

Knocking Down Gm16685 Decreases Liver Granuloma in Murine Schistosomiasis Japonica
 , and
, and 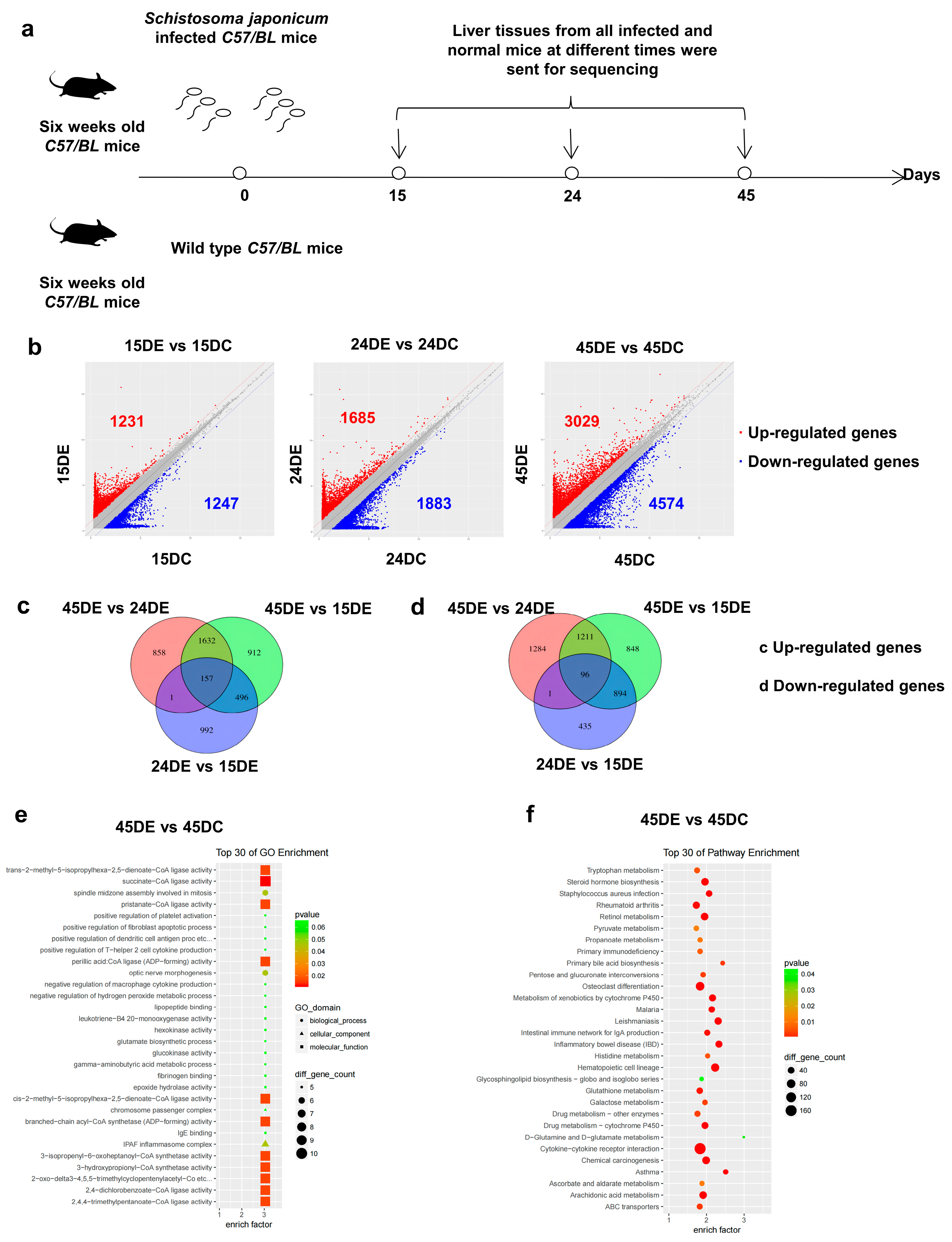{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Schistosomiasis Mouse Model
2.2. Cell Culture
2.3. Quantitative Reverse Transcription-PCR (qRT-PCR)
2.4. Cell Transfection
2.5. Histological Examination
2.6. Adult Counts and Egg Counts
2.7. Isolation of Cells
2.8. RNA Fluorescence in Situ Hybridization (FISH)
2.9. Nucleocytoplasmic Separation
2.10. Protein Determination Method
2.11. Luciferase Reporter Assay
2.12. miRDB
2.13. Statistical Analysis
3. Results
3.1. LncRNA Expression in the Liver of Mice with Schistosomiasis
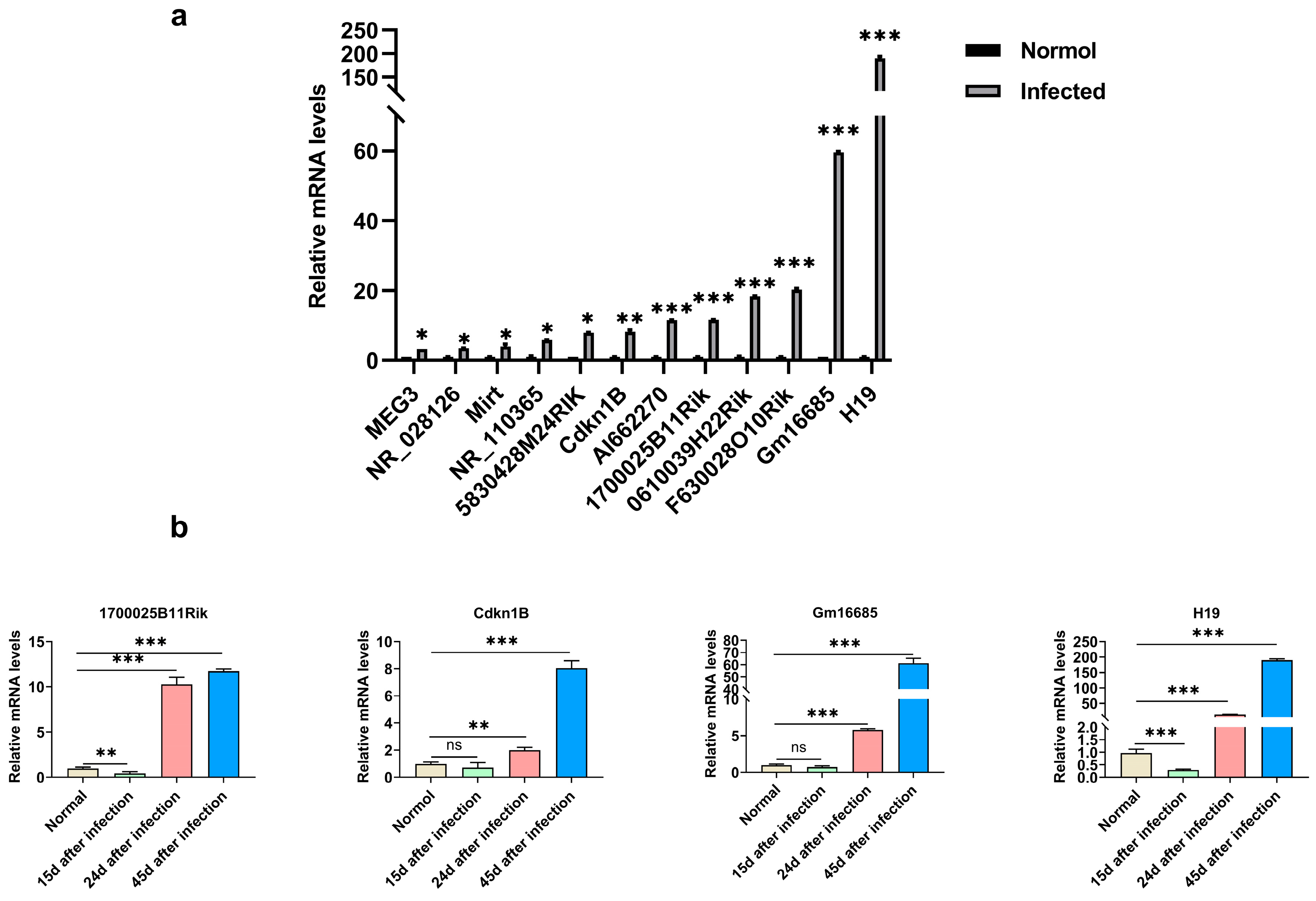
3.2. The Expression of lncRNA-Gm16685 Was Significantly Increased in the Livers of Mice with Schistosomiasis
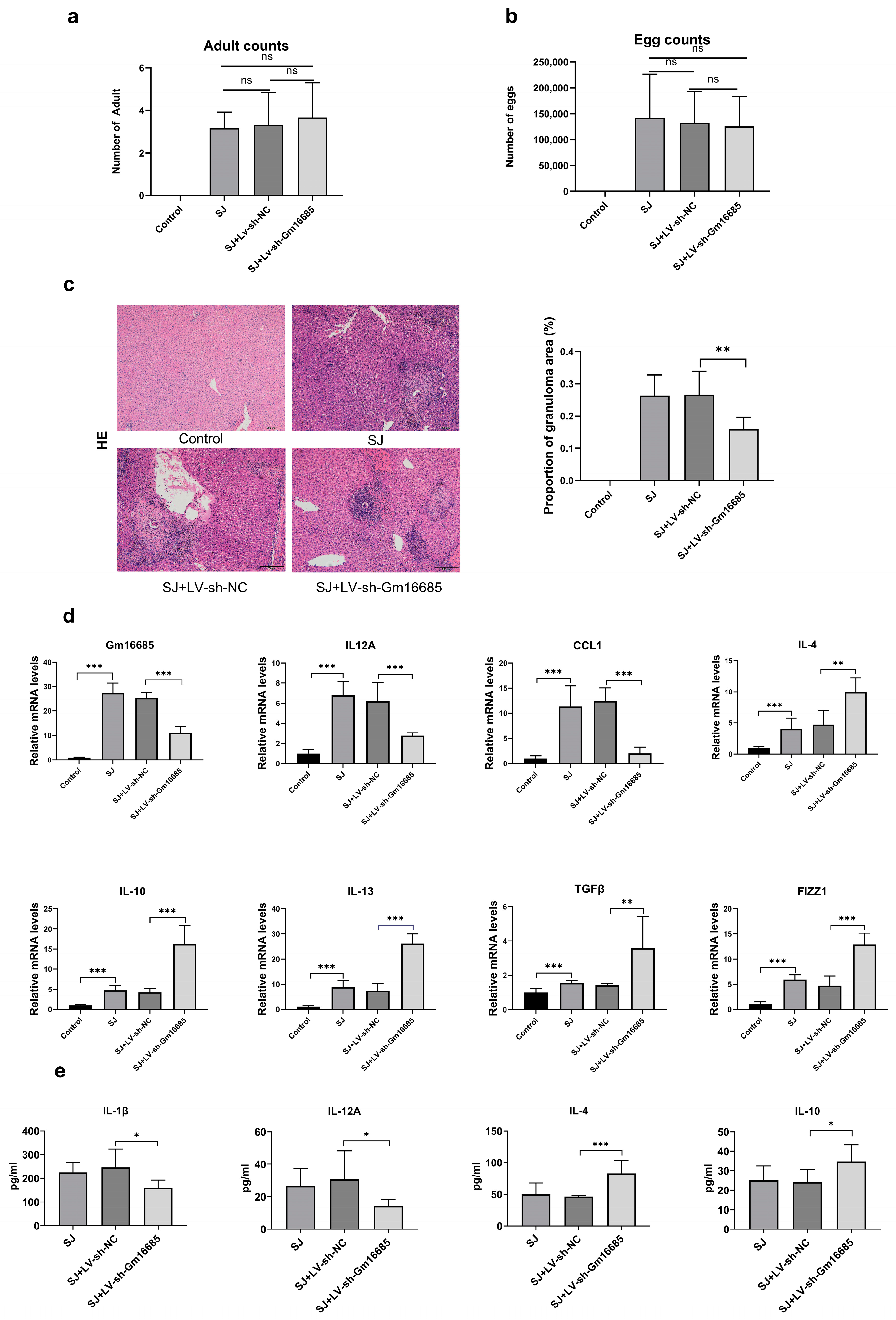
3.3. Knockdown of Gm16685 Alleviates S. japonicum-Induced Hepatic Granulomas
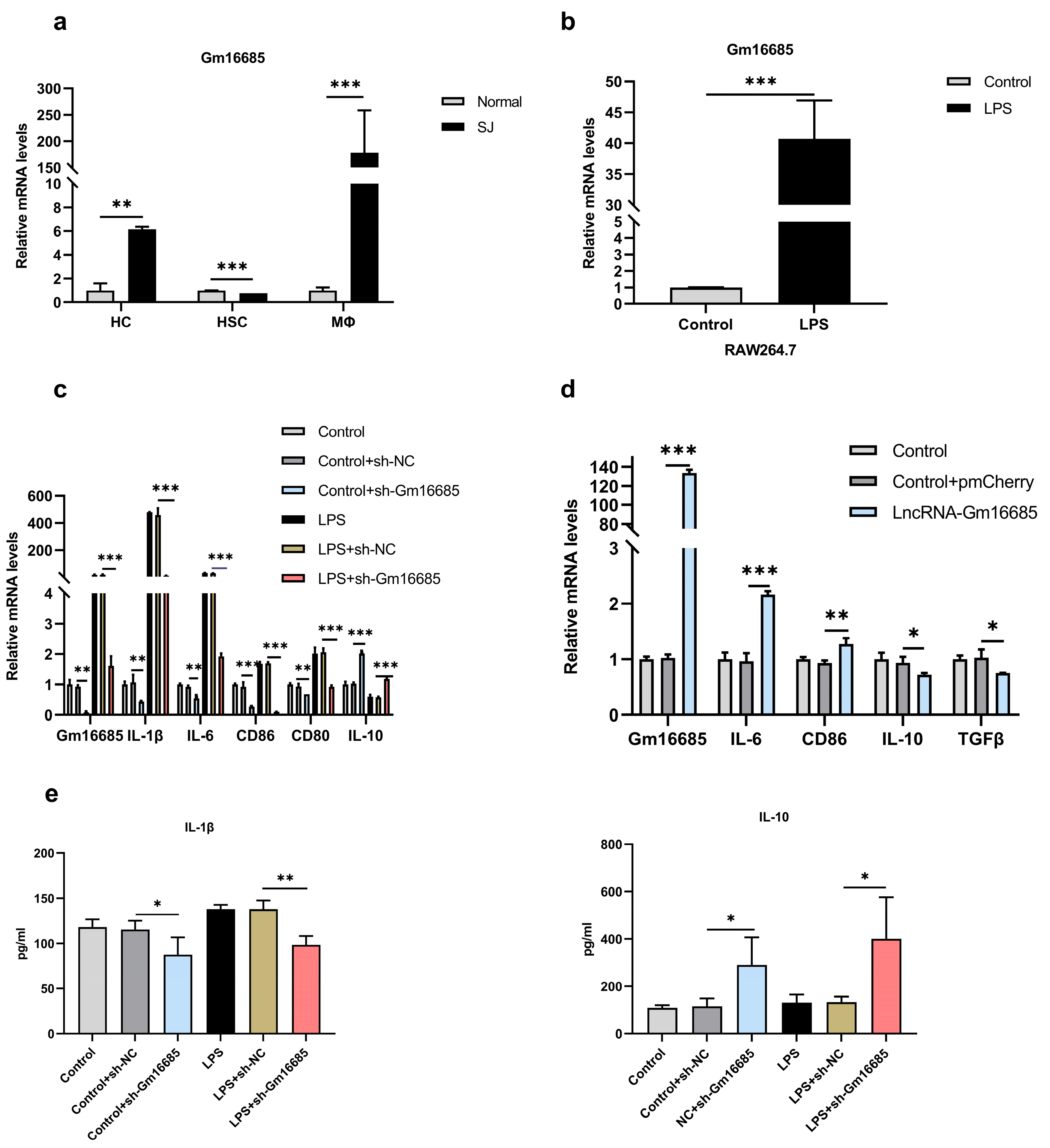
3.4. Knockdown of Gm16685 Promotes the Polarization of Macrophages to the M2 Phenotype In Vitro
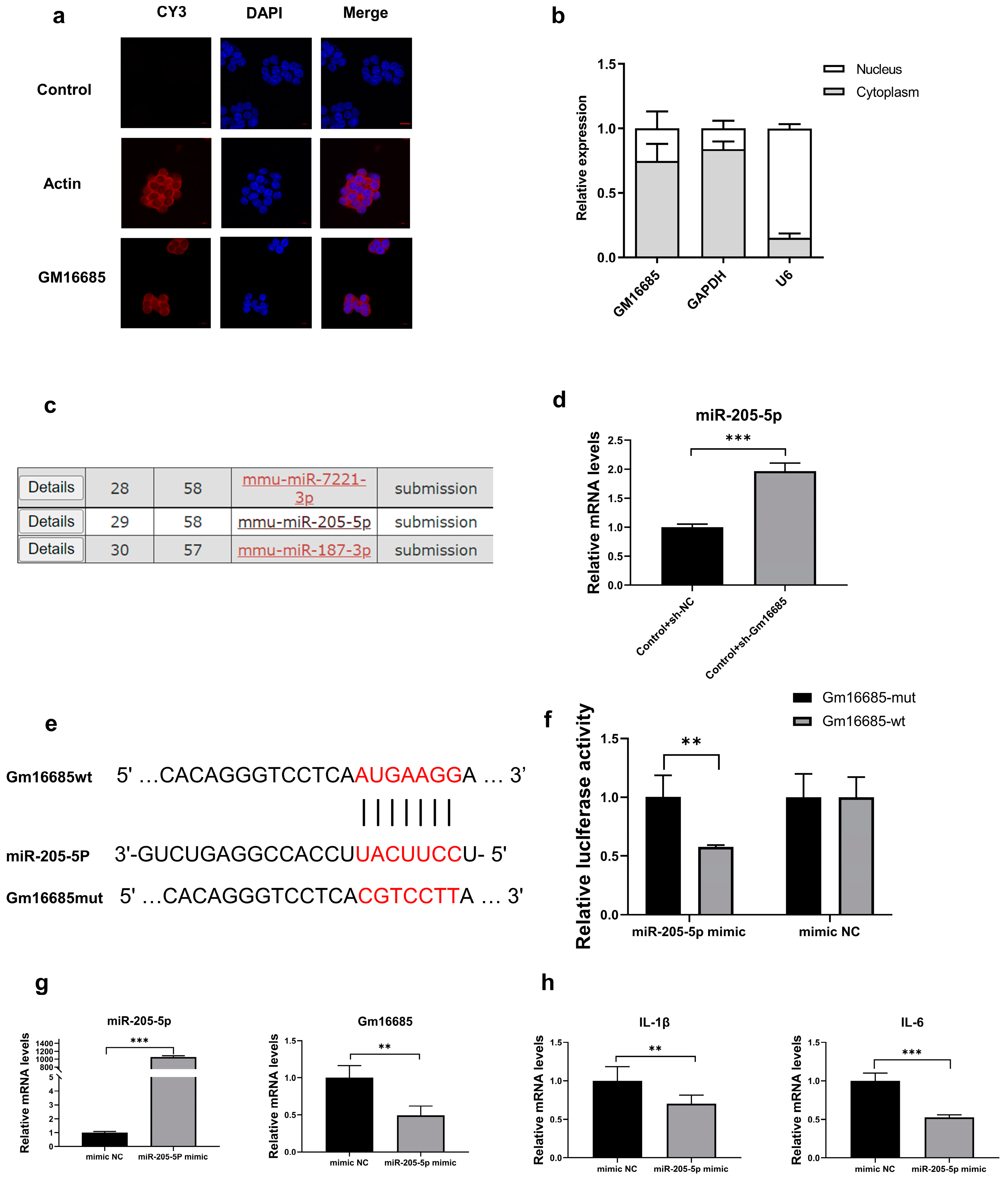
3.5. Further Study on the Mechanism of Action of Gm16685
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Boniatti, J.; Januskaite, P.; Fonseca, L.B.D.; Vicosa, A.L.; Amendoeira, F.C.; Tuleu, C.; Basit, A.W.; Goyanes, A.; Re, M.I. Direct Powder Extrusion 3D Printing of Praziquantel to Overcome Neglected Disease Formulation Challenges in Paediatric Populations. Pharmaceutics 2021, 13, 1114. [Google Scholar] [CrossRef]
