
Tracking the Source of Human Q Fever from a Southern French Village: Sentinel Animals and Environmental Reservoir
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Case Reports
2.2. Animal Sampling
2.3. Serological Analysis
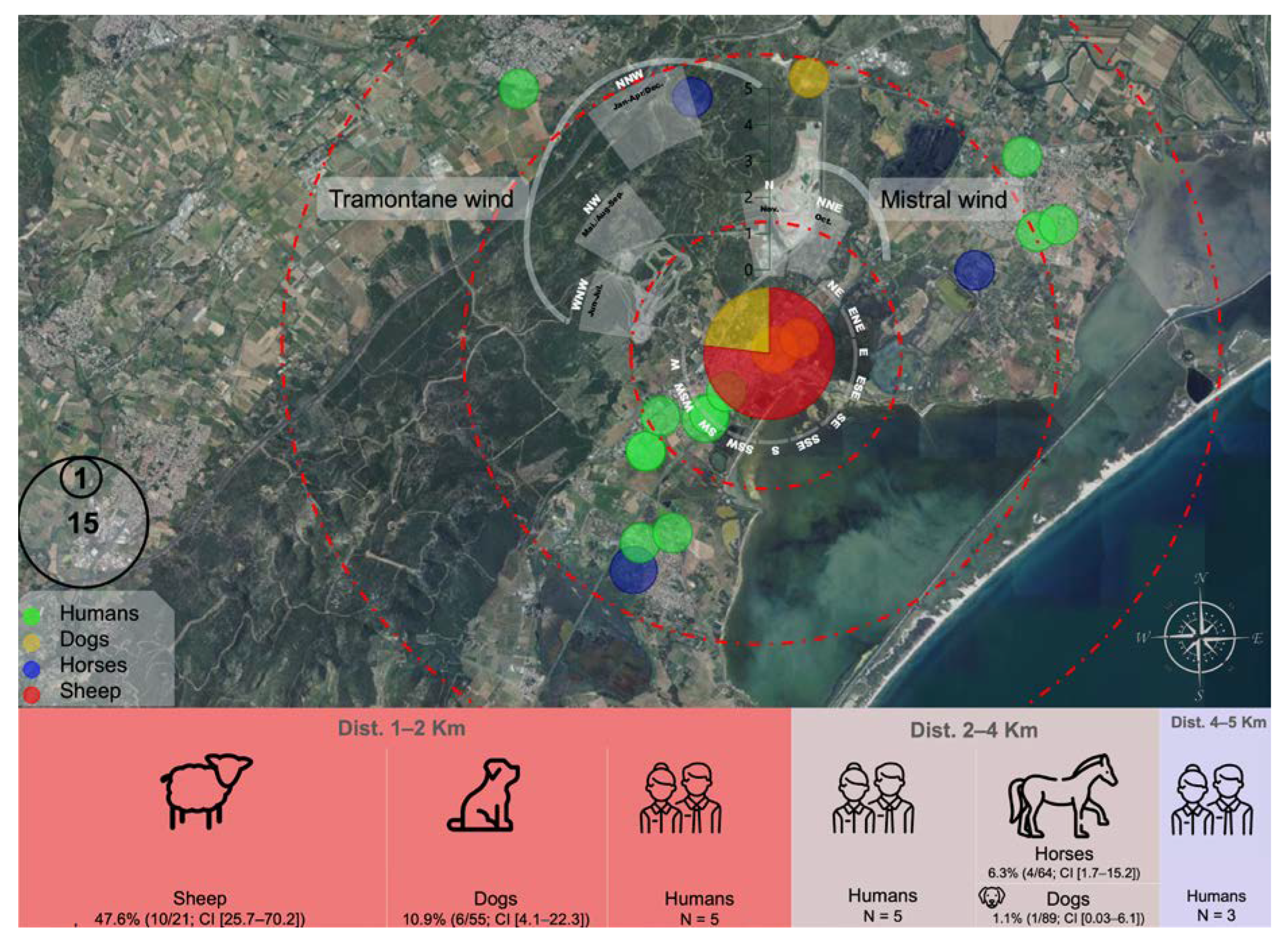
2.4. Geographical Plotting of Q Fever Cases
2.5. Molecular Analysis
2.5.1. Real-Time PCR Detection and Quantification of Bacterial Load
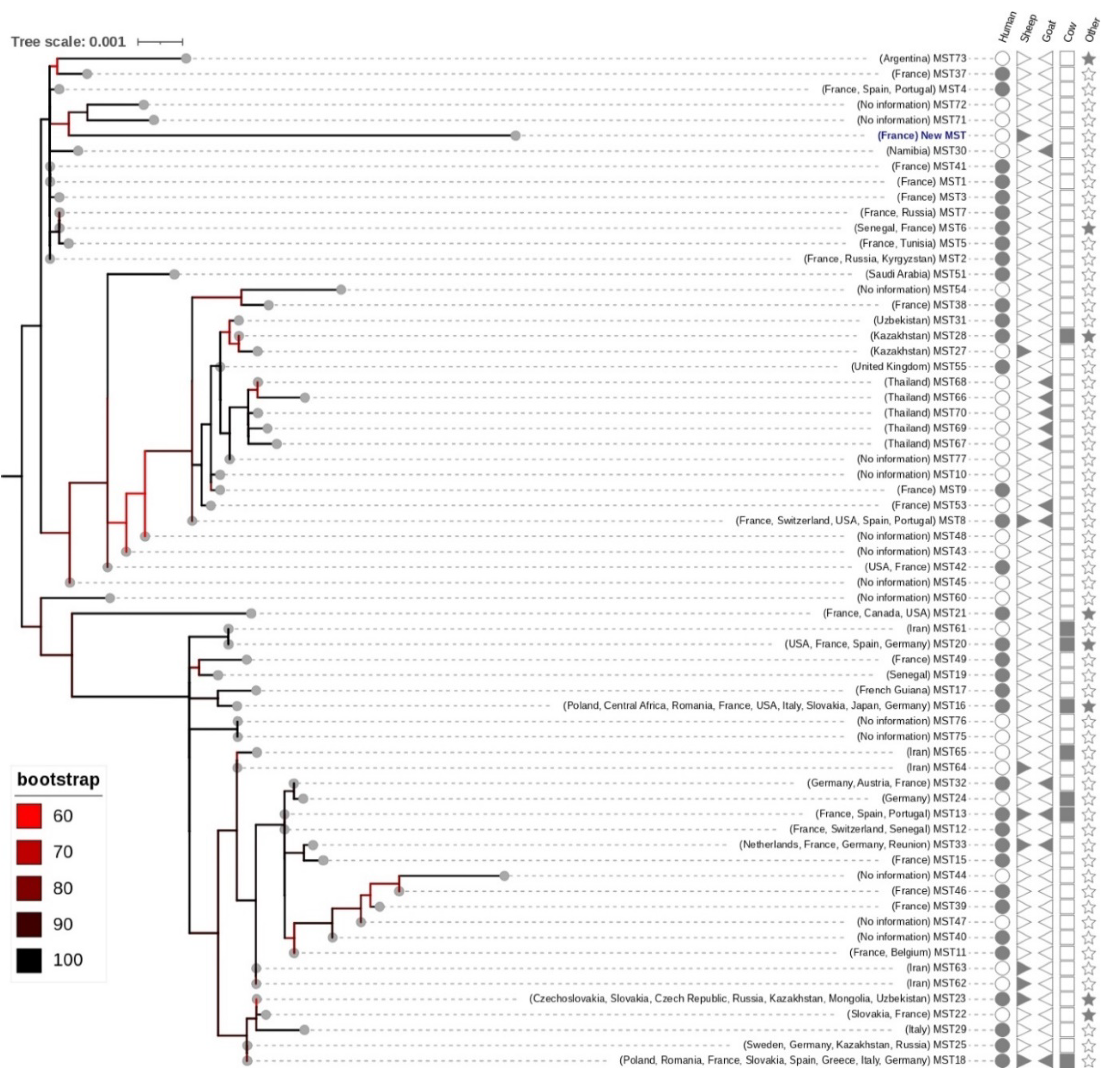
2.5.2. Molecular Typing and Phylogenetic Analysis
MinION Library Preparation and Multiplexed Nanopore Sequencing
Data Analysis and Multi-Loci Sequence Typing
3. Results
3.1. Human Q Fever Cases
3.2. Serological Screening of Q Fever from Animals
3.3. Real-Time PCR Detection and Bacterial Quantification
3.4. Mapping Human and Animal Q Fever Cases
3.5. C. burnetii Genotyping
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Approval and Consent to Participate
References
- Eldin, C.; Mélenotte, C.; Mediannikov, O.; Ghigo, E.; Million, M.; Edouard, S.; Mege, J.L.; Maurin, M.; Raoult, D. From Q fever to Coxiella burnetii infection: A paradigm change. Clin. Microbiol. Rev. 2017, 30, 115–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raoult, D.; Etienne, J.; Massip, P.; Iaocono, S.; Prince, M.A.; Beaurain, P.; Benichou, S.; Auvergnat, J.C.; Mathieu, P.; Bachet, P.H.; et al. Q fever endocarditis in the South of France. J. Infect. Dis. 1987, 155, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Rodolakis, A. Q fever in dairy animals. Ann. N. Y. Acad. Sci. 2009, 1166, 90–93. [Google Scholar] [CrossRef] [PubMed]
- World organization for animal health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals: Mammals, Birds and Bees; WOAH, 2022. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 13 March 2023).
