
The Diversity of Escherichia coli Pathotypes and Vaccination Strategies against This Versatile Bacterial Pathogen


Abstract
:1. Introduction
1.1. Escherichia coli
1.2. Phenomenon of Antibiotic Resistance
2. Diarrheagenic E. coli Pathotypes
2.1. EPEC
2.1.1. Molecular Pathogenesis
2.1.2. Vaccine Strategies against EPEC
2.2. EHEC
2.2.1. Molecular Pathogenesis
2.2.2. Vaccine Strategies against EHEC

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Vaccine | Component of Vaccine | Results/Observations/Outcomes | Animal Model (Year) | References |
|---|---|---|---|---|
| Attenuated bacterial vaccines | Attenuated Salmonella enterica Typhimurium expressing recombinant EspA, intimin and Stx2B. | Significantly higher antibody titers against EspA, intimin and Stx2B, and specific lymphocyte proliferation. | Mice immunized orally (2011). | [142] |
| γ-intimin variant expressed by attenuated Salmonella enterica Typhimurium χ3987 (Δcya,Δcrp,Δasd) and H683 (ΔaroΔasd). | Increased IgG in serum and IgA in feces of mice. Reduced EHEC O157:H7 shedding and colonization post-challenge. | Oral immunization of mice (2012). | [71] | |
| Attenuated EPEC O126:H6. | Reduced mortality in EHEC challenged mouse model and cross-reaction against EspB and intimin EPEC antibodies with EspB and intimin from EHEC. | Mice immunized orally (2016). | [143] | |
| Recombinant bacillus Calmette-Guérin expressing Stx2B (rBCG-Stx2B). | Significant levels of Stx2 IgG in mice. Higher survival rate (>65%) of immunized mice challenged with EHEC. | Mice immunized orally (2012). | [144] | |
| EHEC O157:H7 86-24 strain ΔlerΔstx2 expressing Stx1A Stx2A detoxified. | Lower colonization of EHEC O157:H7 after challenge. | Oral immunization of mice (2009). | [138] | |
| Shiga toxin-based vaccines | cαStx1B and cαStx2A antibodies. | Safety and good tolerance in a human trial single-dose study. | Human volunteers (2009). | [136] |
| one anti-serum albumin VHH and two copies of anti-Stx2B VHH. | Decreased toxicity of EHEC in Stx2 lethal mouse model. | Mice immunized orally (2016). | [137] | |
| Bacterial ghost-based vaccines | Bacterial ghosts of O157:H7 which is unable to cause infection. | Anti-toxicity effect on Vero cell culture. Reduced colonization of EHEC O157:H7 and 93% and 100% survival in orally and rectally immunized mice, respectively. | Orally and rectally immunized mice (2015). | [145] |
| Stx chimeric protein exposing bacterial ghosts of O157:H7 (Stx2Am-Stx1B). | Increased IgG and IgA antibody titers to Stx1A and Stx2B. Survival rate >50% in immunized mice. | Intranasal immunization of mice (2012). | [146] | |
| Peptide-based vaccines | Peptide KT-12 (KASITEIKADKT) conjugated with KLH. | Elevated levels of IgG in subcutaneously immunized mice and IgA in intranasally immunized mice. | Intranasal immunization of mice (2011). | [147] |
| C-terminal region of intimin. | Reduced bacterial adherence to Hep-2 cells and confers protection in immunized mice. | Oral immunization of mice (2011). | [148] | |
| Protein-based vaccines | EspA-Stx1A fusion protein-based vaccine. | Crude toxin Stx2-challenged mice showed 95% survival with high titers of IgG to EspA-Stx1A in treated mice. | Oral immunization of mice (2009). | [149] |
| Stx1B-Stx2-truncated intimin fusion protein. | EHEC O157:H7 challenged immunized mice had a 100% survival rate. | Mice model (2009). | [71] | |
| Plant-based vaccines | Cell line from Nicotiana tabacum (tobacco) NT-1 that expresses inactivated Stx1A. | Stx2-specific IgA in feces of orally immunized mice, and protection against STEC with more than 75% survival rate. | Orally immunized mice model (2018). | [71] |
| Five recombinant EHEC proteins, including NleA, Stx2b, and EspA expressed from Nicotiana benthamiana and transplastomically in Nicotiana tabacum. | Immunized sheep with leaf tissue (feeder) showed less shedding of EHEC O157:H7 when challenged. | Sheep (2018). | [150] | |
| Adjuvant improved vaccines | Adjuvanted EspB and/or C-terminal of γ-intimin protein with MALP-2. | Significantly higher titers of IgA in immunized mice. | Orally immunized mice (2013). | [151] |
| Chimeric Tir-Stx1B-Stx2B adjuvanted with Zot. | Significant increased IgA and IgG and reduced bacterial shedding in feces post- challenge in subcutaneously immunized mice. Partial protection against EHEC. | Subcutaneously immunized mice (2019). | [152] | |
| Polysaccharide-based vaccines | O-specific polysaccharide of EHEC O157:H7 conjugated with recombinant exotoxin A of P. aeruginosa. | Elevated IgG against LPS in vaccinated children with non-collateral reactions to the vaccine. | Human volunteers (2014). | [153] |
| DNA-based vaccines | Stx2AΔAB DNA vaccine. | Immunized mice showed partial protection when challenged with native Stx2. Toxin neutralization is observed in the Vero cell culture. | Intranasally immunized mice (2009). | [71] |
| C-terminal domain of EscC. | Increased IgG in sera and IgA in feces of immunized mice. Reduced bacteria in feces, colon, and cecum post-challenge with EHEC. | Orally immunized mice (2014-2016). | [154,155] | |
| pVAX-efa1 (efa-1′). | Significantly elevated levels of specific mucosal IgA and reduced EHEC colonization post-challenge. | Intranasally immunized mice (2016). | [156] |
2.3. ETEC
2.3.1. Molecular Pathogenesis
2.3.2. Vaccine Strategies against ETEC
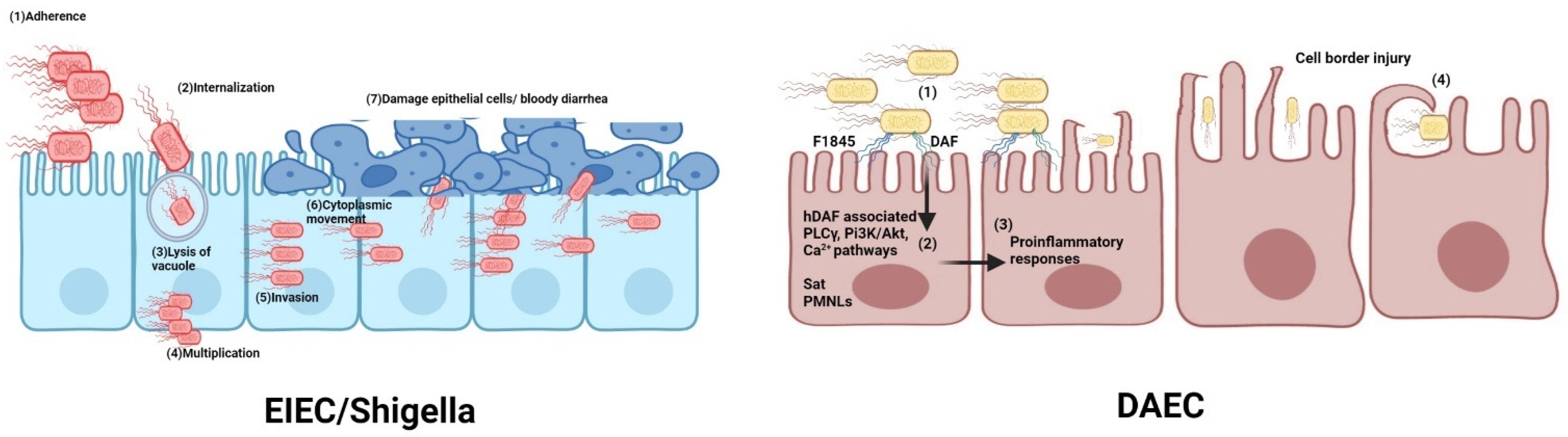
2.4. EIEC
2.4.1. Molecular Pathogenesis
2.4.2. Relationship with Shigella
2.4.3. Vaccine Strategies against EIEC/Shigella
2.5. EAEC and DAEC
2.5.1. Molecular Pathogenesis
2.5.2. Vaccine Strategies against EAEC and DAEC
2.6. AIEC
2.6.1. Molecular Pathogenesis
2.6.2. Relation of AIEC and Crohn’s Disease
2.6.3. Insights for Vaccine Development
3. Extraintestinal Pathogenic E. coli
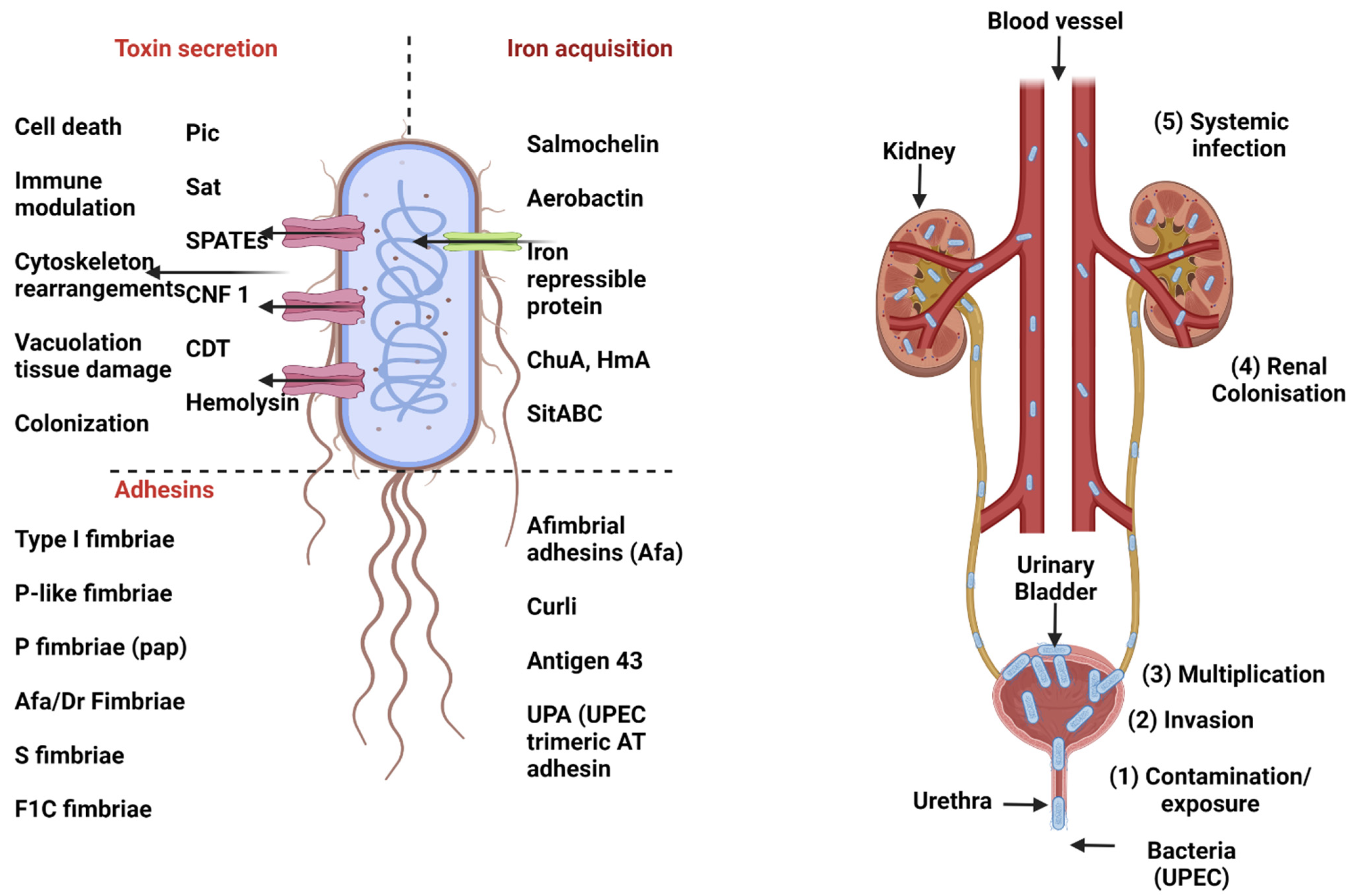
3.1. Uropathogenic E. coli (UPEC)
3.1.1. Urinary Tract Infection (UTI)
3.1.2. Molecular Pathogenesis of UPEC
3.1.3. Vaccine Strategies against UPEC
3.2. MNEC, SEPEC
Vaccine Strategies against MNEC
4. Pathogenic E. coli of Importance to Animal Health
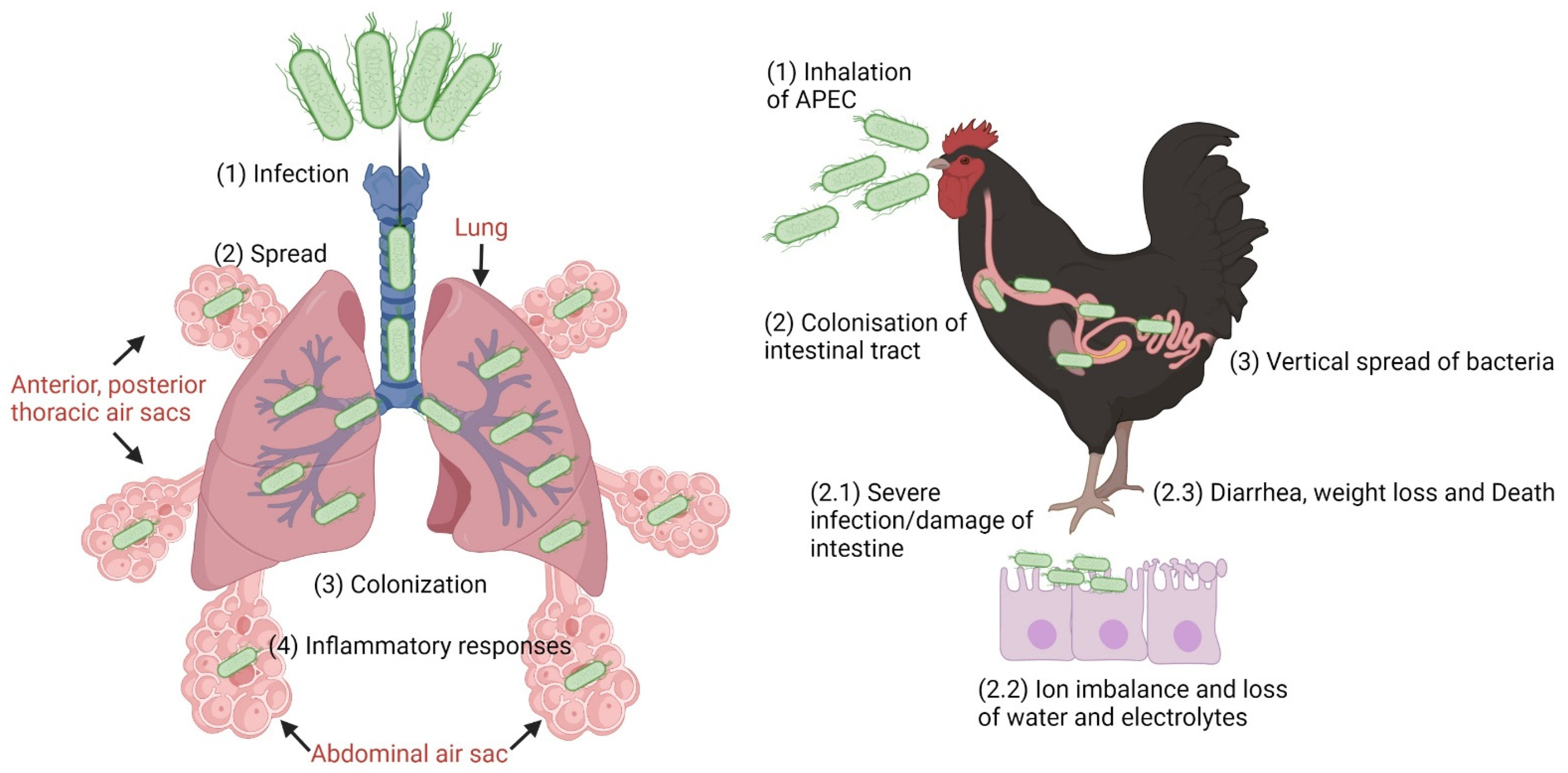
4.1. Avian Pathogenic Escherichia coli (APEC)
4.1.1. APEC Pathogenesis
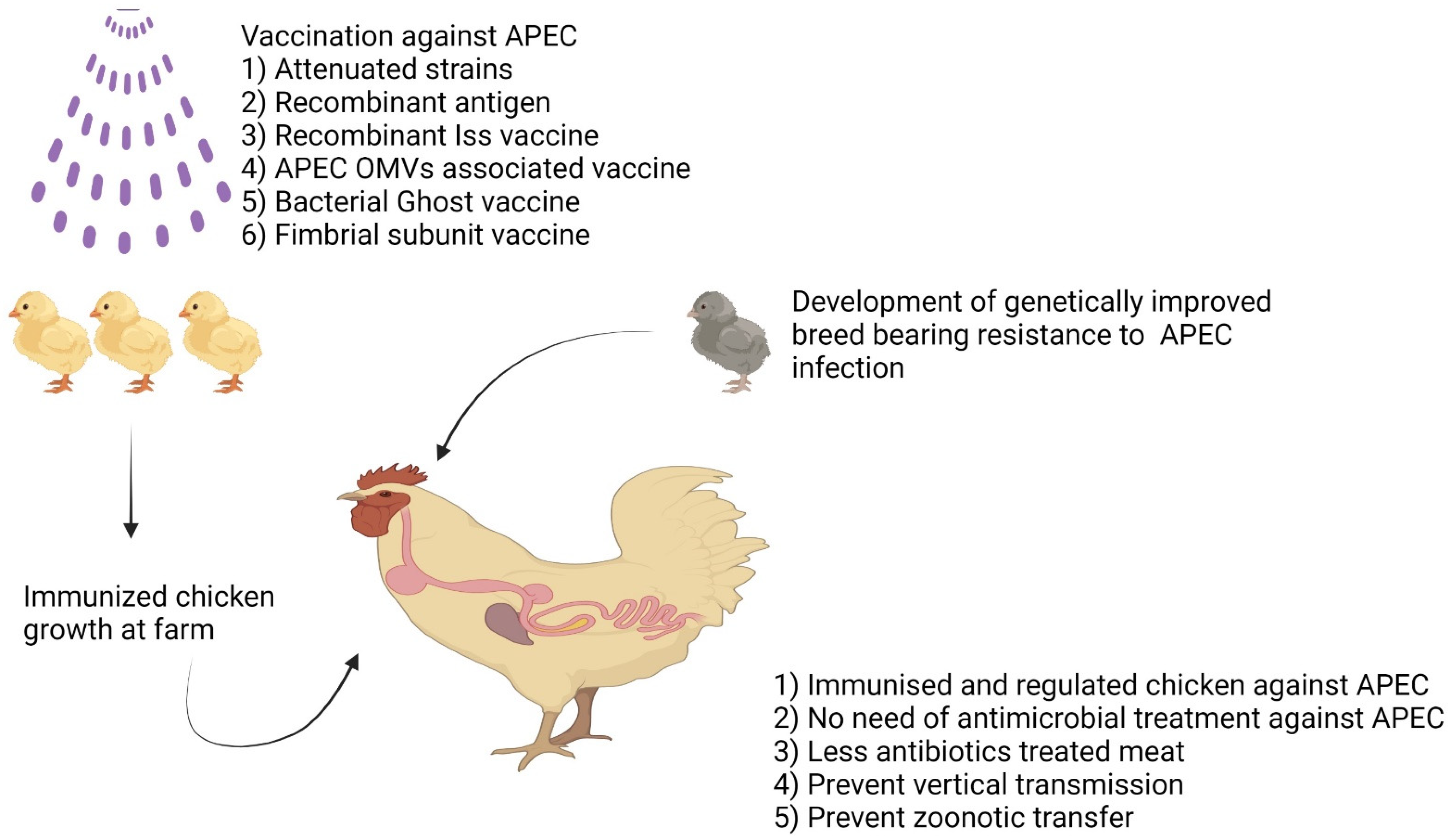
4.1.2. Vaccines and Vaccination Strategies against APEC
4.2. Porcine Colibacillosis
4.2.1. Resistance to Multiple Antibiotics in E. coli Causing Diarrhea in Swine
4.2.2. Need of Vaccines and Vaccine Strategies against Porcine Pathogenic E. coli
4.3. Bovine Colibacillosis
4.4. Mastitis in Cattle and Swine
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blount, Z.D. The natural history of model organisms: The unexhausted potential of E. coli. Elife 2015, 4, e05826. [Google Scholar] [CrossRef]
- Idalia, V.-M.N.; Bernardo, F. Escherichia coli as a model organism and its application in biotechnology. Recent Adv. Physiol. Pathog. Biotechnol. Appl. Tech Open Rij. Croat 2017, 13, 253–274. [Google Scholar]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Leimbach, A.; Hacker, J.; Dobrindt, U. E. coli as an all-rounder: The thin line between commensalism and pathogenicity. Between Pathog. Commensalism 2013, 358, 3–32. [Google Scholar]
- Dale, A.P.; Woodford, N. Extra-intestinal pathogenic Escherichia coli (ExPEC): Disease, carriage and clones. J. Infect. 2015, 71, 615–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The population genetics of pathogenic Escherichia coli. Nat. Rev. Microbiol. 2021, 19, 37–54. [Google Scholar] [CrossRef]
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef]
