
Bioaccessibility of Maillard Reaction Products from Biscuits Formulated from Buckwheat Flours Fermented by Selected Lactic Acid Bacteria

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Fermentation of Raw and Roasted Buckwheat Flours, Preparation of Buckwheat Biscuits from Fermented Flours, and In Vitro Digestion
2.3. Determination of Maillard Reaction Products and Their Bioaccessibility
2.4. Statistical Analysis
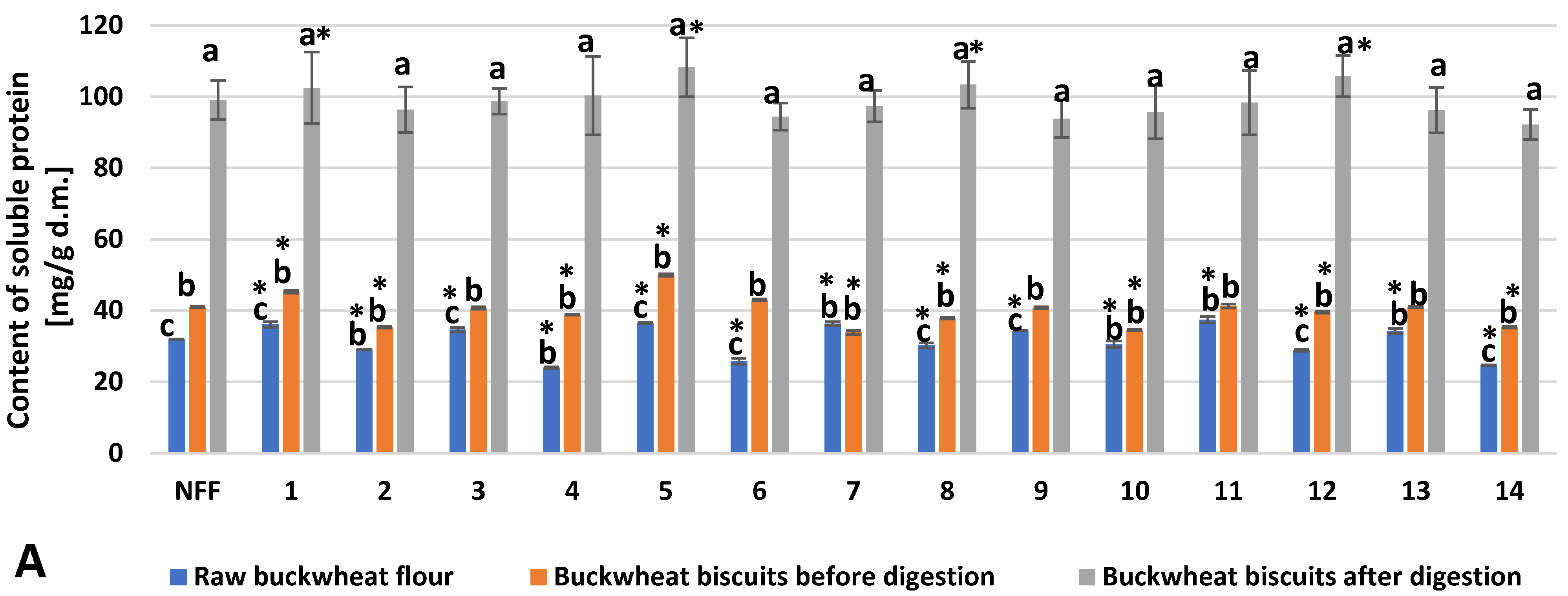
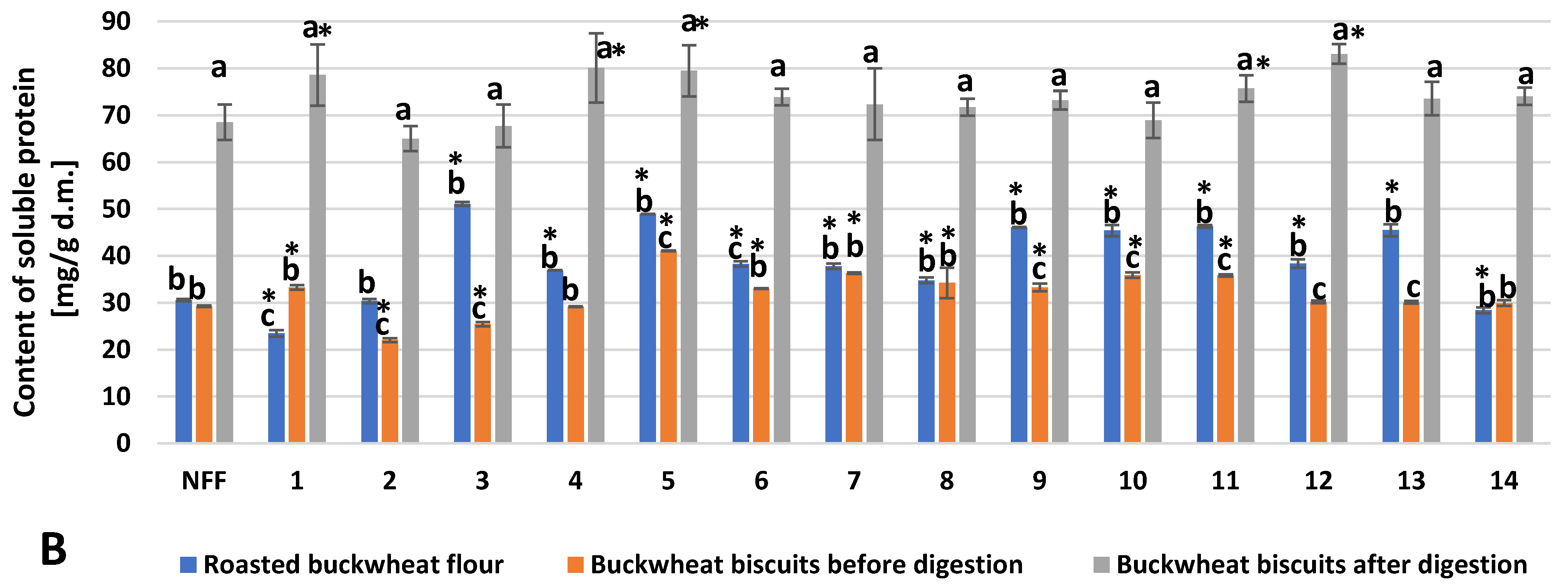
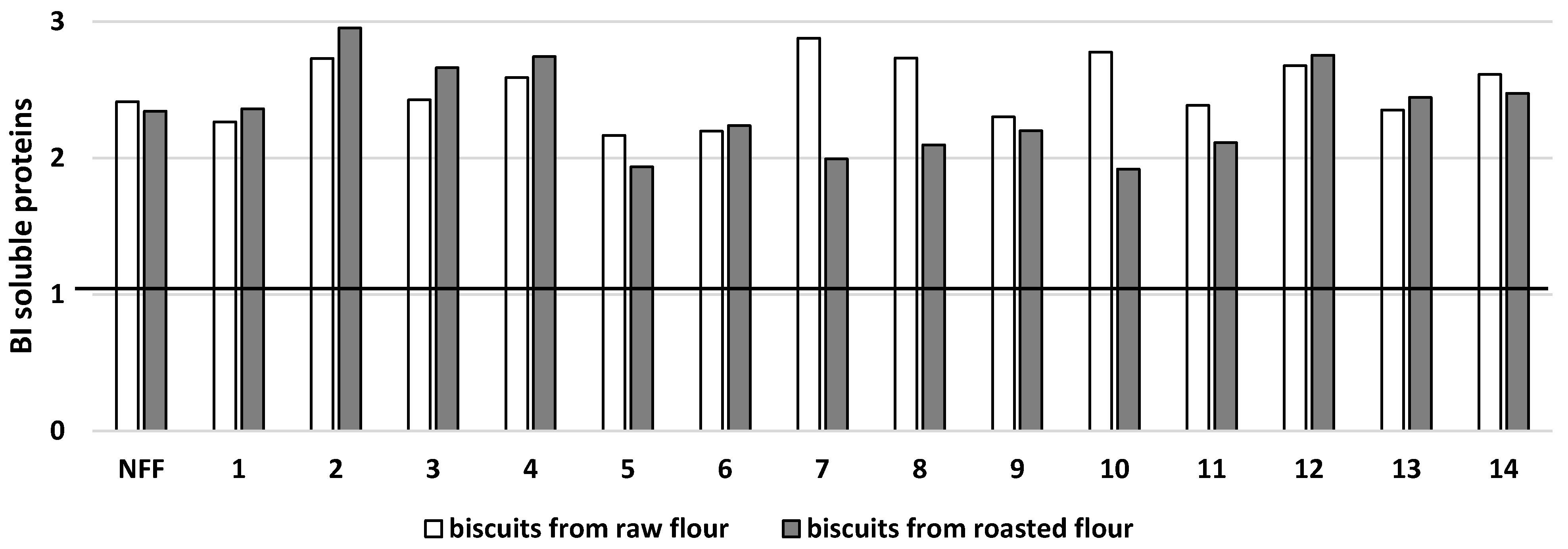
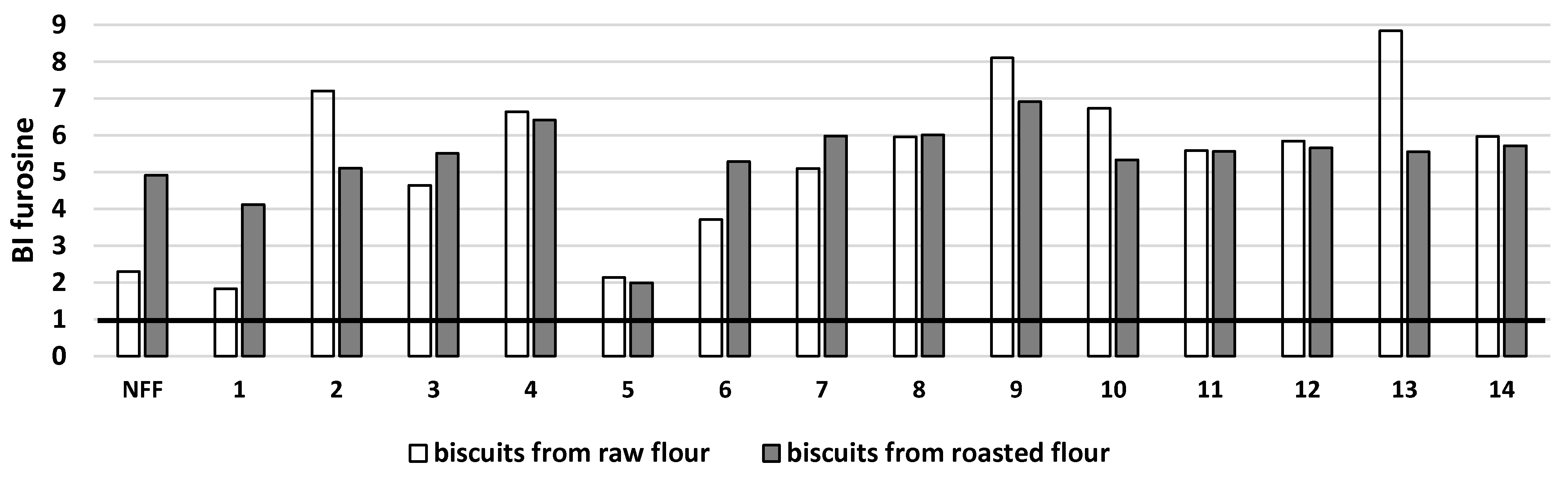
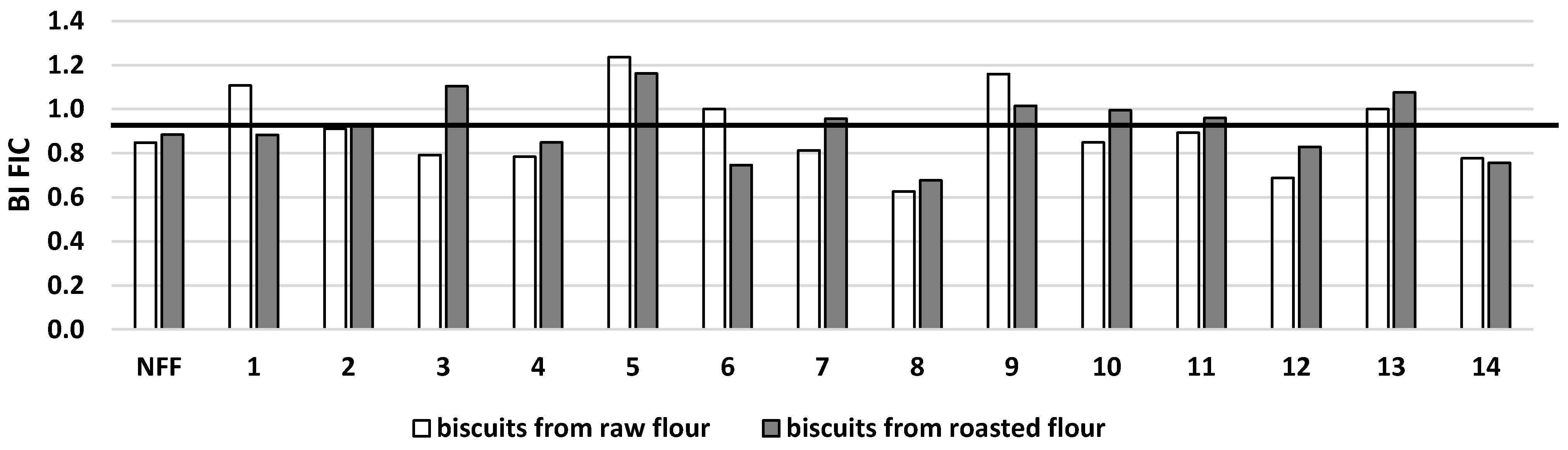
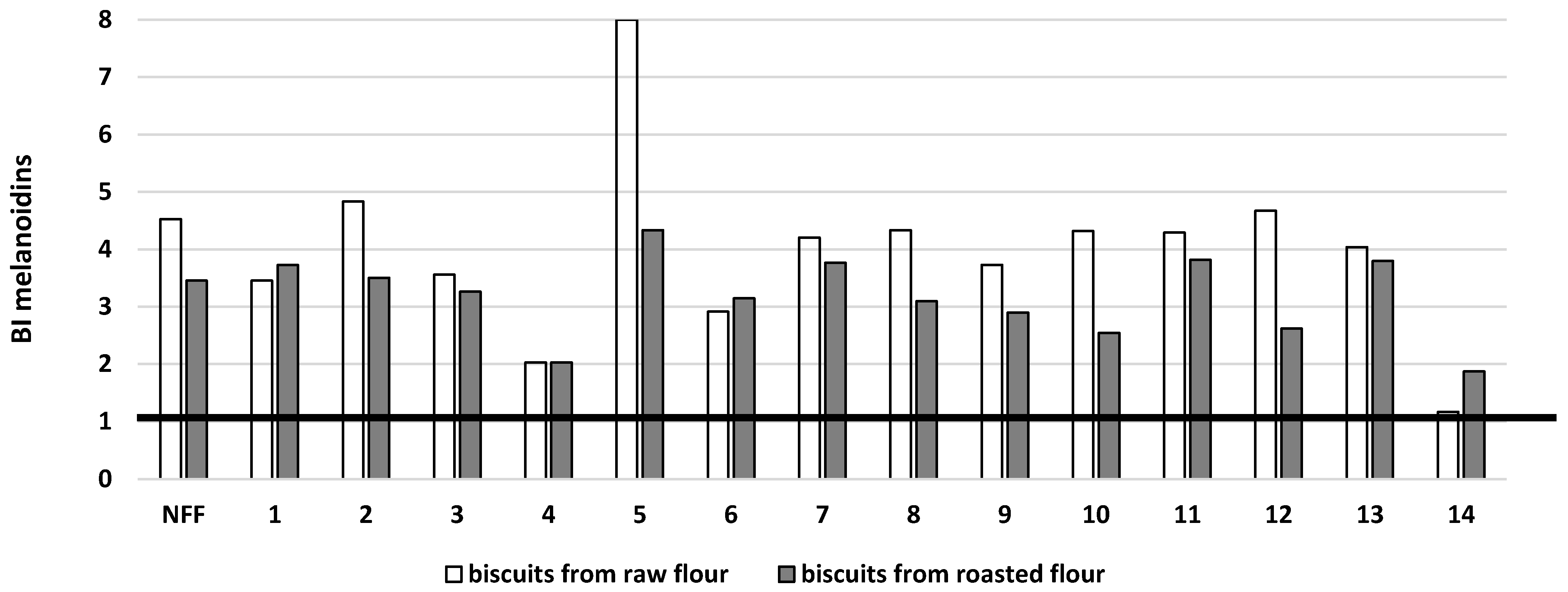
3. Results
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Muller, M.R.; Wolfrum, G.; Stolz, P.; Ehrmann, M.A.; Vogel, R.F. Monitoring the growth of Lactobacillus species during a rye flour fermentation. Food Microbiol. 2001, 18, 217–227. [Google Scholar] [CrossRef]
- Charalampopoulos, D.; Vazquez, J.A.; Pandiella, S.S. Modelling and validation of Lactobacillus plantarum fermentations in cereal-based media with different sugar concentrations and buffering capacities. Biochem. Eng. J. 2009, 44, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Simwaka, J.E.; Chamba, M.V.M.; Huiming, Z.; Masamba, K.G.; Luo, Y. Effect of fermentation on physicochemical and antinutritional factors of complementary foods from millet, sorghum, pumpkin and amaranth seed flours. Int. Food Res. J. 2017, 24, 1869–1879. [Google Scholar]
- Džafić, A.; Oručević Žuljević, S. The Importance of Buckwheat as a Pseudocereal: Content and Stability of Its Main Bioactive Components. In Pseudocereals; Waisundara, V.Y., Ed.; IntechOpen: London, UK, 2022; Chapter 6; pp. 1–27. [Google Scholar]