- Frankel, D.; Richet, H.; Renvoisé, A.; Raoult, D. Q fever in France, 1985-2009. Emerg. Infect. Dis. 2011, 17, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Rahaman, M.R.; Milazzo, A.; Marshall, H.; Bi, P. Is a One Health approach utilized for Q fever control? A comprehensive literature review. Int. J. Environ. Res. Public Health 2019, 16, 730. [Google Scholar] [CrossRef] [Green Version]
- Plummer, P.J.; McClure, J.T.; Menzies, P.; Morley, P.S.; van den Brom, R.; van Metre, D.C. Management of Coxiella burnetii infection in livestock populations and the associated zoonotic risk: A consensus statement. J. Vet. Intern. Med. 2018, 32, 1481–1494. [Google Scholar] [CrossRef] [Green Version]
- De Rooij, M.M.T.; Van Leuken, J.P.G.; Swart, A.; Kretzschmar, M.E.E.; Nielen, M.; De Koeijer, A.A.; Janse, I.; Wouters, I.M.; Heederik, D.J.J. A systematic knowledge synthesis on the spatial dimensions of Q fever epidemics. Zoonoses Public Health 2019, 66, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Maurin, M.; Raoult, D. Q fever. Clin. Microbiol. Rev. 1999, 12, 518–553. [Google Scholar] [CrossRef] [Green Version]
- Eldin, C.; Angelakis, E.; Renvoisé, A.; Raoult, D. Coxiella burnetii DNA, but not viable bacteria, in dairy products in France. Am. J. Trop. Med. Hyg. 2013, 88, 765–769. [Google Scholar] [CrossRef] [Green Version]
- Miller, H.K.; Priestley, R.A.; Kersh, G.J. Comparison of three Coxiella burnetii infectious routes in mice. Virulence 2021, 12, 2562–2570. [Google Scholar] [CrossRef]
- Wittwer, M.; Hammer, P.; Runge, M.; Valentin-Weigand, P.; Neubauer, H.; Henning, K.; Mertens-Scholz, K. Inactivation kinetics of Coxiella burnetii during high-temperature short-time pasteurization of milk. Front. Microbiol. 2022, 12, 753871. [Google Scholar] [CrossRef] [PubMed]
- Noah, D.L.; Noah, D.L.; Crowder, H.R. Biological terrorism against animals and humans: A brief review and primer for action. J. Am. Vet. Med. Assoc. 2002, 221, 40–43. [Google Scholar] [CrossRef] [Green Version]
- Whitney, E.A.S.; Massung, R.F.; Candee, A.J.; Ailes, E.C.; Myers, L.M.; Patterson, N.E.; Berkelman, R.L. Seroepidemiologic and occupational risk survey for Coxiella burnetii antibodies among US veterinarians. Clin. Infect. Dis. 2009, 48, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne Diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Gache, K.; Rousset, E.; Perrin, J.B.; de Cremoux, R.; Hosteing, S.; Jourdain, E.; Guatteo, R.; Nicollet, P.; Touratier, A.; Calavas, D.; et al. Estimation of the frequency of Q fever in sheep, goat and cattle herds in France: Results of a 3-year study of the seroprevalence of Q fever and excretion level of Coxiella burnetii in abortive episodes. Epidemiol. Infect. 2017, 145, 3131–3142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrié, P.; Barry, S.; Rousset, E.; de Crémoux, R.; Sala, C.; Calavas, D.; Perrin, J.B.; Bronner, A.; Gasqui, P.; Gilot-Fromont, E.; et al. Swab Cloths as a tool for revealing environmental contamination by Q fever in ruminant farms. Transbound. Emerg. Dis. 2019, 66, 1202–1209. [Google Scholar] [CrossRef]
- Lurier, T.; Rousset, E.; Gasqui, P.; Sala, C.; Claustre, C.; Abrial, D.; Dufour, P.; de Crémoux, R.; Gache, K.; Delignette-Muller, M.L.; et al. Evaluation using latent class models of the diagnostic performances of three ELISA tests commercialized for the serological diagnosis of Coxiella burnetii Infection in Domestic Ruminants. Vet Res 2021, 52, 56. [Google Scholar] [CrossRef]
- Rousset, E.; Durand, B.; Champion, J.-L.; Prigent, M.; Dufour, P.; Forfait, C.; Marois, M.; Gasnier, T.; Duquesne, V.; Thiéry, R. Efficiency of a Phase 1 Vaccine for the reduction of vaginal Coxiella burnetii shedding in a clinically affected goat herd. Clin. Microbiol. Infect. 2009, 15, 188–189. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, A.; Russo, M. Introducing Microsoft Power BI; Microsoft Press: Unterschleissheim, Germany, 2016; ISBN 1509302751. [Google Scholar]
