

Morphological and Molecular Characterization of Eimeria spp. Infecting Domestic Poultry Gallus gallus in Riyadh City, Saudi Arabia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Design and Collection of Study Animals
2.2. Detection of Coccidiosis Natural Infection
2.3. Microscopic Examination
2.4. DNA Extraction
2.5. Molecular Methods
2.6. Statistical Analysis
3. Results
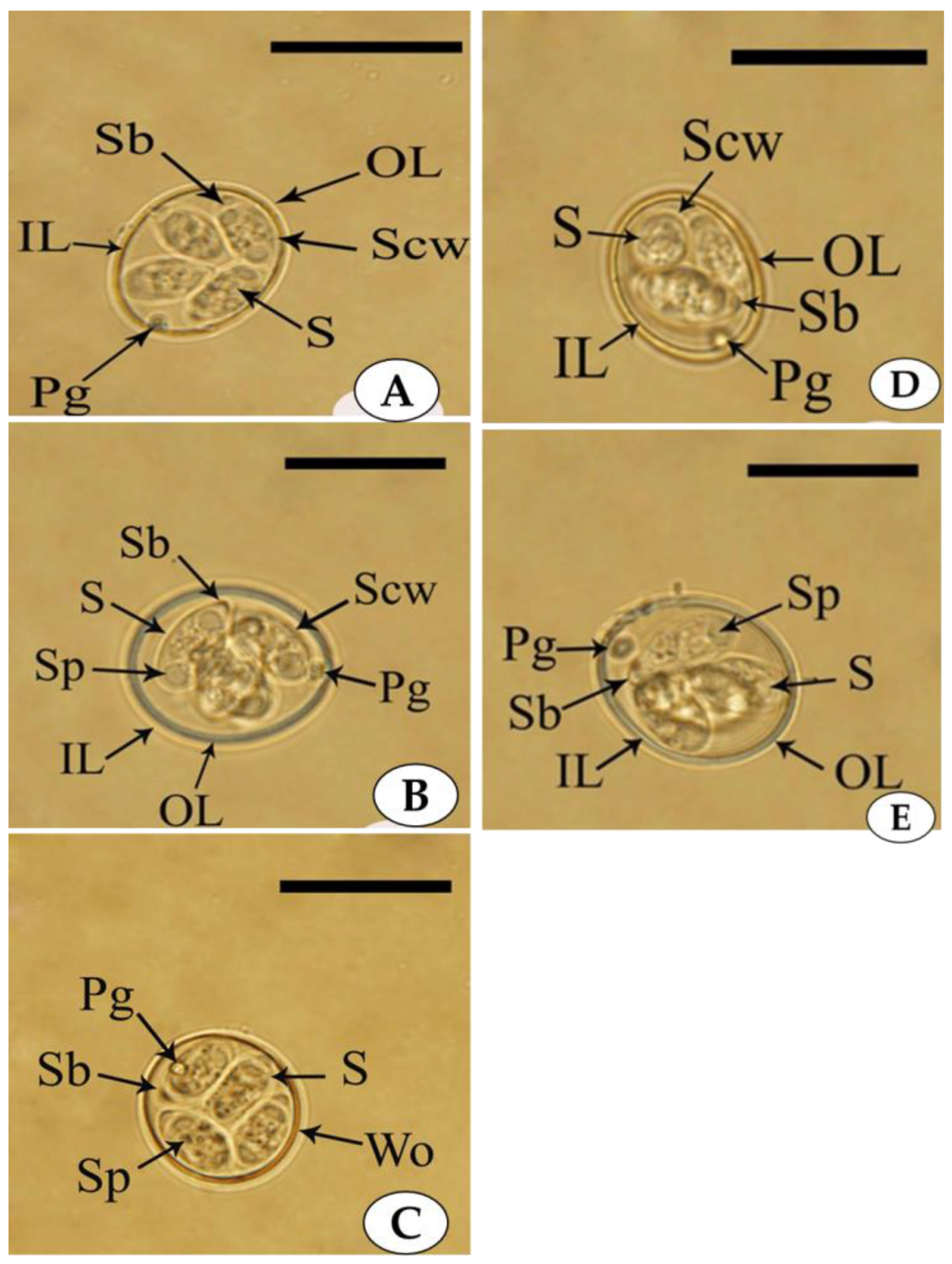
3.1. Morphological Identification
3.1.1. Eimeria tenella
3.1.2. Eimeria maxima
3.1.3. Eimeria praecox
3.1.4. Eimeria acervulina
3.1.5. Eimeria necatrix
3.2. Molecular Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sohaib, M.; Jamil, F. An insight of meat industry in Pakistan with special reference to halal meat: A comprehensive review. Korean J. Food Sci. Anim. Resour. 2017, 37, 329. [Google Scholar] [CrossRef] [PubMed]
- Blake, D.P.; Tomley, F.M. Securing poultry production from the ever-present Eimeria challenge. Trends Parasitol. 2014, 30, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroz-Castañeda, R.E. Avian coccidiosis, new strategies of treatment. In Farm Animals Diseases, Recent Omic Trends and New Strategies of Treatment; Books on Demand: Norderstedt, Germany, 2018; p. 119. [Google Scholar]
- Shirley, M.W.; Smith, A.L.; Tomley, F.M. The biology of avian Eimeria with an emphasis on their control by vaccination. Adv. Parasitol. 2005, 60, 285–330. [Google Scholar] [PubMed]
- Vrba, V.; Poplstein, M.; Pakandl, M. The discovery of the two types of small subunit ribosomal RNA gene in Eimeria mitis contests the existence of E. mivati as an independent species. Vet. Parasitol. 2011, 183, 47–53. [Google Scholar] [CrossRef]
- Williams, R. Intercurrent coccidiosis and necrotic enteritis of chickens: Rational, integrated disease management by maintenance of gut integrity. Avian Pathol. 2005, 34, 159–180. [Google Scholar] [CrossRef]
- Hammond, D.M.; Long, P.L. The Coccidia. Eimeria, Isospora, Toxoplasma, and Related Genera. 1973. Available online: https://www.cabdirect.org/cabdirect/abstract/19752900641 (accessed on 14 March 2023).
- Naidoo, V.; McGaw, L.J.; Bisschop, S.; Duncan, N.; Eloff, J.N. The value of plant extracts with antioxidant activity in attenuating coccidiosis in broiler chickens. Vet. Parasitol. 2008, 153, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Joyner, L.; Long, P. The specific characters of the Eimeria, with special reference to the coccidia of the fowl. Avian Pathol. 1974, 3, 145–157. [Google Scholar] [CrossRef]
- McDougald, L.; Reid, W. Coccidiosis. Dis. Poult. 2003, 28, 1076–1085. [Google Scholar]