- Cardoso, I.A.; de Souza, A.K.L.; Burgess, A.M.G.; Chalmers, I.W.; Hoffmann, K.F.; Nonato, M.C. Characterization of class II fumarase from Schistosoma mansoni provides the molecular basis for selective inhibition. Int. J. Biol. Macromol. 2021, 175, 406–421. [Google Scholar] [CrossRef]
- Abdel Aziz, N.; Musaigwa, F.; Mosala, P.; Berkiks, I.; Brombacher, F. Type 2 immunity: A two-edged sword in schistosomiasis immunopathology. Trends Immunol. 2022, 43, 657–673. [Google Scholar] [CrossRef]
- Liang, S.; Abe, E.M.; Zhou, X.N. Integrating ecological approaches to interrupt schistosomiasis transmission: Opportunities and challenges. Infect. Dis. Poverty 2018, 7, 124. [Google Scholar] [CrossRef]
- Lam, H.Y.P.; Liang, T.R.; Peng, S.Y. Ameliorative effects of Schisandrin B on Schistosoma mansoni-induced hepatic fibrosis in vivo. PLoS Negl. Trop. Dis. 2021, 15, e0009554. [Google Scholar] [CrossRef]
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef]
- Fung, H.S.; Siu, J.C.; Fan, T.W.; Lai, K.C.; Kwan, T.L. Hepatic schistosomiasis. Hong Kong Med. J. 2009, 15, 75–76. [Google Scholar]
- Pesce, J.T.; Ramalingam, T.R.; Mentink-Kane, M.M.; Wilson, M.S.; El Kasmi, K.C.; Smith, A.M.; Thompson, R.W.; Cheever, A.W.; Murray, P.J.; Wynn, T.A. Arginase-1-expressing macrophages suppress Th2 cytokine-driven inflammation and fibrosis. PLoS Pathog. 2009, 5, e1000371. [Google Scholar] [CrossRef] [Green Version]
