
Ticks and Tick-Borne Pathogens in Recreational Greenspaces in North Central Florida, USA
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Site Selection
2.2. Sampling Design and Field Methods
2.3. Tick Identification and Storage
2.4. DNA Extraction
2.5. Polymerase Chain Reaction, Gel Electrophoresis, and Sequencing
2.6. Tick Density and Abundance Analysis
2.7. Infected Tick Microorganism and Pathogen Analysis
3. Results
3.1. Tick Collections and Density Measurements
3.2. Tick-Associated Bacterial and Protozoal Microorganisms
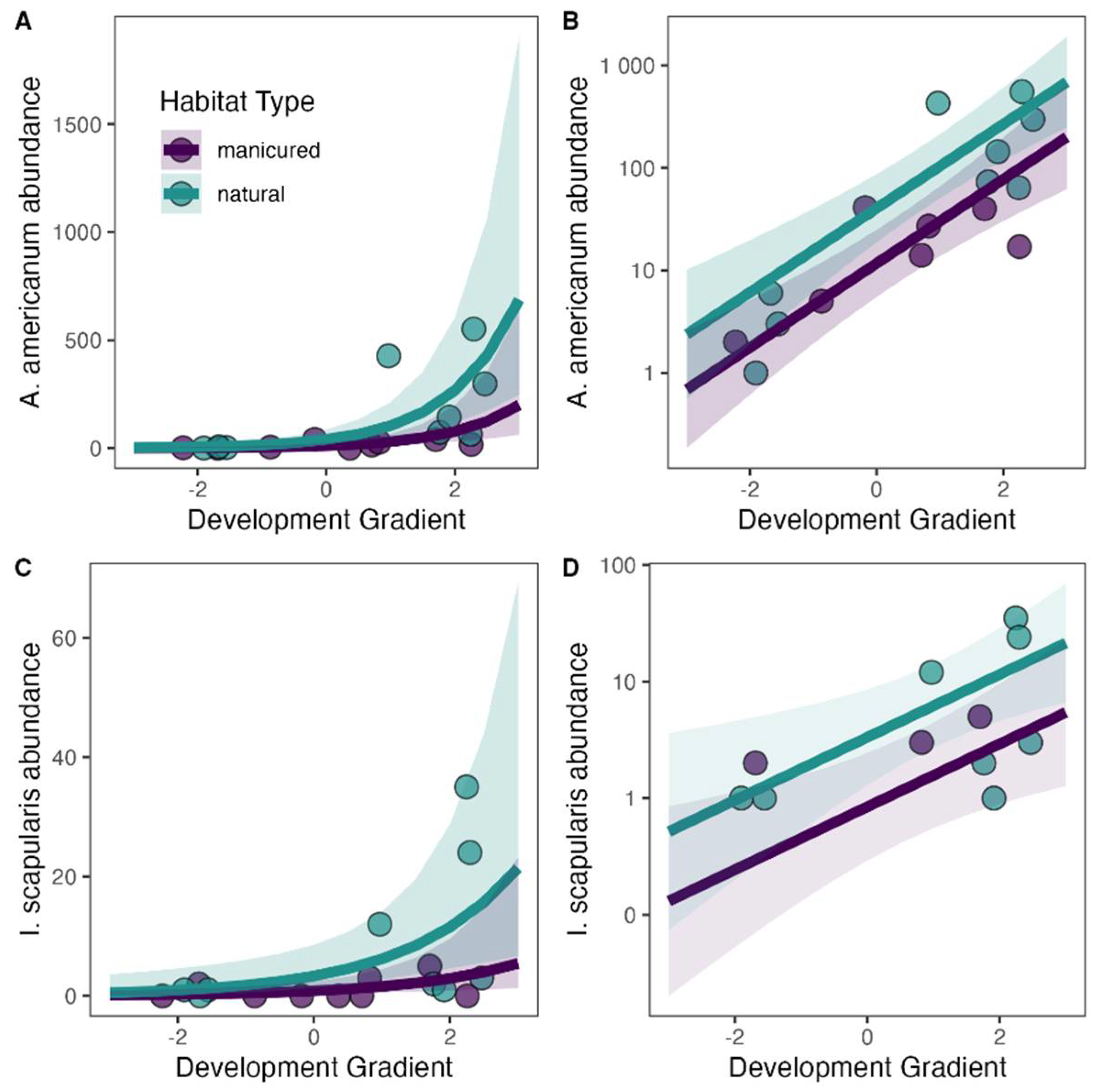
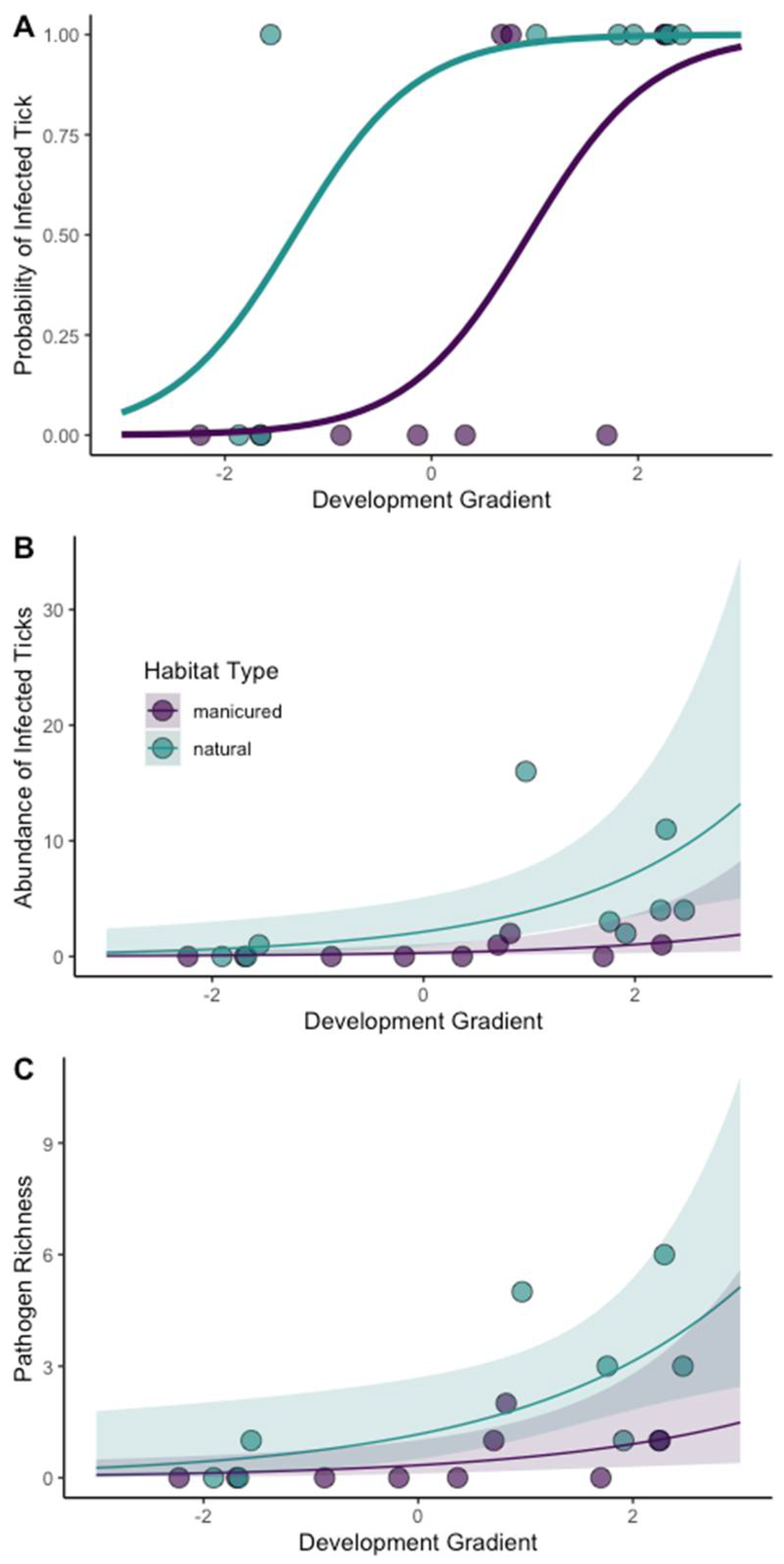
3.3. Effect of Habitat Type and Landcover on Tick Abundance and Pathogen Infection
4. Discussion
4.1. Tick Abundance
4.2. Tick-Borne Bacteria and Protozoa
4.3. Effect of Human Development on Ticks and Tick-Borne Microorganisms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van den Berg, A.E.; Maas, J.; Verheij, R.A.; Groenewegen, P.P. Green Space as a Buffer between Stressful Life Events and Health. Soc. Sci. Med. 2010, 70, 1203–1210. [Google Scholar] [CrossRef] [Green Version]
- Lovell, S.T.; Taylor, J.R. Supplying Urban Ecosystem Services through Multifunctional Green Infrastructure in the United States. Landsc. Ecol. 2013, 28, 1447–1463. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Aronson, M.F.J.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; Scott MacIvor, J. Biodiversity in the City: Fundamental Questions for Understanding the Ecology of Urban Green Spaces for Biodiversity Conservation. BioScience 2017, 67, 799–807. [Google Scholar] [CrossRef] [Green Version]
- Soanes, K.; Lentini, P.E. When Cities Are the Last Chance for Saving Species. Front. Ecol. Environ. 2019, 17, 225–231. [Google Scholar] [CrossRef]
- Mackenstedt, U.; Jenkins, D.; Romig, T. The Role of Wildlife in the Transmission of Parasitic Zoonoses in Peri-Urban and Urban Areas. Int. J. Parasitol. Parasites Wildl. 2015, 4, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Heylen, D.; Lasters, R.; Adriaensen, F.; Fonville, M.; Sprong, H.; Matthysen, E. Ticks and Tick-Borne Diseases in the City: Role of Landscape Connectivity and Green Space Characteristics in a Metropolitan Area. Sci. Total Environ. 2019, 670, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Noden, B.H.; Loss, S.R.; Maichak, C.; Williams, F. Risk of Encountering Ticks and Tick-Borne Pathogens in a Rapidly Growing Metropolitan Area in the U.S. Great Plains. Ticks Tick Borne Dis. 2017, 8, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Noden, B.H.; Roselli, M.A.; Loss, S.R. Effect of Urbanization on Presence, Abundance, and Coinfection of Bacteria and Protozoa in Ticks in the US Great Plains. J. Med. Entomol. 2022, 59, 957–968. [Google Scholar] [CrossRef]
- VanAcker, M.C.; Little, E.A.H.; Molaei, G.; Bajwa, W.I.; Diuk-Wasser, M.A. Enhancement of Risk for Lyme Disease by Landscape Connectivity, New York, NY, USA. Emerg. Infect. Dis. 2019, 25, 1136–1143. [Google Scholar] [CrossRef]
- Simmons, A.E.; Manges, A.B.; Bharathan, T.; Tepe, S.L.; McBride, S.E.; DiLeonardo, M.Q.; Duchamp, J.E.; Simmons, T.W. Lyme Disease Risk of Exposure to Blacklegged Ticks (Acari: Ixodidae) Infected with Borrelia Burgdorferi (Spirochaetales: Spirochaetaceae) in Pittsburgh Regional Parks. J. Med. Entomol. 2020, 57, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Sonenshine, D.E. Range Expansion of Tick Disease Vectors in North America: Implications for Spread of Tick-Borne Disease. Int. J. Environ. Res. Public Health 2018, 15, 478. [Google Scholar] [CrossRef] [Green Version]
