
Growth Dynamic and Threshold Values for Spermicidal Effects of Multidrug-Resistant Bacteria in Extended Boar Semen

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Data Analysis
2.2. Experiments
2.2.1. Chemicals and Media
2.2.2. Semen Collection and Processing
2.2.3. Microbiology
Bacterial Inoculation of Samples
Bacterial Count
2.2.4. Measurement of pH
2.2.5. Spermatology
Computer-Assisted Semen Analysis
Assessment of Sperm Agglutinations
Flowcytometry
2.2.6. Experimental Design
Experiment 1: Growth Dynamic of Resistant Bacteria and Sperm Quality
Experiment 2: Bacterial Growth in Semen Extenders at 17 °C and 5 °C
2.3. Statistical Analysis
3. Results
3.1. Field Data Analysis
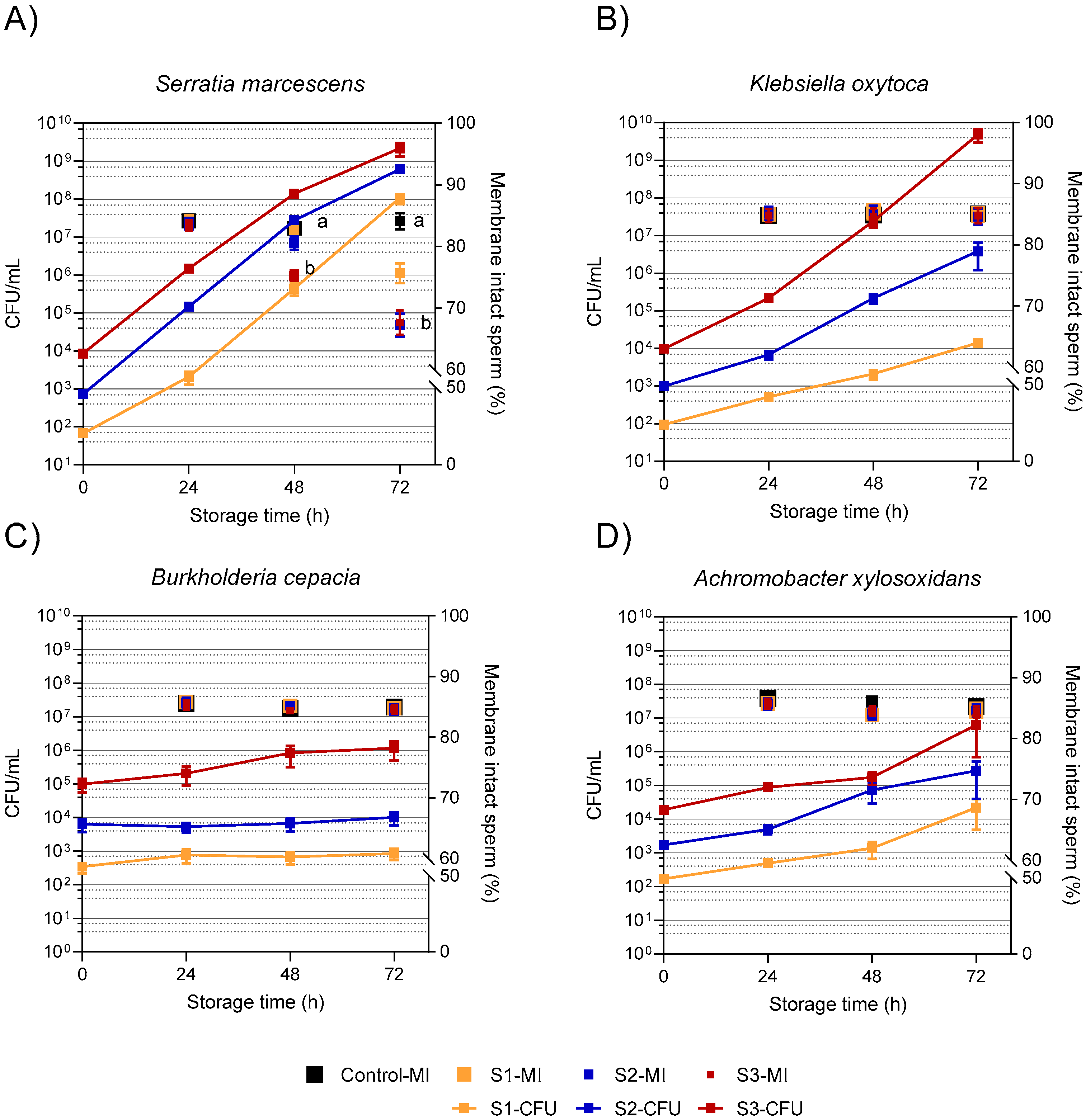
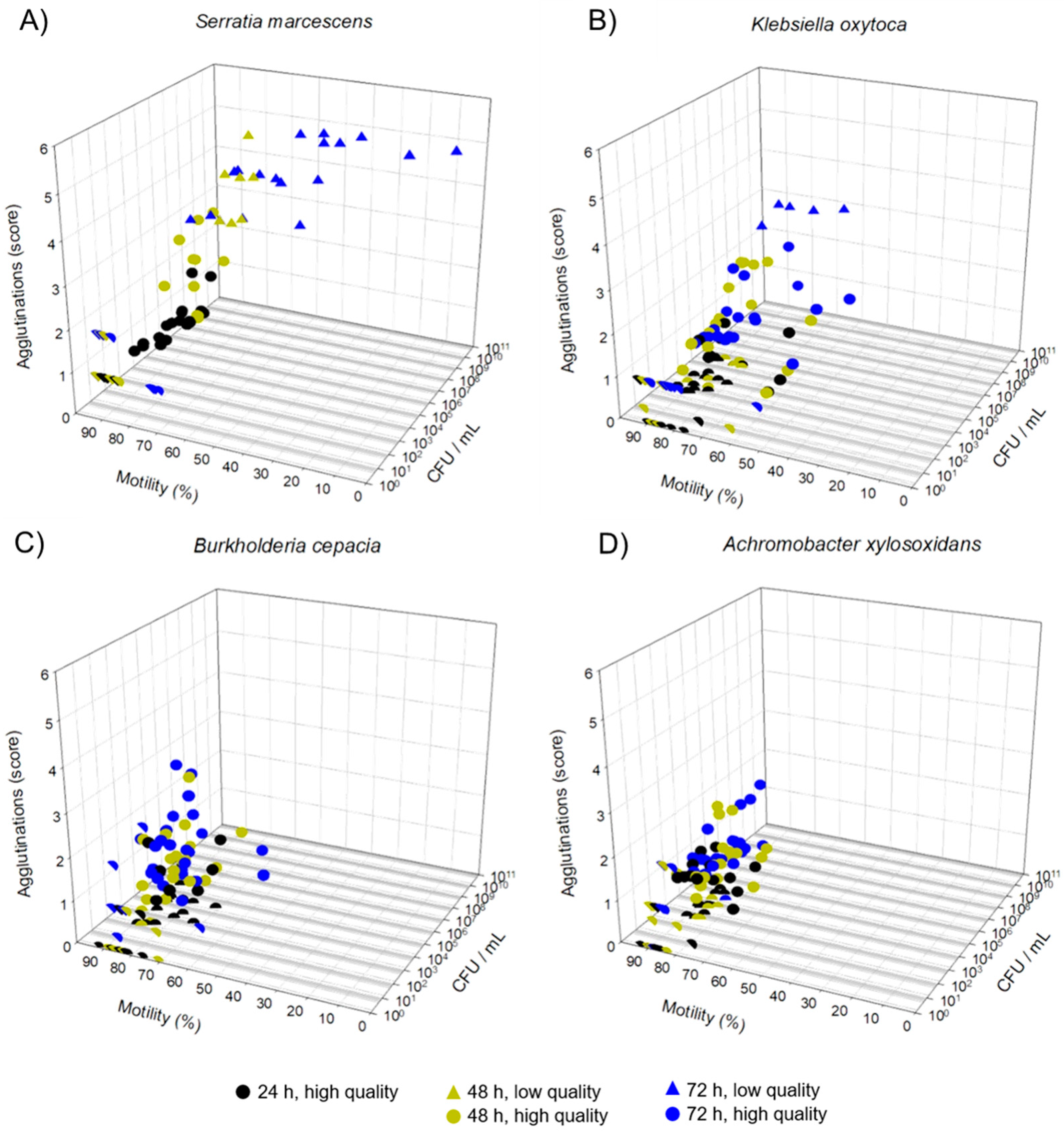
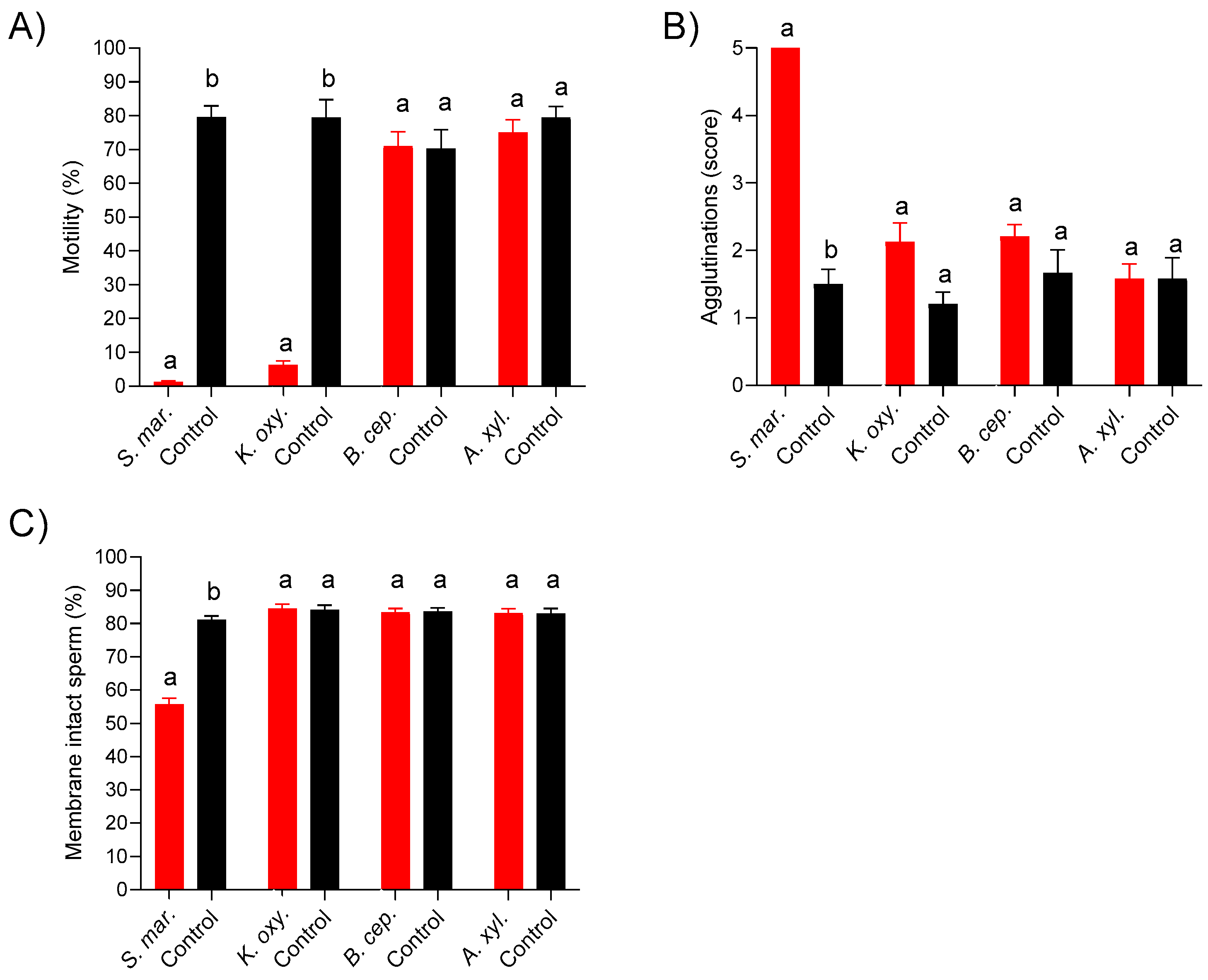
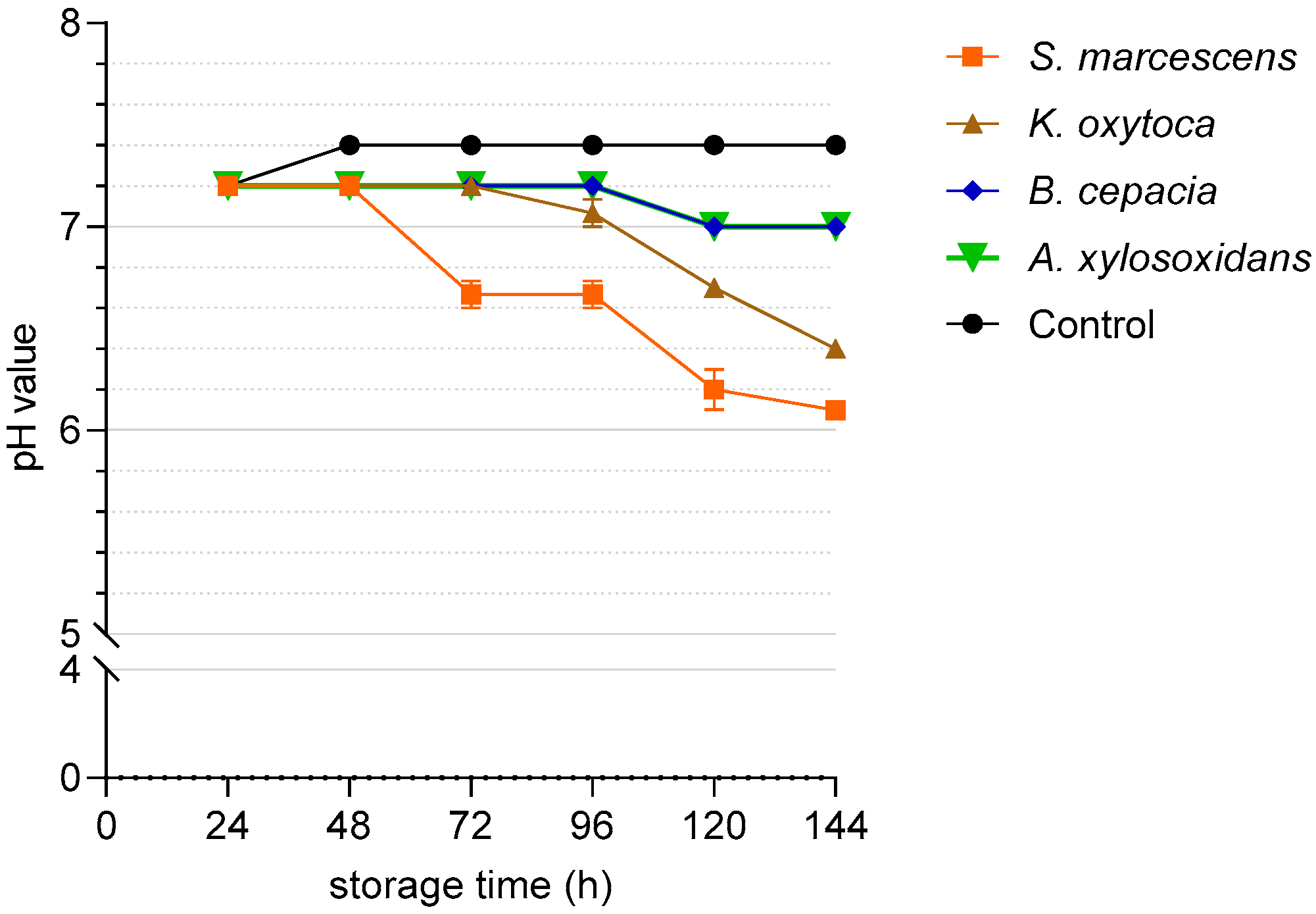
3.2. Experiment 1: Growth Dynamic of Resistant Bacteria and Sperm Quality