- Vannella, K.M.; Barron, L.; Borthwick, L.A.; Kindrachuk, K.N.; Narasimhan, P.B.; Hart, K.M.; Thompson, R.W.; White, S.; Cheever, A.W.; Ramalingam, T.R.; et al. Incomplete deletion of IL-4Ralpha by LysM(Cre) reveals distinct subsets of M2 macrophages controlling inflammation and fibrosis in chronic schistosomiasis. PLoS Pathog. 2014, 10, e1004372. [Google Scholar] [CrossRef]
- Takaki, K.K.; Rinaldi, G.; Berriman, M.; Pagan, A.J.; Ramakrishnan, L. Schistosoma mansoni Eggs Modulate the Timing of Granuloma Formation to Promote Transmission. Cell Host Microbe 2021, 29, 58–67.e55. [Google Scholar] [CrossRef]
- Zhao, J.; Ling, L.; Zhu, W.; Ying, T.; Yu, T.; Sun, M.; Zhu, X.; Du, Y.; Zhang, L. M1/M2 re-polarization of kaempferol biomimetic NPs in anti-inflammatory therapy of atherosclerosis. J. Control Release 2022, 353, 1068–1083. [Google Scholar] [CrossRef]
- Rhee, I. Diverse macrophages polarization in tumor microenvironment. Arch. Pharm. Res. 2016, 39, 1588–1596. [Google Scholar] [CrossRef]
- Faure-Dupuy, S.; Delphin, M.; Aillot, L.; Dimier, L.; Lebosse, F.; Fresquet, J.; Parent, R.; Matter, M.S.; Rivoire, M.; Bendriss-Vermare, N.; et al. Hepatitis B virus-induced modulation of liver macrophage function promotes hepatocyte infection. J. Hepatol. 2019, 71, 1086–1098. [Google Scholar] [CrossRef]
