

Loss of a Functional Mitochondrial Pyruvate Carrier in Komagataella phaffii Does Not Improve Lactic Acid Production from Glycerol in Aerobic Cultivation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
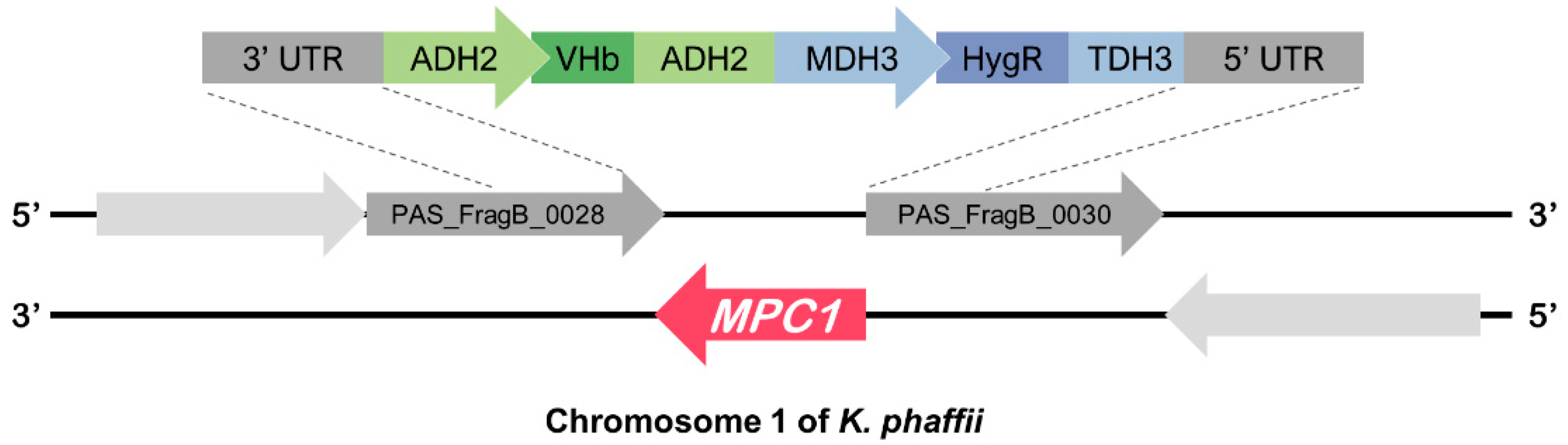
2.1. Construction of the Strains
2.2. Assessment of Growth in Rich, Complex, and Defined Media
2.3. Bioreactor Cultivations
2.3.1. Pre-Inoculum
2.3.2. Cultivation of GLp in the Presence of an Mpc Inhibitor
2.3.3. Small-Scale Batch Cultivation
2.3.4. Batch and Fed-Batch in Medium-Scale Cultivations
2.4. Quantitation of Metabolites on HPLC
2.5. Biomass Determination and Kinetic Parameters of Cultivations
2.6. Quantitative RT-PCR
2.7. Statistical Analysis
3. Results and Discussion
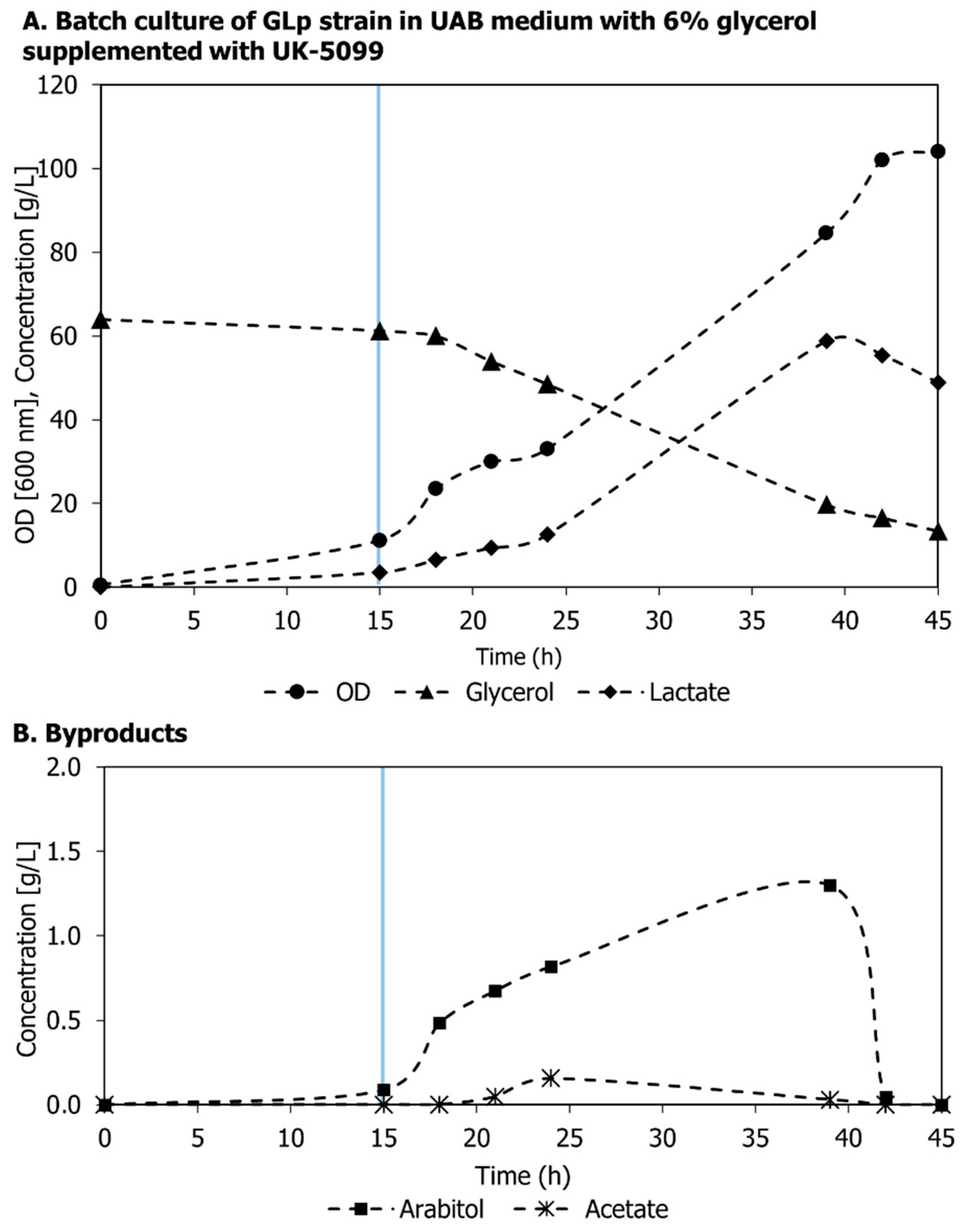
3.1. Effect of the Mpc Inhibitor UK-5099 on Lactic acid Production by GLp
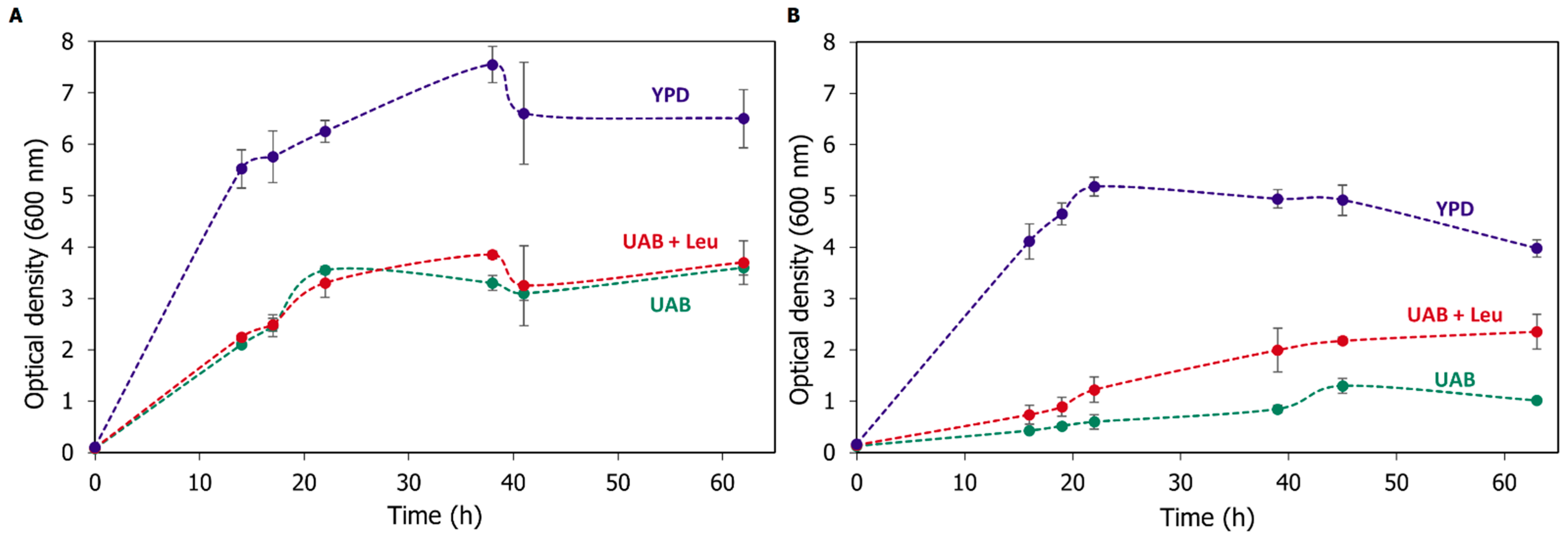
3.2. Auxotrophy to Leucine Indicates the Lack of Mpc1 Activity
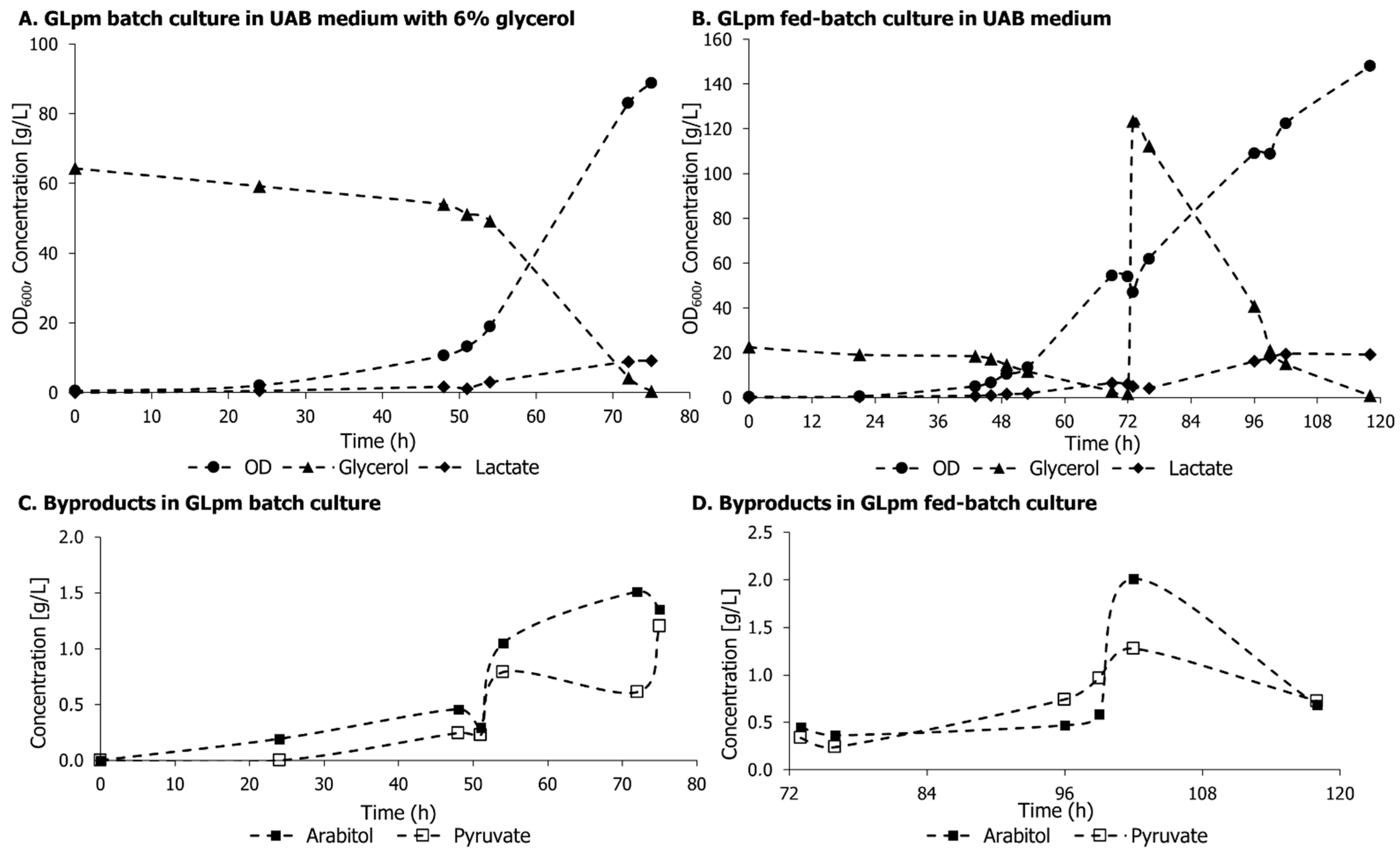
3.3. Lactic Acid Production in GLp and GLpm in Aerobiosis with 4% Glycerol
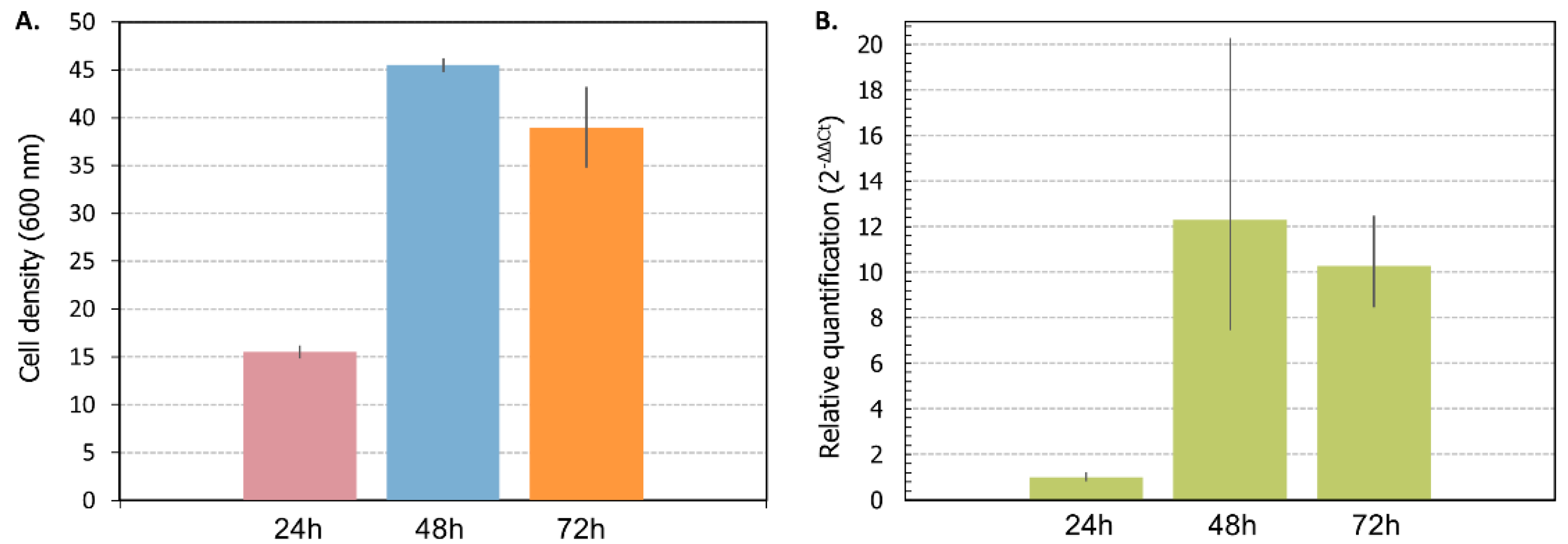
3.4. Relative Expression Level of VHb
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, P.; Anumanthan, A.; Gao, X.-G.; Ilangovan, K.; Suzara, V.V.; Düzgüneş, N.; Renugopalakrishnan, V. Expression of Recombinant Proteins in Pichia Pastoris. Appl. Biochem. Biotechnol. 2007, 142, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Ishigami, M.; Terai, G.; Nakamura, Y.; Hashiba, N.; Nishi, T.; Nakazawa, H.; Hasunuma, T.; Asai, K.; Umetsu, M.; et al. A Streamlined Strain Engineering Workflow with Genome-Wide Screening Detects Enhanced Protein Secretion in Komagataella Phaffii. Commun. Biol. 2022, 5, 561. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sethuraman, N.; Stadheim, T.A.; Zha, D.; Prinz, B.; Ballew, N.; Bobrowicz, P.; Choi, B.-K.; Cook, W.J.; Cukan, M.; et al. Optimization of Humanized IgGs in Glycoengineered Pichia Pastoris. Nat. Biotechnol. 2006, 24, 210–215. [Google Scholar] [CrossRef]
