
Characterization and Genomic Analysis of the Naphthalene-Degrading Delftia tsuruhatensis ULwDis3 Isolated from Seawater

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain
2.2. Chemicals
2.3. Genome Sequencing, Assembly and Annotation
2.4. Growth Media and Conditions
2.5. Enzyme Activity
2.6. Determination of Hydrocarbon Concentration
3. Results
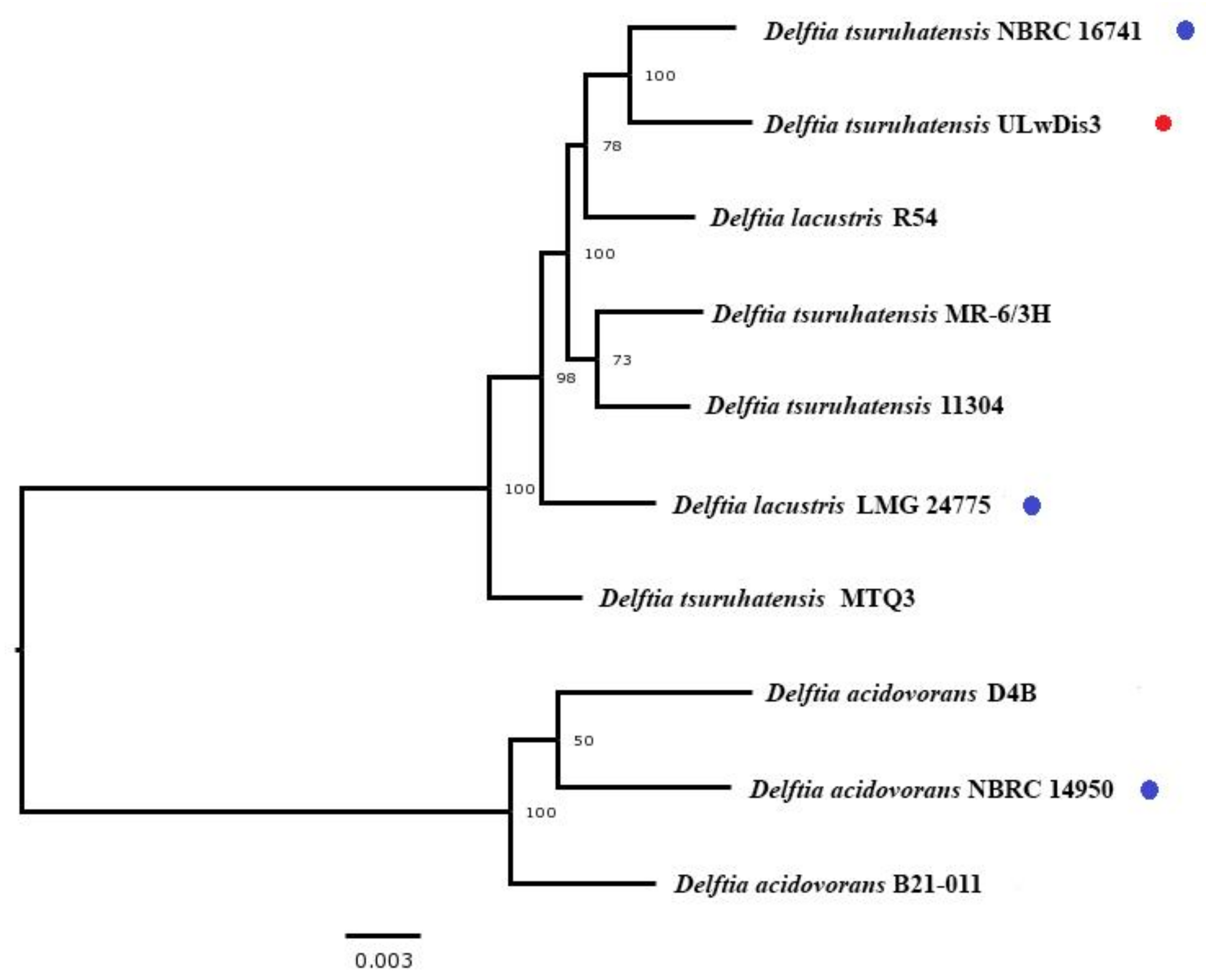
3.1. Nucleotide Sequencing and Annotation
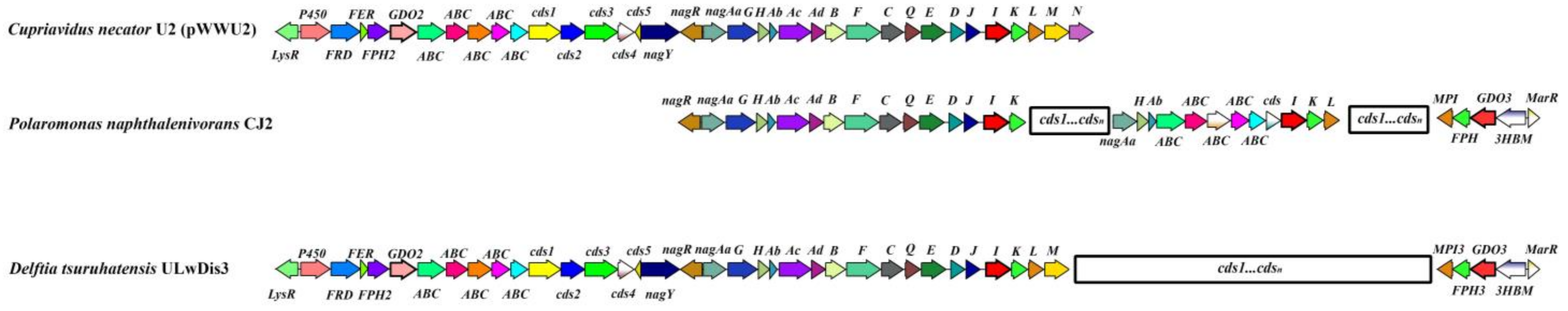
3.2. Genetic Organization of Catabolic Genes of D. tsuruhatensis Strain ULwDis3
3.2.1. Organization of Naphthalene Degradation Genes (nag Genes)
3.2.2. Genes Encoding Gentisate 1.2-Dioxygenases in D. tsuruhatensis Strain ULwDis3
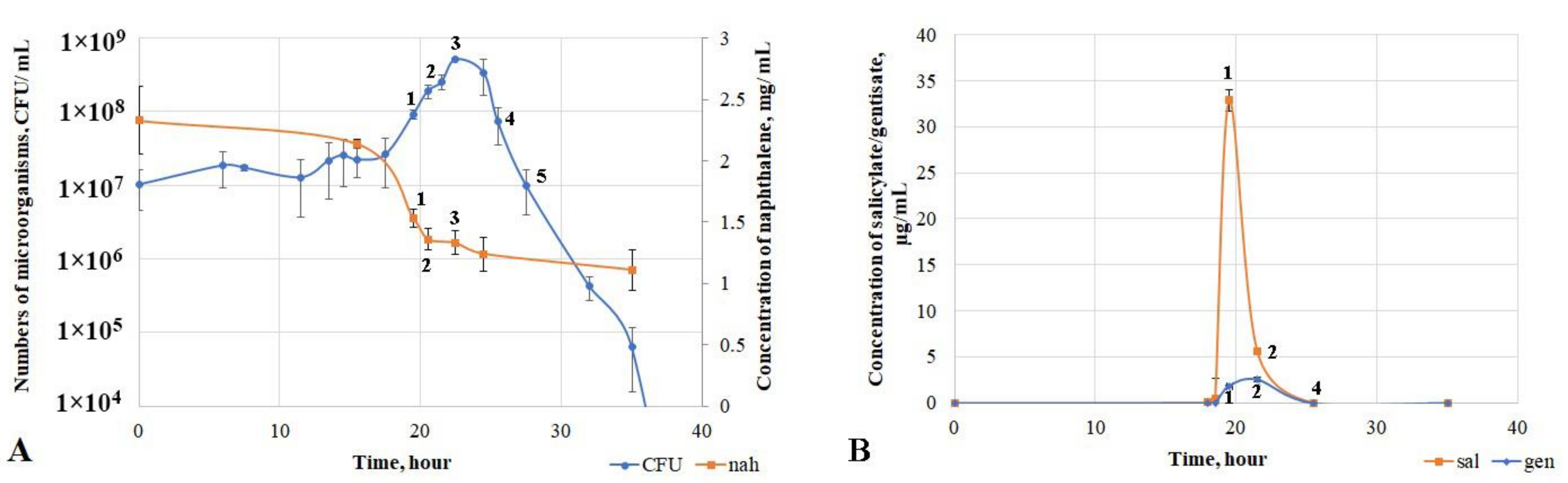
3.3. Physiological and Biochemical Characteristics of D. tsuruhatensis Strain ULwDis3 under Cultivation in Liquid Mineral Medium with Naphthalene or Intermediates of Its Degradation Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sam, K.; Onyena, A.P.; Zabbey, N.; Odoh, C.K.; Nwipie, G.N.; Nkeeh, D.K.; Osuji, L.C.; Little, D.I. Prospects of emerging PAH sources and remediation technologies: Insights from Africa. Environ. Sci. Pollut Res. Int. 2023, 30, 39451–39473. [Google Scholar] [CrossRef] [PubMed]
- Pandolfo, E.; Barra Caracciolo, A.; Rolando, L. Recent Advances in Bacterial Degradation of Hydrocarbons. Water 2023, 15, 375. [Google Scholar] [CrossRef]