- Zielińska, D.; Szawara-Nowak, D.; Michalska, A. Antioxidant capacity of thermally-treated buckwheat. Pol. J. Food Nutr. Sci. 2007, 57, 465–470. [Google Scholar]
- Różańska, M.B.; Siger, A.; Szwengiel, A.; Dziedzic, K.; Mildner-Szkudlarz, S. Maillard Reaction Products in Gluten-Free Bread Made from Raw and Roasted Buckwheat Flour. Molecules 2021, 26, 1361. [Google Scholar] [CrossRef] [PubMed]
- Zieliński, H.; Szawara-Nowak, D.; Bączek, N.; Wronkowska, M. Effect of liquid-state fermentation on the antioxidant and functional properties of raw and roasted buckwheat flours. Food Chem. 2019, 271, 291–297. [Google Scholar] [CrossRef]
- Baráth, Á.; Halasz, A.; Németh, E.; Zalán, Z. Selection of LAB strains for fermented red beet juice production. Eur. Food Res. Technol. 2004, 218, 184–187. [Google Scholar] [CrossRef]
- Carbonell-Capella, J.M.; Buniowska, M.; Barba, F.J.; Esteve, M.J.; Frígola, A. Analytical methods for determining bioavailability and bioaccessibility of bioactive compounds from fruits and vegetables: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 155–171. [Google Scholar] [CrossRef]
- Nooshkam, M.; Varidi, M.; Bashash, M. The Maillard reaction products as food-born antioxidant and antibrowning agents in model and real food systems. Food Chem. 2019, 275, 644–660. [Google Scholar] [CrossRef]
- Zieliński, H.; Honke, J.; Bączek, N.; Majkowska, A.; Wronkowska, M. Bioaccessibility of D-chiro-inositol from water biscuits formulated from buckwheat flours fermented by lactic acid bacteria and fungi. LWT—Food Sci. Technol. 2019, 106, 37–43. [Google Scholar] [CrossRef]
