
Plant Growth-Promoting Rhizobacteria for Sustainable Agricultural Production

Abstract
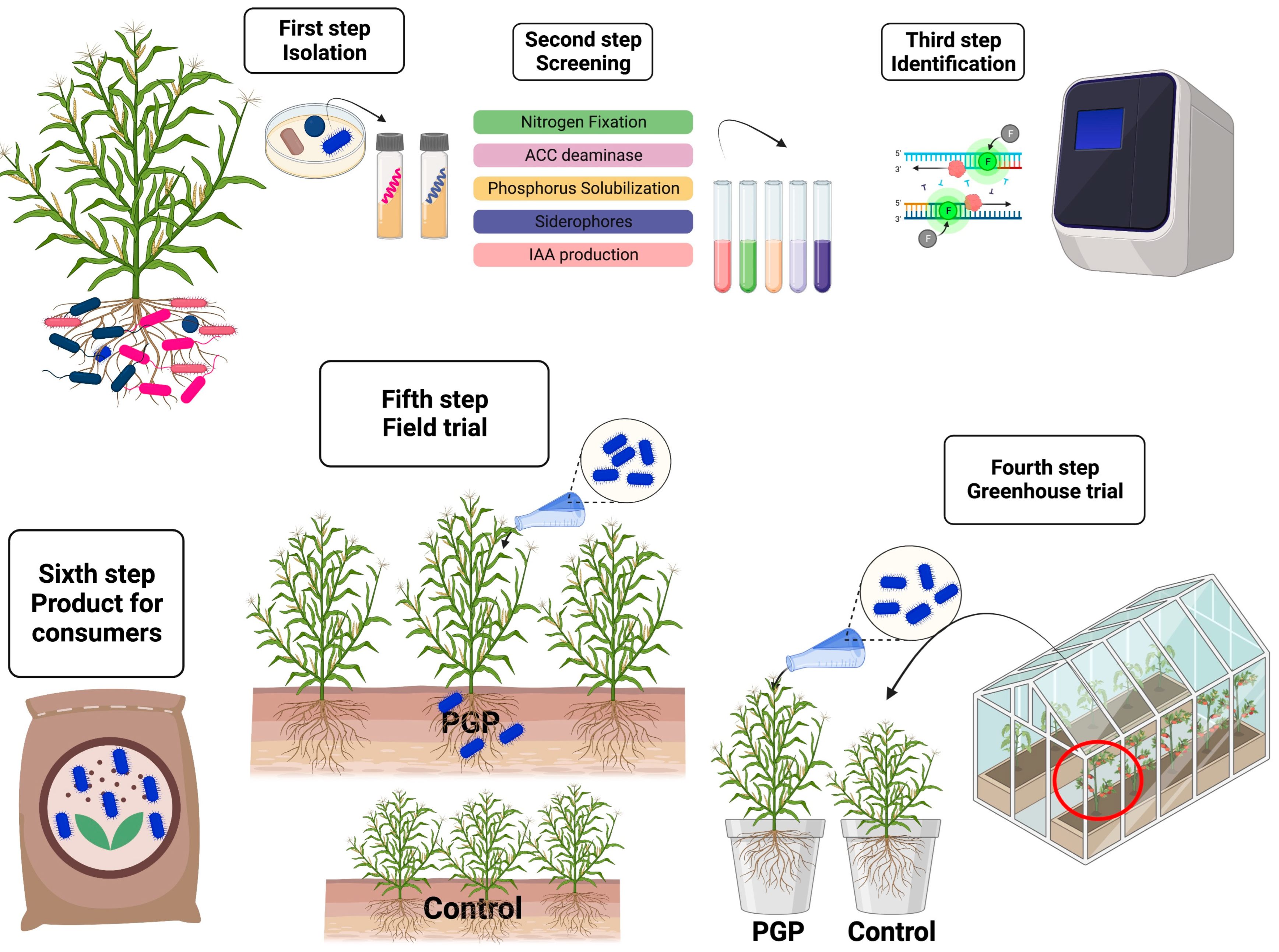
:1. Introduction
2. The Use of Unconventional Fertilizers
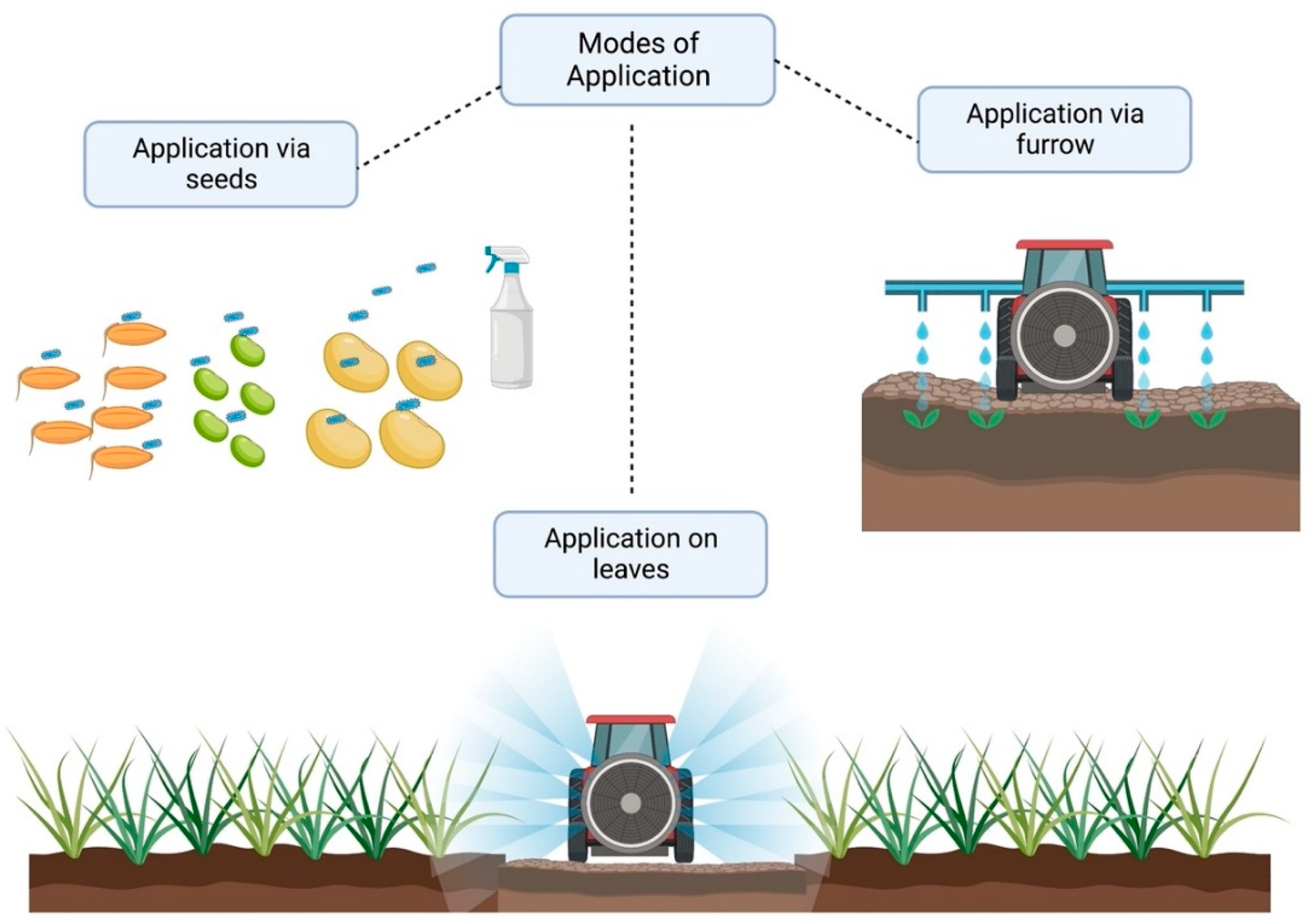
3. Seed Microbiome for Rhizospheric Colonization
4. Rhizospheric Microorganisms
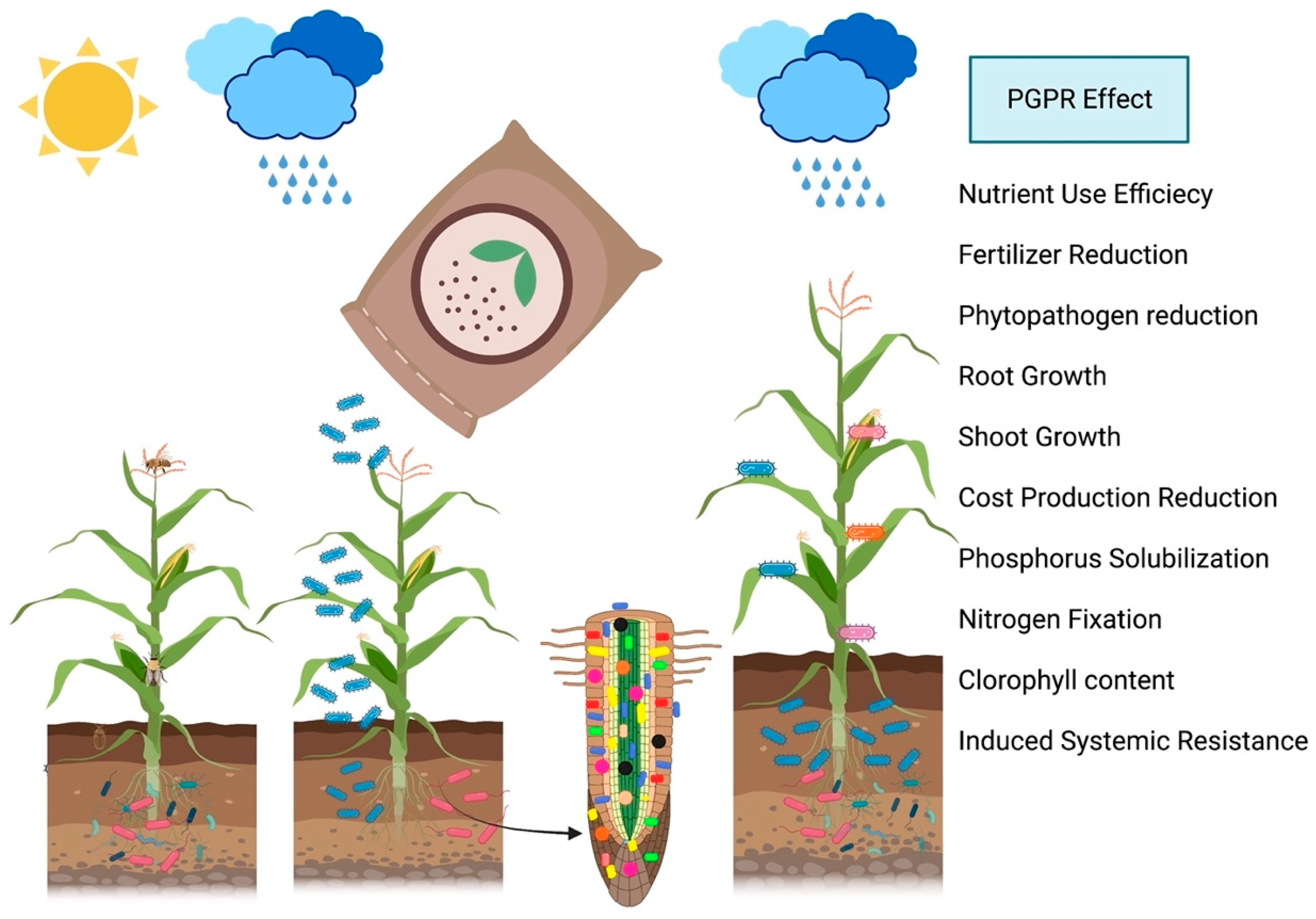
5. Microorganisms Skills to Promote Plant Growth
5.1. Nitrogen Fixation for Reducing Chemical Fertilizers
5.2. Phosphorus Solubilizing and Mineralizing and Siderophore Production
5.3. Phytohormone Production for Reducing Fungicides and Pesticides
6. Nutrient Efficiency Units for Reducing Chemical Fertilizers
7. Future Perspectives
8. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Perez-Montano, F.; Alias-Villegas, C.; Bellogin, R.A.; del Cerro, P.; Espuny, M.R.; Jimenez-Guerrero, I.; Lopez-Baena, F.J.; Ollero, F.J.; Cubo, T. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Vocciante, M.; Grifoni, M.; Fusini, D.; Petruzzelli, G.; Franchi, E. The role of plant growth-promoting rhizobacteria (PGPR) in mitigating plant’s environmental stresses. Appl. Sci. 2022, 12, 1231. [Google Scholar] [CrossRef]
- Bhanse, P.; Kumar, M.; Singh, L.; Awasthi, M.K.; Qureshi, A. Role of plant growth-promoting rhizobacteria in boosting the phytoremediation of stressed soils: Opportunities, challenges, and prospects. Chemosphere 2022, 303, 134954. [Google Scholar] [CrossRef] [PubMed]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef]
- Uzakbaevna, I.A. The Effect of Unconventional Fertilizers on the Growth and Development of Cotton. Int. J. Integr. Educ. 2022, 5, 226–229. [Google Scholar]
- Badr, E.A.; Bakhoum, G.S.; Amin; Howida, G.A.; Khedr, H. Effect of unconventional fertilizers on Root quality and yield components of sugar beet (Beta vulgaris L.) plants. Plants 2022, 11, 2222. [Google Scholar]
- Pajura, R.; Masłoń, A.; Czarnota, J. The Use of Waste to Produce Liquid Fertilizers in Terms of Sustainable Development and Energy Consumption in the Fertilizer Industry—A Case Study from Poland. Energies 2023, 16, 1747. [Google Scholar] [CrossRef]
- Luo, H.; Duan, M.; He, L.; Yang, S.; Zou, Y.; Tang, X. A New Organic-Inorganic Compound Fertilizer for Improving Growth, Yield, and 2-Acetyl-1-Pyrroline Biosynthesis of Fragrant Rice. Agriculture 2021, 11, 1121. [Google Scholar] [CrossRef]
- Dhawi, F. The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming. Metabolites 2023, 13, 247. [Google Scholar] [CrossRef]
- Verma, S.K.; Kharwar, R.N.; White, J.F. The role of seed-vectored endophytes in seedling development and establishment. Symbiosis 2019, 78, 107–113. [Google Scholar] [CrossRef]