- Kurtzman, C. Description of Komagataella Phaffii Sp. Nov. and the Transfer of Pichia Pseudopastoris to the Methylotrophic Yeast Genus Komagataella. Int. J. Syst. Evol. Microbiol. 2005, 55, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Mattanovich, D.; Graf, A.; Stadlmann, J.; Dragosits, M.; Redl, A.; Maurer, M.; Kleinheinz, M.; Sauer, M.; Altmann, F.; Gasser, B. Genome, Secretome and Glucose Transport Highlight Unique Features of the Protein Production Host Pichia Pastoris. Microb. Cell Factories 2009, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.; Swinnen, S.; Thevelein, J.M.; Nevoigt, E. Glycerol Metabolism and Transport in Yeast and Fungi: Established Knowledge and Ambiguities. Environ. Microbiol. 2017, 19, 878–893. [Google Scholar] [CrossRef] [Green Version]
- Kaur, J.; Sarma, A.K.; Jha, M.K.; Gera, P. Valorisation of Crude Glycerol to Value-Added Products: Perspectives of Process Technology, Economics and Environmental Issues. Biotechnol. Rep. 2020, 27, e00487. [Google Scholar] [CrossRef]
- OECD-FAO Biofuels. In Agricultural Outlook 2022–2031; FAO: Québec City, QC, Canada, 2022.
- Castillo Martinez, F.A.; Balciunas, E.M.; Salgado, J.M.; Domínguez González, J.M.; Converti, A.; de Oliveira, R.P.S. Lactic Acid Properties, Applications and Production: A Review. Trends Food Sci. Technol. 2013, 30, 70–83. [Google Scholar] [CrossRef]
- Grand View Research, Inc. Lactic Acid Market Size, Share & Trends Analysis Report by Raw Material (Sugarcane, Corn, Cassava), By Application (PLA, Food & Beverages), By Region, And Segment Forecasts, 2021–2028; Grand View Research: San Francisco, CA, USA, 2021. [Google Scholar]