- Blanton, L.S.; Walker, D.H.; Bouyer, D.H. Rickettsiae and Ehrlichiae within a City Park: Is the Urban Dweller at Risk? Vector Borne Zoonotic Dis. 2014, 14, 168–170. [Google Scholar] [CrossRef] [PubMed]
- Comer, J.A.; Paddock, C.D.; Childs, J.E. Urban Zoonoses Caused by Bartonella, Coxiella, Ehrlichia, and Rickettsia Species. Vector Borne Zoonotic Dis. 2001, 1, 91–118. [Google Scholar] [CrossRef] [PubMed]
- Roselli, M.A. Urban Tick Ecology in Oklahoma City: Tick Distribution, Pathogen Prevalence and Avian Infestation across an Urbanization Gradient. Master’s Thesis, Oklahoma State University, Stillwater, OK, USA, 2019. [Google Scholar]
- Small, M.; Brennan, R.E. Detection of Rickettsia amblyommatis and Ehrlichia chaffeensis in Amblyomma americanum Inhabiting Two Urban Parks in Oklahoma. Vector Borne Zoonotic Dis. 2021, 21, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Diuk-Wasser, M.A.; VanAcker, M.C.; Fernandez, M.P. Impact of Land Use Changes and Habitat Fragmentation on the Eco-Epidemiology of Tick-Borne Diseases. J. Med. Entomol. 2021, 58, 1546–1564. [Google Scholar] [CrossRef]
- Stromdahl, E.Y.; Hickling, G.J. Beyond Lyme: Aetiology of Tick-Borne Human Diseases with Emphasis on the South-Eastern United States. Zoonoses Public Health 2012, 59 (Suppl. S2), 48–64. [Google Scholar] [CrossRef]
- Glass, G.E.; Ganser, C.; Wisely, S.M.; Kessler, W.H. Standardized Ixodid Tick Survey in Mainland Florida. Insects 2019, 10, 235. [Google Scholar] [CrossRef] [Green Version]
- De Jesus, C.E.; Ganser, C.; Kessler, W.H.; White, Z.S.; Bhosale, C.R.; Glass, G.E.; Wisely, S.M. A Survey of Tick-Borne Bacterial Pathogens in Florida. Insects 2019, 10, 297. [Google Scholar] [CrossRef] [Green Version]
- Florida Department of Health Annual Morbidity Statistics Reports; Florida Department of Health: Tallahassee, FL, USA, 2018.
- Lloyd, V.K.; Hawkins, R.G. Under-Detection of Lyme Disease in Canada. Healthcare 2018, 6, 125. [Google Scholar] [CrossRef] [Green Version]
- Carson, D.A.; Kopsco, H.; Gronemeyer, P.; Mateus-Pinilla, N.; Smith, G.S.; Sandstrom, E.N.; Smith, R.L. Knowledge, Attitudes, and Practices of Illinois Medical Professionals Related to Ticks and Tick-Borne Disease. One Health 2022, 15, 100424. [Google Scholar] [CrossRef]
- Dykstra, E.A.; Oltean, H.N.; Kangiser, D.; Marsden-Haug, N.; Rich, S.M.; Xu, G.; Lee, M.-K.; Morshed, M.G.; Graham, C.B.; Eisen, R.J. Ecology and Epidemiology of Tickborne Pathogens, Washington, USA, 2011–2016. Emerg. Infect. Dis. 2020, 26, 648–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulachenski, A.; Ghersi, B.; Lesen, A.; Blum, M. Abandonment, Ecological Assembly and Public Health Risks in Counter-Urbanizing Cities. Sustainability 2016, 8, 491. [Google Scholar] [CrossRef] [Green Version]