- More, M.G.U. Biological Degradation of Naphthalene: A New Era. J. Bioremediat. Biodegrad. 2013, 4, 7. [Google Scholar] [CrossRef]
- Braña, V.; Cagide, C.; Morel, M.A. The Sustainable Use of Delftia in Agriculture, Bioremediation, and Bioproducts Synthesis. In Microbial Models: From Environmental to Industrial Sustainability; Castro-Sowinski, S., Ed.; Springer: Singapore, 2016; Volume 1, pp. 227–247. [Google Scholar] [CrossRef]
- Eren, A.; Güven, K. Isolation and characterization of alkane hydrocarbons-degrading Delftia tsuruhatensis strain D9 from petroleum-contaminated soils. Biotech. Stud. 2022, 31, 36–44. [Google Scholar] [CrossRef]
- Shetty, A.R.; de Gannes, V.; Obi, C.C.; Lucas, S.; Lapidus, A.; Cheng, J.F.; Goodwin, L.A.; Pitluck, S.; Peters, L.; Mikhailova, N.; et al. Complete genome sequence of the phenanthrene-degrading soil bacterium Delftia acidovorans Cs1–4. Stand Genom. Sci. 2015, 10, 55. [Google Scholar] [CrossRef]
- Wu, W.; Huang, H.; Ling, Z.; Yu, Z.; Jiang, Y.; Liu, P.; Li, X. Genome sequencing reveals mechanisms for heavy metal resistance and polycyclic aromatic hydrocarbon degradation in Delftia lacustris strain LZ-C. Ecotoxicology 2016, 25, 234–247. [Google Scholar] [CrossRef]
- Izmalkova, T.Y.; Gafarov, A.B.; Sazonova, O.I.; Sokolov, S.L.; Kosheleva, I.A.; Boronin, A.M. Diversity of Oil-Degrading Microorganisms in the Gulf of Finland (Baltic Sea) in Spring and in Summer. Microbiology 2018, 87, 261–271. [Google Scholar] [CrossRef]