- Karim, M.J.; Begum, N. Morphological and biological characterization of chicken Eimeria with special reference to species identification. Vet. Rev. Pakhribas Agric. Cent. 1994, 9, 7–9. [Google Scholar]
- Shivaramaiah, C.; Barta, J.R.; Hernandez-Velasco, X.; Téllez, G.; Hargis, B.M. Coccidiosis: Recent advance-ments in the immunobiology of Eimeria species, preventive measures, and the importance of vaccination as a control tool against these Apicomplexan parasites. Vet. Med. Res. Rep. 2014, 5, 23. [Google Scholar]
- Awais, M.M.; Akhtar, M.; Iqbal, Z.; Muhammad, F.; Anwar, M.I. Seasonal prevalence of coccidiosis in iindustrialroiler chickens in Faisalabad, Punjab, Pakistan. Trop. Anim. Health Prod. 2012, 44, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Godwin, R.M.; Morgan, J.A. A simple, one-tube assay for the simultaneous detection and diagnosis of ten A ustralian poultry E imeria. Electrophoresis 2014, 35, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Tewari, A.; Maharana, B. Control of poultry coccidiosis: Changing trends. J. Parasit. Dis. 2011, 35, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chere, M.A.; Melese, K.; Megerssa, Y.C. Molecular Characterization of Eimeria Species in Broiler Chickens, Ethiopia. In Veterinary Medicine: Research and Reports; Taylor & Francis: Abingdon, UK, 2022; pp. 153–161. [Google Scholar]
- Long, P.L. Avian Coccidiosis; Academic Press Inc.: Cambridge, MA, USA, 1993. [Google Scholar]
- López-Osorio, S.; Chaparro-Gutiérrez, J.J.; Gómez-Osorio, L.M. Overview of poultry Eimeria life cycle and host-parasite interactions. Front. Vet. Sci. 2020, 7, 384. [Google Scholar] [CrossRef] [PubMed]
- Güven, E.; Beckstead, R.B.; Sırrı, K.; Vatansever, Z.; Karaer, Z. Molecular identification of Eimeria species of broiler chickens in Turkey. Ank. Üniversitesi Vet. Fakültesi Derg. 2013, 60, 245–250. [Google Scholar]
- Allen, P.C.; Lydon, J.; Danforth, H.D. Effects of components of Artemisia annua on coccidia infections in chickens. Poult. Sci. 1997, 76, 1156–1163. [Google Scholar] [CrossRef]
- Amer, M.; Awaad, M.; El-Khateeb, R.M.; Abu-Elezz, N.; Sherein-Said, A.; Ghetas, M.; Kutkat, M. Isolation and identification of Eimeria from field coccidiosis in chickens. J. Am. Sci. 2010, 6, 1107–1114. [Google Scholar]
- Bashtar, A.-R.; Abdel-Ghaffar, F.; Al-Rasheid, K.A.; Mehlhorn, H.; Al Nasr, I. Light microscopic study on Eimeria species infecting Japanese quails reared in Saudi Arabian farms. Parasitol. Res. 2010, 107, 409–416. [Google Scholar] [CrossRef]
- Al-Quraishy, S.; Abdel-Baki, A.; Dkhil, M. Eimeria tenella infection among broiler chicks Gallus domesticus in Riyadh city, Saudi Arabia. J. King Saud Univ.-Sci. 2009, 21, 191–193. [Google Scholar] [CrossRef] [Green Version]