- Box, A. Introduction to Charticulator. In Introducing Charticulator for Power BI; Springer: Berlin/Heidelberg, Germany, 2022; pp. 1–30. [Google Scholar]
- Klee, S.R.; Tyczka, J.; Ellerbrok, H.; Franz, T.; Linke, S.; Baljer, G.; Appel, B. Highly sensitive Real-Time PCR for specific detection and quantification of Coxiella burnetii. BMC Microbiol. 2006, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astobiza, I.; Tilburg, J.J.H.C.; Piñero, A.; Hurtado, A.; García-Pérez, A.L.; Nabuurs-Franssen, M.H.; Klaassen, C.H.W. Genotyping of Coxiella burnetii from domestic ruminants in Northern Spain. BMC Vet. Res. 2012, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Liou, C.-H.; Wu, H.-C.; Liao, Y.-C.; Lauderdale, T.-L.Y.; Huang, I.-W.; Chen, F.-J. NanoMLST: Accurate multilocus sequence typing using Oxford Nanopore technologies MinION with a dual-barcode approach to multiplex large numbers of samples. Microb Genom. 2020, 6, e000336. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baloğlu, B.; Chen, Z.; Elbrecht, V.; Braukmann, T.; MacDonald, S.; Steinke, D. A Workflow for accurate metabarcoding using Nanopore MinION sequencing. Methods Ecol. Evol. 2021, 12, 794–804. [Google Scholar] [CrossRef]
- Srivathsan, A.; Baloğlu, B.; Wang, W.; Tan, W.X.; Bertrand, D.; Ng, A.H.Q.; Boey, E.J.H.; Koh, J.J.Y.; Nagarajan, N.; Meier, R. A Min IONTM-based pipeline for fast and cost-effective DNA barcoding. Mol. Ecol. Resour. 2018, 18, 1035–1049. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2018, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. Sea view version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 274–568. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (ITOL) v5: An online tool for phylogenetic tree displays and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Rousset, E.; Raptopoulo, A.; Poivre, M.; Lafon, J.; Thiéry, R.; Couesnon, A.; Jourdain, E.; Gache, K. Epidemiological investigation on a dairy sheep farm in a professional agricultural high school following an alert of Q fever clustered human cases. In Proceedings of the ESCCAR International congress on Rickettsiae and 9th Meeting of the European Society for Chlamydia Research (ESCR), Lausanne, Switzerland, 23–26 August 2022; p. 171. [Google Scholar]
- Georgiev, M.; Afonso, A.; Neubauer, H.; Needham, H.; Thiéry, R.; Rodolakis, A.; Roest, H.J.; Stärk, K.D.; Stegeman, J.A.; Vellema, P. Q fever in humans and farm animals in four European countries, 1982 to 2010. Eurosurveillance 2013, 18, 20407. [Google Scholar] [CrossRef] [PubMed]
- de Cremoux, R.; Rousset, E.; Touratier, A.; Audusseau, G.; Nicollet, P.; Ribaud, D.; David, V.; le Pape, M. Coxiella burnetii vaginal shedding and antibody responses in dairy goat herds in a context of clinical Q fever outbreaks. FEMS Immunol. Med. Microbiol. 2012, 64, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Pérez, B.; Almería, S.; Tutusaus, J.; Jado, I.; Anda, P.; Monleón, E.; Badiola, J.; Garcia-Ispierto, I.; López-Gatius, F. Coxiella burnetii total immunoglobulin G, phase I and phase II immunoglobulin G antibodies, and bacterial shedding in young dams in persistently infected dairy herds. J. Vet. Diagn. Invest. 2015, 27, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousset, E.; Berri, M.; Durand, B.; Dufour, P.; Prigent, M.; Delcroix, T.; Touratier, A.; Rodolakis, A. Coxiella burnetii shedding routes and antibody response after outbreaks of Q fever-Induced abortion in dairy goat herds. Appl. Environ. Microbiol. 2009, 75, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Guatteo, R.; Beaudeau, F.; Joly, A.; Seegers, H. Coxiella burnetii shedding by dairy cows. Vet. Res. 2007, 38, 849–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, B.U.; Schoneberg, C.; Herms, T.L.; Kleinschmidt, S.; Runge, M.; Ganter, M. Impact of Coxiella burnetii vaccination on humoral immune response, vaginal shedding, and lamb mortality in naturally pre-infected sheep. Front. Vet. Sci. 2022, 9, 1912. [Google Scholar] [CrossRef]