- Brzeziński, M.; Biela, T. Stereocomplexed Polylactides. In Encyclopedia of Polymeric Nanomaterials; Kobayashi, S., Müllen, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–10. ISBN 978-3-642-36199-9. [Google Scholar]
- de Lima, P.B.A.; Mulder, K.C.L.; Melo, N.T.M.; Carvalho, L.S.; Menino, G.S.; Mulinari, E.; de Castro, V.H.; dos Reis, T.F.; Goldman, G.H.; Magalhães, B.S.; et al. Novel Homologous Lactate Transporter Improves L-Lactic Acid Production from Glycerol in Recombinant Strains of Pichia Pastoris. Microb. Cell Factories 2016, 15, 158. [Google Scholar] [CrossRef]
- Pacheco, A.; Talaia, G.; Sá-Pessoa, J.; Bessa, D.; Gonçalves, M.J.; Moreira, R.; Paiva, S.; Casal, M.; Queirós, O. Lactic Acid Production in Saccharomyces Cerevisiae Is Modulated by Expression of the Monocarboxylate Transporters Jen1 and Ady2. FEMS Yeast Res. 2012, 12, 375–381. [Google Scholar] [CrossRef]
- Song, J.-Y.; Park, J.-S.; Kang, C.D.; Cho, H.-Y.; Yang, D.; Lee, S.; Cho, K.M. Introduction of a Bacterial Acetyl-CoA Synthesis Pathway Improves Lactic Acid Production in Saccharomyces Cerevisiae. Metab. Eng. 2016, 35, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kang, C.D.; Lee, S.H.; Park, Y.K.; Cho, K.M. Engineering Cellular Redox Balance in Saccharomyces Cerevisiae for Improved Production of L-Lactic Acid. Biotechnol. Bioeng. 2015, 112, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Zhang, B.; Hua, Y.; Zhu, Y.; Li, W.; Wang, D.; Hong, J. Efficient L-Lactic Acid Production from Corncob Residue Using Metabolically Engineered Thermo-Tolerant Yeast. Bioresour. Technol. 2019, 273, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jang, J.H.; Yeo, H.J.; Seol, J.; Kim, S.R.; Jung, Y.H. Lactic Acid Production from a Whole Slurry of Acid-Pretreated Spent Coffee Grounds by Engineered Saccharomyces Cerevisiae. Appl. Biochem. Biotechnol. 2019, 189, 206–216. [Google Scholar] [CrossRef]