- Baldy-Chudzik, K.; Bok, E.; Mazurek, J. Well-known and new variants of pathogenic Escherichia coli as a consequence of the plastic genome. Postep. Hig. I Med. Dosw. Online 2015, 69, 345–361. [Google Scholar] [CrossRef]
- Köhler, C.-D.; Dobrindt, U. What defines extraintestinal pathogenic Escherichia coli? Int. J. Med. Microbiol. 2011, 301, 642–647. [Google Scholar] [CrossRef]
- Tivendale, K.A.; Logue, C.M.; Kariyawasam, S.; Jordan, D.; Hussein, A.; Li, G.; Wannemuehler, Y.; Nolan, L.K. Avian-pathogenic Escherichia coli strains are similar to neonatal meningitis E. coli strains and are able to cause meningitis in the rat model of human disease. Infect. Immun. 2010, 78, 3412–3419. [Google Scholar] [CrossRef] [Green Version]
- Chaudhuri, R.R.; Henderson, I.R. The evolution of the Escherichia coli phylogeny. Infect. Genet. Evol. 2012, 12, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Manges, A. Escherichia coli and urinary tract infections: The role of poultry-meat. Clin. Microbiol. Infect. 2016, 22, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, L.; Garenaux, A.; Harel, J.; Boulianne, M.; Nadeau, E.; Dozois, C.M. Escherichia coli from animal reservoirs as a potential source of human extraintestinal pathogenic E. coli. FEMS Immunol. Med. Microbiol. 2011, 62, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, K.; Chaudhari, A.A.; Lee, J.H. Avian colibacillosis caused by an intestinal pathogenic Escherichia coli isolate from calf diarrhea. Res. Vet. Sci. 2010, 89, 150–152. [Google Scholar] [CrossRef]
- Meena, P.R.; Yadav, P.; Hemlata, H.; Tejavath, K.; Singh, A.P. Poultry-origin extraintestinal Escherichia coli strains carrying the traits associated with urinary tract infection, sepsis, meningitis and avian colibacillosis in India. J. Appl. Microbiol. 2021, 130, 2087–2101. [Google Scholar] [CrossRef]
- Spurbeck, R.R.; Dinh, P.C., Jr.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Fakhr, M.K.; Nolan, L.K. Comparison of Escherichia coli isolates implicated in human urinary tract infection and avian colibacillosis. Microbiology 2005, 151, 2097–2110. [Google Scholar] [CrossRef] [Green Version]
- Baumgart, L.A.; Lee, J.E.; Salamov, A.; Dilworth, D.J.; Na, H.; Mingay, M.; Blow, M.J.; Zhang, Y.; Yoshinaga, Y.; Daum, C.G. Persistence and plasticity in bacterial gene regulation. Nat. Methods 2021, 18, 1499–1505. [Google Scholar] [CrossRef]
- Vandecraen, J.; Chandler, M.; Aertsen, A.; Van Houdt, R. The impact of insertion sequences on bacterial genome plasticity and adaptability. Crit. Rev. Microbiol. 2017, 43, 709–730. [Google Scholar] [CrossRef] [Green Version]
- Dobrindt, U.; Hacker, J. Whole genome plasticity in pathogenic bacteria. Curr. Opin. Microbiol. 2001, 4, 550–557. [Google Scholar] [CrossRef]
- Darmon, E.; Leach, D.R. Bacterial genome instability. Microbiol. Mol. Biol. Rev. 2014, 78, 1–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, A.; Kaper, J. Pathogenicity islands of intestinal E. coli. Pathog. Isl. Evol. Pathog. Microbes 2002, 1, 31–48. [Google Scholar] [CrossRef]
- Sabaté, M.; Moreno, E.; Pérez, T.; Andreu, A.; Prats, G. Pathogenicity island markers in commensal and uropathogenic Escherichia coli isolates. Clin. Microbiol. Infect. 2006, 12, 880–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaper, J.B.; Mellies, J.L.; Nataro, J.P. Pathogenicity islands and other mobile genetic elements of diarrheagenic Escherichia coli. Pathog. Isl. Other Mob. Virulence Elem. 1999, 33–58. [Google Scholar] [CrossRef]
- Maurelli, A.T.; Fernández, R.E.; Bloch, C.A.; Rode, C.K.; Fasano, A. “Black holes” and bacterial pathogenicity: A large genomic deletion that enhances the virulence of Shigella spp. and enteroinvasive Escherichia coli. Proc. Natl. Acad. Sci. USA 1998, 95, 3943–3948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, R.; Reeves, P.R. Escherichia coli in disguise: Molecular origins of Shigella. Microbes Infect. 2002, 4, 1125–1132. [Google Scholar] [CrossRef]
- Schmidt, H. Shiga-toxin-converting bacteriophages. Res. Microbiol. 2001, 152, 687–695. [Google Scholar] [CrossRef]
- Beutin, L.; Bode, L.; Ozel, M.; Stephan, R. Enterohemolysin production is associated with a temperate bacteriophage in Escherichia coli serogroup O26 strains. J. Bacteriol. 1990, 172, 6469–6475. [Google Scholar] [CrossRef] [Green Version]
- Boyd, E.F.; Brüssow, H. Common themes among bacteriophage-encoded virulence factors and diversity among the bacteriophages involved. Trends Microbiol. 2002, 10, 521–529. [Google Scholar] [CrossRef]
- Khalil, R.K.; Skinner, C.; Patfield, S.; He, X. Phage-mediated Shiga toxin (Stx) horizontal gene transfer and expression in non-Shiga toxigenic Enterobacter and Escherichia coli strains. Pathog. Dis. 2016, 74, ftw037. [Google Scholar] [CrossRef] [Green Version]
- Javadi, M.; Bouzari, S.; Oloomi, M. Horizontal gene transfer and the diversity of Escherichia coli. In Escherichia coli - Recent Advances on Physiology, Pathogenesis and Biotechnological Applications; IntechOpen: London, UK, 2017; pp. 317–332. [Google Scholar] [CrossRef] [Green Version]
- Messerer, M.; Fischer, W.; Schubert, S. Investigation of horizontal gene transfer of pathogenicity islands in Escherichia coli using next-generation sequencing. PLoS ONE 2017, 12, e0179880. [Google Scholar] [CrossRef] [PubMed]
- King, L.A.; Nogareda, F.; Weill, F.-X.; Mariani-Kurkdjian, P.; Loukiadis, E.; Gault, G.; Jourdan-DaSilva, N.; Bingen, E.; Macé, M.; Thevenot, D. Outbreak of Shiga toxin–producing Escherichia coli O104: H4 associated with organic fenugreek sprouts, France, June 2011. Clin. Infect. Dis. 2012, 54, 1588–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beutin, L.; Martin, A. Outbreak of Shiga toxin–producing Escherichia coli (STEC) O104: H4 infection in Germany causes a paradigm shift with regard to human pathogenicity of STEC strains. J. Food Prot. 2012, 75, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Foley, C.; Harvey, E.; Bidol, S.A.; Henderson, T.; Njord, R.; DeSalvo, T.; Haupt, T.; Mba-Jonas, A.; Bailey, C.; Bopp, C. Outbreak of Escherichia coli O104: H4 infections associated with sprout consumption—Europe and North America, May–July 2011. Morb. Mortal. Wkly. Rep. 2013, 62, 1029. [Google Scholar]
- Gati, N.S.; Middendorf-Bauchart, B.; Bletz, S.; Dobrindt, U.; Mellmann, A. Origin and evolution of hybrid Shiga toxin-producing and uropathogenic Escherichia coli strains of sequence type 141. J. Clin. Microbiol. 2019, 58, e01309. [Google Scholar] [CrossRef]
- Gati, N.S.; Temme, I.J.; Middendorf-Bauchart, B.; Kehl, A.; Dobrindt, U.; Mellmann, A. Comparative phenotypic characterization of hybrid Shiga toxin-producing/uropathogenic Escherichia coli, canonical uropathogenic and Shiga toxin-producing Escherichia coli. Int. J. Med. Microbiol. 2021, 311, 151533. [Google Scholar] [CrossRef]
- Bielaszewska, M.; Schiller, R.; Lammers, L.; Bauwens, A.; Fruth, A.; Middendorf, B.; Schmidt, M.A.; Tarr, P.I.; Dobrindt, U.; Karch, H. Heteropathogenic virulence and phylogeny reveal phased pathogenic metamorphosis in Escherichia coli O2: H6. EMBO Mol. Med. 2014, 6, 347–357. [Google Scholar] [CrossRef]
- Marshall, B.M.; Ochieng, D.J.; Levy, S.B. Commensals: Underappreciated reservoir of antibiotic resistance. Microbe 2009, 4, 231–238. [Google Scholar] [CrossRef]
- Tadesse, D.A.; Zhao, S.; Tong, E.; Ayers, S.; Singh, A.; Bartholomew, M.J.; McDermott, P.F. Antimicrobial drug resistance in Escherichia coli from humans and food animals, United States, 1950–2002. Emerg. Infect. Dis. 2012, 18, 741. [Google Scholar] [CrossRef]
