or Not against Coccidiosis and Fed Berry Pomaces" Microorganisms 11, no. 5: 1184. https://doi.org/10.3390/microorganisms11051184