- Ausubel, F.M.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. Short Protocols in Molecular Biology, 4th ed.; John Wiley and Sons, Inc.: New York, NY, USA, 1990. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comp. Biol. J. Comput. Mol. Cell Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods. 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE. 2014, 9, 1–14. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Darling, A.E.; Treangen, T.J.; Messeguer, X.; Perna, N.T. Analyzing patterns of microbial evolution using the mauve genome alignment system. Methods Mol. Biol. 2007, 396, 135–152. [Google Scholar] [CrossRef]
- Olson, R.D.; Assaf, R.; Brettin, T.; Conrad, N.; Cucinell, C.; Davis, J.J.; Dempsey, D.M.; Dickerman, A.; Dietrich, E.M.; Kenyon, R.W.; et al. Introducing the Bacterial and Viral Bioinformatics Resource Center (BV-BRC): A resource combining PATRIC, IRD and ViPR. Nucleic. Acids Res. 2023, 51, 678–689. [Google Scholar] [CrossRef]
- Lee, I.; Ouk Kim, Y.; Park, S.C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Goker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Bacterial and Viral Bioinformatics Resource Center Site. Available online: https://www.bv-brc.org/ (accessed on 4 March 2023).
- The National Center for Biotechnology Information Site. Available online: https://www.ncbi.nlm.nih.gov/genome/?term=delftia (accessed on 10 February 2023).