- Bauer, B.U.; Schoneberg, C.; Herms, T.L.; Runge, M.; Ganter, M. Surveillance of Coxiella burnetii shedding in three naturally infected dairy goat herds after vaccination, focusing on bulk tank milk and dust swabs. Vet. Sci. 2022, 9, 102. [Google Scholar] [CrossRef]
- Joulié, A.; Laroucau, K.; Bailly, X.; Prigent, M.; Gasqui, P.; Lepetitcolin, E.; Blanchard, B.; Rousset, E.; Sidi-Boumedine, K.; Jourdain, E. Circulation of Coxiella burnetii in a naturally infected flock of dairy sheep: Shedding dynamics, environmental contamination, and genotype diversity. Appl. Environ. Microbiol. 2015, 81, 7253–7260. [Google Scholar] [CrossRef] [Green Version]
- Marmion, B.P.; Sukocheva, O.; Storm, P.A.; Lockhart, M.; Turra, M.; Kok, T.; Ayres, J.; Routledge, H.; Graves, S. Q fever: Persistence of antigenic non-viable cell residues of Coxiella burnetii in the host—Implications for post Q fever infection fatigue syndrome and other chronic sequelae. QJM 2009, 102, 673–684. [Google Scholar] [CrossRef]
- Bauer, B.; Prüfer, L.; Walter, M.; Ganter, I.; Frangoulidis, D.; Runge, M.; Ganter, M. Comparison of Coxiella burnetii excretion between sheep and goats naturally infected with one cattle-associated genotype. Pathogens 2020, 9, 652. [Google Scholar] [CrossRef]
- Álvarez-Alonso, R.; Zendoia, I.I.; Barandika, J.F.; Jado, I.; Hurtado, A.; López, C.M.; García-Pérez, A.L. Monitoring Coxiella burnetii infection in naturally infected dairy sheep flocks throughout four lambing seasons and investigation of viable bacteria. Front. Vet. Sci. 2020, 7, 352. [Google Scholar] [CrossRef]
- Álvarez-Alonso, R.; Barandika, J.F.; Ruiz-Fons, F.; Ortega-Araiztegi, I.; Jado, I.; Hurtado, A.; García-Pérez, A.L. Stable levels of Coxiella burnetii prevalence in dairy sheep flocks but changes in genotype distribution after a 10-year period in Northern Spain. Acta Vet. Scand. 2018, 60, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davoust, B.; Marié, J.-L.; Tahir, D.; Dahmani, M.; Dufour, P.; Thiéry, R.; Rousset, E. Seroprevalence of Coxiella burnetii infection in dogs from southeastern France. Int. J. Infect. Dis. 2019, 79, 122. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, I.; Joulié, A.; Pradier, S.; Lecollinet, S.; Beck, C.; Vial, L.; Dufour, P.; Gasqui, P.; Legrand, L.; Edouard, S.; et al. Seroprevalence of horses to Coxiella burnetii in an Q fever endemic area. Vet. Microbiol. 2018, 215, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Van Leuken, J.P.G.; Swart, A.N.; Brandsma, J.; Terink, W.; Van de Kassteele, J.; Droogers, P. Human Q fever incidence is associated to spatiotemporal environmental conditions. One Health 2016, 2, 77–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, L.A.; Goirand, L.; Tissot-Dupont, H.; Giunta, B.; Giraud, C.; Colardelle, C.; Duquesne, V.; Rousset, E.; Aubert, M.; Thiéry, R.; et al. Outbreak of Q fever, Florac, southern France, spring 2007. Vector-Borne Zoonotic Dis. 2011, 11, 341–347. [Google Scholar] [CrossRef]
- Tissot-Dupont, H.; Amadei, M.A.; Nezri, M.; Raoult, D. Wind in November, Q fever in December. Emerg. Infect. Dis. 2004, 10, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Kersh, G.J.; Wolfe, T.M.; Fitzpatrick, K.A.; Candee, A.J.; Oliver, L.D.; Patterson, N.E.; Self, J.S.; Priestley, R.A.; Loftis, A.D.; Massung, R.F. Presence of Coxiella burnetii DNA in the environment of the United States, 2006 to 2008. Appl. Environ. Microbiol. 2010, 76, 4469–4475. [Google Scholar] [CrossRef] [Green Version]