- Zieliński, H.; Wiczkowski, W.; Honke, J.; Piskuła, M.K. In Vitro Expanded Bioaccessibility of Quercetin-3-Rutinoside and Quercetin Aglycone from Buckwheat Biscuits Formulated from Flours Fermented by Lactic Acid Bacteria. Antioxidants 2021, 10, 571. [Google Scholar] [CrossRef]
- Wronkowska, M.; Jeliński, T.; Majkowska, A.; Zieliński, H. Physical properties of buckwheat water biscuits formulated from fermented flours by selected lactic acid bacteria. Pol. J. Food Nutr. Sci. 2018, 68, 25–31. [Google Scholar] [CrossRef]
- Michalska, A.; Amigo-Benavent, M.; Zielinski, H.; del Castillo, M.D. Effect of bread making on formation of Maillard reaction products contributing to the overall antioxidant activity of rye bread. J. Cereal Sci. 2008, 48, 123–132. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Rufián-Henares, J.A.; Morales, F.J. Fast method to determine furosine in breakfast cereals by capillary zone electrophoresis. Eur. Food Res. Technol. 2005, 221, 707–711. [Google Scholar] [CrossRef] [Green Version]
- Zieliński, H.; del Castillo, M.D.; Przygodzka, M.; Ciesarova, Z.; Kukurova, K.; Zielińska, D. Changes in chemical composition and antioxidative properties of rye ginger cakes during their shelf-life. Food Chem. 2012, 135, 2965–2973. [Google Scholar] [CrossRef] [PubMed]
- Damjanovic Desic, S.; Birlouez-Aragon, I. The FAST index—A highly sensitive indicator of the heat impact on infant formula model. Food Chem. 2011, 124, 1043–1049. [Google Scholar] [CrossRef]
- Guerra-Hernández, E.; Corzo, N.; García-Villanova, B. Maillard Reaction Evaluation by Furosine Determination during Infant Cereal Processing. J. Cereal Sci. 1999, 29, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Mesías, M.; Sáez-Escudero, L.; Morales, F.J.; Delgado-Andrade, C. Occurrence of Furosine and Hydroxymethylfurfural in Breakfast Cereals. Evolution of the Spanish Market from 2006 to 2018. Foods 2019, 8, 158. [Google Scholar] [CrossRef] [Green Version]