- Ilmén, M.; Koivuranta, K.; Ruohonen, L.; Rajgarhia, V.; Suominen, P.; Penttilä, M. Production of L-Lactic Acid by the Yeast Candida Sonorensis Expressing Heterologous Bacterial and Fungal Lactate Dehydrogenases. Microb. Cell Fact. 2013, 12, 53. [Google Scholar] [CrossRef] [Green Version]
- Melo, N.T.M.; Mulder, K.C.L.; Nicola, A.M.; Carvalho, L.S.; Menino, G.S.; Mulinari, E.; Parachin, N.S. Effect of Pyruvate Decarboxylase Knockout on Product Distribution Using Pichia Pastoris (Komagataella Phaffii) Engineered for Lactic Acid Production. Bioengineering 2018, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Bender, T.; Martinou, J.-C. The Mitochondrial Pyruvate Carrier in Health and Disease: To Carry or Not to Carry? Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 2436–2442. [Google Scholar] [CrossRef]
- Papa, S.; Francavilla, A.; Paradies, G.; Meduri, B. The Transport of Pyruvate in Rat Liver Mitochondria. FEBS Lett. 1971, 12, 285–288. [Google Scholar] [CrossRef] [Green Version]
- Bricker, D.K.; Taylor, E.B.; Schell, J.C.; Orsak, T.; Boutron, A.; Chen, Y.-C.; Cox, J.E.; Cardon, C.M.; Van Vranken, J.G.; Dephoure, N.; et al. A Mitochondrial Pyruvate Carrier Required for Pyruvate Uptake in Yeast, Drosophila, and Humans. Science 2012, 337, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Herzig, S.; Raemy, E.; Montessuit, S.; Veuthey, J.-L.; Zamboni, N.; Westermann, B.; Kunji, E.R.S.; Martinou, J.-C. Identification and Functional Expression of the Mitochondrial Pyruvate Carrier. Science 2012, 337, 93–96. [Google Scholar] [CrossRef]