- Doronina, N.V.; Trotsenko, Y.A.; Tourova, T.P. Methylarcula marina gen. nov.; sp. nov. and Methylarcula terricola sp. nov.: Novel aerobic, moderately halophilic, facultatively methylotrophic bacteria from coastal saline environments. Int. J. Syst. Evol. Microbiol. 2000, 50, 1849–1859. [Google Scholar] [CrossRef]
- Sambrook, J.R. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Dua, R.D.; Meera, S. Purification and characterisation of naphthalene oxygenase from Corynebacterium renale. Eur. J. Biochem. 1981, 120, 461–465. [Google Scholar] [CrossRef]
- Zhou, N.Y.; Al-Dulayymi, J.; Baird, M.S.; Williams, P.A. Salicylate 5-hydroxylase from Ralstonia sp. strain U2: A monooxygenase with close relationships to and shared electron transport proteins with naphthalene dioxygenase. J. Bacteriol. 2002, 184, 1547–1555. [Google Scholar] [CrossRef]
- Crawford, R.L.; Hutton, S.W.; Chapman, P.J. Purification and properties of gentisate 1,2-dioxygenase from Moraxella osloensis. J. Bacteriol. 1975, 121, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Schlomann, M.; Schmidt, E.; Knackmuss, H.J. Different types of dienelactone hydrolase in 4-fluorobenzoate-utilizing bacteria. J. Bacteriol. 1990, 172, 5112–5118. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.; Rodriguez, R.L.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.V.; Maughan, H.; Cameron, A.D.S.; Yost, C.K. Phylogenomic analysis of the genus Delftia reveals distinct major lineages with ecological specializations. Microb. Genom. 2022, 8, mgen000864. [Google Scholar] [CrossRef]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: Implications for the microbial “pan-genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef]
- Hsiao, W.W.; Ung, K.; Aeschliman, D.; Bryan, J.; Finlay, B.B.; Brinkman, F.S. Evidence of a large novel gene pool associated with prokaryotic genomic islands. PLoS Genet. 2005, 1, e62. [Google Scholar] [CrossRef]
- Perna, N.T.; Plunkett, G., 3rd; Burland, V.; Mau, B.; Glasner, J.D.; Rose, D.J.; Mayhew, G.F.; Evans, P.S.; Gregor, J.; Kirkpatrick, H.A.; et al. Genome sequence of enterohaemorrhagic Escherichia coli O157:H7. Nature 2001, 409, 529–533. [Google Scholar] [CrossRef]
- Fuenmayor, S.L.; Wild, M.; Boyes, A.L.; Williams, P.A. A gene cluster encoding steps in conversion of naphthalene to gentisate in Pseudomonas sp. strain U2. J. Bacteriol. 1998, 180, 2522–2530. [Google Scholar] [CrossRef]
- Jeon, C.O.; Park, M.; Ro, H.S.; Park, W.; Madsen, E.L. The naphthalene catabolic (nag) genes of Polaromonas naphthalenivorans CJ2: Evolutionary implications for two gene clusters and novel regulatory control. Appl. Environ. Microbiol. 2006, 72, 1086–1095. [Google Scholar] [CrossRef]
- Jones, R.M.; Britt-Compton, B.; Williams, P.A. The naphthalene catabolic (nag) genes of Ralstonia sp. strain U2 are an operon that is regulated by NagR, a LysR-type transcriptional regulator. J. Bacteriol. 2003, 185, 5847–5853. [Google Scholar] [CrossRef]
- Kincannon, W.M.; Zahn, M.; Clare, R.; Lusty Beech, J.; Romberg, A.; Larson, J.; Bothner, B.; Beckham, G.T.; McGeehan, J.E.; DuBois, J.L. Biochemical and structural characterization of an aromatic ring-hydroxylating dioxygenase for terephthalic acid catabolism. Proc. Natl. Acad. Sci. USA 2022, 119, e2121426119. [Google Scholar] [CrossRef]
- Cane, P.A.; Williams, P.A. The Plasmid-coded Metabolism of Naphthalene and 2-Methylnaphthalene in Pseudomonas Strains: Phenotypic Changes Correlated with Structural Modification of the Plasmid pWW60–1. Microbiology 1982, 128, 2281–2290. [Google Scholar] [CrossRef]
- Davies, J.I.; Evans, W.C. Oxidative metabolism of naphthalene by soil pseudomonads. The ring-fission mechanism. Biochem. J. 1964, 91, 251–261. [Google Scholar] [CrossRef]
- Serdar, C.M.; Gibson, D.T. Isolation and characterization of altered plasmids in mutant strains of Pseudomonas putida NCIB 9816. Biochem. Biophys. Res. Commun. 1989, 164, 764–771. [Google Scholar] [CrossRef]