- Johnson, W.T. Coccidiosis of the Chicken with Special Reference to Species. 1938. Available online: https://ir.library.oregonstate.edu/downloads/qb98mf848 (accessed on 14 March 2023).
- Khaier, M.A.; Abdelhalim, A.I.; Abukashawa, S.M. Isolation and morphological identification of Eimeria tenella (Family: Eimeriidae) from Khartoum State (Sudan). J. Appl. Ind. Sci. 2015, 3, 177–181. [Google Scholar]
- Edgar, S. Sporulation of oocysts at specific temperatures and notes on the prepatent period of several species of avian coccidia. J. Parasitol. 1955, 41, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Tyzzer, E.E. Coccidiosis in gallinaceous birds. Am. J. Epidemiol. 1929, 10, 269–383. [Google Scholar] [CrossRef]
- Johnson, W.T. Coccidiosis. In Directors Biennial Report 1928–1930; The Oregon Agricultural Experiment Station: Ontario, OR, USA, 1930. [Google Scholar]
- Railliet, A.; Lucet, A. Developpement experimental des coccidies de l’epithelium intestinal du lapin et de la poule. CR Soc. Biol. Paris 1891, 36, 820–823. [Google Scholar]
- Al-Gawad, A.A.; Mahdy, O.A.; El-Massry, A.A.; Al-Aziz, M.S. Studies on coccidia of Egyptian Balady breed chickens. Life Sci. J. 2012, 9, 568–576. [Google Scholar]
- Sun, T.; Mai, S.; Norton, D.; Haycock, J.W.; Ryan, A.J.; Macneil, S. Self-organization of skin cells in three-dimensional electrospun polystyrene scaffolds. Tissue Eng. 2005, 11, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Haug, A.; Gjevre, A.-G.; Thebo, P.; Mattsson, J.G.; Kaldhusdal, M. Coccidial infections in commercial broilers: Epidemiological aspects and comparison of Eimeria species identification by morphometric and polymerase chain reaction techniques. Avian Pathol. 2008, 37, 161–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Pang, W.; Jia, T.; Yan, W.; He, G.; Hao, L.; Bentué, M.; Suo, X. Prevalence of Eimeria species in broil-ers with subclinical signs from fifty farms. Avian Dis. 2009, 53, 301–305. [Google Scholar] [CrossRef]
- Vrba, V.; Blake, D.P.; Poplstein, M. Quantitative real-time PCR assays for detection and quantification of all seven Eimeria species that infect the chicken. Vet. Parasitol. 2010, 174, 183–190. [Google Scholar] [CrossRef]
- Carvalho, F.S.; Wenceslau, A.A.; Teixeira, M.; Carneiro, J.A.M.; Melo, A.D.B.; Albuquerque, G.R. Diagnosis of Eimeria species using traditional and molecular methods in field studies. Vet. Parasitol. 2011, 176, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Lew, A.; Anderson, G.; Minchin, C.; Jeston, P.; Jorgensen, W. Inter-and intra-strain variation and PCR detec-tion of the internal transcribed spacer 1 (ITS-1) sequences of Australian isolates of Eimeria species from chickens. Vet. Parasitol. 2003, 112, 33–50. [Google Scholar] [CrossRef]