- Chokshi, A.; Sifri, Z.; Cennimo, D.; Horng, H. Global contributors to antibiotic resistance. J. Glob. Infect. Dis. 2019, 11, 36. [Google Scholar]
- Ma, F.; Xu, S.; Tang, Z.; Li, Z.; Zhang, L. Use of antimicrobials in food animals and impact of transmission of antimicrobial resistance on humans. Biosaf. Health 2021, 3, 32–38. [Google Scholar] [CrossRef]
- Gopal Rao, G. Risk factors for the spread of antibiotic-resistant bacteria. Drugs 1998, 55, 323–330. [Google Scholar]
- Sengupta, S.; Chattopadhyay, M.K.; Grossart, H.-P. The multifaceted roles of antibiotics and antibiotic resistance in nature. Front. Microbiol. 2013, 4, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, T.H.; Al-Harmoosh, R.A. Mechanisms of antibiotics resistance in bacteria. Sys. Rev. Pharm. 2020, 11, 817–823. [Google Scholar]
- Johnson, J.R.; Johnston, B.; Clabots, C.; Kuskowski, M.A.; Castanheira, M. Escherichia coli sequence type ST131 as the major cause of serious multidrug-resistant E. coli infections in the United States. Clin. Infect. Dis. 2010, 51, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Naseer, U.; Haldorsen, B.; Tofteland, S.; Hegstad, K.; Scheutz, F.; Simonsen, G.S.; Sundsfjord, A.; Group, N.E.S. Molecular characterization of CTX-M-15-producing clinical isolates of Escherichia coli reveals the spread of multidrug-resistant ST131 (O25: H4) and ST964 (O102: H6) strains in Norway. APMIS 2009, 117, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Ayukekbong, J.A.; Ntemgwa, M.; Atabe, A.N. The threat of antimicrobial resistance in developing countries: Causes and control strategies. Antimicrob. Resist. Infect. Control 2017, 6, 1–8. [Google Scholar] [CrossRef]
- Okeke, I.N.; Lamikanra, A.; Edelman, R. Socioeconomic and behavioral factors leading to acquired bacterial resistance to antibiotics in developing countries. Emerg. Infect. Dis. 1999, 5, 18. [Google Scholar] [CrossRef]
- Hassan, M.M. Scenario of Antibiotic Resistance in Developing Countries. Antimicrob. Resist. A One Health Perspect. 2020, 205–230. [Google Scholar] [CrossRef]
- Iseppi, R.; Di Cerbo, A.; Messi, P.; Sabia, C. Antibiotic resistance and virulence traits in vancomycin-resistant enterococci (Vre) and extended-spectrum β-lactamase/ampc-producing (ESBL/ampc) enterobacteriaceae from humans and pets. Antibiotics 2020, 9, 152. [Google Scholar] [CrossRef] [Green Version]
- Mikhayel, M.; Leclercq, S.O.; Sarkis, D.K.; Doublet, B. Occurrence of the Colistin resistance gene mcr-1 and additional antibiotic resistance genes in ESBL/AmpC-producing Escherichia coli from poultry in Lebanon: A nationwide survey. Microbiol. Spectr. 2021, 9, e00025-21. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P. Carbapenemase-producing Enterobacteriaceae: Overview of a major public health challenge. Med. Mal. Infect. 2014, 44, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeifer, Y.; Schlatterer, K.; Engelmann, E.; Schiller, R.A.; Frangenberg, H.R.; Stiewe, D.; Holfelder, M.; Witte, W.; Nordmann, P.; Poirel, L. Emergence of OXA-48-type carbapenemase-producing Enterobacteriaceae in German hospitals. Antimicrob. Agents Chemother. 2012, 56, 2125–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakthavatchalam, Y.D.; Anandan, S.; Veeraraghavan, B. Laboratory detection and clinical implication of oxacillinase-48 like carbapenemase: The hidden threat. J. Glob. Infect. Dis. 2016, 8, 41. [Google Scholar] [PubMed]
- Dautzenberg, M.; Ossewaarde, J.; De Kraker, M.; Van Der Zee, A.; Van Burgh, S.; De Greeff, S.; Bijlmer, H.; Grundmann, H.; Stuart, J.C.; Fluit, A. Successful control of a hospital-wide outbreak of OXA-48 producing Enterobacteriaceae in the Netherlands, 2009 to 2011. Eurosurveillance 2014, 19, 20723. [Google Scholar] [CrossRef] [PubMed]
- Glupczynski, Y.; Huang, T.-D.; Bouchahrouf, W.; de Castro, R.R.; Bauraing, C.; Gérard, M.; Verbruggen, A.-M.; Deplano, A.; Denis, O.; Bogaerts, P. Rapid emergence and spread of OXA-48-producing carbapenem-resistant Enterobacteriaceae isolates in Belgian hospitals. Int. J. Antimicrob. Agents 2012, 39, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Liapis, E.; Pantel, A.; Robert, J.; Nicolas-Chanoine, M.H.; Cavalié, L.; van der Mee-Marquet, N.; De Champs, C.; Aissa, N.; Eloy, C.; Blanc, V. Molecular epidemiology of OXA-48-producing K lebsiella pneumoniae in F rance. Clin. Microbiol. Infect. 2014, 20, O1121–O1123. [Google Scholar] [CrossRef] [Green Version]
- Pitart, C.; Solé, M.; Roca, I.; Fàbrega, A.; Vila, J.; Marco, F. First outbreak of a plasmid-mediated carbapenem-hydrolyzing OXA-48 β-lactamase in Klebsiella pneumoniae in Spain. Antimicrob. Agents Chemother. 2011, 55, 4398–4401. [Google Scholar] [CrossRef] [Green Version]
- Williamson, D.A.; Sidjabat, H.E.; Freeman, J.T.; Roberts, S.A.; Silvey, A.; Woodhouse, R.; Mowat, E.; Dyet, K.; Paterson, D.L.; Blackmore, T. Identification and molecular characterisation of New Delhi metallo-β-lactamase-1 (NDM-1)-and NDM-6-producing Enterobacteriaceae from New Zealand hospitals. Int. J. Antimicrob. Agents 2012, 39, 529–533. [Google Scholar] [CrossRef]
- Farhat, N.; Khan, A.U. Evolving trends of New Delhi Metallo-betalactamse (NDM) variants: A threat to antimicrobial resistance. Infect. Genet. Evol. 2020, 86, 104588. [Google Scholar] [CrossRef]
- Ahmad, N.; Khalid, S.; Ali, S.M.; Khan, A.U. Occurrence of bla NDM variants among Enterobacteriaceae from a neonatal intensive care unit in a northern India hospital. Front. Microbiol. 2018, 9, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angulo, F.J.; Collignon, P.; Powers, J.H.; Chiller, T.M.; Aidara-Kane, A.; Aarestrup, F.M. World Health Organization ranking of antimicrobials according to their importance in human medicine: A critical step for developing risk management strategies for the use of antimicrobials in food production animals. Clin. Infect. Dis. 2009, 49, 132–141. [Google Scholar]
- Collignon, P.C.; Conly, J.M.; Andremont, A.; McEwen, S.A.; Aidara-Kane, A.; World Health Organization Advisory Group, Bogotá Meeting on Integrated Surveillance of Antimicrobial Resistance (WHO-AGISAR); Agerso, Y.; Andremont, A.; Collignon, P.; Conly, J.; et al. World Health Organization ranking of antimicrobials according to their importance in human medicine: A critical step for developing risk management strategies to control antimicrobial resistance from food animal production. Clin. Infect. Dis. 2016, 63, 1087–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naveen, V.; Siddiq, A.; Chandana, G. A study on drug utilization pattern of cephalosporins in general medicine and surgical inpatient department. Int. J. Curr. Pharm. Res. 2018, 10, 33–36. [Google Scholar]
- Founou, R.C.; Founou, L.L.; Essack, S.Y. Clinical and economic impact of antibiotic resistance in developing countries: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0189621. [Google Scholar] [CrossRef] [Green Version]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277. [Google Scholar]
- Ventola, C.L. Cancer immunotherapy, part 1: Current strategies and agents. Pharm. Ther. 2017, 42, 375. [Google Scholar]
- Dadonaite, B.; Ritchie, H.; Roser, M. Diarrheal Diseases. Our World Data. 2018. Available online: https://ourworldindata.org/diarrheal-diseases (accessed on 26 December 2022).
- Roser, M.; Ritchie, H. Burden of Disease. Our World Data. 2021. Available online: https://ourworldindata.org/burden-of-disease (accessed on 26 December 2022).