- Babudieri, B.; Moscovici, C. Research on the behavior of Coxiella burnetii in relation to various physical and chemical agents. Rend. -Ist. Super. Sanita 1950, 13, 739–748. [Google Scholar]
- Welsh, H.H.; Lennette, E.H.; Abinanti, F.R.; Winn, J.F.; Kaplan, W. Q fever studies. XXI. The recovery of Coxiella burnetii from the soil and surface water of premises harboring infected sheep. Am. J. Hyg. 1959, 70, 14–20. [Google Scholar] [PubMed]
- Parker, R.R.; Sussman, O. Spontaneous infection of the brown dog tick, Rhipicephalus sanguineus with Coxiella burnetii. Public Health Reports 1949, 64, 1159–1160. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.R.; Bell, E.J.; Lackman, D.B. Experimental studies of Q fever in cattle. I. Observations on four heifers and two milk cows. Am. J. Hyg. 1948, 48, 191–206. [Google Scholar]
- McCaul, T.F.; Williams, J.C. Developmental cycle of Coxiella burnetii: Structure and morphogenesis of vegetative and sporogenic differentiations. J. Bacteriol. 1981, 147, 1063–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barandika, J.F.; Alvarez-Alonso, R.; Jado, I.; Hurtado, A.; García-Pérez, A.L. Viable Coxiella burnetii in hard cheeses made with unpasteurized milk. Int. J. Food Microbiol. 2019, 303, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Alonso, R.; Basterretxea, M.; Barandika, J.F.; Hurtado, A.; Idiazabal, J.; Jado, I.; Beraza, X.; Montes, M.; Liendo, P.; García-Pérez, A.L. A Q fever outbreak with a high rate of abortions at a dairy goat farm: Coxiella burnetii shedding, environmental contamination, and viability. Appl. Environ. Microbiol. 2018, 84, e01650-18. [Google Scholar] [CrossRef] [Green Version]
- Abeykoon, A.M.H.; Poon, M.; Firestone, S.M.; Stevenson, M.A.; Wiethoelter, A.K.; Vincent, G.A. Performance evaluation and validation of air samplers to detect aerosolized Coxiella burnetii. Microbiol. Spectr. 2022, 10, e00655-22. [Google Scholar] [CrossRef]
- Abeykoon, A.M.H.; Clark, N.J.; Soares Magalhaes, R.J.; Vincent, G.A.; Stevenson, M.A.; Firestone, S.M.; Wiethoelter, A.K. Coxiella burnetii in the environment: A systematic review and critical appraisal of sampling methods. Zoonoses Public Health 2021, 68, 165–181. [Google Scholar] [CrossRef]
- Brooke, R.J.; Kretzschmar, M.E.E.; Mutters, N.T.; Teunis, P.F. Human dose response relation for airborne exposure to Coxiella burnetii. BMC Infect. Dis. 2013, 13, 488. [Google Scholar] [CrossRef] [Green Version]
- Heppell, C.W.; Egan, J.R.; Hall, I. A Human time dose response model for Q fever. Epidemics 2017, 21, 30–38. [Google Scholar] [CrossRef]
- Nelson, M.; Salguero, F.J.; Hunter, L.; Atkins, T.P. A novel marmoset (Callithrix Jacchus) model of human inhalational Q fever. Front. Cell. Infect. Microbiol. 2021, 10, 621635. [Google Scholar] [CrossRef]
- Stein, A.; Louveau, C.; Lepidi, H.; Ricci, F.; Baylac, P.; Davoust, B.; Raoult, D. Q fever pneumonia: Virulence of Coxiella burnetii pathovars in a murine model of aerosol infection. Infect. Immun. 2005, 73, 2469–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Bruin, A.; van Alphen, P.T.W.; van der Plaats, R.Q.J.; ND de Heer, L.; Reusken, C.B.E.M.; van Rotterdam, B.J.; Janse, I. Molecular typing of Coxiella burnetii from animal and environmental matrices during Q fever epidemics in the Netherlands. BMC Vet. Res. 2012, 8, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joulié, A.; Sidi-Boumedine, K.; Bailly, X.; Gasqui, P.; Barry, S.; Jaffrelo, L.; Poncet, C.; Abrial, D.; Yang, E.; Leblond, A. Molecular epidemiology of Coxiella burnetii in French livestock reveals the existence of three main genotype clusters and suggests species-specific associations as well as regional stability. Infect. Genet. Evol. 2017, 48, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Fasemore, A.M.; Helbich, A.; Walter, M.C.; Dandekar, T.; Vergnaud, G.; Förstner, K.U.; Frangoulidis, D. CoxBase: An online platform for epidemiological surveillance, visualization, analysis, and typing of Coxiella burnetii genomic sequences. mSystems 2021, 6, e00403-21. [Google Scholar] [CrossRef]