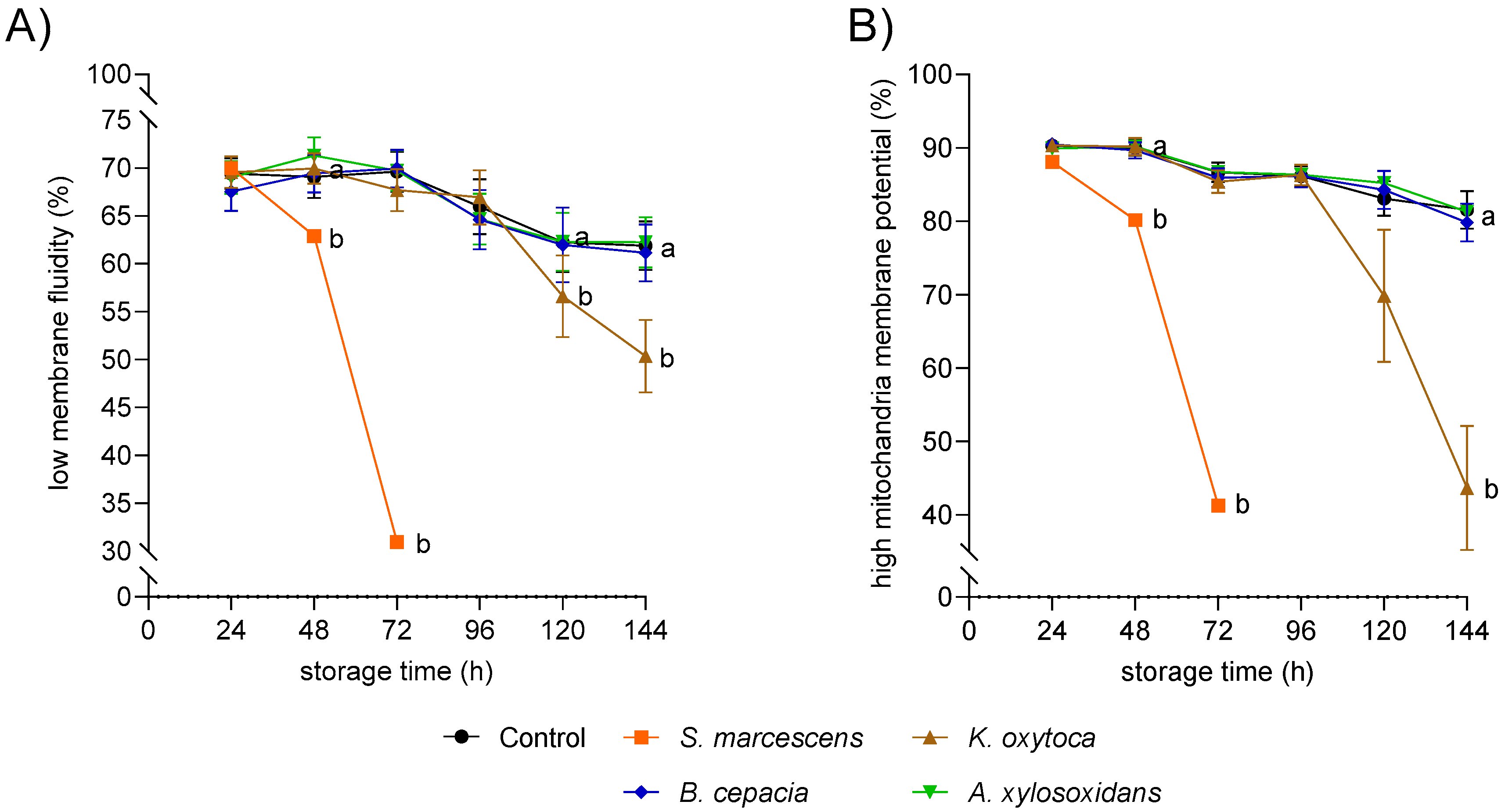
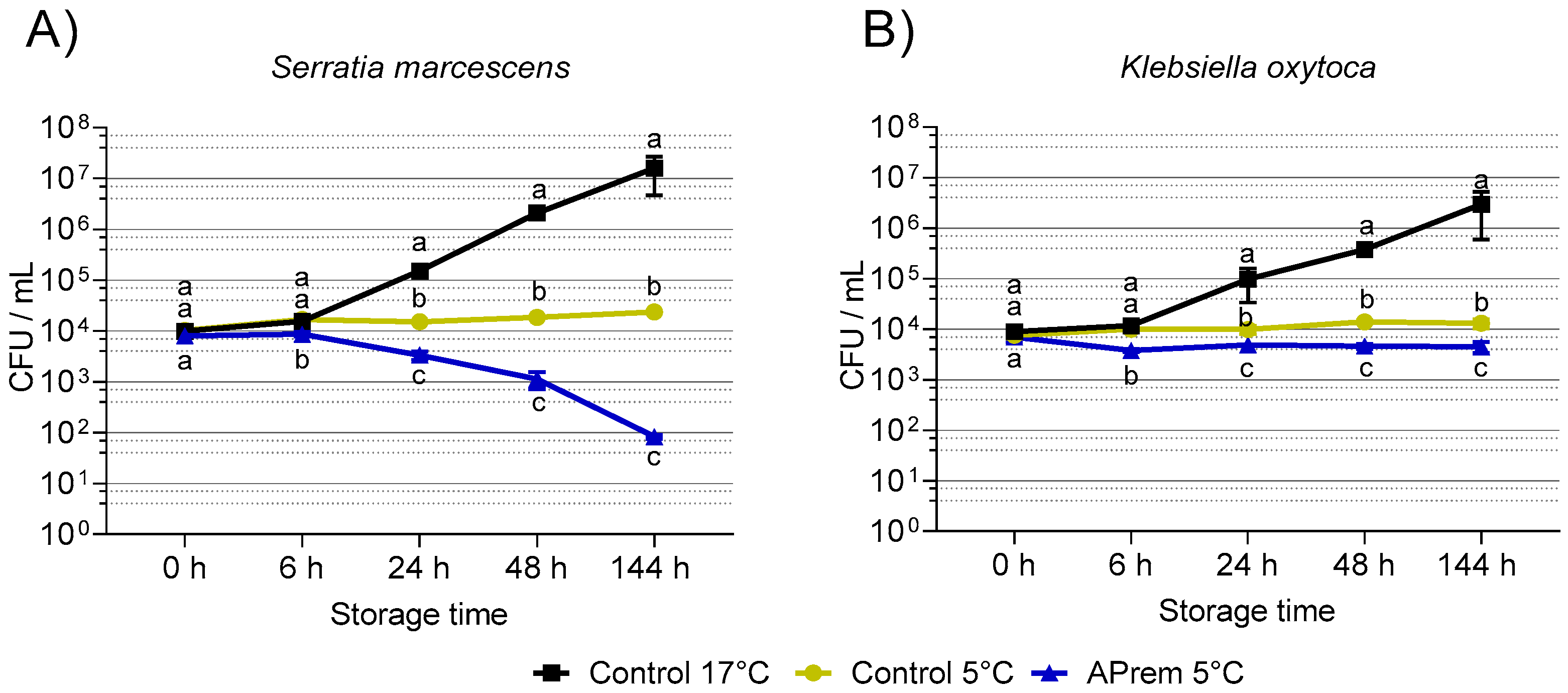
3.3. Experiment 2: Bacterial Growth in Semen Extenders at 17 °C and 5 °C
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waberski, D.; Riesenbeck, A.; Schulze, M.; Weitze, K.F.; Johnson, L. Application of Preserved Boar Semen for Artificial Insemination: Past, Present and Future Challenges. Theriogenology 2019, 137, 2–7. [Google Scholar] [CrossRef]
- AGISAR; WHO. Critically Important Antimicrobials for Human Medicine. 2019. Available online: https://apps.who.int/iris/bitstream/handle/10665/312266/9789241515528-eng.pdf (accessed on 5 September 2022).