- Hansford, K.M.; Wheeler, B.W.; Tschirren, B.; Medlock, J.M. Questing Ixodes Ricinus Ticks and Borrelia Spp. in Urban Green Space across Europe: A Review. Zoonoses Public Health 2022, 69, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Kessler, W.H.; Ganser, C.; Glass, G.E. Modeling the Distribution of Medically Important Tick Species in Florida. Insects 2019, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Ginsberg, H.S.; Ewing, C.P. Comparison of Flagging, Walking, Trapping, and Collecting from Hosts as Sampling Methods for Northern Deer Ticks, Ixodes dammini, and Lone-Star Ticks, Amblyomma americanum (Acari:Ixodidae). Exp. Appl. Acarol. 1989, 7, 313–322. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Lia, R.P.; Capelli, G.; Otranto, D. Efficiency of Flagging and Dragging for Tick Collection. Exp. Appl. Acarol. 2013, 61, 119–127. [Google Scholar] [CrossRef]
- Clifford, C.M.; Anastos, G.; Van der Borght-Elbl, A. The Larval Ixodid Ticks of the Eastern United States (Acarina-Ixodidae); Entomological Society of America: Annapolis, MD, USA, 1961; Volume 26. [Google Scholar]
- Durden, L.A.; Keirans, J.E. Nymphs of the Genus Ixodes (Acari: Ixodidae) of the United States: Taxonomy, Identification Key, Distribution, Hosts, and Medical/Veterinary Importance; Entomological Society of America: Annapolis, MD, USA, 1996. [Google Scholar]
- Keirans, J.E.; Litwak, T.R. Pictorial Key to the Adults of Hard Ticks, Family Ixodidae (Ixodida: Ixodoidea), East of the Mississippi River. J. Med. Entomol. 1989, 26, 435–448. [Google Scholar] [CrossRef]
- Keirans, J.E.; Durden, L.A. Illustrated Key to Nymphs of the Tick Genus Amblyomma (Acari: Ixodidae) Found in the United States. J. Med. Entomol. 1998, 35, 489–495. [Google Scholar] [CrossRef]
- Egizi, A.M.; Robbins, R.G.; Beati, L.; Nava, S.; Vans, C.R.; Occi, J.L.; Fonseca, D.M. A Pictorial Key to Differentiate the Recently Detected Exotic Haemaphysalis longicornis Neumann, 1901 (Acari, Ixodidae) from Native Congeners in North America. Zookeys 2019, 818, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Nadolny, R.M.; Toliver, M.; Gaff, H.D.; Snodgrass, J.G.; Robbins, R.G. Focus Stacking Images of Morphological Character States for Differentiating the Adults of Ixodes affinis and Ixodes scapularis (Acari: Ixodidae) in Areas of Sympatry. J. Med. Entomol. 2021, 58, 1941–1947. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic Identification of Rickettsiae and Estimation of Intraspecies Sequence Divergence for Portions of Two Rickettsial Genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftis, A.D.; Reeves, W.K.; Szumlas, D.E.; Abbassy, M.M.; Helmy, I.M.; Moriarity, J.R.; Dasch, G.A. Surveillance of Egyptian Fleas for Agents of Public Health Significance: Anaplasma, Bartonella, Coxiella, Ehrlichia, Rickettsia, and Yersinia pestis. Am. J. Trop. Med. Hyg. 2006, 75, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayler, K.A.; Wamsley, H.L.; Pate, M.; Barbet, A.F.; Alleman, A.R. Cultivation of Rickettsia amblyommii in Tick Cells, Prevalence in Florida Lone Star Ticks (Amblyomma americanum). Parasit. Vectors 2014, 7, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledger, K.J.; Beati, L.; Wisely, S.M. Survey of Ticks and Tick-Borne Rickettsial and Protozoan Pathogens in Eswatini. Pathogens 2021, 10, 1043. [Google Scholar] [CrossRef] [PubMed]