- Rojas-Lopez, M.; Monterio, R.; Pizza, M.; Desvaux, M.; Rosini, R. Intestinal pathogenic Escherichia coli: Insights for vaccine development. Front. Microbiol. 2018, 9, 440. [Google Scholar] [CrossRef]
- Kliegman, R.M.; Stanton, B.M.; Geme, J.S. Nelson’s textbook of pediatrics (20th edn.), by R. Kliegman, B. Stanton, J. St. Geme, N. Schor (eds). Pediatr. Radiol. 2017, 47, 1364–1365. [Google Scholar] [CrossRef]
- Donnenberg, M.S.; Kaper, J. Enteropathogenic Escherichia coli. Infect. Immun. 1992, 60, 3953–3961. [Google Scholar] [CrossRef] [Green Version]
- Deborah Chen, H.; Frankel, G. Enteropathogenic Escherichia coli: Unravelling pathogenesis. FEMS Microbiol. Rev. 2005, 29, 83–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandes, R.T.; Elias, W.P.; Vieira, M.A.; Gomes, T.A. An overview of atypical enteropathogenic Escherichia coli. FEMS Microbiol. Lett. 2009, 297, 137–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trabulsi, L.R.; Keller, R.; Gomes, T.A.T. Typical and Atypical Enteropathogenic Escherichia coli. Emerg. Infect. Dis. 2002, 8, 508. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, D.K.; Viswalingam, N.; Meganathan, Y.; Kandaswamy, K. Adherence patterns of Escherichia coli in the intestine and its role in pathogenesis. Med. Microecol. 2020, 5, 100025. [Google Scholar] [CrossRef]
- Donnenberg, M.S.; Kaper, J.B.; Finlay, B.B. Interactions between enteropathogenic Escherichia coli and host epithelial cells. Trends Microbiol. 1997, 5, 109–114. [Google Scholar] [CrossRef]
- Bieber, D.; Ramer, S.W.; Wu, C.-Y.; Murray, W.J.; Tobe, T.; Fernandez, R.; Schoolnik, G.K. Type IV pili, transient bacterial aggregates, and virulence of enteropathogenic Escherichia coli. Science 1998, 280, 2114–2118. [Google Scholar] [CrossRef]
- Zahavi, E.E.; Lieberman, J.A.; Donnenberg, M.S.; Nitzan, M.; Baruch, K.; Rosenshine, I.; Turner, J.R.; Melamed-Book, N.; Feinstein, N.; Zlotkin-Rivkin, E. Bundle-forming pilus retraction enhances enteropathogenic Escherichia coli infectivity. Mol. Biol. Cell 2011, 22, 2436–2447. [Google Scholar] [CrossRef] [PubMed]
- Girón, J.A.; Donnenberg, M.S.; Martin, W.C.; Jarvis, K.G.; Kaper, J.B. Distribution of the bundle-forming pilus structural gene (bfpA) among enteropathogenic Escherichia coli. J. Infect. Dis. 1993, 168, 1037–1041. [Google Scholar] [CrossRef]
- Giron, J.A.; Ho, A.S.Y.; Schoolnik, G.K. An inducible bundle-forming pilus of enteropathogenic Escherichia coli. Science 1991, 254, 710–713. [Google Scholar] [CrossRef]
- Vasconcellos, H.L.F.; da Silva, W.D.; Nascimento, I.P.; Kipnis, A. Vaccine Against Entheropathogenic E. coli: A Systematic Review. Int. J. Vaccine Res. 2017, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Vallance, B.; Finlay, B. Exploitation of host cells by enteropathogenic Escherichia coli. Proc. Natl. Acad. Sci. USA 2000, 97, 8799–8806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ide, T.; Laarmann, S.; Greune, L.; Schillers, H.; Oberleithner, H.; Schmidt, M.A. Characterization of translocation pores inserted into plasma membranes by type III-secreted Esp proteins of enteropathogenic Escherichia coli. Cell. Microbiol. 2001, 3, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Gruenheid, S.; DeVinney, R.; Bladt, F.; Goosney, D.; Gelkop, S.; Gish, G.D.; Pawson, T.; Finlay, B.B. Enteropathogenic E. coli Tir binds Nck to initiate actin pedestal formation in host cells. Nat. Cell Biol. 2001, 3, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Goosney, D.L.; Gruenheid, S.; Finlay, B.B. Gut feelings: Enteropathogenic E. coli (EPEC) interactions with the host. Annu. Rev. Cell Dev. Biol. 2000, 16, 173–189. [Google Scholar] [CrossRef]
- Goosney, D.L.; de Grado, M.; Finlay, B.B. Putting E. coli on a pedestal: A unique system to study signal transduction and the actin cytoskeleton. Trends Cell Biol. 1999, 9, 11–14. [Google Scholar] [CrossRef]
- Guignot, J.; Segura, A.; Tran Van Nhieu, G. The serine protease EspC from enteropathogenic Escherichia coli regulates pore formation and cytotoxicity mediated by the type III secretion system. PLoS Pathog. 2015, 11, e1005013. [Google Scholar] [CrossRef] [Green Version]
- Serapio-Palacios, A.; Navarro-Garcia, F. EspC, an autotransporter protein secreted by enteropathogenic Escherichia coli, causes apoptosis and necrosis through caspase and calpain activation, including direct procaspase-3 cleavage. MBio 2016, 7, e00479-16. [Google Scholar] [CrossRef] [Green Version]
- Vasconcellos, H.L.F.; Scaramuzzi, K.; Nascimento, I.P.; Ferreira, J.M.D.C., Jr.; Abe, C.M.; Piazza, R.M.; Kipnis, A.; da Silva, W.D. Generation of recombinant bacillus Calmette–Guérin and Mycobacterium smegmatis expressing BfpA and intimin as vaccine vectors against enteropathogenic Escherichia coli. Vaccine 2012, 30, 5999–6005. [Google Scholar] [CrossRef] [Green Version]
- Mare, A.D.; Ciurea, C.N.; Man, A.; Tudor, B.; Moldovan, V.; Decean, L.; Toma, F. Enteropathogenic Escherichia coli—A summary of the literature. Gastroenterol. Insights 2021, 12, 28–40. [Google Scholar] [CrossRef]
- Loureiro, I.; Frankel, G.; Adu-Bobie, J.; Dougan, G.; Trabulsi, L.R.; Carneiro-Sampaio, M.M. Human Colostrum Contains IgA Antibodies Reactive to EnteropathogenicEscherichia coli Virulence-Associated Proteins: Intimin, BfpA, EspA, and EspB. J. Pediatr. Gastroenterol. Nutr. 1998, 27, 166–171. [Google Scholar] [CrossRef]
- Ellis, R.W.; Brodeur, B.R. New Bacterial Vaccines; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003; pp. 40–63. [Google Scholar]
- Flores, V.M.Q.; de Souza Fernandes, R.C.C.; de Macedo, Z.S.; Medina-Acosta, E. Expression and purification of the recombinant enteropathogenic Escherichia coli vaccine candidates BfpA and EspB. Protein Expr. Purif. 2002, 25, 16–22. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, T.N.; Mitchell, M.C.; Corbishley, A.; Nath, M.; Simmonds, H.; McAteer, S.P.; Mahajan, A.; Low, J.C.; Smith, D.G.; Huntley, J.F. Optimizing the protection of cattle against Escherichia coli O157: H7 colonization through immunization with different combinations of H7 flagellin, Tir, intimin-531 or EspA. PLoS ONE 2015, 10, e0128391. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, T.N.; Mitchell, M.C.; Rosser, T.; McAteer, S.; Low, J.C.; Smith, D.G.; Huntley, J.F.; Mahajan, A.; Gally, D.L. Immunization of cattle with a combination of purified intimin-531, EspA and Tir significantly reduces shedding of Escherichia coli O157: H7 following oral challenge. Vaccine 2010, 28, 1422–1428. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.C.; Campos, I.B.; Abe, C.M.; Trabulsi, L.R.; Elias, W.P.; Ho, P.L.; Oliveira, M.L.S. Immunization of mice with Lactobacillus casei expressing intimin fragments produces antibodies able to inhibit the adhesion of enteropathogenic Escherichia coli to cultivated epithelial cells. FEMS Immunol. Med. Microbiol. 2008, 54, 245–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, J.V.; Garcia, A.B.; Flores, V.M.Q.; de Macedo, Z.S.; Medina-Acosta, E. Phytosecretion of enteropathogenic Escherichia coli pilin subunit A in transgenic tobacco and its suitability for early life vaccinology. Vaccine 2002, 20, 2091–2101. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xia, X.; Liu, Y.; Wan, F. Oral Administration with Live Attenuated Citrobacter rodentium Protects Immunocompromised Mice from Lethal Infection. Infect. Immun. 2022, 90, e00198-22. [Google Scholar] [CrossRef] [PubMed]
- Gohar, A.; Abdeltawab, N.F.; Fahmy, A.; Amin, M.A. Development of safe, effective and immunogenic vaccine candidate for diarrheagenic Escherichia coli main pathotypes in a mouse model. BMC Res. Notes 2016, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Welinder-Olsson, C.; Kaijser, B. Enterohemorrhagic Escherichia coli (EHEC). Scand. J. Infect. Dis. 2005, 37, 405–416. [Google Scholar] [CrossRef]
- Karpman, D.; Ståhl, A.l. Enterohemorrhagic Escherichia coli pathogenesis and the host response. In Enterohemorrhagic Escherichia coli and Other Shiga Toxin-Producing E. coli; Wiley: Hoboken, NJ, USA, 2015; pp. 381–402. [Google Scholar] [CrossRef]
- Scheiring, J.; Andreoli, S.P.; Zimmerhackl, L.B. Treatment and outcome of Shiga-toxin-associated hemolytic uremic syndrome (HUS). Pediatr. Nephrol. 2008, 23, 1749–1760. [Google Scholar] [CrossRef] [Green Version]
- Frankel, G.; Phillips, A.D.; Rosenshine, I.; Dougan, G.; Kaper, J.B.; Knutton, S. Enteropathogenic and enterohaemorrhagic Escherichia coli: More subversive elements. Mol. Microbiol. 1998, 30, 911–921. [Google Scholar] [CrossRef]
- Bavaro, M.F.E. E. coli O157: H7 and other toxigenic strains: The curse of global food distribution. Curr. Gastroenterol. Rep. 2012, 14, 317–323. [Google Scholar] [CrossRef] [PubMed]
- EA, G.G. Animal health and foodborne pathogens: Enterohaemorrhagic O157: H7 strains and other pathogenic Escherichia coli virotypes (EPEC, ETEC, EIEC, EHEC). Pol. J. Vet. Sci. 2002, 5, 103–115. [Google Scholar]
- Meng, J.; LeJeune, J.T.; Zhao, T.; Doyle, M.P. Enterohemorrhagic Escherichia coli. Food Microbiol. Fundam. Front. 2012, 287–309. [Google Scholar] [CrossRef]
- Smith, K.E.; Wilker, P.R.; Reiter, P.L.; Hedican, E.B.; Bender, J.B.; Hedberg, C.W. Antibiotic treatment of Escherichia coli O157 infection and the risk of hemolytic uremic syndrome, Minnesota. Pediatr. Infect. Dis. J. 2012, 31, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar] [CrossRef]
- Manitz, J.; Kneib, T.; Schlather, M.; Helbing, D.; Brockmann, D. Origin detection during food-borne disease outbreaks—A case study of the 2011 EHEC/HUS outbreak in Germany. PLoS Curr. 2014, 2014, 6. [Google Scholar] [CrossRef]
- Braeye, T.; Denayer, S.; De Rauw, K.; Forier, A.; Verluyten, J.; Fourie, L.; Dierick, K.; Botteldoorn, N.; Quoilin, S.; Cosse, P. Lessons learned from a textbook outbreak: EHEC-O157: H7 infections associated with the consumption of raw meat products, June 2012, Limburg, Belgium. Arch. Public Health 2014, 72, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, R.; Gorjão, S.; Seibt, B.; Lima, L.; Barnett, J.; Moss, A.; Wills, J. Tweeting during food crises: A psychosocial analysis of threat coping expressions in Spain, during the 2011 European EHEC outbreak. Int. J. Hum.-Comput. Stud. 2014, 72, 239–254. [Google Scholar] [CrossRef] [Green Version]
- Kanayama, A.; Yahata, Y.; Arima, Y.; Takahashi, T.; Saitoh, T.; Kanou, K.; Kawabata, K.; Sunagawa, T.; Matsui, T.; Oishi, K. Enterohemorrhagic Escherichia coli outbreaks related to childcare facilities in Japan, 2010–2013. BMC Infect. Dis. 2015, 15, 539. [Google Scholar] [CrossRef]
- Meagher, K.D. Policy responses to foodborne disease outbreaks in the United States and Germany. Agric. Hum. Values 2022, 39, 233–248. [Google Scholar] [CrossRef]
- Scharff, R.L. Economic burden from health losses due to foodborne illness in the United States. J. Food Prot. 2012, 75, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.M.; Sreedhar, S.S.; Mickell, J.J.; Chan, J.C. Acute kidney failure: A pediatric experience over 20 years. Arch. Pediatr. Adolesc. Med. 2002, 156, 893–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trachtman, H. HUS and TTP in children. Pediatr. Clin. 2013, 60, 1513–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prager, R.; Lang, C.; Aurass, P.; Fruth, A.; Tietze, E.; Flieger, A. Two novel EHEC/EAEC hybrid strains isolated from human infections. PLoS ONE 2014, 9, e95379. [Google Scholar] [CrossRef]
- Santos, A.C.D.M.; Santos, F.F.; Silva, R.M.; Gomes, T.A.T. Diversity of hybrid-and hetero-pathogenic Escherichia coli and their potential implication in more severe diseases. Front. Cell Infect. Microbiol. 2020, 10, 339. [Google Scholar] [CrossRef]
- Nguyen, Y.; Sperandio, V. Enterohemorrhagic E. coli (EHEC) pathogenesis. Front. Cell. Infect. Microbiol. 2012, 2, 90. [Google Scholar] [CrossRef] [Green Version]
- Tesh, V.; O’brien, A. The pathogenic mechanisms of Shiga toxin and the Shiga-like toxins. Mol. Microbiol. 1991, 5, 1817–1822. [Google Scholar] [CrossRef]
- Karch, H. The role of virulence factors in enterohemorrhagic Escherichia coli (EHEC)-associated hemolytic-uremic syndrome. Semin. Thromb. Hemost. 2001, 27, 207–214. [Google Scholar] [CrossRef]