- Nitsche-Melkus, E.; Bortfeldt, R.; Jung, M.; Schulze, M. Impact of Hygiene on Bacterial Contamination in Extended Boar Semen: An Eight-Year Retrospective Study of 28 European Ai Centers. Theriogenology 2019, 146, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M.; Wallgren, M. Alternatives to Antibiotics in Semen Extenders: A Review. Pathogens 2014, 3, 934–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, M.; Nitsche-Melkus, E.; Hensel, B.; Jung, M.; Jakop, U. Antibiotics and Their Alternatives in Artificial Breeding in Livestock. Anim. Reprod. Sci. 2020, 220, 106284. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M.; Junkes, C.; Mueller, P.; Speck, S.; Ruediger, K.; Dathe, M.; Mueller, K. Effects of Cationic Antimicrobial Peptides on Liquid-Preserved Boar Spermatozoa. PLoS ONE 2014, 9, e100490. [Google Scholar] [CrossRef] [Green Version]
- Speck, S.; Courtiol, A.; Junkes, C.; Dathe, M.; Müller, K.; Schulze, M. Cationic Synthetic Peptides: Assessment of Their Antimicrobial Potency in Liquid Preserved Boar Semen. PLoS ONE 2014, 9, e105949. [Google Scholar] [CrossRef] [Green Version]
- Feugang, J.M.; Rhoads, C.E.; Mustapha, P.A.; Tardif, S.; Parrish, J.J.; Willard, S.T.; Ryan, P.L. Treatment of Boar Sperm with Nanoparticles for Improved Fertility. Theriogenology 2019, 137, 75–81. [Google Scholar] [CrossRef]
- Pérez-Duran, F.; Acosta-Torres, L.S.; Serrano-Díaz, P.N.; Toscano-Torres, I.A.; Olivo-Zepeda, I.B.; García-Caxin, E.; Nuñez-Anita, R.E. Toxicity and Antimicrobial Effect of Silver Nanoparticles in Swine Sperms. Syst. Biol. Reprod. Med. 2020, 66, 1–9. [Google Scholar] [CrossRef]
- Barone, F.; Ventrella, D.; Zannoni, A.; Forni, M.; Bacci, M. Can Microfiltered Seminal Plasma Preserve the Morphofunctional Characteristics of Porcine Spermatozoa in the Absence of Antibiotics? A Preliminary Study. Reprod. Domest. Anim. 2016, 51, 604–610. [Google Scholar] [CrossRef]
- Morrell, J.; Núñez-González, A.; Crespo-Félez, I.; Martínez-Martínez, S.; Alborcia, M.-J.M.; Fernández-Alegre, E.; Dominguez, J.; Gutiérrez-Martín, C.; Martínez-Pastor, F. Removal of Bacteria from Boar Semen Using a Low-Density Colloid. Theriogenology 2018, 126, 272–278. [Google Scholar] [CrossRef]
- Althouse, G.; Kuster, C.; Clark, S.; Weisiger, R. Field Investigations of Bacterial Contaminants and Their Effects on Extended Porcine Semen. Theriogenology 2000, 53, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Bussalleu, E.; Yeste, M.; Sepúlveda, L.; Torner, E.; Pinart, E.; Bonet, S. Effects of Different Concentrations of Enterotoxigenic and Verotoxigenic E. Coli on Boar Sperm Quality. Anim. Reprod. Sci. 2011, 127, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, L.; Bussalleu, E.; Yeste, M.; Bonet, S. Effects of Different Concentrations of Pseudomonas Aeruginosa on Boar Sperm Quality. Anim. Reprod. Sci. 2014, 150, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Pinart, E.; Domènech, E.; Bussalleu, E.; Yeste, M.; Bonet, S. A Comparative Study of the Effects of Escherichia Coli and Clostridium Perfringens Upon Boar Semen Preserved in Liquid Storage. Anim. Reprod. Sci. 2017, 177, 65–78. [Google Scholar] [CrossRef]
- Maroto Martín, L.O.; Muñoz, E.C.; De Cupere, F.; Van Driessche, E.; Echemendia-Blanco, D.; Rodríguez, J.M.M.; Beeckmans, S. Bacterial Contamination of Boar Semen Affects the Litter Size. Anim. Reprod. Sci. 2010, 120, 95–104. [Google Scholar] [CrossRef]
- Althouse, G.C.; Lu, K.G. Bacteriospermia in Extended Porcine Semen. Theriogenology 2005, 63, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Úbeda, J.L.; Ausejo, R.; Dahmani, Y.; Falceto, M.V.; Usan, A.; Malo, C.; Perez-Martinez, F.C. Adverse Effects of Members of the Enterobacteriaceae Family on Boar Sperm Quality. Theriogenology 2013, 80, 565–570. [Google Scholar] [CrossRef]