- Bender, T.; Pena, G.; Martinou, J.-C. Regulation of Mitochondrial Pyruvate Uptake by Alternative Pyruvate Carrier Complexes. EMBO J. 2015, 34, 911–924. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wakai, S.; Sasakura, N.; Tsutsumi, H.; Hata, Y.; Ogino, C.; Kondo, A. Pyruvate Metabolism Redirection for Biological Production of Commodity Chemicals in Aerobic Fungus Aspergillus Oryzae. Metab. Eng. 2020, 61, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Melo, N.T.M.; Pontes, G.C.; Procópio, D.P.; de Gois e Cunha, G.C.; Eliodório, K.P.; Paes, H.C.; Basso, T.O.; Parachin, N.S. Evaluation of Product Distribution in Chemostat and Batch Fermentation in Lactic Acid-Producing Komagataella Phaffii Strains Utilizing Glycerol as Substrate. Microorganisms 2020, 8, 781. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hughes, D.E.; Bailey, J.E. Intracellular Expression of Vitreoscilla Hemoglobin Alters the Aerobic Metabolism of Saccharomyces Cerevisiae. Biotechnol. Prog. 1994, 10, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Chien, L.-J.; Lee, C.-K. Expression of Bacterial Hemoglobin in the Yeast, Pichia Pastoris, with a Low O2-Induced Promoter. Biotechnol. Lett. 2005, 27, 1491–1497. [Google Scholar] [CrossRef]
- Prielhofer, R.; Barrero, J.J.; Steuer, S.; Gassler, T.; Zahrl, R.; Baumann, K.; Sauer, M.; Mattanovich, D.; Gasser, B.; Marx, H. GoldenPiCS: A Golden Gate-Derived Modular Cloning System for Applied Synthetic Biology in the Yeast Pichia Pastoris. BMC Syst. Biol. 2017, 11, 123. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Liu, D. Improved Production of Aspergillus Usamii Endo-β-1,4-Xylanase in Pichia Pastoris via Combined Strategies. Biomed. Res. Int. 2016, 2016, 3265895. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Letchworth, G.J. High Efficiency Transformation by Electroporation of Pichia Pastoris Pretreated with Lithium Acetate and Dithiothreitol. BioTechniques 2004, 36, 152–154. [Google Scholar] [CrossRef]