- Lydy, S.L.; Williams-Newkirk, A.J.; Dugan, E.J.; Hensley, J.R.; Dasch, G.A. Novel PCR Exclusion Assay to Detect Spotted Fever Group Rickettsiae in the Lone Star Tick (Amblyomma americanum). Ticks Tick Borne Dis. 2020, 11, 101453. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, E.; Kock, R.; McKeever, D.; Gakuya, F.; Musyoki, C.; Chege, S.M.; Mutinda, M.; Kariuki, E.; Davidson, Z.; Low, B.; et al. Prevalence of Theileria equi and Babesia caballi as well as the Identification of Associated Ticks in Sympatric Grevy’s Zebras (Equus Grevyi) and Donkeys (Equus Africanus Asinus) in Northern Kenya. J. Wildl. Dis. 2015, 51, 137–147. [Google Scholar] [CrossRef]
- Barbour, A.G.; Maupin, G.O.; Teltow, G.J.; Carter, C.J.; Piesman, J. Identification of an Uncultivable Borrelia Species in the Hard Tick Amblyomma americanum: Possible Agent of a Lyme Disease-like Illness. J. Infect. Dis. 1996, 173, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Bunikis, J.; Garpmo, U.; Tsao, J.; Berglund, J.; Fish, D.; Barbour, A.G. Sequence Typing Reveals Extensive Strain Diversity of the Lyme Borreliosis Agents Borrelia burgdorferi in North America and Borrelia afzelii in Europe. Microbiology 2004, 150, 1741–1755. [Google Scholar] [CrossRef] [Green Version]
- De Jesus, C.; Bhosale, C.; Wilson, K.; White, Z.; Wisely, S.M. Reptile Host Associations of Ixodes scapularis in Florida and Implications for Borrelia spp. Ecology. Pathogens 2021, 10, 999. [Google Scholar] [CrossRef]
- Tabara, K.; Arai, S.; Kawabuchi, T.; Itagaki, A.; Ishihara, C.; Satoh, H.; Okabe, N.; Tsuji, M. Molecular Survey of Babesia microti, Ehrlichia Species and Candidatus Neoehrlichia mikurensis in Wild Rodents from Shimane Prefecture, Japan. Microbiol. Immunol. 2007, 51, 359–367. [Google Scholar] [CrossRef]
- Roux, V.; Rydkina, E.; Eremeeva, M.; Raoult, D. Citrate Synthase Gene Comparison, a New Tool for Phylogenetic Analysis, and Its Application for the Rickettsiae. Int. J. Syst. Bacteriol. 1997, 47, 252–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parola, P.; Cornet, J.-P.; Sanogo, Y.O.; Miller, R.S.; Van Thien, H.; Gonzalez, J.-P.; Raoult, D.; Telford, S.R., III; Wongsrichanalai, C. Detection of Ehrlichia spp., Anaplasma spp., Rickettsia spp. and Other Eubacteria in Ticks from the Thai-Myanmar Border and Vietnam. J. Clin. Microbiol. 2003, 41, 1600–1608. [Google Scholar] [CrossRef] [Green Version]
- Loftis, A.D.; Mixson, T.R.; Stromdahl, E.Y.; Yabsley, M.J.; Garrison, L.E.; Williamson, P.C.; Fitak, R.R.; Fuerst, P.A.; Kelly, D.J.; Blount, K.W. Geographic Distribution and Genetic Diversity of the Ehrlichia sp. from Panola Mountain in Amblyomma americanum. BMC Infect. Dis. 2008, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Jackson, H.B.; Fahrig, L. What Size Is a Biologically Relevant Landscape? Landsc. Ecol. 2012, 27, 929–941. [Google Scholar] [CrossRef]
- Jackson, H.B.; Fahrig, L. Are Ecologists Conducting Research at the Optimal Scale? Glob. Ecol. Biogeogr. 2015, 24, 52–63. [Google Scholar] [CrossRef]
- Matula, D.W.; Sokal, R.R. Properties of Gabriel Graphs Relevant to Geographic Variation Research and the Clustering of Points in the Plane. Geogr. Anal. 1980, 12, 205–222. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S. In Statistics and Computing; Springer: New York, NY, USA, 2002. [Google Scholar]
- Bivand, R.; Bernat, A.; Carvalho, M.; Chun, Y.; Dormann, C.; Dray, S.; Halbersma, R.; Lewin-Koh, N.; Ma, J.; Millo, G.; et al. The Spdep Package, Version 05–83; The R Project for Statistical Computing: Vienna, Austria, 2013.
