
Identification of Fusarium oxysporum f. sp. lactucae Race 1 as the Causal Agent of Lettuce Fusarium Wilt in Greece, Commercial Cultivars’ Susceptibility, and Temporal Expression of Defense-Related Genes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Pathogen Isolation
2.2. Molecular Identification of Fungal Strains
2.2.1. DNA Extraction
2.2.2. PCR Amplification
2.2.3. Sequencing and Phylogenetic Analysis
2.3. Pathogenicity Tests
2.3.1. Plant Material
2.3.2. Inoculation Procedure
2.3.3. Estimation of Disease Index
2.4. Defense Gene Expression Measurements
2.4.1. Plant Material for Gene Expression Measurements and RNA Preparation
2.4.2. Selection of Defense-Related Genes for Expression Measurements
2.4.3. Quantification of Gene Expression Levels with RT-qPCR
3. Results
3.1. Pathogen Identification and Phylogenetic Analysis of the Isolates
3.2. Race Determination with Race 1- and Race 4-Specific PCR Primers
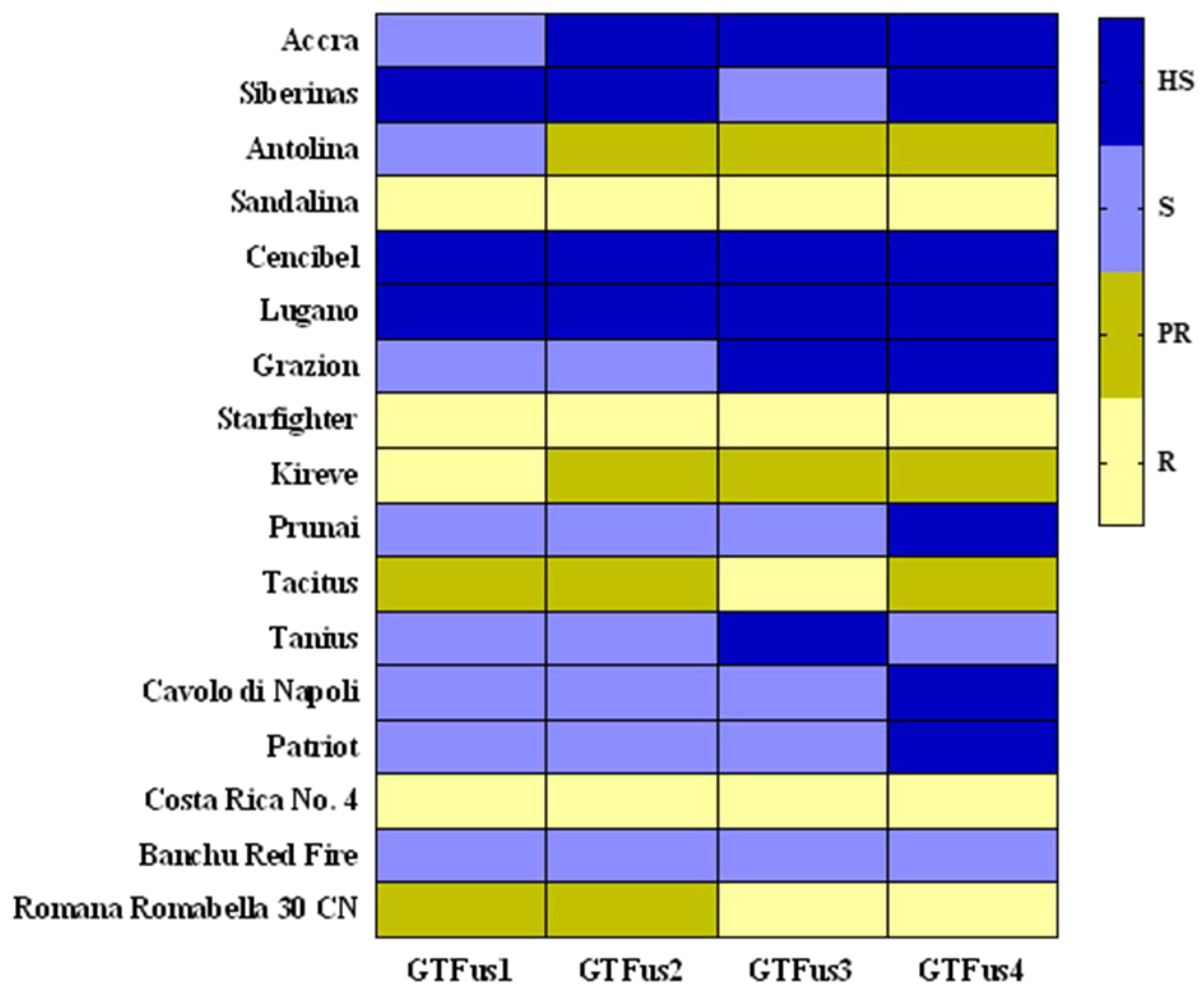
3.3. Pathogenicity Test and Race Determination Based on a Set of Differential Lettuce Cultivars
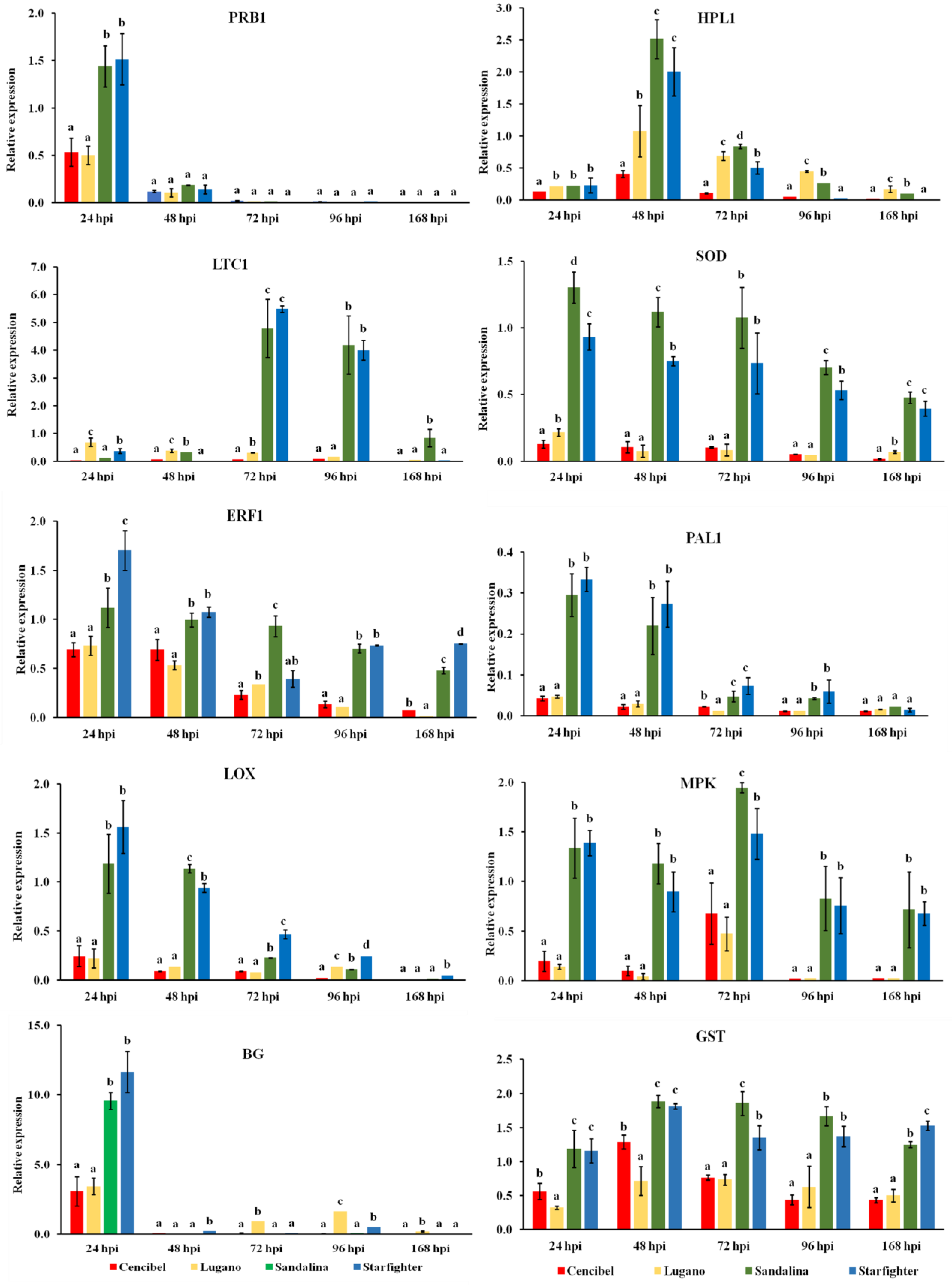
3.4. Defense Genes’ Expression in the Resistant and Susceptible Lettuce Cultivars
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matuo, T.; Motohashi, S. On Fusarium oxysporum f. sp. lactucae n. f. causing root rot of lettuce. Trans. Mycol. Soc. Japan 1967, 8, 13–15. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 22 February 2023).
- Gullino, M.L.; Gilardi, G.; Garibaldi, A. Ready-to-eat salad crops: A plant pathogen’s heaven. Plant Dis. 2019, 103, 2153–2170. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, A.; Gilardi, G.; Gullino, M.L. First report of Fusarium oxysporum on lettuce in Europe. Plant Dis. 2002, 86, 1052. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, M.; Dematheis, F.; Gullino, M.L.; Garibaldi, A. Identification of race 1 of Fusarium oxysporum f. sp. lactucae on lettuce by inter-retrotranposon sequence-characterized amplified region technique. Phytopathology 2007, 97, 987–996. [Google Scholar] [CrossRef]
- Fujinaga, M.; Ogiso, H.; Tsuchiya, N.; Saito, H. Physiological specialization of Fusarium oxysporum f. sp. lactucae, a causal organism of fusarium root of crisphead lettuce in Japan. J. Gen. Plant Pathol. 2001, 667, 205–206. [Google Scholar] [CrossRef]
- Fujinaga, M.; Ogiso, H.; Tuchiya, N.; Saito, H.; Yamanaka, S.; Nozue, M.; Kojima, M. Race 3, a new race of Fusarium oxysporum f. sp. lactucae determined by a differential system with commercial cultivars. J. Gen. Plant Pathol. 2003, 69, 23–28. [Google Scholar] [CrossRef]