- Kuźniar, A.; Włodarczyk, K.; Grządziel, J.; Woźniak, M.; Furtak, K.; Gałązka, A.; Dziadczyk, E.; Skórzyńska-Polit, E.; Wolińska, A. New insight into the composition of wheat seed microbiota. Int. J. Mol. Sci. 2020, 21, 4634. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Urtis-Flores, C.A.; Loeza-Lara, P.D.; del Orozco-Mosqueda, M.C.; Glick, B.R. Rhizosphere colonization determinants by plant growth-promoting rhizobacteria (PGPR). Biology 2021, 10, 475. [Google Scholar] [CrossRef] [PubMed]
- Samreen, T.; Naveed, M.; Nazir, M.Z.; Asghar, H.N.; Khan, M.I.; Zahir, Z.A.; Kanwal, S.; Jeevan, B.; Sharma, D.; Meena, V.S. Seed associated bacterial and fungal endophytes: Diversity, life cycle, transmission, and application potential. Appl. Soil Ecol. 2021, 168, 104191. [Google Scholar] [CrossRef]
- Kumar, A.; Droby, S.; White, J.F.; Singh, V.K.; Singh, S.K.; Zhimo, V.Y.; Biasi, A. Endophytes and seed priming: Agricultural applications and future prospects. In Microbial Endophytes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 107–124. [Google Scholar]
- Li, H.; Parmar, S.; Sharma, V.K.; White, J.F. Seed endophytes and their potential applications. In Seed Endophytes; Verma, S.K., White, J.F., Jr., Eds.; Springer: Cham, Switzerland, 2019; pp. 35–54. [Google Scholar]
- Kandel, S.L.; Joubert, P.M.; Doty, S.L. Bacterial endophyte colonization and distribution within plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [PubMed]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial seed endophytes: Genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 2015, 7, 40–50. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.; Reddy, M.; El Enshasy, H. Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: Recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Pal, G.; Kumar, K.; Verma, A.; Verma, S.K. Seed inhabiting bacterial endophytes of maize promote seedling establishment and provide protection against fungal disease. Microbiol. Res. 2022, 255, 126926. [Google Scholar] [CrossRef]
- Jana, S.K.; Islam, M.M.; Hore, S.; Mandal, S. Rice seed endophytes transmit into the plant seedling, promote plant growth and inhibit fungal phytopathogens. Plant Growth Regul. 2023, 99, 373–388. [Google Scholar] [CrossRef]
- Bowen, G.; Rovira, A. The rhizosphere and its management to improve plant growth. Adv. Agron. 1999, 66, 1–102. [Google Scholar]
- Vetterlein, D.; Carminati, A.; Kögel-Knabner, I.; Bienert, G.P.; Smalla, K.; Oburger, E.; Schnepf, A.; Banitz, T.; Tarkka, M.T.; Schlüter, S. Rhizosphere spatiotemporal organization—A key to rhizosphere functions. Front. Agron. 2020, 2, 8. [Google Scholar] [CrossRef]
- Hassan, M.K.; McInroy, J.A.; Kloepper, J.W. The interactions of rhizodeposits with plant growth-promoting rhizobacteria in the rhizosphere: A review. Agriculture 2019, 9, 142. [Google Scholar] [CrossRef]