- Halestrap, A.P. The Mitochondrial Pyruvate Carrier. Kinetics and Specificity for Substrates and Inhibitors. Biochem. J. 1975, 148, 85–96. [Google Scholar] [CrossRef] [Green Version]
- McCommis, K.S.; Kovacs, A.; Weinheimer, C.J.; Shew, T.M.; Koves, T.R.; Ilkayeva, O.R.; Kamm, D.R.; Pyles, K.D.; King, M.T.; Veech, R.L.; et al. Nutritional Modulation of Heart Failure in Mitochondrial Pyruvate Carrier–Deficient Mice. Nat. Metab. 2020, 2, 1232–1247. [Google Scholar] [CrossRef]
- Tavoulari, S.; Thangaratnarajah, C.; Mavridou, V.; Harbour, M.E.; Martinou, J.-C.; Kunji, E.R. The Yeast Mitochondrial Pyruvate Carrier Is a Hetero-Dimer in Its Functional State. EMBO J. 2019, 38, e100785. [Google Scholar] [CrossRef] [PubMed]
- Kohlhaw, G.B. Leucine Biosynthesis in Fungi: Entering Metabolism through the Back Door. Microbiol. Mol. Biol. Rev. 2003, 67, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, K.; Matsuda, F.; Okamoto, K.; Ishii, J.; Kondo, A.; Shimizu, H. Repression of Mitochondrial Metabolism for Cytosolic Pyruvate-Derived Chemical Production in Saccharomyces Cerevisiae. Microb. Cell Fact. 2019, 18, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Liu, G.; Chen, Y.; Jiang, S.; Ma, Y.; Zheng, P.; Guo, X.; Xiao, D. Enhanced Production of Ethyl Lactate in Saccharomyces Cerevisiae by Genetic Modification. J. Agric. Food Chem. 2020, 68, 13863–13870. [Google Scholar] [CrossRef]
- Park, S.-H.; Kim, S.; Hahn, J.-S. Improvement of Isobutanol Production in Saccharomyces Cerevisiae by Increasing Mitochondrial Import of Pyruvate through Mitochondrial Pyruvate Carrier. Appl. Microbiol. Biotechnol. 2016, 100, 7591–7598. [Google Scholar] [CrossRef]