- Kalita, A.; Kakati, P.; Sarmah, P. Isolation and Molecular Identification of Eimeria Species Circulating in Broiler Chicken Flocks of Assam (India). Available online: https://www.luvas.edu.in/haryana-veterinarian/download/harvet2021-dec/23.pdf (accessed on 14 March 2023).
- Razmi, G.R.; Kalideri, G.A. Prevalence of subclinical coccidiosis in broiler-chicken farms in the municipality of Mashhad, Khorasan, Iran. Prev. Vet. Med. 2000, 44, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Hamidinejat, H.; Shapouri, M.S.; Mayahi, M.; Borujeni, M.P. Characterization of Eimeria species in commer-cial broilers by PCR based on ITS1 regions of rDNA. Iran. J. Parasitol. 2010, 5, 48. [Google Scholar]
- Siddiki, A.Z.; Mina, S.; Hasan, M.A.; Akbar, M.T.; Alam, R.; Islam, M.A.; Ira, I.Z.; Ayesa, B. Molecular charac-terization of Eimeria spp. from chicken by Polymerase Chain Reaction based on species-specific SCAR markers. J. Agric. Vet. Sci. 2014, 7, 13–17. [Google Scholar]
- Gadelhaq, S.M.; Arafa, W.M.; Aboelhadid, S.M. Molecular characterization of Eimeria species naturally in-fecting Egyptian Baldi Chickens. Iran. J. Parasitol. 2015, 10, 87. [Google Scholar] [PubMed]
- Fernandez, S.; Pagotto, A.; Furtado, M.; Katsuyama, A.; Madeira, A.M.B.N.; Gruber, A. A multiplex PCR assay for the simultaneous detection and discrimination of the seven Eimeria species that infect domestic fowl. Parasitology 2003, 127, 317–325. [Google Scholar] [CrossRef]
- Tatar, G.; Elsurer, R.; Simsek, H.; Balaban, Y.H.; Hascelik, G.; Ozcebe, O.I.; Buyukasik, Y.; Sokmensuer, C. Screening of tissue transglutaminase antibody in healthy blood donors for celiac disease screening in the Turkish population. Dig. Dis. Sci. 2004, 49, 1479–1484. [Google Scholar] [CrossRef]
- Mohammed, O.; Hussein, H. Eimeria idmii sp. n.(Apicomplexa: Eimeriidae) from the Arabian mountain gazelle, Gazella gazella, in Saudi Arabia. J. Helminthol. Soc. Wash. 1992, 59, 120–124. [Google Scholar]
- Kumar, S.; Garg, R.; Moftah, A.; Clark, E.L.; Macdonald, S.E.; Chaudhry, A.S.; Sparagano, O.; Banerjee, P.S.; Kundu, K.; Tomley, F.M. An optimised protocol for molecular identification of Eimeria from chickens. Veter-Ina. Parasitol. 2014, 199, 24–31. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Parasite | Primer Sequence 5′ → 3′ | Product Size | Annealing Temperature °C |
|---|---|---|---|