- Claerbout, J.; Venneman, S.; Vandevelde, I.; Decombel, A.; Bleyaert, P.; Volckaert, A.; Neukermans, J.; Hofte, M. First report of Fusarium oxysporum f. sp. lactucae race 4 on lettuce in Belgium. Plant Dis. 2018, 102, 1037. [Google Scholar] [CrossRef]
- Gilardi, G.; Franco Ortega, S.; Rijswick, P.; Ortu, G.; Gullino, M.L.; Garibaldi, A. A new race of Fusarium oxysporum f. sp. lactucae of lettuce. Plant Pathol. 2017, 66, 677–688. [Google Scholar] [CrossRef]
- Taylor, A.; Jackson, A.C.; Clarkson, J.P. First report of Fusarium oxysporum f. sp. lactucae race 4 causing lettuce wilt in England and Ireland. Plant Dis. 2019, 103, 1033. [Google Scholar] [CrossRef]
- Gilardi, G.; Garibaldi, A.; Matic, S.; Senatore, M.T.; Pipponzi, S.; Prodi, A.; Gullino, M.L. First report of Fusarium oxysporum f. sp. lactucae race 4 on lettuce in Italy. Plant Dis. 2019, 103, 2680. [Google Scholar] [CrossRef]
- Ogiso, H.; Fujinaga, M.; Saito, H.; Takehara, T.; Yamanaka, S. Physiological races and vegetative compatibility groups of Fusarium oxysporum f. sp. lactucae isolated from crisphead lettuce in Japan. J. Gen. Plant Pathol. 2002, 68, 292–299. [Google Scholar] [CrossRef]
- Yamauchi, N.; Shimazu, J.; Horiuchi, S.; Satou, M.; Horiuchi, S.; Shirakawa, T. Physiological races and vegetative compatibility groups of butterhead lettuce isolates of Fusarium oxysporum f. sp. lactucae in Japan. J. Gen. Plant Pathol. 2004, 70, 308–313. [Google Scholar] [CrossRef]
- Fujinaga, M.; Ogiso, H.; Shinohara, H.; Tsushima, S.; Nishimura, N.; Togawa, M.; Saito, H.; Nozue, M. Phylogenetic relationships between the lettuce root rot pathogen Fusarium oxysporum f. sp. lactucae races 1, 2, and 3 based on the sequence of the intergenic spacer region of its ribosomal DNA. J. Gen. Plant Pathol. 2005, 71, 402–407. [Google Scholar] [CrossRef]
- Pasquali, M.; Dematheis, F.; Gilardi, G.; Gullino, M.L.; Garibaldi, A. Vegetative compatibility groups of Fusarium oxysporum f. sp. lactucae from lettuce. Plant Dis. 2005, 89, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Mbofung, G.Y.; Hong, S.; Pryor, B. Phylogeny of Fusarium oxysporum f. sp. lactucae inferred from mitochondrial small subunit, elongation factor 1-alpha, and nuclear ribosomal intergenic spacer sequence data. Phytopathology 2007, 97, 87–98. [Google Scholar] [CrossRef]