- Barron, L.; Wynn, T.A. Macrophage activation governs schistosomiasis-induced inflammation and fibrosis. Eur. J. Immunol. 2011, 41, 2509–2514. [Google Scholar] [CrossRef] [Green Version]
- Feyder, M.; Goff, L.A. Investigating long noncoding RNAs using animal models. J. Clin. Investig. 2016, 126, 2783–2791. [Google Scholar] [CrossRef] [Green Version]
- Begolli, R.; Sideris, N.; Giakountis, A. LncRNAs as Chromatin Regulators in Cancer: From Molecular Function to Clinical Potential. Cancers 2019, 11, 1524. [Google Scholar] [CrossRef] [Green Version]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef]
- Carlevaro-Fita, J.; Lanzos, A.; Feuerbach, L.; Hong, C.; Mas-Ponte, D.; Pedersen, J.S.; Drivers, P.; Functional Interpretation, G.; Johnson, R.; Consortium, P. Cancer LncRNA Census reveals evidence for deep functional conservation of long noncoding RNAs in tumorigenesis. Commun. Biol. 2020, 3, 56. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.L.; Huang, L.Z.; Wang, Y.; Luo, W.S.; Li, F.; Xiao, J.; Qin, S.R.; Wang, Z.Y.; Song, X.W.; Wang, Y.; et al. Single-cell RNA-sequencing analysis identifies host long noncoding RNA MAMDC2-AS1 as a co-factor for HSV-1 nuclear transport. Int. J. Biol. Sci. 2020, 16, 1586–1603. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zhang, J.; Wen, L.; Lin, A. Membrane-lipid associated lncRNA: A new regulator in cancer signaling. Cancer Lett. 2018, 419, 27–29. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Kowluru, R.A. Long Noncoding RNA MALAT1 and Regulation of the Antioxidant Defense System in Diabetic Retinopathy. Diabetes 2021, 70, 227–239. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, S.; Shang, J.; Jiang, Y.; Dai, Y.; Xu, B.; Yu, Y.; Liang, Z.; Yang, Y. LncRNA: Shedding light on mechanisms and opportunities in fibrosis and aging. Ageing Res. Rev. 2019, 52, 17–31. [Google Scholar] [CrossRef]
- Hu, B.; Ma, X.; Fu, P.; Sun, Q.; Tang, W.; Sun, H.; Yang, Z.; Yu, M.; Zhou, J.; Fan, J.; et al. The mRNA-miRNA-lncRNA Regulatory Network and Factors Associated with Prognosis Prediction of Hepatocellular Carcinoma. Genom. Proteom. Bioinform. 2021, 19, 913–925. [Google Scholar] [CrossRef]
- DiStefano, J.K.; Gerhard, G.S. Long Noncoding RNAs and Human Liver Disease. Annu. Rev. Pathol. 2022, 17, 1–21. [Google Scholar] [CrossRef]
- Xia, T.; Giri, B.R.; Liu, J.; Du, P.; Li, X.; Li, X.; Li, S.; Cheng, G. RNA sequencing analysis of altered expression of long noncoding RNAs associated with Schistosoma japonicum infection in the murine liver and spleen. Parasites Vectors 2020, 13, 601. [Google Scholar] [CrossRef]
- Kho, C. Tough Decoy-Mediated Cardiac Gene Suppression. Methods Mol. Biol. 2022, 2573, 13–30. [Google Scholar] [CrossRef]
- Guo, K.; Qian, K.; Shi, Y.; Sun, T.; Wang, Z. LncRNA-MIAT promotes thyroid cancer progression and function as ceRNA to target EZH2 by sponging miR-150-5p. Cell Death Dis. 2021, 12, 1097. [Google Scholar] [CrossRef]