- Nuccio, E.E.; Starr, E.; Karaoz, U.; Brodie, E.L.; Zhou, J.; Tringe, S.G.; Malmstrom, R.R.; Woyke, T.; Banfield, J.F.; Firestone, M.K. Niche differentiation is spatially and temporally regulated in the rhizosphere. ISME J. 2020, 14, 999–1014. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Cambon, M.C.; Vacher, C.; Mitter, B.; Samad, A.; Sessitsch, A. The plant endosphere world–bacterial life within plants. Environ. Microbiol. 2021, 23, 1812–1829. [Google Scholar] [CrossRef] [PubMed]
- del Carmen Orozco-Mosqueda, M.; Fadiji, A.E.; Babalola, O.O.; Glick, B.R.; Santoyo, G. Rhizobiome engineering: Unveiling complex rhizosphere interactions to enhance plant growth and health. Microbiol. Res. 2022, 263, 127137. [Google Scholar] [CrossRef] [PubMed]
- Brunel, C.; Pouteau, R.; Dawson, W.; Pester, M.; Ramirez, K.S.; van Kleunen, M. Towards unraveling macroecological patterns in rhizosphere microbiomes. Trends Plant Sci. 2020, 25, 1017–1029. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of plant growth promoting rhizobacteria in grain legumes: Growth promotion and crop production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef]
- Rabari, A.; Ruparelia, J.; Jha, C.K.; Sayyed, R.; Mitra, D.; Priyadarshini, A.; Senapati, A.; Panneerselvam, P.; Mohapatra, P.K.D. Articulating beneficial rhizobacteria mediated plant defenses through induced systemic resistance. Pedosphere 2022, 8, 10. [Google Scholar] [CrossRef]
- Salwan, R.; Sharma, M.; Sharma, A.; Sharma, V. Insights into Plant Beneficial Microorganism-Triggered Induced Systemic Resistance. Plant Stress 2023, 7, 100140. [Google Scholar] [CrossRef]
- Khan, M.; Bhargava, P.; Goel, R. Quorum sensing molecules of Rhizobacteria: A trigger for developing systemic resistance in plants. Plant Growth Promot. Rhizobacteria Sustain. Stress Manag. 2019, 12, 117–138. [Google Scholar]
- Khatoon, Z.; Huang, S.; Rafique, M.; Fakhar, A.; Kamran, M.A.; Santoyo, G. Unlocking the potential of plant growth-promoting rhizobacteria on soil health and the sustainability of agricultural systems. J. Environ. Manag. 2020, 273, 111118. [Google Scholar] [CrossRef]
- Ahluwalia, O.; Singh, P.C.; Bhatia, R. A review on drought stress in plants: Implications, mitigation and the role of plant growth promoting rhizobacteria. Resour. Environ. Sustain. 2021, 5, 100032. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salt-tolerant plant growth promoting rhizobacteria for enhancing crop productivity of saline soils. Front. Microbiol. 2019, 10, 2791. [Google Scholar] [CrossRef] [PubMed]
- Bhavya, K.; Geetha, A. Plant growth promoting rhizobacteria. Adv. Agric. Sci. 2021, 61, 87. [Google Scholar]
- Riaz, U.; Murtaza, G.; Anum, W.; Samreen, T.; Sarfraz, M.; Nazir, M.Z. Plant Growth-Promoting Rhizobacteria (PGPR) as biofertilizers and biopesticides. In Microbiota and Biofertilizers; Springer: Berlin/Heidelberg, Germany, 2021; pp. 181–196. [Google Scholar]