- Schulze, M.; Ammon, C.; Rüdiger, K.; Jung, M.; Grobbel, M. Analysis of Hygienic Critical Control Points in Boar Semen Production. Theriogenology 2015, 83, 430–437. [Google Scholar] [CrossRef]
- Tvrdá, E.; Bučko, O.; Rojková, K.; Ďuračka, M.; Kunová, S.; Kováč, J.; Benko, F.; Kačániová, M. The Efficiency of Selected Extenders against Bacterial Contamination of Boar Semen in a Swine Breeding Facility in Western Slovakia. Animals 2021, 11, 3320. [Google Scholar] [CrossRef]
- Costinar, L.; Herman, V.; Pitoiu, E.; Iancu, I.; Degi, J.; Hulea, A.; Pascu, C. Boar Semen Contamination: Identification of Gram-Negative Bacteria and Antimicrobial Resistance Profile. Animals 2021, 12, 43. [Google Scholar] [CrossRef]
- Althouse, G.; Pierdon, M.; Lu, K. Thermotemporal Dynamics of Contaminant Bacteria and Antimicrobials in Extended Porcine Semen. Theriogenology 2008, 70, 1317–1323. [Google Scholar] [CrossRef]
- Gòdia, M.; Ramayo-Caldas, Y.; Zingaretti, L.M.; Darwich, L.; López, S.; Rodríguez-Gil, J.E.; Yeste, M.; Sánchez, A.; Clop, A. A Pilot Rna-Seq Study in 40 Pietrain Ejaculates to Characterize the Porcine Sperm Microbiome. Theriogenology 2020, 157, 525–533. [Google Scholar] [CrossRef]
- Johnson, L.A.; Weitze, K.F.; Fiser, P.; Maxwell, W.M.C. Storage of Boar Semen. Anim. Reprod. Sci. 2000, 62, 143–172. [Google Scholar] [CrossRef] [PubMed]
- Höfner, L.; Luther, A.-M.; Palladini, A.; Fröhlich, T.; Waberski, D. Tolerance of Stored Boar Spermatozoa to Autologous Seminal Plasma: A Proteomic and Lipidomic Approach. Int. J. Mol. Sci. 2020, 21, 6474. [Google Scholar] [CrossRef]
- Jäkel, H.; Scheinpflug, K.; Mühldorfer, K.; Gianluppi, R.; Lucca, M.S.; Mellagi, A.P.G.; Bortolozzo, F.P.; Waberski, D. In Vitro Performance and in Vivo Fertility of Antibiotic-Free Preserved Boar Semen Stored at 5 °C. J. Anim. Sci. Biotechnol. 2021, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Luther, A.-M.; Nguyen, T.Q.; Verspohl, J.; Waberski, D. Antimicrobially Active Semen Extenders Allow the Reduction of Antibiotic Use in Pig Insemination. Antibiotics 2021, 10, 1319. [Google Scholar] [CrossRef]
- Gatti, J.-L.; Chevrier, C.; Paquignon, M.; Dacheux, J.-L. External Ionic Conditions, Internal Ph and Motility of Ram and Boar Spermatozoa. J. Reprod. Fertil. 1993, 98, 439–449. [Google Scholar] [CrossRef] [Green Version]
- PPark, Y.-J.; Shin, D.-H.; Pang, W.-K.; Ryu, D.-Y.; Rahman, S.; Adegoke, E.O.; Pang, M.-G. Short-Term Storage of Semen Samples in Acidic Extender Increases the Proportion of Females in Pigs. BMC Veter-Res. 2021, 17, 362. [Google Scholar] [CrossRef]
- Riesenbeck, A.; Schulze, M.; Rüdiger, K.; Henning, H.; Waberski, D. Quality Control of Boar Sperm Processing: Implications from European Ai Centres and Two Spermatology Reference Laboratories. Reprod. Domest. Anim. 2015, 50 (Suppl. 2), 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, M.; Jung, M.; Hensel, B. Science-Based Quality Control in Boar Semen Production. Mol. Reprod. Dev. 2022, 1–9. [Google Scholar] [CrossRef]
- Gączarzewicz, D.; Udała, J.; Piasecka, M.; Błaszczyk, B.; Stankiewicz, T. Bacterial Contamination of Boar Semen and Its Relationship to Sperm Quality Preserved in Commercial Extender Containing Gentamicin Sulfate. Pol. J. Veter- Sci. 2016, 19, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Pastor, F.; Lacalle, E.; Martínez-Martínez, S.; Fernández-Alegre, E.; Álvarez-Fernández, L.; Martinez-Alborcia, M.J.; Bolarin, A.; Morrell, J.M. Low Density Porcicoll Separates Spermatozoa from Bacteria and Retains Sperm Quality. Theriogenology 2021, 165, 28–36. [Google Scholar] [CrossRef]