- Bartoń, K. MuMIn: Multi-Model Inference, R Package Version 1.43. 17; The R Project for Statistical Computing: Vienna, Austria, 2022.
- Wickham, H. ggplot2; In Use R! Springer: Berlin, Germany, 2016. [Google Scholar]
- Marks Stowe, D.A.; Birkenheuer, A.J.; Grindem, C.B. Pathology in Practice. Intraerythrocytic Infection with Organisms Consistent with a Large Babesia Sp. J. Am. Vet. Med. Assoc. 2012, 241, 1029–1031. [Google Scholar] [CrossRef] [PubMed]
- Wikander, Y.M.; Anantatat, T.; Kang, Q.; Reif, K.E. Prevalence of Cytauxzoon Felis Infection-Carriers in Eastern Kansas Domestic Cats. Pathogens 2020, 9, 854. [Google Scholar] [CrossRef]
- Dahlgren, F.S.; Mandel, E.J.; Krebs, J.W.; Massung, R.F.; McQuiston, J.H. Increasing Incidence of Ehrlichia chaffeensis and Anaplasma phagocytophilum in the United States, 2000–2007. Am. J. Trop. Med. Hyg. 2011, 85, 124–131. [Google Scholar] [CrossRef]
- Qurollo, B.A.; Davenport, A.C.; Sherbert, B.M.; Grindem, C.B.; Birkenheuer, A.J.; Breitschwerdt, E.B. Infection with Panola Mountain Ehrlichia sp. in a Dog with Atypical Lymphocytes and Clonal T-Cell Expansion. J. Vet. Intern. Med. 2013, 27, 1251–1255. [Google Scholar] [CrossRef]
- Reeves, W.K.; Loftis, A.D.; Nicholson, W.L.; Czarkowski, A.G. The First Report of Human Illness Associated with the Panola Mountain Ehrlichia Species: A Case Report. J. Med. Case Rep. 2008, 2, 139. [Google Scholar] [CrossRef] [Green Version]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on Tick-Borne Rickettsioses around the World: A Geographic Approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [Green Version]
- Haus, J.M.; Demchur, J.A.; Dion, J.R.; Habecker, P.L.; Bowman, J.L. Theileriosis in Multiple Neonatal White-Tailed Deer (Odocoileus virginianus) in Delaware, USA. J. Wildl. Dis. 2018, 54, 885–888. [Google Scholar] [CrossRef]
- Scott, J.D.; Sajid, M.S.; Pascoe, E.L.; Foley, J.E. Detection of Babesia odocoilei in Humans with Babesiosis Symptoms. Diagnostics 2021, 11, 947. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, A.; Pastor, A.R.; Berkvens, C.N.; Gara-Boivin, C.; Hébert, M.; Léveillé, A.N.; Barta, J.R.; Smith, D.A. Babesia odocoilei as a Cause of Mortality in Captive Cervids in Canada. Can. Vet. J. 2018, 59, 52–58. [Google Scholar]
- Eisen, L. Tick Species Infesting Humans in the United States. Ticks Tick Borne Dis. 2022, 13, 102025. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Luo, C.-Y.; Ribbe, F.; Pearson, P.; Ledizet, M.; Rich, S.M. Borrelia miyamotoi in Human-Biting Ticks, United States, 2013–2019. Emerg. Infect. Dis. 2021, 27, 3193–3195. [Google Scholar] [CrossRef]
- Keirans, J.E.; Needham, G.R.; Oliver, J.H. The Ixodes ricinus Complex Worldwide: Diagnosis of the Species in the Complex, Hosts and Distribution. In Proceedings of IXth International Congress; Ohio Biological Survey: Columbus, OH, USA, 1999. [Google Scholar]
- Oliver, J.H., Jr.; Lin, T.; Gao, L.; Clark, K.L.; Banks, C.W.; Durden, L.A.; James, A.M.; Chandler, F.W., Jr. An Enzootic Transmission Cycle of Lyme Borreliosis Spirochetes in the Southeastern United States. Proc. Natl. Acad. Sci. USA 2003, 100, 11642–11645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadolny, R.M.; Wright, C.L.; Hynes, W.L.; Sonenshine, D.E.; Gaff, H.D. Ixodes affinis (Acari: Ixodidae) in Southeastern Virginia and Implications for the Spread of Borrelia Burgdorferi, the Agent of Lyme Disease. J. Vector Ecol. 2011, 36, 464–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, P.; Saleh, M.N.; Sundstrom, K.D.; Ientile, M.; Little, S.E. Ixodes Spp. from Dogs and Cats in the United States: Diversity, Seasonality, and Prevalence of Borrelia burgdorferi and Anaplasma phagocytophilum. Vector Borne Zoonotic Dis. 2021, 21, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Paddock, C.D.; Finley, R.W.; Wright, C.S.; Robinson, H.N.; Schrodt, B.J.; Lane, C.C.; Ekenna, O.; Blass, M.A.; Tamminga, C.L.; Ohl, C.A.; et al. Rickettsia parkeri Rickettsiosis and Its Clinical Distinction from Rocky Mountain Spotted Fever. Clin. Infect. Dis. 2008, 47, 1188–1196. [Google Scholar] [CrossRef] [Green Version]