- Pintore, I.; Gilardi, G.; Gullino, M.L.; Garibaldi, A. Analysis of vegetative compatibility groups of Italian and Dutch isolates of Fusarium oxysporum f. sp. lactucae. J. Plant Pathol. 2017, 99, 517–521. [Google Scholar]
- Gordon, T.R.; Martyn, R.D. The evolutionary biology of Fusarium oxysporum. Annu. Rev. Phytopathol. 1997, 35, 111–128. [Google Scholar] [CrossRef]
- Mbofung, G.Y.; Pryor, B.M. A PCR-based assay for detection of Fusarium oxysporum f. sp. lactucae in lettuce seed. Plant Dis. 2010, 94, 860–866. [Google Scholar] [CrossRef]
- Paugh, K.R.; Gordon, T.R. The population of Fusarium oxysporum f. sp. lactucae in California and Arizona. Plant Dis. 2020, 104, 1811–1816. [Google Scholar] [CrossRef]
- Edel-Hermann, V.; Lecomte, C. Current status of Fusarium oxysporum formae speciales and races. Phytopathology 2019, 109, 512–530. [Google Scholar] [CrossRef]
- Ma, L.-J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, M.S.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Wong, C.L.; Muzzi, F.; Vlaardingerbroek, I.; Kidd, B.N.; Schenk, P.M. Root defense analysis against Fusarium oxysporum reveals new regulators to confer resistance. Sci. Rep. 2014, 4, 5584. [Google Scholar] [CrossRef]
- Lyons, R.; Stiller, J.; Powell, J.; Rusu, A.; Manners, J.M.; Kazan, K. Fusarium oxysporum triggers tissue-specific transcriptional reprogramming in Arabidopsis thaliana. PLoS ONE 2015, 10, e0121902. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef]
- Truman, W.; Bennett, M.H.; Kubigsteltig, I.; Turnbull, C.; Grant, M. Arabidopsis systemic immunity uses conserved defense signaling pathways and is mediated by jasmonates. Proc. Natl. Acad. Sci. USA 2007, 104, 1075–1080. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Li, J.; Xie, C.; Jian, W.; Yang, X. An overview of the molecular genetics of plant resistance to the Verticillium wilt pathogen Verticillium dahliae. Int. J. Mol. Sci. 2020, 21, 1120. [Google Scholar] [CrossRef]
- Beckman, C.H.; Roberts, E.M. On the nature and genetic basis for resistance and tolerance to fungal wilt diseases of plants. Adv. Bot. Res. 1995, 21, 35–77. [Google Scholar] [CrossRef]
- Berrocal-Lobo, M.; Molina, A. Arabidopsis defense response against Fusarium oxysporum. Trends Plant Sci. 2008, 13, 145–150. [Google Scholar] [CrossRef]
- Di, X.; Gomila, J.; Takken, F.L.W. Involvement of salicylic acid, ethylene and jasmonic acid signalling pathways in the susceptibility of tomato to Fusarium oxysporum. Mol. Plant Pathol. 2017, 18, 1024–1035. [Google Scholar] [CrossRef]