- Wickramasinghe, W.; Girija, D.; Gopal, K.S.; Kesevan, S. Multi-phasic nitrogen fixing plant growth promoting rhizobacteria as biofertilizer for rice cultivation. Res. J. Agric. Sci. 2021, 12, 399–404. [Google Scholar]
- Hussain, M.B.; Shah, S.H.; Matloob, A.; Mubaraka, R.; Ahmed, N.; Ahmad, I.; Jamshaid, M.U. Rice Interactions with Plant Growth Promoting Rhizobacteria. In Modern Techniques of Rice Crop Production; Springer: Berlin/Heidelberg, Germany, 2022; pp. 231–255. [Google Scholar]
- Soumare, A.; Diédhiou, A.G.; Arora, N.K.; Tawfeeq Al-Ani, L.K.; Ngom, M.; Fall, S.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L.; Sy, M.O. Potential role and utilization of plant growth promoting microbes in plant tissue culture. Front. Microbiol. 2021, 12, 649878. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liao, Z.; Chew, H.; Wang, L.; Lin, B.; Chen, C.; Lu, G.; Lin, Z. Effect of Pennisetum giganteum z.x. lin mixed nitrogen-fixing bacterial fertilizer on the growth, quality, soil fertility and bacterial community of pakchoi (Brassica chinensis L.). PLoS ONE 2020, 15, e0228709. [Google Scholar] [CrossRef]
- Song, X.; Zhang, J.; Peng, C.; Li, D. Replacing nitrogen fertilizer with nitrogen-fixing cyanobacteria reduced nitrogen leaching in red soil paddy fields. Agric. Ecosyst. Environ. 2021, 312, 107320. [Google Scholar] [CrossRef]
- Tapia-García, E.Y.; Hernández-Trejo, V.; Guevara-Luna, J.; Rojas-Rojas, F.U.; Arroyo-Herrera, I.; Meza-Radilla, G.; Vásquez-Murrieta, M.S.; Estrada-de Los Santos, P. Plant growth-promoting bacteria isolated from wild legume nodules and nodules of Phaseolus vulgaris L. trap plants in central and southern Mexico. Microbiol. Res. 2020, 239, 126522. [Google Scholar] [CrossRef]
- Sheoran, S.; Kumar, S.; Kumar, P.; Meena, R.S.; Rakshit, S. Nitrogen fixation in maize: Breeding opportunities. Theor. Appl. Genet. 2021, 134, 1263–1280. [Google Scholar] [CrossRef]
- Pandey, A.; Sharma, E.; Palni, L.M.S. Influence of bacterial inoculation on maize in upland farming systems of the Sikkim Himalaya. Soil Biol. Biochem. 1998, 30, 379–384. [Google Scholar] [CrossRef]
- Khan, A.A.; Jilani, G.; Akhtar, M.S.; Naqvi, S.M.S.; Rasheed, M. Phosphorus solubilizing bacteria: Occurrence, mechanisms and their role in crop production. J. Agric. Biol. Sci. 2009, 1, 48–58. [Google Scholar]
- Wan, W.; Qin, Y.; Wu, H.; Zuo, W.; He, H.; Tan, J.; Wang, Y.; He, D. Isolation and characterization of phosphorus solubilizing bacteria with multiple phosphorus sources utilizing capability and their potential for lead immobilization in soil. Front. Microbiol. 2020, 11, 752. [Google Scholar] [CrossRef] [PubMed]