- Frangoulidis, D.; Kahlhofer, C.; Said, A.S.; Osman, A.Y.; Chitimia-Dobler, L.; Shuaib, Y.A. High prevalence and new genotype of Coxiella burnetii in ticks infesting camels in Somalia. Pathogens 2021, 10, 741. [Google Scholar] [CrossRef]
- Miller, H.K.; Binder, A.M.; Peterson, A.; Theel, E.S.; Volpe, J.M.; Couturier, M.R.; Cherry, C.C.; Kersh, G.J. Trends in Q fever serologic testing by immunofluorescence from four large reference laboratories in the United States, 2012–2016. Sci. Rep. 2018, 8, 16670. [Google Scholar] [CrossRef] [Green Version]
- Tissot-Dupont, H.; Torres, S.; Nezri, M.; Raoult, D. Hyperendemic focus of Q fever related to sheep and wind. Am. J. Epidemiol. 1999, 150, 67–74. [Google Scholar] [CrossRef]
- Thill, P.; Eldin, C.; Dahuron, L.; Berlioz-Artaud, A.; Demar, M.; Nacher, M.; Beillard, E.; Djossou, F.; Epelboin, L. High endemicity of Q fever in French Guiana: A cross sectional study (2007–2017). PLoS Negl. Trop. Dis. 2022, 16, e0010349. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Date | Patient Id | Gender | Age | Distance from the Sheepfold (m) |
|---|---|---|---|---|
| June 2019 | 1 | M | 30–40 | 4220 |
| September 2019 | 2 | M | 60–70 | 4850 |
| November 2019 | 3 | F | 70–80 | 2710 |
| May 2020 | 4 | M | 50–60 | 2100 |
| June 2020 | 5 | M | 40–50 | 3920 |
| June 2020 | 6 | M | 40–50 | 4200 |
| July 2020 | 7 | F | 50–60 | 2100 |
| September 2020 | 8 | M | 60–70 | 1130 |
| December 2020 | 9 | F | 60–70 | 766 |
| March 2021 | 10 | M | 50–60 | 1210 |
| May 2021 | 11 | F | 60–70 | 1670 |
| December 2021 | 12 | F | 50–60 | 1210 |
| June 2022 | 13 | F | 50–60 | 3060 |
| Origin of Animals | Location (Commune) | Distance from the Sheepfold (m) | Sheep | Goats | Dogs | Horses | All Animals | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | Positive | % | No. | Positive | % | No. | Positive | % | No. | Positive | % | No. | Positive | % | |||
| Sheepfold | A | 0 | 21 | 10 | 47.6 | 3 | 3 | 100 | 24 | 13 | 54.1 | ||||||
| Kennel 1 | A | 80 | - | - | 6 | 4 | 66.6 | 6 | 4 | 66.6 | |||||||
| Owner 1 | A | 445 | - | - | 6 | 0 | 0 | 2 | 1 | 50 | 4 | 1 | 25 | 12 | 2 | 16.6 | |
| Kennel 2 | A | 548 | - | - | 4 | 0 | 0 | 4 | 0 | 0 | |||||||
| Owner 2 | A | 989 | - | - | 1 | 0 | 0 | 1 | 0 | 0 | |||||||
| Kennel 3 | A | 1140 | - | - | 34 | 4 | 11.7 | 34 | 4 | 11.7 | |||||||
| Owner 3 | B | 1210 | - | - | 2 | 1 | 50 | 2 | 1 | 50 | |||||||
| Owner 4 | A | 1350 | - | - | 1 | 0 | 0 | 1 | 0 | 0 | |||||||
| Owner 5 | A | 1440 | - | - | 1 | 1 | 100 | 1 | 1 | 100 | |||||||
| Owner 6 | A | 1590 | - | - | 1 | 0 | 0 | 1 | 0 | 0 | |||||||
| Stable 1 | C | 2960 | - | - | 1 | 0 | 0 | 4 | 0 | 0 | 14 | 1 | 7 | 19 | 1 | 5.2 | |
| Kennel 4 | B | 3240 | - | - | 10 | 0 | 0 | 10 | 0 | 0 | |||||||
| Stable 2 | B | 3400 | - | - | 10 | 3 | 30 | 10 | 3 | 30 | |||||||
| Stable 3 | D | 3500 | - | - | 40 | 2 | 5 | 40 | 2 | 5 | |||||||
| Kennel 5 | C | 3730 | - | - | 70 | 5 | 7 | 70 | 5 | 7 | |||||||
| Kennel 6 | B | 3760 | - | - | 5 | 0 | 0 | 5 | 0 | 0 | |||||||
| Owner 7 | B | 4250 | - | - | 3 | 0 | 0 | 3 | 0 | 0 | |||||||
| Owner 8 | B | 4820 | - | - | 2 | 0 | 0 | 2 | 0 | 0 | |||||||
| Stable 4 | B | 5080 | - | - | 1 | 0 | 0 | 22 | 1 | 4.5 | 23 | 1 | 4.43 | ||||
| Stable 5 | D | 6330 | - | - | 16 | 1 | 6.2 | 16 | 1 | 6.2 | |||||||
| TOTAL | 21 | 10 | 47.6 | 7 | 0 | 0 | 150 | 19 | 12.6 | 106 | 9 | 8.49 | 284 | 38 | 13.38 | ||