- Yıltırak, S.; Kocadağlı, T.; Çelik, E.E.; Kanmaz, E.O.; Gökmen, V. Effects of sprouting and fermentation on the formation of Maillard reaction products in different cereals heated as wholemeal. Food Chem. 2022, 389, 133075. [Google Scholar] [CrossRef]
- Birlouez-Aragon, I.; Nicolas, M.; Metais, A.; Marchond, N.; Grenier, J.; Calvo, D. A Rapid Fluorimetric Method to Estimate the Heat Treatment of Liquid Milk. Int. Dairy J. 1998, 8, 771–777. [Google Scholar] [CrossRef]
- Leclère, J.; Birlouez-Aragon, I. The Fluorescence of Advanced Maillard Products Is a Good Indicator of Lysine Damage during the Maillard Reaction. J. Agric. Food Chem. 2001, 49, 4682–4687. [Google Scholar] [CrossRef] [PubMed]
- Langner, E.; Rzeski, W. Biological Properties of melanoidins: A review. Int. J. Food Prop. 2014, 17, 344–353. [Google Scholar] [CrossRef]
- Brudzynski, K.; Miotto, D. The relationship between the content of Maillard reaction-like products and bioactivity of Canadian honeys. Food Chem. 2011, 124, 869–874. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | ||||||||||
| Strain/Sample | Furosine (mg/g d.m.) | Free FIC (FI/mg d.m.) | Tryptophan (FI/mg d.m.) | FAST Index (%) | Browning Index (AU) | |||||
| Before | After | Before | After | Before | After | Before | After | Before | After | |
| Nonfermented raw flour | 3.28 ± 0.04 b | 7.53 ± 0.51 a | 78.9 ± 3.9 a | 66.9 ± 0.8 b | 30.1 ± 0.9 b | 120.1 ± 3.4 a | 262.2 ± 18.4 a | 55.7 ± 2.1 b | 0.21 ± 0.01 b | 0.96 ± 0.02 a |
| Raw flour fermented by: | ||||||||||