- Nath, D.; Maurya, B.R.; Meena, V.S. Documentation of five potassium-and phosphorus-solubilizing bacteria for their K and P-solubilization ability from various minerals. Biocatal. Agric. Biotechnol. 2017, 10, 174–181. [Google Scholar] [CrossRef]
- Wang, L.; Lin, H.; Dong, Y.; Li, B.; He, Y. Effects of endophytes inoculation on rhizosphere and endosphere microecology of Indian mustard (Brassica juncea) grown in vanadium-contaminated soil and its enhancement on phytoremediation. Chemosphere 2020, 240, 124891. [Google Scholar] [CrossRef]
- Liu, J.; Qi, W.; Li, Q.; Wang, S.-G.; Song, C.; Yuan, X.-z. Exogenous phosphorus-solubilizing bacteria changed the rhizosphere microbial community indirectly. 3 Biotech 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pantigoso, H.A.; Manter, D.K.; Fonte, S.J.; Vivanco, J.M. Root exudate-derived compounds stimulate the phosphorus solubilizing ability of bacteria. Sci. Rep. 2023, 13, 4050. [Google Scholar] [CrossRef] [PubMed]
- Kour, D.; Rana, K.L.; Yadav, N.; Yadav, A.N. Bioprospecting of phosphorus solubilizing bacteria from Renuka Lake ecosystems, lesser Himalayas. J. Appl. Biol. Biotechnol. 2019, 7, 1–6. [Google Scholar]
- Da Silva, L.I.; Pereira, M.C.; de Carvalho, A.M.X.; Buttrós, V.H.; Pasqual, M.; Dória, J. Phosphorus-Solubilizing Microorganisms: A Key to Sustainable Agriculture. Agriculture 2023, 13, 462. [Google Scholar] [CrossRef]
- Din, M.; Nelofer, R.; Salman, M.; Khan, F.H.; Khan, A.; Ahmad, M.; Jalil, F.; Din, J.U.; Khan, M. Production of nitrogen fixing Azotobacter (SR-4) and phosphorus solubilizing Aspergillus niger and their evaluation on Lagenaria siceraria and Abelmoschus esculentus. Biotechnol. Rep. 2019, 22, e00323. [Google Scholar] [CrossRef]
- Gandhi, N.U.; Chandra, S.B. A comparative analysis of three classes of bacterial non-specific acid phosphatases and archaeal phosphoesterases: Evolutionary perspective. Acta Inform. Med. 2012, 20, 167. [Google Scholar] [CrossRef]
- Rejsek, K.; Vranova, V.; Formanek, P. Determination of the proportion of total soil extracellular acid phosphomonoesterase (EC 3.1. 3.2) activity represented by roots in the soil of different forest ecosystems. Sci. World J. 2012, 2012, 250805. [Google Scholar] [CrossRef] [PubMed]
- Silva, U.C.; Cuadros-Orellana, S.; Silva, D.R.; Freitas-Júnior, L.F.; Fernandes, A.C.; Leite, L.R.; Oliveira, C.A.; Dos Santos, V.L. Genomic and phenotypic insights into the potential of rock phosphate solubilizing bacteria to promote millet growth in vivo. Front. Microbiol. 2021, 11, 574550. [Google Scholar] [CrossRef] [PubMed]
- McLaren, T.I.; Smernik, R.J.; McLaughlin, M.J.; Doolette, A.L.; Richardson, A.E.; Frossard, E. The chemical nature of soil organic phosphorus: A critical review and global compilation of quantitative data. Adv. Agron. 2020, 160, 51–124. [Google Scholar]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Sultana, S.; Alam, S.; Karim, M.M. Screening of siderophore-producing salt-tolerant rhizobacteria suitable for supporting plant growth in saline soils with iron limitation. J. Agric. Food Res. 2021, 4, 100150. [Google Scholar] [CrossRef]