- Gurjar, G.; Giri, A.; Gupta, V. Gene expression profiling during wilting in chickpea caused by Fusarium oxysporum f. sp. ciceri. Am. J. Plant Sci. 2012, 3, 190–201. [Google Scholar] [CrossRef]
- Jadhav, P.R.; Mahatma, M.K.; Mahatma, L.; Jha, S.; Parekh, V.B.; Khandelwal, V. Expression analysis of key genes of phenylpropanoid pathway and phenol profiling during Ricinus communis–Fusarium oxysporum f. sp. ricini interaction. Ind. Crops Prod. 2013, 50, 456–461. [Google Scholar] [CrossRef]
- Patil, V.R.; Patel, R.M.; Parekh, V.B.; Pathak, J.; Saripalli, G. Differential gene expression analyses of ten defence response genes during Fusarium wilt infection in resistant and susceptible pigeonpea cultivars. Plant Stress 2021, 2, 100043. [Google Scholar] [CrossRef]
- De Cremer, K.; Mathys, J.; Vos, C.; Froenicke, L.; Michelmore, R.W.; Cammue, B.P.A.; De Coninck, B. RNA seq-based transcriptome analysis of Lactuca sativa infected by the fungal necrotroph Botrytis cinerea. Plant Cell Environ. 2013, 36, 1992–2007. [Google Scholar] [CrossRef] [PubMed]
- Almatwari, A.H.A.; Hassandokht, M.; Soltani, F.; Mirzadi Gohari, A.; Javan-Nikkhah, M. Temporal expression profiles of defense-related genes involved in Lactuca sativa-Sclerotinia sclerotiorum interactions. Plant Pathol. J. 2021, 103, 61–69. [Google Scholar] [CrossRef]
- Breen, S.; Williams, S.J.; Outram, M.; Kobe, B.; Solomon, P.S. Emerging insights into the functions of pathogenesis-related protein 1. Trends Plant Sci. 2017, 22, 871–879. [Google Scholar] [CrossRef]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef]
- Matsui, K.; Minami, A.; Hornung, E.; Shibata, H.; Kishimoto, K.; Ahnert, V.; Kindl, H.; Kajiwara, T.; Feussner, I. Biosynthesis of fatty acid derived aldehydes is induced upon mechanical wounding and its products show fungicidal activities in cucumber. Phytochemistry 2006, 67, 649–657. [Google Scholar] [CrossRef]
- Yadav, H.; Dreher, D.; Athmer, B.; Porzel, A.; Gavrin, A.; Baldermann, S.; Tissier, A.; Hause, B. Medicago TERPENE SYNTHASE 10 is involved in defense against an oomycete root pathogen. Plant Physiol. 2019, 180, 1598–1613. [Google Scholar] [CrossRef]
- Maurya, R.; Namdeo, M. Superoxide dismutase: A key enzyme for the survival of intracellular pathogens in host. In Reactive Oxygen Species; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Lorenzo, O.; Piqueras, R.; Sánchez-Serrano, J.J.; Solano, R. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell 2003, 15, 165–178. [Google Scholar] [CrossRef]