- Xicohtencatl-Cortes, J.; Saldaña, Z.; Deng, W.; Castañeda, E.; Freer, E.; Tarr, P.I.; Finlay, B.B.; Puente, J.L.; Girón, J.A. Bacterial macroscopic rope-like fibers with cytopathic and adhesive properties. J. Biol. Chem. 2010, 285, 32336–32342. [Google Scholar] [CrossRef]
- Dutta, P.R.; Cappello, R.; Navarro-García, F.; Nataro, J.P. Functional comparison of serine protease autotransporters of Enterobacteriaceae. Infect. Immun. 2002, 70, 7105–7113. [Google Scholar] [CrossRef] [Green Version]
- Tse, C.M.; In, J.G.; Yin, J.; Donowitz, M.; Doucet, M.; Foulke-Abel, J.; Ruiz-Perez, F.; Nataro, J.P.; Zachos, N.C.; Kaper, J.B. Enterohemorrhagic E. coli (EHEC)—Secreted serine protease EspP stimulates electrogenic ion transport in human colonoid monolayers. Toxins 2018, 10, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, A.; Brockmeyer, J. Prevalence, biogenesis, and functionality of the serine protease autotransporter EspP. Toxins 2012, 5, 25–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, H.; Fairbrother, J.M.; Désautels, C.; Mabrouk, T.; Dugourd, D.; Dezfulian, H.; Harel, J. Presence of the LEE (locus of enterocyte effacement) in pig attaching and effacing Escherichia coli and characterization of eae, espA, espB and espD genes of PEPEC (pig EPEC) strain 1390. Microb. Pathog. 2000, 28, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Dean-Nystrom, E.A.; Bosworth, B.T.; Cray, W.C., Jr.; Moon, H.W. Pathogenicity of Escherichia coli O157: H7 in the intestines of neonatal calves. Infect. Immun. 1997, 65, 1842–1848. [Google Scholar] [CrossRef] [Green Version]
- Tzipori, S.; Gunzer, F.; Donnenberg, M.S.; de Montigny, L.; Kaper, J.B.; Donohue-Rolfe, A. The role of the eaeA gene in diarrhea and neurological complications in a gnotobiotic piglet model of enterohemorrhagic Escherichia coli infection. Infect. Immun. 1995, 63, 3621–3627. [Google Scholar] [CrossRef] [Green Version]
- Dean-Nystrom, E.A.; Pohlenz, J.F.; Moon, H.W.; O’Brien, A.D. Escherichia coli O157: H7 causes more-severe systemic disease in suckling piglets than in colostrum-deprived neonatal piglets. Infect. Immun. 2000, 68, 2356–2358. [Google Scholar] [CrossRef] [Green Version]
- Karpman, D.; Connell, H.; Svensson, M.; Scheutz, F.; Aim, P.; Svanborg, C. The role of lipopolysaccharide and Shiga-like toxin in a mouse model of Escherichia coli O157: H7 infection. J. Infect. Dis. 1997, 175, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, H.; Takahashi, M.; Yamaguchi, H.; Osaki, T.; Komatsu, A.; Fujioka, Y.; Kamiya, S. Experimental infection of germ-free mice with hyper-toxigenic enterohaemorrhagic Escherichia coli O157: H7, strain 6. J. Med. Microbiol. 2002, 51, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Conlan, J.W.; Perry, M.B. Susceptibility of three strains of conventional adult mice to intestinal colonization by an isolate of Escherichia coli O157: H7. Can. J. Microbiol. 1998, 44, 800–805. [Google Scholar] [CrossRef]
- Woods, J.B.; Schmitt, C.K.; Darnell, S.C.; Meysick, K.C.; O’Brien, A.D. Ferrets as a model system for renal disease secondary to intestinal infection with Escherichia coli O157: H7 and other Shiga toxin-producing E. coli. J. Infect. Dis. 2002, 185, 550–554. [Google Scholar] [CrossRef] [Green Version]
- Bitzan, M.; Poole, R.; Mehran, M.; Sicard, E.; Brockus, C.; Thuning-Roberson, C.; Rivière, M. Safety and pharmacokinetics of chimeric anti-Shiga toxin 1 and anti-Shiga toxin 2 monoclonal antibodies in healthy volunteers. Antimicrob. Agents Chemother. 2009, 53, 3081–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejías, M.P.; Hiriart, Y.; Lauché, C.; Fernández-Brando, R.J.; Pardo, R.; Bruballa, A.; Ramos, M.V.; Goldbaum, F.A.; Palermo, M.S.; Zylberman, V. Development of camelid single chain antibodies against Shiga toxin type 2 (Stx2) with therapeutic potential against Hemolytic Uremic Syndrome (HUS). Sci. Rep. 2016, 6, 24913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Sun, Y.; Feng, S.; Zhu, L.; Guo, X.; Qi, C. Towards an attenuated enterohemorrhagic Escherichia coli O157: H7 vaccine characterized by a deleted ler gene and containing apathogenic Shiga toxins. Vaccine 2009, 27, 5929–5935. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.A. LEEways: Tales of EPEC, ATEC and EHEC. Cell Microbiol. 2010, 12, 1544–1552. [Google Scholar] [CrossRef]
- Calderon Toledo, C.; Arvidsson, I.; Karpman, D. Cross-reactive protection against enterohemorrhagic Escherichia coli infection by enteropathogenic E. coli in a mouse model. Infect. Immun. 2011, 79, 2224–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrine, M.; Matambisso, G.; Nobela, N.; Vubil, D.; Massora, S.; Acácio, S.; Nhampossa, T.; Alonso, P.; Mandomando, I. Low frequency of enterohemorrhagic, enteroinvasive and diffusely adherent Escherichia coli in children under 5 years in rural Mozambique: A case-control study. BMC Infect. Dis. 2020, 20, 659. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Ning, Y.; Wang, H.; Xiao, D.; Tang, B.; Luo, P.; Cheng, Y.; Jiang, M.; Li, N.; Zou, Q. Vaccination of attenuated EIS-producing Salmonella induces protective immunity against enterohemorrhagic Escherichia coli in mice. Vaccine 2011, 29, 7395–7403. [Google Scholar] [CrossRef]
- Cepeda Molero, M.E. Generation of Enteropathogenic E. coli Strains Lacking the Repertoire of Effectors Translocated by the Type III Protein Secretion System and Their Characterization in the Infection of Cultured Cell Lines and Human Intestinal Biopsies. 2016. Available online: https://dialnet.unirioja.es/servlet/dctes?codigo=67208 (accessed on 15 December 2022).
- Fujii, J.; Naito, M.; Yutsudo, T.; Matsumoto, S.; Heatherly, D.P.; Yamada, T.; Kobayashi, H.; Yoshida, S.-i.; Obrig, T. Protection by a recombinant Mycobacterium bovis Bacillus Calmette-Guerin vaccine expressing Shiga toxin 2 B subunit against Shiga toxin-producing Escherichia coli in mice. Clin. Vaccine Immunol. 2012, 19, 1932–1937. [Google Scholar] [CrossRef] [Green Version]
- Cai, K.; Tu, W.; Liu, Y.; Li, T.; Wang, H. Novel fusion antigen displayed-bacterial ghosts vaccine candidate against infection of Escherichia coli O157: H7. Sci. Rep. 2015, 5, 17479. [Google Scholar] [CrossRef]
- Mayr, U.B.; Kudela, P.; Atrasheuskaya, A.; Bukin, E.; Ignatyev, G.; Lubitz, W. Rectal single dose immunization of mice with Escherichia coli O157: H7 bacterial ghosts induces efficient humoral and cellular immune responses and protects against the lethal heterologous challenge. Microb. Biotechnol. 2012, 5, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-h.; He, K.-w.; Zhang, S.-x.; Lu, W.-c.; Zhao, P.-d.; Luan, X.-t.; Ye, Q.; Wen, L.-b.; Li, B.; Guo, R.-l. Subcutaneous and intranasal immunization with Stx2B–Tir–Stx1B–Zot reduces colonization and shedding of Escherichia coli O157: H7 in mice. Vaccine 2011, 29, 3923–3929. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.s.; Zhou, Y.; Yu, Y.; Peng, L.J.; Zhao, W.; Zheng, X.L. B-cell epitope KT-12 of enterohemorrhagic Escherichia coli O157: H7: A novel peptide vaccine candidate. Microbiol. Immunol. 2011, 55, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Feng, Y.; Luo, P.; Gu, J.; Yu, S.; Zhang, W.-J.; Liu, Y.-Q.; Wang, Q.-X.; Zou, Q.-M.; Mao, X.-H. Fusion expression and immunogenicity of EHEC EspA-Stx2Al protein: Implications for the vaccine development. J. Microbiol. 2009, 47, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Mendoza, S.; Nieto-Gómez, R. Green therapeutic biocapsules: Using plant cells to orally deliver biopharmaceuticals. Trends Biotechnol. 2018, 36, 1054–1067. [Google Scholar] [CrossRef]
- Garcia-Angulo, V.A.; Kalita, A.; Torres, A.G. Advances in the development of enterohemorrhagic Escherichia coli vaccines using murine models of infection. Vaccine 2013, 31, 3229–3235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanifar, J.; Salmanian, A.H.; Haji Hosseini, R.; Amani, J.; Kazemi, R. Chitosan nano-structure loaded with recombinant E. coli O157: H7 antigens as a vaccine candidate can effectively increase immunization capacity. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2593–2604. [Google Scholar] [CrossRef]
- Szu, S.C.; Ahmed, A. Clinical studies of Escherichia coli O157: H7 conjugate vaccines in adults and young children. Microbiol. Spectr. 2014, 2, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Tapia, D.; Ross, B.N.; Kalita, A.; Kalita, M.; Hatcher, C.L.; Muruato, L.A.; Torres, A.G. From in silico protein epitope density prediction to testing Escherichia coli O157: H7 vaccine candidates in a murine model of colonization. Front. Cell Infect. Microbiol. 2016, 6, 94. [Google Scholar] [CrossRef] [Green Version]
- García-Angulo, V.A.; Kalita, A.; Kalita, M.; Lozano, L.; Torres, A.G. Comparative genomics and immunoinformatics approach for the identification of vaccine candidates for enterohemorrhagic Escherichia coli O157: H7. Infect. Immun. 2014, 82, 2016–2026. [Google Scholar] [CrossRef]
- Riquelme-Neira, R.; Rivera, A.; Sáez, D.; Fernández, P.; Osorio, G.; del Canto, F.; Salazar, J.C.; Vidal, R.M.; Oñate, A. Vaccination with DNA encoding truncated enterohemorrhagic Escherichia coli (EHEC) factor for adherence-1 gene (efa-1′) confers protective immunity to mice infected with E. coli O157: H7. Front. Cell Infect. Microbiol. 2016, 5, 104. [Google Scholar] [CrossRef] [Green Version]
- Qadri, F.; Svennerholm, A.-M.; Faruque, A.; Sack, R.B. Enterotoxigenic Escherichia coli in developing countries: Epidemiology, microbiology, clinical features, treatment, and prevention. Clin. Microbiol. Rev. 2005, 18, 465–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoh, A.I.; Osode, A.Ν. Enterotoxigenic Escherichia coli (ETEC): A recurring decimal in infants’ and travelers’ diarrhea. Rev. Environ. Health 2008, 23, 135–148. [Google Scholar] [CrossRef]
- Vicente, A.C.; Teixeira, L.F.; Iniguez-Rojas, L.; Luna, M.d.G.; Silva, L.; Andrade, J.R.d.C.; Guth, B.E.C. Outbreaks of cholera-like diarrhoea caused by enterotoxigenic Escherichia coli in the Brazilian Amazon Rainforest. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, J.M.; Hardwidge, P.R.; Munson, G.P.; Rasko, D.A.; Sommerfelt, H.; Steinsland, H. Molecular mechanisms of enterotoxigenic Escherichia coli infection. Microbes Infect. 2010, 12, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, J.M.; Kuhlmann, F.M. Enterotoxigenic Escherichia coli infections. Curr. Infect. Dis. Rep. 2019, 21, 9. [Google Scholar] [CrossRef]
- Khalil, I.; Walker, R.; Porter, C.K.; Muhib, F.; Chilengi, R.; Cravioto, A.; Guerrant, R.; Svennerholm, A.-M.; Qadri, F.; Baqar, S. Enterotoxigenic Escherichia coli (ETEC) vaccines: Priority activities to enable product development, licensure, and global access. Vaccine 2021, 39, 4266–4277. [Google Scholar] [CrossRef]
- Hosangadi, D.; Smith, P.G.; Kaslow, D.C.; Giersing, B.K. WHO consultation on ETEC and Shigella burden of disease, Geneva, 6–7th April 2017: Meeting report. Vaccine 2019, 37, 7381–7390. [Google Scholar] [CrossRef]
- McGregor, A.C.; Wright, S.G. Gastrointestinal symptoms in travellers. Clin. Med. 2015, 15, 93. [Google Scholar] [CrossRef] [Green Version]
- Steffen, R. Epidemiology of traveler’s diarrhea. Clin. Infect. Dis. 2005, 41, S536–S540. [Google Scholar] [CrossRef]
- Olson, S.; Hall, A.; Riddle, M.S.; Porter, C.K. Travelers’ diarrhea: Update on the incidence, etiology and risk in military and similar populations—1990–2005 versus 2005–2015, does a decade make a difference? Trop. Dis. Travel Med. Vaccines 2019, 5, 1–15. [Google Scholar] [CrossRef]
- Girón, J.A.; Levine, M.M.; Kaper, J.B. Longus: A long pilus ultrastructure produced by human enterotoxigenic Escherichia coli. Mol. Microbiol. 1994, 12, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Spangler, B.D. Structure and function of cholera toxin and the related Escherichia coli heat-labile enterotoxin. Microbiol. Rev. 1992, 56, 622–647. [Google Scholar] [CrossRef] [PubMed]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Khan, A. Basic facts of mastitis in dairy animals: A review. Pak. Vet. J. 2006, 26, 204. [Google Scholar]