| Lactiplantibacillus plantarum IB | 6.58 ± 0.05 *b | 12.05 ± 0.79 *a | 69.0 ± 3.0 *a | 76.4 ± 6.0 * a | 17.7 ± 5.3 *b | 65.5 ± 23.7 *a | 471.9 ± 27.4 *a | 116.7 ± 9.4 *b | 0.63 ± 0.25 *b | 2.18 ± 0.79 *a |
| Lactiplantibacillus plantarum W42 | 0.72 ± 0.03 *b | 5.16 ± 0.51 *a | 67.3 ± 3.0 *a | 61.3 ± 1.9 b | 31.2 ± 1.8 b | 124.5 ± 3.7 a | 215.3 ± 18.6 a | 49.2 ± 1.6 b | 0.22 ± 0.01 b | 1.06 ± 0.36 a |
| Lactobacillus delbrucki subsp. bulgaricus 151 | 0.94 ± 0.02 *b | 4.38 ± 0.40 *a | 96.4 ± 0.5 *a | 76.3 ± 1.2 * b | 33.1 ± 1.3 b | 119.4 ± 4.8 a | 291.7 ± 11.5 a | 63.9 ± 3.4 b | 0.23 ± 0.01 b | 0.84 ± 0.10 a |
| Lacticaseibacillus casei Lcy | 0.79 ± 0.02 *b | 5.25 ± 0.47 *a | 80.9 ± 3.0 a | 63.4 ± 0.1 b | 32.1 ± 0.7 b | 123.4 ± 2.7 a | 251.8 ± 8.3 a | 51.4 ± 1.1 b | 0.47 ± 0.01 *b | 0.96 ± 0.05 a |
| Streptococcus thermophilus MK-10 | 6.64 ± 0.01 *b | 14.20 ± 1.29 *a | 79.8 ± 0.7 b | 98.6 ± 2.3 *a | 14.8 ± 0.6 *b | 74.9 ± 5.3 *a | 538.7 ± 22.2 *a | 131.6 ± 12.5 *b | 0.21 ± 0.02 b | 1.74 ± 0.08 *a |
| Lactobacillus acidophilus La5 | 0.99 ± 0.02 *b | 3.67 ± 0.17 *a | 68.7 ± 2.8 *a | 68.7 ± 2.0 a | 30.5 ± 2.4 b | 115.4 ± 2.8 a | 225.3 ± 8.8 a | 59.5 ± 2.4 b | 0.23 ± 0.01 b | 0.66 ± 0.08 *a |
| Lactobacillus acidophilus V | 1.08 ± 0.02 *b | 5.50 ± 0.66 *a | 94.9 ± 1.3 *a | 77.1 ± 4.2 *b | 33.7 ± 1.3 b | 126.2 ± 2.9 a | 281.9 ± 15.1 a | 61.1 ± 2.5 b | 0.22 ± 0.02 b | 0.93 ± 0.07 a |
| Lactobacillus acidophilus 145 | 0.93 ± 0.02 *b | 5.54 ± 0.50 *a | 105.5 ± 0.5 *a | 66.1 ± 8.8 b | 39.6 ± 1.6 *b | 119.5 ± 16.2 a | 266.2 ± 10.0 a | 55.3 ± 1.2 b | 0.23 ± 0.01 b | 1.00 ± 0.17 a |
| Lacticaseibacillus casei 2K | 0.71 ± 0.01 *b | 5.74 ± 0.12 * a | 59.0 ± 1.6 *b | 68.4 ± 2.1 a | 25.1 ± 0.9 *b | 129.6 ± 6.2 a | 234.7 ± 11.5 a | 52.8 ± 2.1 b | 0.26 ± 0.01 b | 0.96 ± 0.13 a |