- Zvirin, T.; Herman, R.; Brotman, Y.; Denisov, Y.; Belausov, E.; Freeman, S.; Perl-Treves, R. Differential colonization and defence responses of resistant and susceptible melon lines infected by Fusarium oxysporum race 1·2. Plant Pathol. 2010, 59, 576–585. [Google Scholar] [CrossRef]
- Bowles, D.J. Defense-related proteins in higher plants. Annu. Rev. Biochem. 1990, 59, 873–907. [Google Scholar] [CrossRef] [PubMed]
- Bent, A.F. Plant mitogen-activated protein kinase cascades: Negative regulatory roles turn out positive. Proc. Natl. Acad. Sci. USA 2001, 98, 784–786. [Google Scholar] [CrossRef] [PubMed]
- Tena, G.; Asai, T.; Chiu, W.L.; Sheen, J. Plant mitogen-activated protein kinase signaling cascades. Curr. Opin. Plant Biol. 2001, 4, 392–400. [Google Scholar] [CrossRef]
- Lozovaya, V.V.; Waranyuwat, A.; Widholm, J.M. β-l,3-glucanase and resistance to Aspergillus flavus infection in maize. Crop Sci. 1998, 38, 1255–1260. [Google Scholar] [CrossRef]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-transferase enzymes in plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1836. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual, 1st ed.; Blackwell Publishing: Ames, IA, USA, 2006. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Anderson, J.B.; Stasovski, E. Molecular phylogeny of northern hemisphere species of Armillaria. Mycologia 1992, 84, 505–516. [Google Scholar] [CrossRef]
- Appel, D.J.; Gordon, T.R. Relationships among pathogenic and nonpathogenic isolates of Fusarium oxysporum based on the partial sequence of the intergenic spacer region of the ribosomal DNA. MPMI 1996, 9, 125–138. [Google Scholar] [CrossRef]
- Cabral, C.S.; Brunelli, K.R.; Costa, H.; Fonseca, M.E.D.N.; Boiteux, L.S.; Reis, A. Identification of Fusarium oxysporum f. sp. lactucae race 1 as the causal agent of lettuce wilt in Brazil. Trop. Plant Pathol. 2014, 39, 197–202. [Google Scholar] [CrossRef]
- Cabral, C.S.; de Fonseca, N.M.E.; Brunelli, K.R.; Rossato, M.; Costa, H.; Boiteux, L.S.; Reis, A. Relationships among Brazilian and worldwide isolates of Fusarium oxysporum f. sp. lactucae race 1 inferred from ribosomal intergenic spacer (IGS-rDNA) region and EF-1α gene sequences. Eur. J. Plant Pathol. 2018, 152, 81–94. [Google Scholar] [CrossRef]