- Delaporte-Quintana, P.; Lovaisa, N.C.; Rapisarda, V.A.; Pedraza, R.O. The plant growth promoting bacteria Gluconacetobacter diazotrophicus and Azospirillum brasilense contribute to the iron nutrition of strawberry plants through siderophores production. Plant Growth Regul. 2020, 91, 185–199. [Google Scholar] [CrossRef]
- Ferreira, C.M.; Soares, H.M.; Soares, E.V. Promising bacterial genera for agricultural practices: An insight on plant growth-promoting properties and microbial safety aspects. Sci. Total Environ. 2019, 682, 779–799. [Google Scholar] [CrossRef]
- Ghazy, N.; El-Nahrawy, S. Siderophore production by Bacillus subtilis MF497446 and Pseudomonas koreensis MG209738 and their efficacy in controlling Cephalosporium maydis in maize plant. Arch. Microbiol. 2021, 203, 1195–1209. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, S.; Upadhyay, N. Effects of organophosphate pesticides on siderophore producing soils microorganisms. Biocatal. Agric. Biotechnol. 2019, 21, 101359. [Google Scholar] [CrossRef]
- Sukul, P.; Kumar, J.; Rani, A.; Abdillahi, A.M.; Rakesh, R.B.; Kumar, M.H. Functioning of plant growth promoting rhizobacteria (PGPR) and their mode of actions: An overview from chemistry point of view. Plant Arch. 2021, 21, 628–634. [Google Scholar] [CrossRef]
- Khan, S.; Shahid, M.; Khan, M.S.; Syed, A.; Bahkali, A.H.; Elgorban, A.M.; Pichtel, J. Fungicide-tolerant plant growth-promoting rhizobacteria mitigate physiological disruption of white radish caused by fungicides used in the field cultivation. Int. J. Environ. Res. Public Health 2020, 17, 7251. [Google Scholar] [CrossRef] [PubMed]
- Al-Enazi, N.M.; AlTami, M.S.; Alhomaidi, E. Unraveling the potential of pesticide-tolerant Pseudomonas sp. augmenting biological and physiological attributes of Vigna radiata (L.) under pesticide stress. RSC Adv. 2022, 12, 17765–17783. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Kang, J.P.; Ahn, J.C.; Kim, Y.J.; Piao, C.H.; Yang, D.U.; Yang, D.C. Siderophore-producing rhizobacteria reduce heavy metal-induced oxidative stress in Panax ginseng Meyer. J. Ginseng Res. 2021, 45, 218–227. [Google Scholar] [CrossRef]
- Gao, B.; Chai, X.; Huang, Y.; Wang, X.; Han, Z.; Xu, X.; Wu, T.; Zhang, X.; Wang, Y. Siderophore production in pseudomonas SP. strain SP3 enhances iron acquisition in apple rootstock. J. Appl. Microbiol. 2022, 133, 720–732. [Google Scholar] [CrossRef] [PubMed]
- Ahirwar, N.K.; Singh, R.; Chaurasia, S.; Chandra, R.; Ramana, S. Effective role of beneficial microbes in achieving the sustainable agriculture and eco-friendly environment development goals: A review. Front. Microbiol. 2020, 5, 111–123. [Google Scholar] [CrossRef]