| Lactobacillus delbrucki subsp. bulgaricus K | 0.89 ± 0.02 *b | 6.00 ± 0.13 a | 69.6 ± 1.8 *a | 59.1 ± 9.2 * a | 26.8 ± 0.0 b | 104.0 ± 12.7 *a | 259.5 ± 6.4 a | 56.8 ± 2.1 b | 0.23 ± 0.01 b | 0.99 ± 0.05 a |
| Lacticaseibacillus rhamnosus GG | 1.13 ± 0.02 *b | 6.30 ± 0.25 a | 61.8 ± 0.8 *a | 55.2 ± 7.5 * a | 24.1 ± 1.3 *b | 103.5 ± 15.1 *a | 256.5 ± 16.9 a | 53.4 ± 8.3 b | 0.21 ± 0.01 b | 0.92 ± 0.05 a |
| Lacticaseibacillus rhamnosus 8/4 | 0.98 ± 0.02 *b | 5.76 ± 0.26 * a | 107.7 ± 3.9 *a | 74.1 ± 2.1 * b | 35.7 ± 2.7 *b | 119.0 ± 6.0 a | 301.2 ± 23.3 *a | 62.2 ± 1.8 b | 0.24 ± 0.02 b | 1.12 ± 0.17 a |
| Lacticaseibacillus rhamnosus K | 0.74 ± 0.02 *b | 6.50 ± 0.31 a | 67.8 ± 0.6 *a | 67.8 ± 4.8 a | 28.3 ± 0.7 b | 125.4 ± 8.9 a | 239.5 ± 7.9 a | 54.1 ± 3.0 b | 0.21 ± 0.01 b | 0.85 ± 0.06 a |
| Ligilactobacillus salivarius AWH | 0.60 ± 0.02 *b | 3.58 ± 0.20 * a | 88.7 ± 2.3 *a | 68.9 ± 2.3 b | 31.4 ± 1.7 *b | 109.1 ± 10.9 a | 282.8 ± 22.8 a | 63.1 ± 5.1 b | 1.05 ± 0.02 *b | 1.23 ± 0.09 a |
| (B) | ||||||||||
| Strain/sample | Furosine (mg/g d.m.) | Free FIC (FI/mg d.m.) | Tryptophan (FI/mg d.m.) | FAST Index (%) | Browning Index (AU) | |||||
| Before | After | Before | After | Before | After | Before | After | Before | After | |
| Nonfermented roasted flour | 1.06 ± 0.04 b | 5.20 ± 0.83 a | 55.9 ± 0.6 a | 49.4 ± 1.6 b | 26.1 ± 0.7 b | 89.8 ± 3.7 a | 214.0 ± 4.6 a | 55.0 ± 0.5 b | 0.32 ± 0.01 b | 1.09 ± 0.04 a |
| Roasted flour fermented by: | ||||||||||
| Lactiplantibacillus plantarum IB | 1.00 ± 0.03 b | 4.13 ± 0.23 a | 60.9 ± 1.7 *a | 53.8 ± 2.5 b | 29.6 ± 0.5 *b | 86.6 ± 2.4 a | 205.7 ± 7.7 a | 62.1 ± 1.3 b | 0.31 ± 0.02 b | 1.17 ± 0.15 a |