- Schmid, S.; Henning, H.; Petrunkina, A.M.; Weitze, K.F.; Waberski, D. Response to Capacitating Stimuli Indicates Extender-Related Differences in Boar Sperm Function. J. Anim. Sci. 2013, 91, 5018–5025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drobnis, E.Z.; Crowe, L.M.; Berger, T.; Anchordoguy, T.J.; Overstreet, J.W.; Crowe, J.H. Cold Shock Damage Is Due to Lipid Phase Transitions in Cell Membranes: A Demonstration Using Sperm as a Model. J. Exp. Zool. 1993, 265, 432–437. [Google Scholar] [CrossRef] [PubMed]
- White, I.G. Lipids and Calcium Uptake of Sperm in Relation to Cold Shock and Preservation: A Review. Reprod. Fertil. Dev. 1993, 5, 639–658. [Google Scholar] [CrossRef] [PubMed]
- Schmid, S.; Henning, H.; Oldenhof, H.; Wolkers, W.F.; Petrunkina, A.M.; Waberski, D. The Specific Response to Capacitating Stimuli Is a Sensitive Indicator of Chilling Injury in Hypothermically Stored Boar Spermatozoa. Andrology 2013, 1, 376–386. [Google Scholar] [CrossRef]
- Waberski, D.; Luther, A.-M.; Grünther, B.; Jäkel, H.; Henning, H.; Vogel, C.; Peralta, W.; Weitze, K.F. Sperm Function in Vitro and Fertility after Antibiotic-Free, Hypothermic Storage of Liquid Preserved Boar Semen. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Menezes, T.A.; Mellagi, A.P.G.; da Silva Oliveira, G.; Bernardi, M.L.; Wentz, I.; Ulguim, R.D.R.; Bortolozzo, F.P. Antibiotic-Free Extended Boar Semen Preserved under Low Temperature Maintains Acceptable in-Vitro Sperm Quality and Reduces Bacterial Load. Theriogenology 2020, 149, 131–138. [Google Scholar] [CrossRef]
- Jäkel, H.; Henning, H.; Luther, A.-M.; Rohn, K.; Waberski, D. Assessment of Chilling Injury in Hypothermic Stored Boar Spermatozoa by Multicolor Flow Cytometry. Cytom. A 2021, 99, 1033–1041. [Google Scholar] [CrossRef]
- Rowe, M.; Veerus, L.; Trosvik, P.; Buckling, A.; Pizzari, T. The Reproductive Microbiome: An Emerging Driver of Sexual Selection, Sexual Conflict, Mating Systems, and Reproductive Isolation. Trends Ecol. Evol. 2020, 35, 220–234. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | AI Centers (n) | Semen Samples (n) | High Quality, No Bacterial Isolation (n) | High Quality, Bacterial Isolation (n) | Low Quality, No Bacterial Isolation (n) | Low Quality, Bacterial Isolation (n) | Bacteria Type |
|---|---|---|---|---|---|---|---|
| 2016 | 26 | 513 | 437 | 7 * | 67 | 2 | B. species, E. coli, K. oxytoca |
| 2017 | 24 | 480 | 466 | 0 | 14 | 0 | - |
| 2018 | 24 | 479 | 424 | 0 | 55 | 0 | - |
| 2019 | 23 | 450 | 436 | 2 ** | 12 | 0 | A. xylosoxidans, S. marcescens |
| 2020 | 22 | 440 | 427 | 5 ** | 7 | 1 | K. oxytoca |
| 2021 | 22 | 437 | 396 | 0 | 41 | 0 | - |
| 2022 | 21 | 420 | 362 | 0 | 58 | 0 | - |
| Sum | 3219 | 2948 (91.6%) | 14 (0.4%) | 254 (7.9%) | 3 (0.1%) |
| Bacteria (All Samples/Low Quality Samples) | ||||
|---|---|---|---|---|
| Bacterial Count CFU/mL | Serratia marcescens | Klebsiella oxytoca | Burkholderia cepacia | Achromobacter xylosoxidans |
| All (n)/Low (n) | All (n)/Low (n) | All (n)/Low (n) | All (n)/Low (n) | |
| <106 | 18/0 | 51/0 | 69/0 | 69/0 |
| 106–107 | 6/0 | 8/0 | 3/0 | 2/0 |
| 107–108 | 9/5 | 7/1 | - | 1/0 |
| >108 | 21/21 | 6/4 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luther, A.-M.; Beckermann, C.; Nguyen, T.Q.; Verspohl, J.; Waberski, D. Growth Dynamic and Threshold Values for Spermicidal Effects of Multidrug-Resistant Bacteria in Extended Boar Semen. Microorganisms 2023, 11, 788. https://doi.org/10.3390/microorganisms11030788
Luther A-M, Beckermann C, Nguyen TQ, Verspohl J, Waberski D. Growth Dynamic and Threshold Values for Spermicidal Effects of Multidrug-Resistant Bacteria in Extended Boar Semen. Microorganisms. 2023; 11(3):788. https://doi.org/10.3390/microorganisms11030788
Chicago/Turabian StyleLuther, Anne-Marie, Christina Beckermann, Thu Quynh Nguyen, Jutta Verspohl, and Dagmar Waberski. 2023. "Growth Dynamic and Threshold Values for Spermicidal Effects of Multidrug-Resistant Bacteria in Extended Boar Semen" Microorganisms 11, no. 3: 788. https://doi.org/10.3390/microorganisms11030788