- Chuah, C.; Jones, M.K.; Burke, M.L.; Owen, H.C.; Anthony, B.J.; McManus, D.P.; Ramm, G.A.; Gobert, G.N. Spatial and temporal transcriptomics of Schistosoma japonicum-induced hepatic granuloma formation reveals novel roles for neutrophils. J. Leukoc. Biol. 2013, 94, 353–365. [Google Scholar] [CrossRef]
- Mederacke, I.; Dapito, D.H.; Affo, S.; Uchinami, H.; Schwabe, R.F. High-yield and high-purity isolation of hepatic stellate cells from normal and fibrotic mouse livers. Nature Protocols 2015, 10, 305–315. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Yang, D.Y.; Fan, X.L.; Zhang, M.W.; Li, Y.; Gu, X.B.; Yang, M.Y. The Roles and Mechanisms of lncRNAs in Liver Fibrosis. Int. J. Mol. Sci. 2020, 21, 1482. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Han, X.H.; Zhang, Z.; Zheng, L.N.; Hu, Z.M.; Yao, Q.B.; Cui, H.M.; Shu, G.M.; Si, M.J.; Li, C.; et al. The liver-enriched lnc-LFAR1 promotes liver fibrosis by activating TGF beta and Notch pathways. Nature Commun. 2017, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Shi, Z.M.; Zhang, M.X.; Dong, X.Y.; Zheng, L.N.; Li, G.T.; Han, X.H.; Yao, Z.; Han, T.; Hong, W. Silencing lncRNA Lfar1 alleviates the classical activation and pyoptosis of macrophage in hepatic fibrosis. Cell Death Dis. 2020, 11, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, K.; Lan, J.; Xu, L.; Feng, X.; Liao, H.; Xie, K.; Wu, H.; Zeng, Y. Long noncoding RNA TLNC1 promotes the growth and metastasis of liver cancer via inhibition of p53 signaling. Mol. Cancer 2022, 21, 105. [Google Scholar] [CrossRef]