| Lactiplantibacillus plantarum W42 | 0.96 ± 0.01 *b | 4.91 ± 0.28 a | 61.0 ± 1.1 *a | 56.9 ± 1.9 *b | 27.8 ± 1.4 b | 91.9 ± 0.6 a | 219.7 ± 13.3 a | 61.9 ± 2.4 b | 0.30 ± 0.03 b | 1.05 ± 0.03 a |
| Lactobacillus delbrucki subsp. bulgaricus 151 | 0.91 ± 0.02 *b | 5.03 ± 0.78 a | 51.2 ± 2.8 a | 56.5 ± 2.0 *a | 21.2 ± 0.3 *b | 86.9 ± 1.4 a | 241.5 ± 12.4 *a | 65.1 ± 3.1 *b | 0.31 ± 0.02 b | 1.00 ± 0.10 a |
| Lacticaseibacillus casei Lcy | 0.95 ± 0.04 *b | 6.07 ± 0.20 a | 68.2 ± 1.8 *a | 57.9 ± 0.9 *b | 30.6 ± 0.5 *b | 89.3 ± 1.3 a | 222.7 ± 2.9 a | 64.9 ± 1.7 *b | 0.53 ± 0.03 *b | 1.07 ± 0.06 a |
| Streptococcus thermophilus MK-10 | 6.16 ± 0.04 *b | 12.26 ± 0.83 *a | 59.4 ± 3.0 b | 69.0 ± 1.2 *a | 17.3 ± 1.9 *b | 79.0 ± 6.3 a | 349.9 ± 26.2 *a | 88.1 ± 9.0 *b | 0.29 ± 0.03 b | 1.24 ± 0.11 a |
| Lactobacillus acidophilus La5 | 1.07 ± 0.02 b | 5.66 ± 0.13 a | 72.5 ± 1.8 *a | 54.1 ± 3.2 b | 31.1 ± 1.5 *b | 85.3 ± 4.7 a | 234.2 ± 15.0 a | 63.4 ± 0.9 *b | 0.32 ± 0.01 b | 1.00 ± 0.03 a |
| Lactobacillus acidophilus V | 0.96 ± 0.03 *b | 5.74 ± 0.05 a | 57.9 ± 0.4 a | 55.4 ± 0.8 b | 23.6 ± 0.8 *b | 90.9 ± 1.0 a | 245.1 ± 6.5 *a | 61.0 ± 1.0 b | 0.29 ± 0.03 b | 1.08 ± 0.04 a |
| Lactobacillus acidophilus 145 | 0.94 ± 0.01 *b | 5.66 ± 0.43 a | 76.0 ± 1.6 *a | 51.4 ± 2.1 b | 29.1 ± 1.1 *b | 85.5 ± 4.1 a | 261.7 ± 10.1 *a | 60.2 ± 2.1 b | 0.34 ± 0.02 b | 1.04 ± 0.04 a |
| Lacticaseibacillus casei 2K | 0.77 ± 0.01 *b | 5.35 ± 0.26 a | 52.0 ± 0.6 a | 52.8 ± 3.4 a | 22.0 ± 0.6 *b | 91.5 ± 6.4 a | 236.3 ± 3.9 a | 57.6 ± 0.4 b | 0.34 ± 0.01 b | 0.99 ± 0.11 a |
| Lactobacillus delbrucki subsp. bulgaricus K | 0.97 ± 0.01 *b | 5.16 ± 0.26 a | 55.8 ± 1.7 a | 55.5 ± 1.6 a | 24.7 ± 0.7 b | 89.7 ± 3.5 a | 225.7 ± 3.7 a | 61.8 ± 1.3 b | 0.39 ± 0.01 *b | 0.98 ± 0.05 a |