- Patel, S.K.; Dotson, J.; Allen, K.P.; Fleckenstein, J.M. Identification and molecular characterization of EatA, an autotransporter protein of enterotoxigenic Escherichia coli. Infect. Immun. 2004, 72, 1786–1794. [Google Scholar] [CrossRef] [Green Version]
- Qadri, F.; Akhtar, M.; Bhuiyan, T.R.; Chowdhury, M.I.; Ahmed, T.; Rafique, T.A.; Khan, A.; Rahman, S.I.; Khanam, F.; Lundgren, A. Safety and immunogenicity of the oral, inactivated, enterotoxigenic Escherichia coli vaccine ETVAX in Bangladeshi children and infants: A double-blind, randomised, placebo-controlled phase 1/2 trial. Lancet Infect. Dis. 2020, 20, 208–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, E.B.; Lawson, L.B.; Freytag, L.C.; Clements, J.D. Characterization of a mutant Escherichia coli heat-labile toxin, LT (R192G/L211A), as a safe and effective oral adjuvant. Clin. Vaccine Immunol. 2011, 18, 546–551. [Google Scholar] [CrossRef] [Green Version]
- Clements, J.D.; Norton, E.B. The mucosal vaccine adjuvant LT (R192G/L211A) or dmLT. MSphere 2018, 3, e00215-18. [Google Scholar] [CrossRef] [Green Version]
- Leach, S.; Lundgren, A.; Carlin, N.; Löfstrand, M.; Svennerholm, A.-M. Cross-reactivity and avidity of antibody responses induced in humans by the oral inactivated multivalent enterotoxigenic Escherichia coli (ETEC) vaccine ETVAX. Vaccine 2017, 35, 3966–3973. [Google Scholar] [CrossRef]
- Barry, E.M.; Levine, M.M. A tale of two bacterial enteropathogens and one multivalent vaccine. Cell Microbiol. 2019, 21, e13067. [Google Scholar] [CrossRef]
- Lundgren, A.; Bourgeois, L.; Carlin, N.; Clements, J.; Gustafsson, B.; Hartford, M.; Holmgren, J.; Petzold, M.; Walker, R.; Svennerholm, A.-M. Safety and immunogenicity of an improved oral inactivated multivalent enterotoxigenic Escherichia coli (ETEC) vaccine administered alone and together with dmLT adjuvant in a double-blind, randomized, placebo-controlled Phase I study. Vaccine 2014, 32, 7077–7084. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.K.; Stephens, J.C.; Beavis, J.C.; Greenwood, J.; Gewert, C.; Randall, R.; Freeman, D.; Darsley, M.J. Generation and characterization of a live attenuated enterotoxigenic Escherichia coli combination vaccine expressing six colonization factors and heat-labile toxin subunit B. Clin. Vaccine Immunol. 2011, 18, 2128–2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.K.; Terry, T.D.; Sack, D.A.; Londoño-Arcila, P.; Darsley, M.J. Construction and characterization of genetically defined aro omp mutants of enterotoxigenic Escherichia coli and preliminary studies of safety and immunogenicity in humans. Infect. Immun. 2001, 69, 4969–4979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, M.M.; Girón, J.A.; Noriega, F.R. Fimbrial vaccines. In Fimbriae; CRC Press: Boca Raton, FL, USA, 2020; pp. 255–270. [Google Scholar]
- Barry, E.M.; Levine, M.M. Multivalent Shigella Enterotoxigenic Escherichia coli Vaccine. In New Generation Vaccines; CRC Press: Boca Raton, FL, USA, 2016; pp. 751–755. [Google Scholar]
- Barry, E.; Cassels, F.; Riddle, M.; Walker, R.; Wierzba, T. Vaccines against Shigella and Enterotoxigenic Escherichia coli: A summary of the 2018 VASE Conference. Vaccine 2019, 37, 4768–4774. [Google Scholar] [CrossRef] [PubMed]
- Koprowski, H.; Levine, M.M.; Anderson, R.J.; Losonsky, G.; Pizza, M.; Barry, E.M. Attenuated Shigella flexneri 2a vaccine strain CVD 1204 expressing colonization factor antigen I and mutant heat-labile enterotoxin of enterotoxigenic Escherichia coli. Infect. Immun. 2000, 68, 4884–4892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svennerholm, A.-M.; Glenn, G. Vaccines against enterotoxigenic Escherichia coli. In New Generation Vaccines; CRC Press: Boca Raton, FL, USA, 2016; pp. 742–750. [Google Scholar] [CrossRef]
- Girardi, P.; Harutyunyan, S.; Neuhauser, I.; Glaninger, K.; Korda, O.; Nagy, G.; Nagy, E.; Szijártó, V.; Pall, D.; Szarka, K. Evaluation of the Safety, Tolerability and Immunogenicity of ShigETEC, an Oral Live Attenuated Shigella-ETEC Vaccine in Placebo-Controlled Randomized Phase 1 Trial. Vaccines 2022, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Chowdhury, M.I.; Bhuiyan, T.R.; Kaim, J.; Ahmed, T.; Rafique, T.A.; Khan, A.; Rahman, S.I.; Khanam, F.; Begum, Y.A. Evaluation of the safety and immunogenicity of the oral inactivated multivalent enterotoxigenic Escherichia coli vaccine ETVAX in Bangladeshi adults in a double-blind, randomized, placebo-controlled Phase I trial using electrochemiluminescence and ELISA assays for immunogenicity analyses. Vaccine 2019, 37, 5645–5656. [Google Scholar] [PubMed]
- Seo, H.; Zhang, W. Development of effective vaccines for enterotoxigenic Escherichia coli. Lancet Infect. Dis. 2020, 20, 150–152. [Google Scholar] [CrossRef] [Green Version]
- Harro, C.; Sack, D.; Bourgeois, A.L.; Walker, R.; DeNearing, B.; Feller, A.; Chakraborty, S.; Buchwaldt, C.; Darsley, M.J. A combination vaccine consisting of three live attenuated enterotoxigenic Escherichia coli strains expressing a range of colonization factors and heat-labile toxin subunit B is well tolerated and immunogenic in a placebo-controlled double-blind phase I trial in healthy adults. Clin. Vaccine Immunol. 2011, 18, 2118–2127. [Google Scholar]
- Tobias, J.; Lebens, M.; Bölin, I.; Wiklund, G.; Svennerholm, A.-M. Construction of non-toxic Escherichia coli and Vibrio cholerae strains expressing high and immunogenic levels of enterotoxigenic E. coli colonization factor I fimbriae. Vaccine 2008, 26, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Randall, A.; Vickers, T.J.; Molina, D.; Harro, C.D.; DeNearing, B.; Brubaker, J.; Sack, D.A.; Bourgeois, A.L.; Felgner, P.L. Interrogation of a live-attenuated enterotoxigenic Escherichia coli vaccine highlights features unique to wild-type infection. NPJ Vaccines 2019, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobias, J.; Holmgren, J.; Hellman, M.; Nygren, E.; Lebens, M.; Svennerholm, A.-M. Over-expression of major colonization factors of enterotoxigenic Escherichia coli, alone or together, on non-toxigenic E. coli bacteria. Vaccine 2010, 28, 6977–6984. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Bartels, S.; Qadri, F.; Fleckenstein, J.M. Enterotoxigenic Escherichia coli elicits immune responses to multiple surface proteins. Infect. Immun. 2010, 78, 3027–3035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, K.; Hamilton, D.; Ostmann, M.M.; Fleckenstein, J.M. Vaccination with EtpA glycoprotein or flagellin protects against colonization with enterotoxigenic Escherichia coli in a murine model. Vaccine 2009, 27, 4601–4608. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Hamilton, D.J.; Munson, G.P.; Fleckenstein, J.M. Outer membrane vesicles induce immune responses to virulence proteins and protect against colonization by enterotoxigenic Escherichia coli. Clin. Vaccine Immunol. 2011, 18, 1803–1808. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Iqbal, J.; Gómez-Duarte, O.G. Murine immunization with CS21 pili or LngA major subunit of enterotoxigenic Escherichia coli (ETEC) elicits systemic and mucosal immune responses and inhibits ETEC gut colonization. Vet. Microbiol. 2017, 202, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Leitner, D.R.; Lichtenegger, S.; Temel, P.; Zingl, F.G.; Ratzberger, D.; Roier, S.; Schild-Prüfert, K.; Feichter, S.; Reidl, J.; Schild, S. A combined vaccine approach against Vibrio cholerae and ETEC based on outer membrane vesicles. Front. Microbiol. 2015, 6, 823. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.A.; Roy, K.; Woo-Rasberry, V.; Hamilton, D.J.; Kansal, R.; Qadri, F.; Fleckenstein, J.M. Directed evaluation of enterotoxigenic Escherichia coli autotransporter proteins as putative vaccine candidates. PLoS Negl. Trop. Dis. 2011, 5, e1428. [Google Scholar] [CrossRef] [Green Version]
- Behrens, R.H.; Cramer, J.P.; Jelinek, T.; Shaw, H.; von Sonnenburg, F.; Wilbraham, D.; Weinke, T.; Bell, D.J.; Asturias, E.; Pauwells, H.L.E. Efficacy and safety of a patch vaccine containing heat-labile toxin from Escherichia coli against travellers’ diarrhoea: A phase 3, randomised, double-blind, placebo-controlled field trial in travellers from Europe to Mexico and Guatemala. Lancet Infect. Dis. 2014, 14, 197–204. [Google Scholar] [CrossRef]
- Rahjerdi, A.K.; Jafari, M.; Motamedi, M.J.; Amani, J.; Salmanian, A.H. Immunogenic Evaluation of Bivalent Vaccine Candidate against Enterohemorrhagic and Enterotoxigenic Escherichia coli. Iran. J. Immunol. 2019, 16, 200–211. [Google Scholar]
- Pasqua, M.; Michelacci, V.; Di Martino, M.L.; Tozzoli, R.; Grossi, M.; Colonna, B.; Morabito, S.; Prosseda, G. The intriguing evolutionary journey of enteroinvasive E. coli (EIEC) toward pathogenicity. Front. Microbiol. 2017, 8, 2390. [Google Scholar] [CrossRef]
- Ud-Din, A.; Wahid, S. Relationship among Shigella spp. and enteroinvasive Escherichia coli (EIEC) and their differentiation. Braz. J. Microbiol. 2014, 45, 1131–1138. [Google Scholar] [CrossRef]
- Lan, R.; Alles, M.C.; Donohoe, K.; Martinez, M.B.; Reeves, P.R. Molecular evolutionary relationships of enteroinvasive Escherichia coli and Shigella spp. Infect. Immun. 2004, 72, 5080–5088. [Google Scholar] [CrossRef] [Green Version]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef]
- Newitt, S.; MacGregor, V.; Robbins, V.; Bayliss, L.; Chattaway, M.A.; Dallman, T.; Ready, D.; Aird, H.; Puleston, R.; Hawker, J. Two linked enteroinvasive Escherichia coli outbreaks, Nottingham, UK, June 2014. Emerg. Infect. Dis. 2016, 22, 1178. [Google Scholar] [CrossRef] [Green Version]
- Lagerqvist, N.; Löf, E.; Enkirch, T.; Nilsson, P.; Roth, A.; Jernberg, C. Outbreak of gastroenteritis highlighting the diagnostic and epidemiological challenges of enteroinvasive Escherichia coli, County of Halland, Sweden, November 2017. Eurosurveillance 2020, 25, 1900466. [Google Scholar] [CrossRef] [Green Version]
- Herzig, C.T.; Fleischauer, A.T.; Lackey, B.; Lee, N.; Lawson, T.; Moore, Z.S.; Hergert, J.; Mobley, V.; MacFarquhar, J.; Morrison, T. Notes from the Field: Enteroinvasive Escherichia coli Outbreak Associated with a Potluck Party—North Carolina, June–July 2018. Morb. Mortal. Wkly. Rep. 2019, 68, 183. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, R.; Wang, Q.; Howard, P.; Sintchenko, V. Genome Sequences of Enteroinvasive Escherichia coli Sequence Type 6, 99, and 311 Strains Acquired in Asia Pacific. Microbiol. Resour. Announc. 2019, 8, e00944-19. [Google Scholar] [CrossRef] [Green Version]
- Prats, G.; Llovet, T. Enteroinvasive Escherichia coli. Pathogenesis and epidemiology. Microbiologia 1995, 11, 91–96. [Google Scholar]
- Schnupf, P.; Sansonetti, P.J. Shigella pathogenesis: New insights through advanced methodologies. Microbiol. Spectr. 2019, 7, 15–39. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Riddle, M.S.; Platts-Mills, J.A.; Pavlinac, P.; Zaidi, A.K. Shigellosis. Lancet 2018, 391, 801–812. [Google Scholar] [CrossRef]
- Lambrecht, N.J.; Bridges, D.; Wilson, M.L.; Adu, B.; Eisenberg, J.N.; Folson, G.; Baylin, A.; Jones, A.D. Associations of bacterial enteropathogens with systemic inflammation, iron deficiency, and anemia in preschool-age children in southern Ghana. PLoS ONE 2022, 17, e0271099. [Google Scholar] [CrossRef]
- Echeverria, P.; Sethabutr, O.; Pitarangsi, C. Microbiology and diagnosis of infections with Shigella and enteroinvasive Escherichia coli. Rev. Infect. Dis. 1991, 13, S220–S225. [Google Scholar] [CrossRef]
- Escobar-Páramo, P.; Giudicelli, C.; Parsot, C.; Denamur, E. The evolutionary history of Shigella and enteroinvasive Escherichia coli revised. J. Mol. Evol. 2003, 57, 140–148. [Google Scholar] [CrossRef]
- Belotserkovsky, I.; Sansonetti, P.J. Shigella and enteroinvasive Escherichia coli. In Escherichia coli a Versatile Pathogen; Springer: Cham, Switzerland, 2018; Volume 416, pp. 1–26. [Google Scholar] [CrossRef]