- Wan, C.; Jin, F.; Du, Y.; Yang, K.; Yao, L.; Mei, Z.; Huang, W. Genistein improves schistosomiasis liver granuloma and fibrosis via dampening NF-kB signaling in mice. Parasitol. Res. 2017, 116, 1165–1174. [Google Scholar] [CrossRef]
- Song, Y.; Liu, C.; Liu, X.; Trottier, J.; Beaudoin, M.; Zhang, L.; Pope, C.; Peng, G.; Barbier, O.; Zhong, X.; et al. H19 promotes cholestatic liver fibrosis by preventing ZEB1-mediated inhibition of epithelial cell adhesion molecule. Hepatology 2017, 66, 1183–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akincilar, S.C.; Wu, L.L.; Ng, Q.F.; Chua, J.Y.H.; Unal, B.; Noda, T.; Chor, W.H.J.; Ikawa, M.; Tergaonkar, V. NAIL: An evolutionarily conserved lncRNA essential for licensing coordinated activation of p38 and NF kappa B in colitis. Gut 2021, 70, 1857–1871. [Google Scholar] [CrossRef] [PubMed]
- Pagan, A.J.; Ramakrishnan, L. The Formation and Function of Granulomas. Annu. Rev. Immunol. 2018, 36, 639–665. [Google Scholar] [CrossRef]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Chen, L.L. Linking Long Noncoding RNA Localization and Function. Trends Biochem. Sci. 2016, 41, 761–772. [Google Scholar] [CrossRef]
- Guo, C.J.; Ma, X.K.; Xing, Y.H.; Zheng, C.C.; Xu, Y.F.; Shan, L.; Zhang, J.; Wang, S.; Wang, Y.; Carmichael, G.G.; et al. Distinct Processing of lncRNAs Contributes to Non-conserved Functions in Stem Cells. Cell 2020, 181, 621–636.e622. [Google Scholar] [CrossRef]