| Lacticaseibacillus rhamnosus GG | 0.96 ± 0.04 *b | 5.35 ± 0.85 a | 49.3 ± 2.0 *a | 47.3 ± 2.1 a | 22.1 ± 0.3 *b | 82.0 ± 1.1 a | 223.0 ± 11.1 a | 57.7 ± 2.2 b | 0.25 ± 0.05 *b | 0.97 ± 0.09 a |
| Lacticaseibacillus rhamnosus 8/4 | 0.99 ± 0.04 *b | 5.60 ± 0.58 a | 74.1 ± 3.7 *a | 61.4 ± 3.8 *b | 26.2 ± 0.6 b | 97.0 ± 6.1 a | 283.0 ± 9.4 *a | 63.4 ± 0.2 *b | 0.38 ± 0.02 b | 0.98 ± 0.03 a |
| Lacticaseibacillus rhamnosus K | 1.02 ± 0.02 b | 5.65 ± 0.29 a | 52.5 ± 2.9 a | 56.5 ± 4.1 *a | 23.2 ± 0.2 *b | 91.1 ± 6.2 a | 225.8 ± 11.2 a | 62.1 ± 0.8 b | 0.25 ± 0.03 *b | 0.94 ± 0.05 a |
| Ligilactobacillus salivarius AWH | 0.95 ± 0.00 *b | 5.43 ± 0.17 a | 67.8 ± 1.0 *a | 51.3 ± 3.8 b | 28.7 ± 1.0 *b | 84.1 ± 6.4 a | 236.7 ± 7.9 a | 61.0 ± 0.6 b | 0.50 ± 0.01 *b | 0.95 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wronkowska, M.; Szawara-Nowak, D.; Piskuła, M.K.; Zieliński, H. Bioaccessibility of Maillard Reaction Products from Biscuits Formulated from Buckwheat Flours Fermented by Selected Lactic Acid Bacteria. Microorganisms 2023, 11, 883. https://doi.org/10.3390/microorganisms11040883
Wronkowska M, Szawara-Nowak D, Piskuła MK, Zieliński H. Bioaccessibility of Maillard Reaction Products from Biscuits Formulated from Buckwheat Flours Fermented by Selected Lactic Acid Bacteria. Microorganisms. 2023; 11(4):883. https://doi.org/10.3390/microorganisms11040883
Chicago/Turabian StyleWronkowska, Małgorzata, Dorota Szawara-Nowak, Mariusz Konrad Piskuła, and Henryk Zieliński. 2023. "Bioaccessibility of Maillard Reaction Products from Biscuits Formulated from Buckwheat Flours Fermented by Selected Lactic Acid Bacteria" Microorganisms 11, no. 4: 883. https://doi.org/10.3390/microorganisms11040883