- Whitman, T.J.; Richards, A.L.; Paddock, C.D.; Tamminga, C.L.; Sniezek, P.J.; Jiang, J.; Byers, D.K.; Sanders, J.W. Rickettsia parkeri Infection after Tick Bite, Virginia. Emerg. Infect. Dis. 2007, 13, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Paddock, C.D.; Goddard, J. The Evolving Medical and Veterinary Importance of the Gulf Coast Tick (Acari: Ixodidae). J. Med. Entomol. 2015, 52, 230–252. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.B.; Yabsley, M.J.; Garrison, L.E.; Freye, J.D.; Dunlap, B.G.; Dunn, J.R.; Mead, D.G.; Jones, T.F.; Moncayo, A.C. Rickettsia parkeri in Amblyomma americanum Ticks, Tennessee and Georgia, USA. Emerg. Infect. Dis. 2009, 15, 1471–1473. [Google Scholar] [CrossRef] [PubMed]
- Gaines, D.N.; Operario, D.J.; Stroup, S.; Stromdahl, E.; Wright, C.; Gaff, H.; Broyhill, J.; Smith, J.; Norris, D.E.; Henning, T.; et al. Ehrlichia and Spotted Fever Group Rickettsiae Surveillance in Amblyomma americanum in Virginia through Use of a Novel Six-Plex Real-Time PCR Assay. Vector Borne Zoonotic Dis. 2014, 14, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Wright, C.L.; Sonenshine, D.E.; Gaff, H.D.; Hynes, W.L. Rickettsia parkeri Transmission to Amblyomma americanum by Cofeeding with Amblyomma maculatum (Acari: Ixodidae) and Potential for Spillover. J. Med. Entomol. 2015, 52, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Goddard, J. Experimental Infection of Lone Star Ticks, Amblyomma americanum(L.), with Rickettsia Parkeri and Exposure of Guinea Pigs to the Agent. J. Med. Entomol. 2003, 40, 686–689. [Google Scholar] [CrossRef]
- Cohen, S.B.; Yabsley, M.J.; Freye, J.D.; Dunlap, B.G.; Rowland, M.E.; Huang, J.; Dunn, J.R.; Jones, T.F.; Moncayo, A.C. Prevalence of Ehrlichia chaffeensis and Ehrlichia ewingii in Ticks from Tennessee. Vector Borne Zoonotic Dis. 2010, 10, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Loftis, A.D.; Reeves, W.K.; Spurlock, J.P.; Mahan, S.M.; Troughton, D.R.; Dasch, G.A.; Levin, M.L. Infection of a Goat with a Tick-Transmitted Ehrlichia from Georgia, U.S.A., that Is Closely Related to Ehrlichia ruminantium. J. Vector Ecol. 2006, 31, 213–223. [Google Scholar] [CrossRef]
- Waldrup, K.A.; Kocan, A.A.; Barker, R.W.; Wagner, G.G. Transmission of Babesia odocoilei in White-Tailed Deer (Odocoileus virginianus) by Ixodes scapularis (Acari: Ixodidae). J. Wildl. Dis. 1990, 26, 390–391. [Google Scholar] [CrossRef] [Green Version]
- Steiner, F.E.; Pinger, R.R.; Vann, C.N.; Abley, M.J.; Sullivan, B.; Grindle, N.; Clay, K.; Fuqua, C. Detection of Anaplasma phagocytophilum and Babesia odocoilei DNA in Ixodes scapularis (Acari: Ixodidae) Collected in Indiana. J. Med. Entomol. 2006, 43, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shock, B.C.; Moncayo, A.; Cohen, S.; Mitchell, E.A.; Williamson, P.C.; Lopez, G.; Garrison, L.E.; Yabsley, M.J. Diversity of Piroplasms Detected in Blood-Fed and Questing Ticks from Several States in the United States. Ticks Tick Borne Dis. 2014, 5, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Livengood, J.; Hutchinson, M.L.; Thirumalapura, N.; Tewari, D. Detection of Babesia, Borrelia, Anaplasma, and Rickettsia spp. in Adult Black-Legged Ticks (Ixodes scapularis) from Pennsylvania, United States, with a Luminex Multiplex Bead Assay. Vector Borne Zoonotic Dis. 2020, 20, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Zembsch, T.E.; Bron, G.M.; Paskewitz, S.M. Evidence for Vertical Transmission of Babesia odocoilei (Piroplasmida: Babesiidae) in Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2021, 58, 2484–2487. [Google Scholar] [CrossRef]
- Birkenheuer, A.J.; Le, J.A.; Valenzisi, A.M.; Tucker, M.D.; Levy, M.G.; Breitschwerdt, E.B. Cytauxzoon felis Infection in Cats in the Mid-Atlantic States: 34 Cases (1998–2004). J. Am. Vet. Med. Assoc. 2006, 228, 568–571. [Google Scholar] [CrossRef]
- Köster, L.S.; Lobetti, R.G.; Kelly, P. Canine Babesiosis: A Perspective on Clinical Complications, Biomarkers, and Treatment. Vet. Med. Auckl 2015, 6, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Gallatin, L.L.; Irizarry-Rovira, A.R.; Renninger, M.L.; Holman, P.J.; Wagner, G.G.; Sojka, J.E.; Christian, J.A. Babesia odocoilei Infection in Elk. J. Am. Vet. Med. Assoc. 2003, 223, 1027–1032, 986. [Google Scholar] [CrossRef]