- Sang, L.; Yang, L.; Ge, Q.; Xie, S.; Zhou, T.; Lin, A. Subcellular distribution, localization, and function of noncoding RNAs. Wiley Interdiscip. Rev. RNA 2022, 13, e1729. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.J.; Zhao, Y.H.; Yin, K.W.; Zhang, X.Q.; Liu, J. Dexmedetomidine inhibits inflammatory response and oxidative stress through regulating miR-205-5p by targeting HMGB1 in cerebral ischemic/reperfusion. Immunopharmacol. Immunotoxicol. 2021, 43, 478–486. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, R.; Tang, X.; Lin, H.; Xing, C.; Xu, N.; Dai, B.; Wang, P.; Shao, W.; Liu, M.; Shen, J.; et al. Knocking Down Gm16685 Decreases Liver Granuloma in Murine Schistosomiasis Japonica. Microorganisms 2023, 11, 796. https://doi.org/10.3390/microorganisms11030796
Zhao R, Tang X, Lin H, Xing C, Xu N, Dai B, Wang P, Shao W, Liu M, Shen J, et al. Knocking Down Gm16685 Decreases Liver Granuloma in Murine Schistosomiasis Japonica. Microorganisms. 2023; 11(3):796. https://doi.org/10.3390/microorganisms11030796
Chicago/Turabian StyleZhao, Ruyu, Xiaoxue Tang, Huiyao Lin, Chen Xing, Na Xu, Bingxin Dai, Pingping Wang, Wei Shao, Miao Liu, Jijia Shen, and et al. 2023. "Knocking Down Gm16685 Decreases Liver Granuloma in Murine Schistosomiasis Japonica" Microorganisms 11, no. 3: 796. https://doi.org/10.3390/microorganisms11030796