- Hacisalihoglu, G. Zinc (Zn): The last nutrient in the alphabet and shedding light on Zn efficiency for the future of crop production under suboptimal Zn. Plants 2020, 9, 1471. [Google Scholar] [CrossRef]
- Mosttafiz, S.; Rahman, M.; Rahman, M. Biotechnology: Role of microbes in sustainable agriculture and environmental health. Internet J. Microbiol. 2012, 10, 1–6. [Google Scholar]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.-Y.; Qin, F. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Cipriano, M.A.P.; de Freitas-Iório, R.P.; Dimitrov, M.R.; de Andrade, S.A.L.; Kuramae, E.E.; da Silveira, A.P.D. Plant-growth endophytic bacteria improve nutrient use efficiency and modulate foliar N-metabolites in sugarcane seedling. Microorganisms 2021, 9, 479. [Google Scholar] [CrossRef]
- Rahim, A.; Ranjha, A.; Waraich, E. Effect of phosphorus application and irrigation scheduling on wheat yield and phosphorus use efficiency. Soil Environ. 2010, 29, 15–22. [Google Scholar]
- Wang, J.; Li, R.; Zhang, H.; Wei, G.; Li, Z. Beneficial bacteria activate nutrients and promote wheat growth under conditions of reduced fertilizer application. BMC Microbiol. 2020, 20, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.B.; Sahai, V.; Goyal, D.; Prasad, M.; Yadav, A.; Shrivastav, P.; Ali, A.; Dantu, P.K. Identification, characterization and evaluation of multifaceted traits of plant growth promoting rhizobacteria from soil for sustainable approach to agriculture. Curr. Microbiol. 2020, 77, 3633–3642. [Google Scholar] [CrossRef] [PubMed]
- Bashir, S.; Anwar, S.; Ahmad, B.; Sarfraz, Q.; Khatk, W.; Islam, M. Response of wheat crop to phosphorus levels and application methods. J. Environ. Earth Sci. 2015, 5, 151–155. [Google Scholar]
- Gao, Y.; Fang, Z.; Van Zwieten, L.; Bolan, N.; Dong, D.; Quin, B.F.; Meng, J.; Li, F.; Wu, F.; Wang, H. A critical review of biochar-based nitrogen fertilizers and their effects on crop production and the environment. Biochar 2022, 4, 1–19. [Google Scholar] [CrossRef]
- Badri, D.V.; Weir, T.L.; Van der Lelie, D.; Vivanco, J.M. Rhizosphere chemical dialogues: Plant-microbe interactions. Curr. Opin. Biotechnol. 2009, 20, 642–650. [Google Scholar] [CrossRef]
- Kumar, A.; Patel, J.; Meena, V.S.; Ramteke, P. Plant growth-promoting rhizobacteria: Strategies to improve abiotic stresses under sustainable agriculture. J. Plant Nutr. 2019, 42, 1402–1415. [Google Scholar] [CrossRef]
- Sun, C.X.; Bei, K.; Liu, Y.H.; Pan, Z.Y. Humic Acid Improves Greenhouse Tomato Quality and Bacterial Richness in Rhizosphere Soil. ACS Omega 2022, 7, 29823–29831. [Google Scholar] [CrossRef]