- Haarasilta, S.; Taskinen, L. Location of Three Key Enzymes of Gluconeogenesis in Baker’s Yeast. Arch. Microbiol. 1977, 113, 159–161. [Google Scholar] [CrossRef]
- Palmieri, L.; Vozza, A.; Agrimi, G.; Marco, V.D.; Runswick, M.J.; Palmieri, F.; Walker, J.E. Identification of the Yeast Mitochondrial Transporter for Oxaloacetate and Sulfate *. J. Biol. Chem. 1999, 274, 22184–22190. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Chu, J.; Zhang, S.; Zhuang, Y.; Qian, J.; Wang, Y.; Hu, X. Intracellular Expression of Vitreoscilla Hemoglobin Improves S-Adenosylmethionine Production in a Recombinant Pichia Pastoris. Appl. Microbiol. Biotechnol. 2007, 74, 1205–1212. [Google Scholar] [CrossRef]
- Wu, J.-M.; Hsu, T.-A.; Lee, C.-K. Expression of the Gene Coding for Bacterial Hemoglobin Improves Beta-Galactosidase Production in a Recombinant Pichia Pastoris. Biotechnol. Lett. 2003, 25, 1457–1462. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Description | Reference | |
|---|---|---|---|
| Strain | |||
| GLp | GS115: pGAP-LDH Bos taurus Δpdc1 | [19] | |
| GLpm | GLp: VHb V. stercoraria Δmpc1 | This work | |
| Plasmid | |||
| mpc-VHb-HygR | This work | ||
| Primers | Sequence (5′→3′) | ||
| MPC1F | CTCAGATCGATAGAGTGCAAG | MPC1F with MPC1R: 543 bp amplicon in wild type and no amplicon in transformant. RAD9F with MPC1R: amplification occurs only if insertion occurs in the locus. | This work |
| MPC1R | GGAGAAGCTCCATTCGAC | This work | |
| RAD9F | CTCTATGCCTTGAACTATGTCG | This work | |
| qVHbF | CATCTTGCCAGCCGTTAAGAAG | Relative quantification of the heterologous hemoglobin expression. | This work |
| qVHbR | AACAACTCCTGACCGACGATAG | This work | |
| qACT1F | TGTTGGTTGTCCTCGGTTGA | Constitutive control for quantitative PCR. | This work |
| qACT1R | TGAGCTTGGATTCGGCAGAT | This work | |
| Strain | YP | UAB | UAB + Leucine |
|---|---|---|---|