| Serological Analyses (PrioCHECK TM Q Fever) | Molecular Analysis Using IS1111 qPCR | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sheep Number | Gender | Age (Years) | First Screening: 7 April 2022 | Second Screening: 7 June 2022 | Nasal Swabs | Vaginal Swabs | Rectal Swabs | ||||||
| Interp. | S/P% | Interp. | S/P% | Ct Value | IS Copy Number/ Reaction | Genome Equivalent Copy Number/mL | Ct Value | IS Copy Number/ Reaction | Genome Equivalent Copy Number/mL | Ct Value | |||
| OVG1 | F | 3 | POS++ | 177 | POS++ | 176 | ND | ND | ND | ||||
| OVG2 | F | 3 | NEG | 26.2 | NEG | 19.4 | ND | ND | ND | ||||
| OVG3 | F | 7 | POS++ | 185.6 | POS++ | 188.9 | 26.6 | 75,600 | 756,000 | ND | ND | ||
| OVG4 | F | 3 | NEG | 6.2 | NEG | 6.6 | 31.3 | 3,090 | 31,000 | ND | ND | ||
| OVG5 | F | 8 | POS++ | 47 | NEG | 35.6 | 34.3 | 422 | 4200 | ND | ND | ||
| OVG6 | F | 9 | POS++ | 46.4 | NEG | 31.6 | ND | ND | ND | ||||
| OVG7 | F | 3 | POS++ | 113 | POS++ | 91.3 | ND | ND | ND | ||||
| OVG8 | F | 9 | NEG | 14.7 | NEG | 16 | ND | ND | ND | ||||
| OVG9 | F | 1 | POS+++ | 232.1 | POS+++ | 250.9 | ND | ND | ND | ||||
| OVG10 | F | 8 | NEG | 17.6 | NEG | 17.3 | ND | ND | ND | ||||
| OVG11 | F | 3 | NEG | 24.9 | |||||||||
| OVG12 | F | 3 | NEG | 10.8 | NEG | 5.6 | ND | 36.94 | 70.9 | 800 | ND | ||
| OVG13 | F | 3 | POS+++ | 205.1 | POS+++ | 175.8 | 34.3 | 407 | 4000 | ND | ND | ||
| OVG14 | F | 3 | NEG | 15 | |||||||||
| OVG15 | F | 3 | POS+ | 70.8 | POS+ | 50.8 | ND | ND | ND | ||||
| OVG16 | M | 8 | NEG | 9.5 | NEG | 9.1 | 29.4 | 11,100 | 111,000 | ND | |||
| OVG17 | F | 3 | POS++ | 190.4 | POS+++ | 288.5 | ND | ND | ND | ||||
| OVG18 | F | 6 | POS+ | 82.7 | POS+ | 99.2 | ND | ND | ND | ||||
| OVM19 | M | 3 months | NEG | 3.30 | ND | ND | |||||||
| OVM20 | M | NEG | 2.70 | ND | ND | ||||||||
| OVM21 | F | NEG | 3 | ND | ND | ND | |||||||
| Sample Type | Environmental Swabs in the Sheepfold | IS1111 qPCR | ||
|---|---|---|---|---|
| Ct Value | Genome Equivalent Copy Number /mL | Class per Sample | ||
| Swab | Manure | 35.57 | 170 | [<1000 bact.] |
| Cobweb | 26.89 | 61,320 | [10 × 104–10 × 105] | |
| Wooden board | 26.07 | 106,000 | [10 × 104–10 × 105] | |
| Concrete floor | 29.19 | 13,000 | [10 × 104–10 × 105] | |
| Cloth (SodiBox®) | Watering hole | 33.78 | 39,000 | [10 × 105–10 × 107] |
| Left feeder | 31.25 | 222,000 | [10 × 105–10 × 107] | |
| Right feeder | 30.02 | 521,000 | [10 × 105–10 × 107] | |
| Contamination Area | Q Fever Seroprevalence in Animal | Human Cases | ||
|---|---|---|---|---|
| Number of Animals | Number of Positive Cases | % | ||
| 2 km radius around the sheepfold | 86 | 25 | 29 | 5 |
| Between 2 and 4 km around the sheepfold | 154 | 11 | 7.14 | 5 |
| Between 4 and 6 km around the sheepfold | 44 | 2 | 4.54 | 3 |
| TOTAL | 284 | 38 | 13.38 | 13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laidoudi, Y.; Rousset, E.; Dessimoulie, A.-S.; Prigent, M.; Raptopoulo, A.; Huteau, Q.; Chabbert, E.; Navarro, C.; Fournier, P.-E.; Davoust, B. Tracking the Source of Human Q Fever from a Southern French Village: Sentinel Animals and Environmental Reservoir. Microorganisms 2023, 11, 1016. https://doi.org/10.3390/microorganisms11041016
Laidoudi Y, Rousset E, Dessimoulie A-S, Prigent M, Raptopoulo A, Huteau Q, Chabbert E, Navarro C, Fournier P-E, Davoust B. Tracking the Source of Human Q Fever from a Southern French Village: Sentinel Animals and Environmental Reservoir. Microorganisms. 2023; 11(4):1016. https://doi.org/10.3390/microorganisms11041016
Chicago/Turabian StyleLaidoudi, Younes, Elodie Rousset, Anne-Sophie Dessimoulie, Myriam Prigent, Alizée Raptopoulo, Quentin Huteau, Elisabeth Chabbert, Catherine Navarro, Pierre-Edouard Fournier, and Bernard Davoust. 2023. "Tracking the Source of Human Q Fever from a Southern French Village: Sentinel Animals and Environmental Reservoir" Microorganisms 11, no. 4: 1016. https://doi.org/10.3390/microorganisms11041016