- Habe, H.; Omori, T. Genetics of polycyclic aromatic hydrocarbon metabolism in diverse aerobic bacteria. Biosci. Biotechnol Biochem. 2003, 67, 225–243. [Google Scholar] [CrossRef]
- Li, W.; Shi, J.; Wang, X.; Han, Y.; Tong, W.; Ma, L.; Liu, B.; Cai, B. Complete nucleotide sequence and organization of the naphthalene catabolic plasmid pND6-1 from Pseudomonas sp. strain ND6. Gene 2004, 336, 231–240. [Google Scholar] [CrossRef]
- Izmalkova, T.Y.; Sazonova, O.I.; Nagornih, M.O.; Sokolov, S.L.; Kosheleva, I.A.; Boronin, A.M. The organization of naphthalene degradation genes in Pseudomonas putida strain AK5. Res. Microbiol. 2013, 164, 244–253. [Google Scholar] [CrossRef]
- Park, M.; Jeon, Y.; Jang, H.H.; Ro, H.S.; Park, W.; Madsen, E.L.; Jeon, C.O. Molecular and biochemical characterization of 3-hydroxybenzoate 6-hydroxylase from Polaromonas naphthalenivorans CJ2. Appl. Environ. Microbiol. 2007, 73, 5146–5152. [Google Scholar] [CrossRef]
- Ishiyama, D.; Vujaklija, D.; Davies, J. Novel pathway of salicylate degradation by Streptomyces sp. strain WA46. Appl. Environ. Microbiol. 2004, 70, 1297–1306. [Google Scholar] [CrossRef]
- Goetz, F.E.; Harmuth, L.J. Gentisate pathway in Salmonella typhimurium: Metabolism of m-hydroxybenzoate and gentisate. FEMS. Microbiol. Lett. 1992, 76, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.C.; Cooper, R.A. Catabolism of 3-hydroxybenzoate by the gentisate pathway in Klebsiella pneumoniae M5a1. Arch. Microbiol. 1990, 154, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Poh, C.L.; Bayly, R.C. Evidence for isofunctional enzymes used in m-cresol and 2,5-xylenol degradation via the gentisate pathway in Pseudomonas alcaligenes. J. Bacteriol. 1980, 143, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Gevers, D.; Vandepoele, K.; Simillon, C.; Van de Peer, Y. Gene duplication and biased functional retention of paralogs in bacterial genomes. Trends. Microbiol. 2004, 12, 148–154. [Google Scholar] [CrossRef]
- van der Meer, J.R. Evolution of novel metabolic pathways for the degradation of chloroaromatic compounds. Antonie Van Leeuwenhoek 1997, 71, 159–178. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type Strain | ANI, % | DDH, % |
|---|---|---|
| Delftia tsuruhatensis NBRC 16741 | 98.6 * | 79.3 * |
| Delftia acidovorans NBRC 14950 | 95.1 | 71.6 |
| Delftia lacustris LMG 24775 | 98.33 | 75.0 |
| Substrate | Point | Time, Hours | Enzyme Activity, μmol × mg−1 Protein | ||
|---|---|---|---|---|---|
| Naphthalene 1.2-Dioxygenase | Salicylate 5-Hydroxylase | Gentisate 1.2-Dioxygenase | |||
| Naphthalene | 1 | 19.5 | 11 | ˂1 | 32 |
| 2 | 21.5 | 49 | 1148 | 269 | |
| 3 | 22.5 | 16 | 858 | 2110 | |
| 4 | 25.5 | ˂1 | ˂1 | 251 | |
| 5 | 27.5 | ˂1 | ˂1 | 77 | |
| Succinate | 13 | ˂1 | 48 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sazonova, O.I.; Ivanova, A.A.; Delegan, Y.A.; Streletskii, R.A.; Vershinina, D.D.; Sokolov, S.L.; Vetrova, A.A. Characterization and Genomic Analysis of the Naphthalene-Degrading Delftia tsuruhatensis ULwDis3 Isolated from Seawater. Microorganisms 2023, 11, 1092. https://doi.org/10.3390/microorganisms11041092
Sazonova OI, Ivanova AA, Delegan YA, Streletskii RA, Vershinina DD, Sokolov SL, Vetrova AA. Characterization and Genomic Analysis of the Naphthalene-Degrading Delftia tsuruhatensis ULwDis3 Isolated from Seawater. Microorganisms. 2023; 11(4):1092. https://doi.org/10.3390/microorganisms11041092
Chicago/Turabian StyleSazonova, Olesya I., Anastasia A. Ivanova, Yanina A. Delegan, Rostislav A. Streletskii, Diana D. Vershinina, Sergei L. Sokolov, and Anna A. Vetrova. 2023. "Characterization and Genomic Analysis of the Naphthalene-Degrading Delftia tsuruhatensis ULwDis3 Isolated from Seawater" Microorganisms 11, no. 4: 1092. https://doi.org/10.3390/microorganisms11041092