- Doyle, M. Foodborne Bacterial Pathogens; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Andrade, A.; Girón, J.A.; Amhaz, J.M.; Trabulsi, L.R.; Martinez, M.B. Expression and characterization of flagella in nonmotile enteroinvasive Escherichia coli isolated from diarrhea cases. Infect. Immun. 2002, 70, 5882–5886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasser, A.; Mosadegh, M.; Azimi, T.; Shariati, A. Molecular mechanisms of Shigella effector proteins: A common pathogen among diarrheic pediatric population. Mol. Cell Pediatr. 2022, 9, 1–21. [Google Scholar] [CrossRef]
- Jin, Q.; Yuan, Z.; Xu, J.; Wang, Y.; Shen, Y.; Lu, W.; Wang, J.; Liu, H.; Yang, J.; Yang, F. Genome sequence of Shigella flexneri 2a: Insights into pathogenicity through comparison with genomes of Escherichia coli K12 and O157. Nucleic Acids Res. 2002, 30, 4432–4441. [Google Scholar] [CrossRef]
- Wei, J.; Goldberg, M.; Burland, V.; Venkatesan, M.; Deng, W.; Fournier, G.; Mayhew, G.; Plunkett, G., III; Rose, D.; Darling, A. Complete genome sequence and comparative genomics of Shigella flexneri serotype 2a strain 2457T. Infect. Immun. 2003, 71, 2775–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansonetti, P.; d’Hauteville, H.; Écobichon, C.T.; Pourcel, C. Molecular comparison of virulence plasmids in Shigella and enteroinvasive Escherichia coli. Ann. l’Institut Pasteur/Microbiol. 1983, 134, 295–318. [Google Scholar] [CrossRef]
- Bravo, V.; Puhar, A.; Sansonetti, P.; Parsot, C.; Toro, C.S. Distinct mutations led to inactivation of type 1 fimbriae expression in Shigella spp. PLoS ONE 2015, 10, e0121785. [Google Scholar] [CrossRef]
- Ramos, H.C.; Rumbo, M.; Sirard, J.-C. Bacterial flagellins: Mediators of pathogenicity and host immune responses in mucosa. Trends Microbiol. 2004, 12, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.; Wierzba, T.; Walker, R.I. Status of vaccine research and development for Shigella. Vaccine 2016, 34, 2887–2894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLennan, C.A.; Grow, S.; Ma, L.-F.; Steele, A.D. The Shigella vaccines pipeline. Vaccines 2022, 10, 1376. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Li, B.; Xu, T.; Yu, H.; Chen, J.; Yu, H.; Li, S.; Zeng, L.; Huang, X.; Liu, Q. Outer Membrane Vesicles Derived from Salmonella enterica Serotype Typhimurium Can Deliver Shigella flexneri 2a O-Polysaccharide Antigen To Prevent Shigella flexneri 2a Infection in Mice. Appl. Environ. Microbiol. 2021, 87, e00968-0021. [Google Scholar] [CrossRef]
- Camacho, A.I.; Irache, J.M.; Gamazo, C. Recent progress towards development of a Shigella vaccine. Expert Rev. Vaccines 2013, 12, 43–55. [Google Scholar] [CrossRef]
- Pastor, Y.; Camacho, A.I.; Zúñiga-Ripa, A.; Merchán, A.; Rosas, P.; Irache, J.M.; Gamazo, C. Towards a subunit vaccine from a Shigella flexneri ΔtolR mutant. Vaccine 2018, 36, 7509–7519. [Google Scholar] [CrossRef] [PubMed]
- Camacho, A.; Irache, J.; De Souza, J.; Sánchez-Gómez, S.; Gamazo, C. Nanoparticle-based vaccine for mucosal protection against Shigella flexneri in mice. Vaccine 2013, 31, 3288–3294. [Google Scholar] [CrossRef] [PubMed]
- Barel, L.-A.; Mulard, L.A. Classical and novel strategies to develop a Shigella glycoconjugate vaccine: From concept to efficacy in human. Hum. Vaccines Immunother. 2019, 15, 1338–1356. [Google Scholar] [CrossRef]
- Levine, M.M.; Kotloff, K.L.; Barry, E.M.; Pasetti, M.F.; Sztein, M.B. Clinical trials of Shigella vaccines: Two steps forward and one step back on a long, hard road. Nat. Rev. Microbiol. 2007, 5, 540–553. [Google Scholar] [CrossRef] [Green Version]
- Alexander, W.A.; Hartman, A.B.; Oaks, E.V.; Venkatesan, M.M. Construction and characterization of virG (icsA)-deleted Escherichia coli K12-Shigella flexneri hybrid vaccine strains. Vaccine 1996, 14, 1053–1061. [Google Scholar] [CrossRef]
- Harutyunyan, S.; Neuhauser, I.; Mayer, A.; Aichinger, M.; Szijártó, V.; Nagy, G.; Nagy, E.; Girardi, P.; Malinoski, F.J.; Henics, T. Characterization of shigetec, a novel live attenuated combined vaccine against shigellae and etec. Vaccines 2020, 8, 689. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, M.M.; Ballou, C.; Barnoy, S.; McNeal, M.; El-Khorazaty, J.; Frenck, R.; Baqar, S. Antibody in Lymphocyte Supernatant (ALS) responses after oral vaccination with live Shigella sonnei vaccine candidates WRSs2 and WRSs3 and correlation with serum antibodies, ASCs, fecal IgA and shedding. PLoS ONE 2021, 16, e0259361. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Moon, Y.-h.; Kim, H.; Rho, S.; Shin, Y.K.; Song, M.; Walker, R.; Czerkinsky, C.; Kim, D.W.; Kim, J.-O. Cross-protective Shigella whole-cell vaccine with a truncated O-polysaccharide chain. Front. Microbiol. 2018, 9, 2609. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Alaimo, C. The Ongoing Journey of a Shigella Bioconjugate Vaccine. Vaccines 2022, 10, 212. [Google Scholar] [CrossRef]
- Phalipon, A.; Mulard, L.A. Toward a Multivalent Synthetic Oligosaccharide-Based Conjugate Vaccine against Shigella: State-of-the-Art for a Monovalent Prototype and Challenges. Vaccines 2022, 10, 403. [Google Scholar] [CrossRef]
- Mo, Y.; Fang, W.; Li, H.; Chen, J.; Hu, X.; Wang, B.; Feng, Z.; Shi, H.; He, Y.; Huang, D. Safety and Immunogenicity of a Shigella Bivalent Conjugate Vaccine (ZF0901) in 3-Month-to 5-Year-Old Children in China. Vaccines 2021, 10, 33. [Google Scholar] [CrossRef]
- Nataro, J.P.; Kaper, J.B.; Robins-Browne, R.; Prado, V.; Vial, P.; Levine, M.M. Patterns of adherence of diarrheagenic Escherichia coli to HEp-2 cells. Pediatr. Infect. Dis. J. 1987, 6, 829–831. [Google Scholar] [CrossRef]
- Harrington, S.M.; Dudley, E.G.; Nataro, J.P. Pathogenesis of enteroaggregative Escherichia coli infection. FEMS Microbiol. Lett. 2006, 254, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Czeczulin, J.R.; Balepur, S.; Hicks, S.; Phillips, A.; Hall, R.; Kothary, M.H.; Navarro-Garcia, F.; Nataro, J.P. Aggregative adherence fimbria II, a second fimbrial antigen mediating aggregative adherence in enteroaggregative Escherichia coli. Infect. Immun. 1997, 65, 4135–4145. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, C. Enteroaggregative Escherichia coli. In Escherichia coli a Versatile Pathogen; Springer: Cham, Switzerland, 2018; pp. 27–50. [Google Scholar] [CrossRef]
- Fasano, A.; Noriega, F.; Liao, F.; Wang, W.; Levine, M. Effect of shigella enterotoxin 1 (ShET1) on rabbit intestine in vitro and in vivo. Gut 1997, 40, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Savarino, S.J.; McVeigh, A.; Watson, J.; Cravioto, A.; Molina, J.; Echeverria, P.; Bhan, M.K.; Levine, M.M.; Fasano, A. Enteroaggregative Escherichia coli heat-stable enterotoxin is not restricted to enteroaggregative E. coli. J. Infect. Dis. 1996, 173, 1019–1022. [Google Scholar] [CrossRef] [Green Version]
- Eslava, C.; Navarro-García, F.; Czeczulin, J.R.; Henderson, I.R.; Cravioto, A.; Nataro, J.P. Pet, an autotransporter enterotoxin from enteroaggregative Escherichia coli. Infect. Immun. 1998, 66, 3155–3163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villaseca, J.M.; Navarro-García, F.; Mendoza-Hernández, G.; Nataro, J.P.; Cravioto, A.; Eslava, C. Pet toxin from enteroaggregative Escherichia coli produces cellular damage associated with fodrin disruption. Infect. Immun. 2000, 68, 5920–5927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, S.M.; Sheikh, J.; Henderson, I.R.; Ruiz-Perez, F.; Cohen, P.S.; Nataro, J.P. The Pic protease of enteroaggregative Escherichia coli promotes intestinal colonization and growth in the presence of mucin. Infect. Immun. 2009, 77, 2465–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, A.G.; Fraga, T.R.; Granados Martínez, A.P.; Kondo, M.Y.; Juliano, M.A.; Juliano, L.; Navarro-Garcia, F.; Isaac, L.; Barbosa, A.S.; Elias, W.P. The serine protease Pic from enteroaggregative Escherichia coli mediates immune evasion by the direct cleavage of complement proteins. J. Infect. Dis. 2015, 212, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Perez, F.; Wahid, R.; Faherty, C.S.; Kolappaswamy, K.; Rodriguez, L.; Santiago, A.; Murphy, E.; Cross, A.; Sztein, M.B.; Nataro, J.P. Serine protease autotransporters from Shigella flexneri and pathogenic Escherichia coli target a broad range of leukocyte glycoproteins. Proc. Natl. Acad. Sci. USA 2011, 108, 12881–12886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Servin, A.L. Pathogenesis of human diffusely adhering Escherichia coli expressing Afa/Dr adhesins (Afa/Dr DAEC): Current insights and future challenges. Clin. Microbiol. Rev. 2014, 27, 823–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Bouguénec, C.; Servin, A.L. Diffusely adherent Escherichia coli strains expressing Afa/Dr adhesins (Afa/Dr DAEC): Hitherto unrecognized pathogens. FEMS Microbiol. Lett. 2006, 256, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Servin, A.L. Pathogenesis of Afa/Dr diffusely adhering Escherichia coli. Clin. Microbiol. Rev. 2005, 18, 264–292. [Google Scholar] [CrossRef] [Green Version]
- Meza-Segura, M.; Zaidi, M.B.; Vera-Ponce de León, A.; Moran-Garcia, N.; Martinez-Romero, E.; Nataro, J.P.; Estrada-Garcia, T. New insights into DAEC and EAEC pathogenesis and phylogeny. Front. Cell Infect. Microbiol. 2020, 10, 572951. [Google Scholar] [CrossRef] [PubMed]
- Mirsepasi-Lauridsen, H.C.; Vallance, B.A.; Krogfelt, K.A.; Petersen, A.M. Escherichia coli pathobionts associated with inflammatory bowel disease. Clin. Microbiol. Rev. 2019, 32, e00060-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultsz, C.; Moussa, M.; Van Ketel, R.; Tytgat, G.N.; Dankert, J. Frequency of pathogenic and enteroadherent Escherichia coli in patients with inflammatory bowel disease and controls. J. Clin. Pathol. 1997, 50, 573–579. [Google Scholar] [CrossRef] [Green Version]
- Canizalez-Roman, A.; Flores-Villaseñor, H.M.; Gonzalez-Nuñez, E.; Velazquez-Roman, J.; Vidal, J.E.; Muro-Amador, S.; Alapizco-Castro, G.; Díaz-Quiñonez, J.A.; León-Sicairos, N. Surveillance of diarrheagenic Escherichia coli strains isolated from diarrhea cases from children, adults and elderly at Northwest of Mexico. Front. Microbiol. 2016, 7, 1924. [Google Scholar] [CrossRef] [Green Version]
- Mohamed Ali, M. Evaluation of antimicrobial susceptibility &rapid urine screening tests in asymptomatic urinary tract infection in pregnant women in Karbala. Karbala J. Pharm. Sci. 2011, 2, 22–34. [Google Scholar]
- Bouzari, S.; Dashti, A.; Jafari, A.; Oloomi, M. Immune response against adhesins of enteroaggregative Escherichia coli immunized by three different vaccination strategies (DNA/DNA, Protein/Protein, and DNA/Protein) in mice. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 215–225. [Google Scholar] [CrossRef]
- Saeedi, P.; Yazdanparast, M.; Behzadi, E.; Salmanian, A.H.; Mousavi, S.L.; Nazarian, S.; Amani, J. A review on strategies for decreasing E. coli O157: H7 risk in animals. Microb. Pathog. 2017, 103, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Goluszko, P.; Goluszko, E.; Nowicki, B.; Nowicki, S.; Popov, V.; Wang, H.-Q. Vaccination with purified Dr Fimbriae reduces mortality associated with chronic urinary tract infection due to Escherichia coli bearing Dr adhesin. Infect. Immun. 2005, 73, 627–631. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.J.; Thompson, A.P.; O’Driscoll, A.; Clarke, D.J. Pathogenesis of adherent–invasive Escherichia coli. Future Microbiol. 2013, 8, 1289–1300. [Google Scholar] [CrossRef]
- Palmela, C.; Chevarin, C.; Xu, Z.; Torres, J.; Sevrin, G.; Hirten, R.; Barnich, N.; Ng, S.C.; Colombel, J.-F. Adherent-invasive Escherichia coli in inflammatory bowel disease. Gut 2018, 67, 574–587. [Google Scholar] [CrossRef]
- Chervy, M.; Barnich, N.; Denizot, J. Adherent-Invasive E. coli: Update on the Lifestyle of a Troublemaker in Crohn’s Disease. Int. J. Mol. Sci. 2020, 21, 3734. [Google Scholar] [CrossRef]
- Martinez-Medina, M.; Aldeguer, X.; Lopez-Siles, M.; González-Huix, F.; López-Oliu, C.; Dahbi, G.; Blanco, J.E.; Blanco, J.; Garcia-Gil, J.L.; Darfeuille-Michaud, A. Molecular diversity of Escherichia coli in the human gut: New ecological evidence supporting the role of adherent-invasive E. coli (AIEC) in Crohn’s disease. Inflamm. Bowel Dis. 2009, 15, 872–882. [Google Scholar] [CrossRef]