- Bartlett, S.L.; Abou-Madi, N.; Messick, J.B.; Birkenheuer, A.; Kollias, G.V. Diagnosis and Treatment of Babesia odocoilei in Captive Reindeer (Rangifer Tarandus Tarandus) and Recognition of Three Novel Host Species. J. Zoo Wildl. Med. 2009, 40, 152–159. [Google Scholar] [CrossRef]
- Pattullo, K.M.; Wobeser, G.; Lockerbie, B.P.; Burgess, H.J. Babesia odocoilei Infection in a Saskatchewan Elk (Cervus elaphus canadensis) Herd. J. Vet. Diagn. Invest. 2013, 25, 535–540. [Google Scholar] [CrossRef] [Green Version]
- Milnes, E.L.; Thornton, G.L.; Delnatte, P.; Léveillé, A.N.; Barta, J.R.; Smith, D.A.; Nemeth, N.M. Molecular Detection of Babesia odocoilei in Wild, Farmed, and Zoo Cervids in Onatario, Cananda. J. Wildl. Dis. 2019, 55, 335–342. [Google Scholar] [CrossRef]
- Reichard, M.V.; Sanders, T.L.; Weerarathne, P.; Meinkoth, J.H.; Miller, C.A.; Scimeca, R.C.; Almazán, C. Cytauxzoonosis in North America. Pathogens 2021, 10, 1170. [Google Scholar] [CrossRef]
- Allen, K.E.; Thomas, J.E.; Wohltjen, M.L.; Reichard, M.V. Transmission of Cytauxzoon felis to Domestic Cats by Amblyomma americanum Nymphs. Parasit. Vectors 2019, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Shock, B.C.; Murphy, S.M.; Patton, L.L.; Shock, P.M.; Olfenbuttel, C.; Beringer, J.; Prange, S.; Grove, D.M.; Peek, M.; Butfiloski, J.W.; et al. Distribution and Prevalence of Cytauxzoon felis in Bobcats (Lynx rufus), the Natural Reservoir, and Other Wild Felids in Thirteen States. Vet. Parasitol. 2011, 175, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Cauvin, A.; Hood, K.; Shuman, R.; Orange, J.; Blackburn, J.K.; Sayler, K.A.; Wisely, S.M. The Impact of Vector Control on the Prevalence of Theileria cervi in Farmed Florida White-Tailed Deer, Odocoileus virginianus. Parasit. Vectors 2019, 12, 100. [Google Scholar] [CrossRef] [PubMed]
- Moutailler, S.; Valiente Moro, C.; Vaumourin, E.; Michelet, L.; Tran, F.H.; Devillers, E.; Cosson, J.-F.; Gasqui, P.; Van, V.T.; Mavingui, P.; et al. Co-Infection of Ticks: The Rule Rather Than the Exception. PLoS Negl. Trop. Dis. 2016, 10, e0004539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Chamorro, A.; Hodžić, A.; King, K.C.; Cabezas-Cruz, A. Ecological and Evolutionary Perspectives on Tick-Borne Pathogen Co-Infections. Curr. Res. Parasitol. Vector Borne Dis. 2021, 1, 100049. [Google Scholar] [CrossRef]
- Stromdahl, E.Y.; Vince, M.A.; Billingsley, P.M.; Dobbs, N.A.; Williamson, P.C. Rickettsia amblyommii Infecting Amblyomma americanum Larvae. Vector Borne Zoonotic Dis. 2008, 8, 15–24. [Google Scholar] [CrossRef]
- Spielman, A.; Wilson, M.L.; Levine, J.F.; Piesman, J. Ecology of Ixodes dammini-Borne Human Babesiosis and Lyme Disease. Annu. Rev. Entomol. 1985, 30, 439–460. [Google Scholar] [CrossRef]
- Sonenshine, D.E.; Michael Roe, R. Biology of Ticks Volume 2; OUP USA: New York, NY, USA, 2013; ISBN 9780199744060. [Google Scholar]
- Beier, P.; Noss, R.F. Do Habitat Corridors Provide Connectivity? Conserv. Biol. 1998, 12, 1241–1252. [Google Scholar] [CrossRef]
- Kruize, H.; van der Vliet, N.; Staatsen, B.; Bell, R.; Chiabai, A.; Muiños, G.; Higgins, S.; Quiroga, S.; Martinez-Juarez, P.; Aberg Yngwe, M.; et al. Urban Green Space: Creating a Triple Win for Environmental Sustainability, Health, and Health Equity through Behavior Change. Int. J. Environ. Res. Public Health 2019, 16, 4403. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tick Species and Life Stage | Habitat Type | |
|---|---|---|
| Natural (Mean ± SE) | Manicured (Mean ± SE) | |
| Amblyomma americanum adult | 5.82 ± 2.24 | 0.41 ± 0.12 |
| Amblyomma americanum nymph | 8.70 ± 3.47 | 0.94 ± 0.37 |
| Amblyomma maculatum nymph | 0.01 ± 0.01 | 0.01 ± 0.01 |
| Dermacentor variabilis adult | 0.15 ± 0.11 | 0.02 ± 0.01 |
| Haemaphysalis leporispalustris nymph | 0.01 ± 0.01 | 0 |
| Ixodes affinis adult | 0.01 ± 0.01 | 0 |
| Ixodes scapularis adult | 0.73 ± 0.35 | 0.09 ± 0.05 |
| Tick Species and Life Stage | Microorganism | n. Positive/n. Tested (%) | Wilson 95% CI | Gene Tested | Reference(s) to Medical or Veterinary Importance |
|---|---|---|---|---|---|
| Amblyomma americanum adult/nymph | Babesia sp. | 16/1076 (1.5%) | 0.9–2.4% | 18s | Unknown pathogenicity |
| Babesia sp. (Coco) | 7/1076 (0.7%) | 0.3–1.3% | 18s | [57] | |
| Borrelia lonestari | 14/1076 (1.3%) | 0.8–2.2% | flaB | Unknown pathogenicity | |