- O’Donnell, K.; Gueidan, C.; Sink, S.; Johnston, P.R.; Crous, P.W.; Glenn, A.; Riley, R.; Zitomer, N.C.; Colyer, P.; Waalwijk, C.; et al. A two-locus DNA sequence database for typing plant and human pathogens within the Fusarium oxysporum species complex. Fungal Genet. Biol. 2009, 46, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xiao, M.; Kong, F.; Chen, S.; Dou, H.-T.; Sorrell, T.; Li, R.-Y.; Xu, Y.-C. Accurate and practical identification of 20 Fusarium species by seven-locus sequence analysis and reverse line blot hybridization, and an in vitro antifungal susceptibility study. J. Clin. Microbiol. 2011, 49, 1890–1898. [Google Scholar] [CrossRef] [PubMed]
- Gilardi, G.; Demarchi, S.; Gullino, M.L.; Garibaldi, A. Varietal resistance to control fusarium wilts of leafy vegetables under greenhouse. Commun. Agric. Appl. Biol. 2014, 79, 21–27. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression using real-time quantitative PCR. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, M.M.; Martinez, M.C.; León, M.; Armengol, J.; Monserrat, A. First report of Fusarium wilt of lettuce caused by Fusarium oxysporum f. sp. lactucae race 1 in Spain. Plant Dis. 2020, 104, 1858. [Google Scholar] [CrossRef]
- Herrero, M.L.; Nagy, N.E.; Solheim, H. First report of Fusarium oxysporum f. sp. lactucae race 1 causing Fusarium wilt of lettuce in Norway. Plant Dis. 2021, 105, 2239. [Google Scholar] [CrossRef]
- Scott, J.C.; Kirkpatrick, S.C.; Gordon, T.R. Variation in susceptibility of lettuce cultivars to Fusarium wilt caused by Fusarium oxysporum f. sp. lactucae. Plant Pathol. 2010, 59, 139–146. [Google Scholar] [CrossRef]
- Gordon, T.R.; Koike, S.T. Management of fusarium wilt in lettuce. J. Crop Prot. 2015, 73, 45–49. [Google Scholar] [CrossRef]
- Llorens, A.; Hinojo, M.J.; Mateo, R.; Medina, A.; Valle-Algarra, F.M.; Gonzalez-Jaen, M.T.; Jimenez, M. Variability and characterization of mycotoxin-producing Fusarium spp. isolates by PCR-RFLP analysis of the IGS-rDNA region. Antonie Leeuwenhoek 2006, 89, 465–478. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Gard, B.; Gilardi, G. IPM for protecting leafy vegetables under greenhouses. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Springer International Publishing: New York, NY, USA, 2020; Volume 9, pp. 567–596. [Google Scholar] [CrossRef]
- Gilardi, G.; Gullino, M.L.; Garibaldi, A. Emerging foliar and soil-borne pathogens of leafy vegetable crops: A possible threat to Europe. EPPO Bull. 2018, 481, 116–127. [Google Scholar] [CrossRef]