| GLp (parental) | 0.245 ± 0.008 a | 0.207 ± 0.005 b | 0.209 ± 0.008 b |
| GLpm (Δmpc1) | 0.185 ± 0.000 c | 0.074 ± 0.000 d | 0.097 ± 0.001 e |
| Strain | Specific Growth Rate (h−1) | Titer of Lactic Acid (g L−1) | Productivity (g L−1 h−1) | YP/S (g L−1/g L−1) | YX/S (g L−1/g L−1) | Medium |
|---|---|---|---|---|---|---|
| GLp (parental) | 0.308 ± 0.001 | 10.57 ± 0.55 | 0.145 ± 0.025 | 0.230 ± 0.003 | 0.523 ± 0.021 | 4% glycerol YP |
| GLpm (Δmpc1) | 0.165 ± 0.006 | 10.25 ± 0.49 | 0.153 ± 0.007 | 0.278 ± 0.021 | 0.645 ± 0.062 | 4% glycerol YP |
| GLpm | 0.058 ± 0.007 | 9.11 ± 0.72 | 0.122 ± 0.010 | 0.143 ± 0.011 | 0.506 ± 0.009 | 6% glycerol UAB |
| GLpm * | 0.021 ± 0.003 | 19.57 ± 3.07 | 0.163 ± 0.034 | 0.117 ± 0.025 | 0.302 ± 0.030 | 2% glycerol UAB fed−10% glycerol |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira Junqueira, A.C.; Moreira Melo, N.T.; Skorupa Parachin, N.; Costa Paes, H. Loss of a Functional Mitochondrial Pyruvate Carrier in Komagataella phaffii Does Not Improve Lactic Acid Production from Glycerol in Aerobic Cultivation. Microorganisms 2023, 11, 483. https://doi.org/10.3390/microorganisms11020483
de Oliveira Junqueira AC, Moreira Melo NT, Skorupa Parachin N, Costa Paes H. Loss of a Functional Mitochondrial Pyruvate Carrier in Komagataella phaffii Does Not Improve Lactic Acid Production from Glycerol in Aerobic Cultivation. Microorganisms. 2023; 11(2):483. https://doi.org/10.3390/microorganisms11020483
Chicago/Turabian Stylede Oliveira Junqueira, Ana Caroline, Nadielle Tamires Moreira Melo, Nádia Skorupa Parachin, and Hugo Costa Paes. 2023. "Loss of a Functional Mitochondrial Pyruvate Carrier in Komagataella phaffii Does Not Improve Lactic Acid Production from Glycerol in Aerobic Cultivation" Microorganisms 11, no. 2: 483. https://doi.org/10.3390/microorganisms11020483