- Wang, H.; Ma, S.; Xia, Q.; Zhao, Z.; Chen, X.; Shen, X.; Yin, C.; Mao, Z. The interaction of the pathogen Fusarium proliferatum with Trichoderma asperellum characterized by transcriptome changes in apple rootstock roots. Physiol. Mol. Plant Pathol. 2022, 121, 101894. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| PGPR | Abilities | Condition | Results | References |
|---|---|---|---|---|
| G. diazotrophicus and A. brasilense | Siderophore production | Hydroponia | Increased iron in the plants | [60] |
| Pseudomonas sp. | Siderophore production and PGPR properties | Vase with vermiculite | Iron acquisition | [78] |
| Lysinibacillus sp. and Paenibacillus dendritiformis | Seedling protection | Maize in vase | Improved seed germination | [19] |
| Bacillus subtilis | Seedling protection | Pot experiment | Protection against Cephalosporium maydis | [62] |
| Bacillus sp. | IAA production Phosphorus solubilization | Greenhouse | Increased rice seedling Increase P availability | [79] |
| Mesorhizobium panacihumi | Siderophore production | Pot tests | Reduced soil contamination | [67] |
| Bacillus sp. Citrobacter sp. Citrobacter sp. Flavobacterium sp. and Pantoea sp. | Phytohormones production; siderophore; hydrogen cyanide | Greenhouse | Reduced fungal phytopathogens | [20] |
| Pseudomonas sp. | Protection against fungicide | Greenhouse | Improved cultivation of radish plants | [65] |
| Pseudomonas fluorescens Rhizobium leguminosarum Bacillus brevis Azotobacter vinelandii | Siderophore production | Chamber | Protection against organophosphate pesticides | [80] |
| Anabaena azotica | Nitrogen fixation | Field experiment | Chemical nitrogen reduction | [41] |
| Brucella sp. and Pseudomonas brassicae | Siderophore production | Field experiment | High production in iron-deficient soil | [81] |
| Bacillus aryabhattai | Siderophore production | Field experiment | Increased production in iron-deficient soil | [59] |
| Rhodopseudomonas palustris | Phosphorus solubilization | Greenhouse | Improved soil fertility | [82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Andrade, L.A.; Santos, C.H.B.; Frezarin, E.T.; Sales, L.R.; Rigobelo, E.C. Plant Growth-Promoting Rhizobacteria for Sustainable Agricultural Production. Microorganisms 2023, 11, 1088. https://doi.org/10.3390/microorganisms11041088
de Andrade LA, Santos CHB, Frezarin ET, Sales LR, Rigobelo EC. Plant Growth-Promoting Rhizobacteria for Sustainable Agricultural Production. Microorganisms. 2023; 11(4):1088. https://doi.org/10.3390/microorganisms11041088
Chicago/Turabian Stylede Andrade, Luana Alves, Carlos Henrique Barbosa Santos, Edvan Teciano Frezarin, Luziane Ramos Sales, and Everlon Cid Rigobelo. 2023. "Plant Growth-Promoting Rhizobacteria for Sustainable Agricultural Production" Microorganisms 11, no. 4: 1088. https://doi.org/10.3390/microorganisms11041088