- Scott, J.C.; McRoberts, D.N.; Gordon, T.R. Colonization of lettuce cultivars and rotation crops by Fusarium oxysporum f. sp. lactucae, the cause of fusarium wilt of lettuce. Plant Pathol. 2014, 63, 548–553. [Google Scholar] [CrossRef]
- Matheron, M.E.; McCreight, J.D.; Tickes, B.R.; Porchas, M. Effect of planting date, cultivar, and stage of plant development on incidence of Fusarium wilt of lettuce in desert production fields. Plant Dis. 2005, 89, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Gilardi, G.; Vasileiadou, A.; Garibaldi, A.; Gullino, M.L. Low temperatures favour Fusarium wilt development by race 4 of Fusarium oxysporum f. sp. lactucae. J. Plant Pathol. J. 2021, 103, 973–979. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Name | Species Name | Geographical Origin | GenBank Accession Numbers x | Ref. | |
|---|---|---|---|---|---|
| TEF1-α | IGS | ||||
| GTFus1 | Fusarium oxysporum f. sp. lactucae race 1 | Greece | OQ466113 | OQ507251 | * |
| GTFus2 | F. oxysporum f. sp. lactucae race 1 | Greece | OQ466114 | OQ507252 | |
| GTFus3 | F. oxysporum f. sp. lactucae race 1 | Greece | OQ466115 | OQ507253 | |
| GTFus4 | F. oxysporum f. sp. lactucae race 1 | Greece | OQ466116 | OQ507254 | |
| Fuslat 1.14 | F. oxysporum f. sp. lactucae race 1 | Italy | KU840921.1 | KU840914.1 | [9] |
| Fuslat 3.14 | F. oxysporum f. sp. lactucae race 1 | Italy | KU840820.1 | KU840913.1 | |
| GL1815 | F. oxysporum f. sp. lactucae race 1 | California, USA | MH412701.1 | MH412706.1 | [20] |
| GL1639 | F. oxysporum f. sp. lactucae race 1 | California, USA | MH412702.1 | MH412707.1 | |
| JCP024 | F. oxysporum f. sp. lactucae race 1 | California, USA | MH412703.1 | MH412708.1 | |
| JCP164 | F. oxysporum f. sp. lactucae race 1 | Arizona, USA | MH412704.1 | MH412709.1 | |
| BMP1300 | F. oxysporum f. sp. lactucae race 1 | Arizona, USA | DQ837658.1 | DQ831864.1 | [16] |
| BMP1301 | F. oxysporum f. sp. lactucae race 1 | Arizona, USA | DQ837659.1 | DQ831865.1 | |
| BMP1306 | F. oxysporum f. sp. lactucae race 1 | Arizona, USA | DQ837660.1 | DQ831866.1 | |
| Fus-171 | F. oxysporum f. sp. lactucae race 1 | Brazil | KY561356.1 | KY352887.1 | [54] |
| Fus-172 | F. oxysporum f. sp. lactucae race 1 | Brazil | KY561357.1 | KY352888.1 | |
| S1 | F. oxysporum f. sp. lactucae race 1 | Japan | DQ837657.1 | DQ831863.1 | [16] |
| F9501 | F. oxysporum f. sp. lactucae race 2 | Japan | DQ837693.1 | DQ831893.1 | |
| FK09701 | F. oxysporum f. sp. lactucae race 2 | Japan | DQ837694.1 | DQ831892.1 | |