| Genus Eimeria | EF1: AAGTTGCGTAAATAGAGCCCTC ER1: AGACATCCATTGCTGAAAG | 400–600 bp | 56 |
| Eimeria acervulina (AC) | EAF: GGCTTGGATGATGTTTGCTG EAR: CGAACGCAATAACACACGCT | 321 bp | 72 |
| Eimeria praecox (PR) | EPFA: AAAA/GCAA/CAGCGATTCAAG EPRA: CCAAGCGATTTCATCATT/CGGGGA/G | 116 bp | 61 |
| Eimeria maxima (MA) | EMFA1: CT/ACACCACTCACAATGAGGCAC EMR1: GTGAT/ATCGTTC/TGG/AG/AAGTTTGC | 145 bp | 70 |
| Eimeia necatrix (NE) | ENF: TACATCCCAATCTTTGAATCG ENR: GGCATACTAGCTTCGAGCAAC | 383 bp | 61 |
| Eimeria tenella (TE) | ETF: AATTTAGTCCATCGCAACCCT ETR: CGAGCGCTCTGCATACGACA | 278 bp | 65 |
| Eimeria Species | NO. Chickens | Infected % |
|---|---|---|
| E. tenella | 13 | 10.84 |
| E. necatrix | 7 | 5.84 |
| E. acervulina | 5 | 4.16 |
| E. maxima | 3 | 2.5 |
| E. praecopx | 2 | 1.66 |
| Species | Oocyst | Sporocyst | Source | ||||
|---|---|---|---|---|---|---|---|
| Measurements | M | MC | PG | SB | SR | ||
| Eimeria praecox | 19–25 ×15.1–18.7 | − | − | + | + | − | [28] |
| 23.69 × 19.205 | − | − | + | + | − | [21] | |
| 19.7–25.7 × 15.6–19.7 | − | − | + | + | − | [26] | |
| Eimeria tenella | 20.0–26.5 ×17.0–22.0 | − | − | + | + | − | [29] |
| 21.39 × 18.745 | − | − | + | + | − | [21] | |
| 15–19 × 20–25 | − | − | + | + | − | [24] | |
| Eimeria maxima | 21.5–40.2 × 16.0–29.3 | − | − | + | + | − | [27] |
| 29.9 × 23.8 | − | − | + | + | − | [30] | |
| 21.4–42.5 × 16.5–29.8 | − | − | + | + | − | [26] | |
| Eimeria acervulina | 17.3–20.2 × 13.7–16.3 | − | − | + | + | − | [26] |
| 21.96 × 17.48 | − | − | + | + | − | [21] | |
| 18.3–27.54 × 13.7–20.40 | − | − | + | + | − | [27] | |
| Eimeria necatrix | 13.2–22.5 × 10.0–18.7 | − | − | + | + | − | [28] |
| 21.27 × 17.71 | − | − | + | + | − | [21] | |
| 13.7–22.7 × 11.3–18.3 | − | − | + | + | − | [26] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mares, M.M.; Al-Quraishy, S.; Abdel-Gaber, R.; Murshed, M. Morphological and Molecular Characterization of Eimeria spp. Infecting Domestic Poultry Gallus gallus in Riyadh City, Saudi Arabia. Microorganisms 2023, 11, 795. https://doi.org/10.3390/microorganisms11030795
Mares MM, Al-Quraishy S, Abdel-Gaber R, Murshed M. Morphological and Molecular Characterization of Eimeria spp. Infecting Domestic Poultry Gallus gallus in Riyadh City, Saudi Arabia. Microorganisms. 2023; 11(3):795. https://doi.org/10.3390/microorganisms11030795
Chicago/Turabian StyleMares, Mohammed M., Saleh Al-Quraishy, Rewaida Abdel-Gaber, and Mutee Murshed. 2023. "Morphological and Molecular Characterization of Eimeria spp. Infecting Domestic Poultry Gallus gallus in Riyadh City, Saudi Arabia" Microorganisms 11, no. 3: 795. https://doi.org/10.3390/microorganisms11030795