| Cytauxzoon felis | 1/1076 (0.1%) | 0.0–0.5% | 18s | [58] | |
| Ehrlichia ewingii | 10/1076 (0.9%) | 0.5–1.7% | groEL | [59] | |
| Ehrlichia chaffeensis | 7/1076 (0.6%) | 0.3–1.3% | groEL | [59] | |
| Panola Mountain Ehrlichia sp. | 9/1076 (0.8%) | 0.4–1.6% | groEL; gltA | [60,61] | |
| Hepatozoon sp A. | 19/1076 (1.8%) | 1.1–2.7% | 18s | Unknown pathogenicity | |
| Rickettsia amblyommatis | 583/1076 (54.2%) | 51.2–57.1% | ompA; 17kda | Unknown pathogenicity | |
| Rickettsia parkeri | 3/1076 (0.3%) | 0.1–0.8% | PLA; 17kda | [62] | |
| Theileria cervi | 48/1076 (4.5%) | 3.4–5.9% | 18s | [63] | |
| Amblyomma maculatum nymph | Rickettsia andeanae | 1/2 (50.0%) | 9.5–90.5% | ompA; gltA | Unknown pathogenicity |
| Borrelia spp., Anaplasmataceae spp. Apicomplexa spp. | 0/2 (0.0%) | NA | flaB; ospA; IGS; groEL; 18s | ||
| Dermacentor variabilis adult | Rickettsia rhiphicephali | 2/18 (11.1%) | 3.1–32.8% | ompA; gltA; PLA | Unknown pathogenicity |
| Theileria cervi | 1/18 (5.6%) | 1.0–25.8% | 18s | [63] | |
| Borrelia spp., Anaplasmataceae spp. | 0/18 (0.0%) | NA | flaB; ospA; IGS; groEL | ||
| Haemaphysalis leporispalustris nymph | Rickettsia spp., Borrelia spp., Anaplasmataceae spp., Apicomplexa spp. | 0/1 (0.0%) | NA | ompA; flaB; ospA; IGS; groEL; 18s | |
| Ixodes affinis adult | Rickettsia sp. | 1/1 (100.0%) | 20.6–100.0% | ompA; gltA | Unknown pathogenicity |
| Borrelia spp., Anaplasmataceae spp. Apicomplexa spp. | 0/1 (0.0%) | NA | flaB; ospA; IGS; groEL; 18s | ||
| Ixodes scapularis adult | Babesia odocoilei | 8/89 (9.0%) | 4.6–16.7% | 18s | [64,65] |
| Cryptoplasma (Allocryptoplasma) sp. | 13/89 (14.6%) | 8.7–23.4% | groEL; 16s | Unknown pathogenicity | |
| Hepatozoon sp. B. | 1/89 (1.1%) | 0.2–6.1% | 18s | Unknown pathogenicity | |
| Rickettsia spp. endosymbiont | 28/89 (32.5%) | 22.8–41.7% | ompA | Unknown pathogenicity | |
| Borrelia spp. | 0/89 (0.0%) | NA | flaB, ospA, IGS |
| Microorganism | (n. Positive Pools/Total Pools) % Positives [Wilson 95% CI] | (n. Positive/Total Larvae) Minimum Infection Rate [Wilson 95% CI] |
|---|---|---|
| Borrelia lonestari | (2/21) 9.5% [2.7–28.9%] | (2/940) 0.2% [0.1–0.8%] |
| Rickettsia amblyommatis | (14/21) 66.6% [45.4–82.8%] | (14/940) 1.5% [0.9–2.5%] |
| Rickettsia parkeri | (2/21) 9.5% [2.6–28.9%] | (2/940) 0.2% [0.1–0.8%] |
| Species | Life Stage | n. of Triple Infections | n. of Dual Infection | n. of Single Infections | n. Tested |
|---|---|---|---|---|---|
| Amblyomma americanum | adult | 6 | 40 | 292 | 498 |
| Amblyomma americanum | nymph | 0 | 31 | 266 | 578 |
| Amblyomma. maculatum | nymph | 0 | 0 | 1 | 2 |
| Dermacentor variabilis | adult | 0 | 0 | 3 | 18 |
| Ixodes affinis | adult | 0 | 0 | 1 | 1 |
| Ixodes scapularis | adult | 0 | 6 | 38 | 89 |
| Species | Intercept | Habitat Type | Development Gradient | Pseudo R2 |
|---|---|---|---|---|
| (a) Amblyomma americanum abundance | 2.461 *** (0.377) | 1.238 *** (0.525) | 0.945 *** (0.165) | 62.1 |
| (b) Ixodes scapularis abundance | −0.169 (0.531) | 1.371 ** (0.666) | 0.620 *** (0.216) | 49.0 |
| (c) presence of infected tick | −1.591 (1.198) | 3.835 (2.702) | 1.690 * (0.971) | 54.9 |
| (d) abundance of infected ticks | −1.187 * (0.625) | 1.945 *** (0.688) | 0.608 *** (0.211) | 61.0 |
| (e) pathogen richness | −1.082 ** (0.550) | 1.238 ** (0.600) | 0.493 ** (0.197) | 54.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhosale, C.R.; Wilson, K.N.; Ledger, K.J.; White, Z.S.; Dorleans, R.; De Jesus, C.E.; Wisely, S.M. Ticks and Tick-Borne Pathogens in Recreational Greenspaces in North Central Florida, USA. Microorganisms 2023, 11, 756. https://doi.org/10.3390/microorganisms11030756
Bhosale CR, Wilson KN, Ledger KJ, White ZS, Dorleans R, De Jesus CE, Wisely SM. Ticks and Tick-Borne Pathogens in Recreational Greenspaces in North Central Florida, USA. Microorganisms. 2023; 11(3):756. https://doi.org/10.3390/microorganisms11030756
Chicago/Turabian StyleBhosale, Chanakya R., Kristen N. Wilson, Kimberly J. Ledger, Zoe S. White, Rayann Dorleans, Carrie E. De Jesus, and Samantha M. Wisely. 2023. "Ticks and Tick-Borne Pathogens in Recreational Greenspaces in North Central Florida, USA" Microorganisms 11, no. 3: 756. https://doi.org/10.3390/microorganisms11030756