| MAFF744085 | F. oxysporum f. sp. lactucae race 3 | Japan | KU840924.1 | KU840917.1 | [9] |
| MAFF744086 | F. oxysporum f. sp. lactucae race 3 | Japan | KU840925.1 | KU840918.1 | |
| L. sativa 04750888 | F. oxysporum f. sp. lactucae race 4 | Netherlands | KU840922.1 | KU840915.1 | |
| L. sativa 04750896 | F. oxysporum f. sp. lactucae race 4 | Netherlands | KU840923.1 | KU840916.1 | |
| FOV14 | F. oxysporum f. sp. vasinfectum | California, USA | DQ837695.1 | DQ831885.1 | [16] |
| FOLR2 | F. oxysporum f. sp. lycopersici | California, USA | DQ837692.1 | DQ831894.1 | |
| TX388 | F. oxysporum f. sp. melonis | Texas, USA | DQ837696.1 | DQ83188.1 | |
| FOA50 | F. oxysporum f. sp. asparagi | Australia | DQ837691.1 | DQ831886.1 | |
| NRRL22546 | F. oxysporum f. sp. medicaginis | SE Asia | DQ837690.1 | DQ831901.1 | |
| NRRL26871 | F. oxysporum f. sp. spinaciae | Japan | DQ837687.1 | DQ831888.1 | |
| NRRL53158 | F. oxysporum f. sp. conglutinans | North Carolina, USA | FJ985443.1 | FJ985678.1 | [55] |
| NRRL26222 | F. oxysporum f. sp. dianthi | Israel | FJ985284.1 | FJ985490.1 | |
| NRRL36570 | F. oxysporum f. sp. radicis-lycopersici | Unknown | FJ985358.1 | FJ985591.1 | |
| NRRL36107 | F. oxysporum f. sp. cubense | Honduras | FJ985326.1 | FJ985559.1 | |
| NRRL22553 | F. oxysporum f. sp. raphani | Germany | FJ985273.1 | FJ985463.1 | |
| PUF016 | F. subglutinans | China | HQ165847.1 | HQ165883.1 | [56] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tziros, G.T.; Karaoglanidis, G.S. Identification of Fusarium oxysporum f. sp. lactucae Race 1 as the Causal Agent of Lettuce Fusarium Wilt in Greece, Commercial Cultivars’ Susceptibility, and Temporal Expression of Defense-Related Genes. Microorganisms 2023, 11, 1082. https://doi.org/10.3390/microorganisms11041082
Tziros GT, Karaoglanidis GS. Identification of Fusarium oxysporum f. sp. lactucae Race 1 as the Causal Agent of Lettuce Fusarium Wilt in Greece, Commercial Cultivars’ Susceptibility, and Temporal Expression of Defense-Related Genes. Microorganisms. 2023; 11(4):1082. https://doi.org/10.3390/microorganisms11041082
Chicago/Turabian StyleTziros, George T., and George S. Karaoglanidis. 2023. "Identification of Fusarium oxysporum f. sp. lactucae Race 1 as the Causal Agent of Lettuce Fusarium Wilt in Greece, Commercial Cultivars’ Susceptibility, and Temporal Expression of Defense-Related Genes" Microorganisms 11, no. 4: 1082. https://doi.org/10.3390/microorganisms11041082