- Conte, M.P.; Longhi, C.; Marazzato, M.; Conte, A.L.; Aleandri, M.; Lepanto, M.S.; Zagaglia, C.; Nicoletti, M.; Aloi, M.; Totino, V. Adherent-invasive Escherichia coli (AIEC) in pediatric Crohn’s disease patients: Phenotypic and genetic pathogenic features. BMC Res. Notes 2014, 7, 748. [Google Scholar] [CrossRef] [PubMed]
- Kamali Dolatabadi, R.; Feizi, A.; Halaji, M.; Fazeli, H.; Adibi, P. The prevalence of adherent-invasive Escherichia coli and its association with inflammatory bowel diseases: A systematic review and meta-analysis. Front. Med. 2021, 8, 730243. [Google Scholar] [CrossRef]
- Darfeuille-Michaud, A.; Boudeau, J.; Bulois, P.; Neut, C.; Glasser, A.-L.; Barnich, N.; Bringer, M.-A.; Swidsinski, A.; Beaugerie, L.; Colombel, J.-F. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 2004, 127, 412–421. [Google Scholar] [CrossRef]
- Leccese, G.; Bibi, A.; Mazza, S.; Facciotti, F.; Caprioli, F.; Landini, P.; Paroni, M. Probiotic Lactobacillus and Bifidobacterium strains counteract adherent-invasive Escherichia coli (AIEC) virulence and hamper IL-23/Th17 axis in ulcerative colitis, but not in crohn’s disease. Cells 2020, 9, 1824. [Google Scholar] [CrossRef]
- Denizot, J.; Sivignon, A.; Barreau, F.; Darcha, C.; Chan, C.H.; Stanners, C.P.; Hofman, P.; Darfeuille-Michaud, A.; Barnich, N. Adherent-invasive Escherichia coli induce claudin-2 expression and barrier defect in CEABAC10 mice and Crohn’s disease patients. Inflamm. Bowel Dis. 2012, 18, 294–304. [Google Scholar] [CrossRef]
- Bringer, M.-A.; Darfeuille-Michaud, A. Bacterial Adhesion to Intestinal Mucosa. In Mucosal Immunology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 949–953. [Google Scholar]
- Barnich, N.; Carvalho, F.A.; Glasser, A.-L.; Darcha, C.; Jantscheff, P.; Allez, M.; Peeters, H.; Bommelaer, G.; Desreumaux, P.; Colombel, J.-F. CEACAM6 acts as a receptor for adherent-invasive E. coli, supporting ileal mucosa colonization in Crohn disease. J. Clin. Investig. 2007, 117, 1566–1574. [Google Scholar] [CrossRef] [Green Version]
- Rolhion, N.; Hofman, P.; Darfeuille-Michaud, A. The endoplasmic reticulum stress response chaperone Gp96, a host receptor for Crohn disease-associated adherent-invasive Escherichia coli. Gut Microbes 2011, 2, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Rolhion, N.; Barnich, N.; Bringer, M.-A.; Glasser, A.-L.; Ranc, J.; Hébuterne, X.; Hofman, P.; Darfeuille-Michaud, A. Abnormally expressed ER stress response chaperone Gp96 in CD favours adherent-invasive Escherichia coli invasion. Gut 2010, 59, 1355–1362. [Google Scholar] [CrossRef]
- Gibold, L.; Garenaux, E.; Dalmasso, G.; Gallucci, C.; Cia, D.; Mottet-Auselo, B.; Faïs, T.; Darfeuille-Michaud, A.; Nguyen, H.T.T.; Barnich, N. The Vat-AIEC protease promotes crossing of the intestinal mucus layer by Crohn’s disease-associated Escherichia coli. Cell Microbiol. 2016, 18, 617–631. [Google Scholar] [CrossRef]
- Prudent, V.; Demarre, G.; Vazeille, E.; Wery, M.; Quenech’Du, N.; Ravet, A.; Dauverd-Girault, J.; Van Dijk, E.; Bringer, M.-A.; Descrimes, M. The Crohn’s disease-related bacterial strain LF82 assembles biofilm-like communities to protect itself from phagolysosomal attack. Commun. Biol. 2021, 4, 627. [Google Scholar] [CrossRef]
- Chassaing, B.; Koren, O.; Carvalho, F.A.; Ley, R.E.; Gewirtz, A.T. AIEC pathobiont instigates chronic colitis in susceptible hosts by altering microbiota composition. Gut 2014, 63, 1069–1080. [Google Scholar] [CrossRef]
- Nadalian, B.; Yadegar, A.; Houri, H.; Olfatifar, M.; Shahrokh, S.; Asadzadeh Aghdaei, H.; Suzuki, H.; Zali, M.R. Prevalence of the pathobiont adherent-invasive Escherichia coli and inflammatory bowel disease: A systematic review and meta-analysis. J. Gastroenterol. Hepatol. 2021, 36, 852–863. [Google Scholar] [CrossRef]
- Negroni, A.; Colantoni, E.; Vitali, R.; Palone, F.; Pierdomenico, M.; Costanzo, M.; Cesi, V.; Cucchiara, S.; Stronati, L. NOD2 induces autophagy to control AIEC bacteria infectiveness in intestinal epithelial cells. Inflamm. Res. 2016, 65, 803–813. [Google Scholar] [CrossRef]
- Homer, C.R.; Richmond, A.L.; Rebert, N.A.; Achkar, J.P.; McDonald, C. ATG16L1 and NOD2 interact in an autophagy-dependent antibacterial pathway implicated in Crohn’s disease pathogenesis. Gastroenterology 2010, 139, 1630–1641.e2. [Google Scholar] [CrossRef] [Green Version]
- Alvarez Dorta, D.; Sivignon, A.; Chalopin, T.; Dumych, T.I.; Roos, G.; Bilyy, R.O.; Deniaud, D.; Krammer, E.M.; De Ruyck, J.; Lensink, M.F. The antiadhesive strategy in Crohn′ s disease: Orally active mannosides to decolonize pathogenic Escherichia coli from the gut. ChemBioChem 2016, 17, 936–952. [Google Scholar] [CrossRef]
- Sivignon, A.; Yan, X.; Alvarez Dorta, D.; Bonnet, R.; Bouckaert, J.; Fleury, E.; Bernard, J.; Gouin, S.G.; Darfeuille-Michaud, A.; Barnich, N. Development of heptylmannoside-based glycoconjugate antiadhesive compounds against adherent-invasive Escherichia coli bacteria associated with crohn’s disease. MBio 2015, 6, e01298-15. [Google Scholar] [CrossRef] [Green Version]
- Chalopin, T.; Dorta, D.A.; Sivignon, A.; Caudan, M.; Dumych, T.; Bilyy, R.; Deniaud, D.; Barnich, N.; Bouckaert, J.; Gouin, S. Second generation of thiazolylmannosides, FimH antagonists for E. coli-induced Crohn’s disease. Org. Biomol. Chem. 2016, 14, 3913–3925. [Google Scholar] [CrossRef]
- Yan, X.; Sivignon, A.; Yamakawa, N.; Crepet, A.; Travelet, C.; Borsali, R.; Dumych, T.; Li, Z.; Bilyy, R.; Deniaud, D. Glycopolymers as antiadhesives of E. coli strains inducing inflammatory bowel diseases. Biomacromolecules 2015, 16, 1827–1836. [Google Scholar] [CrossRef]
- Boudeau, J.; Glasser, A.L.; Julien, S.; Colombel, J.F.; Darfeuille-Michaud, A. Inhibitory effect of probiotic Escherichia coli strain Nissle 1917 on adhesion to and invasion of intestinal epithelial cells by adherent–invasive E. coli strains isolated from patients with Crohn’s disease. Aliment. Pharmacol. Ther. 2003, 18, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Van den Abbeele, P.; Marzorati, M.; Derde, M.; De Weirdt, R.; Joan, V.; Possemiers, S.; Van de Wiele, T. Arabinoxylans, inulin and Lactobacillus reuteri 1063 repress the adherent-invasive Escherichia coli from mucus in a mucosa-comprising gut model. NPJ Biofilms Microbiomes 2016, 2, 16016. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S. Targeting Siderophores to Reduce Adherent-Invasive E. coli Colonization: A Potential Therapy for Inflammatory Bowel Disease; UC San Diego: San Diego, CA, USA, 2021; Available online: https://escholarship.org/uc/item/3mw9g605 (accessed on 15 December 2022).
- Gerner, R.R.; Hossain, S.; Sargun, A.; Siada, K.; Norton, G.J.; Zheng, T.; Neumann, W.; Nuccio, S.-P.; Nolan, E.M.; Raffatellu, M. Siderophore Immunization Restricted Colonization of Adherent-Invasive Escherichia coli and Ameliorated Experimental Colitis. Mbio 2022, 13, e02184-22. [Google Scholar] [CrossRef] [PubMed]
- Roussel, C.; Sivignon, A.; De Vallee, A.; Garrait, G.; Denis, S.; Tsilia, V.; Ballet, N.; Vandekerckove, P.; Van de Wiele, T.; Barnich, N. Anti-infectious properties of the probiotic Saccharomyces cerevisiae CNCM I-3856 on enterotoxigenic E. coli (ETEC) strain H10407. Appl. Microbiol. Biotechnol. 2018, 102, 6175–6189. [Google Scholar] [CrossRef] [PubMed]
- Mantegazza, C.; Molinari, P.; D’Auria, E.; Sonnino, M.; Morelli, L.; Zuccotti, G.V. Probiotics and antibiotic-associated diarrhea in children: A review and new evidence on Lactobacillus rhamnosus GG during and after antibiotic treatment. Pharmacol. Res. 2018, 128, 63–72. [Google Scholar] [CrossRef]
- Nguyen, N.K.; Deehan, E.C.; Zhang, Z.; Jin, M.; Baskota, N.; Perez-Muñoz, M.E.; Cole, J.; Tuncil, Y.E.; Seethaler, B.; Wang, T. Gut microbiota modulation with long-chain corn bran arabinoxylan in adults with overweight and obesity is linked to an individualized temporal increase in fecal propionate. Microbiome 2020, 8, 118. [Google Scholar] [CrossRef]
- Canfora, E.E.; van der Beek, C.M.; Hermes, G.D.; Goossens, G.H.; Jocken, J.W.; Holst, J.J.; van Eijk, H.M.; Venema, K.; Smidt, H.; Zoetendal, E.G. Supplementation of diet with galacto-oligosaccharides increases bifidobacteria, but not insulin sensitivity, in obese prediabetic individuals. Gastroenterology 2017, 153, 87–97.e3. [Google Scholar] [CrossRef]
- Kittana, H.H.B. Deciphering Interactions Between the Gut Microbiota and Host Immune System During Intestinal Inflammation; The University of Nebraska-Lincoln: Lincoln, NE, USA, 2018. [Google Scholar]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Zhang, Y.; Rowehl, L.; Krumsiek, J.M.; Orner, E.P.; Shaikh, N.; Tarr, P.I.; Sodergren, E.; Weinstock, G.M.; Boedeker, E.C.; Xiong, X. Identification of candidate adherent-invasive E. coli signature transcripts by genomic/transcriptomic analysis. PLoS ONE 2015, 10, e0130902. [Google Scholar] [CrossRef] [Green Version]
- Kotsiliti, E. Phage therapy suppresses gut inflammation in IBD. Nat. Biotechnol. 2022, 40, 1327. [Google Scholar] [CrossRef] [PubMed]
- Skyberg, J.A.; Johnson, T.J.; Johnson, J.R.; Clabots, C.; Logue, C.M.; Nolan, L.K. Acquisition of avian pathogenic Escherichia coli plasmids by a commensal E. coli isolate enhances its abilities to kill chicken embryos, grow in human urine, and colonize the murine kidney. Infect. Immun. 2006, 74, 6287–6292. [Google Scholar] [CrossRef] [Green Version]
- Bauchart, P.; Germon, P.; Brée, A.; Oswald, E.; Hacker, J.; Dobrindt, U. Pathogenomic comparison of human extraintestinal and avian pathogenic Escherichia coli–search for factors involved in host specificity or zoonotic potential. Microb. Pathog. 2010, 49, 105–115. [Google Scholar] [CrossRef]
- Moulin-Schouleur, M.; Répérant, M.; Laurent, S.; Brée, A.; Mignon-Grasteau, S.; Germon, P.; Rasschaert, D.; Schouler, C. Extraintestinal pathogenic Escherichia coli strains of avian and human origin: Link between phylogenetic relationships and common virulence patterns. J. Clin. Microbiol. 2007, 45, 3366–3376. [Google Scholar] [CrossRef] [Green Version]
- Habouria, H.; Pokharel, P.; Maris, S.; Garénaux, A.; Bessaiah, H.; Houle, S.; Veyrier, F.J.; Guyomard-Rabenirina, S.; Talarmin, A.; Dozois, C.M. Three new serine-protease autotransporters of Enterobacteriaceae (SPATEs) from extra-intestinal pathogenic Escherichia coli and combined role of SPATEs for cytotoxicity and colonization of the mouse kidney. Virulence 2019, 10, 568–587. [Google Scholar] [CrossRef] [Green Version]
- Tabasi, M.; Karam, M.R.A.; Habibi, M.; Mostafavi, E.; Bouzari, S. Genotypic characterization of virulence factors in Escherichia coli isolated from patients with acute cystitis, pyelonephritis and asymptomatic bacteriuria. J. Clin. Diagn. Res. JCDR 2016, 10, DC01. [Google Scholar]
- Foxman, B. The epidemiology of urinary tract infection. Nat. Rev. Urol. 2010, 7, 653–660. [Google Scholar]
- Lloyd, A.L.; Rasko, D.A.; Mobley, H.L. Defining genomic islands and uropathogen-specific genes in uropathogenic Escherichia coli. J. Bacteriol. 2007, 189, 3532–3546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Garcia, F.; Gutierrez-Jimenez, J.; Garcia-Tovar, C.; Castro, L.A.; Salazar-Gonzalez, H.; Cordova, V. Pic, an autotransporter protein secreted by different pathogens in the Enterobacteriaceae family, is a potent mucus secretagogue. Infect. Immun. 2010, 78, 4101–4109. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, B.K.; Mulvey, M.A. The UPEC pore-forming toxin α-hemolysin triggers proteolysis of host proteins to disrupt cell adhesion, inflammatory, and survival pathways. Cell Host Microbe 2012, 11, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Gao, S.; Huan, H.; Xu, X.; Zhu, X.; Yang, W.; Gao, Q.; Liu, X. Comparison of virulence factors and expression of specific genes between uropathogenic Escherichia coli and avian pathogenic E. coli in a murine urinary tract infection model and a chicken challenge model. Microbiology 2009, 155, 1634–1644. [Google Scholar] [CrossRef] [Green Version]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) infections: Virulence factors, bladder responses, antibiotic, and non-antibiotic antimicrobial strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef] [Green Version]
- Schmiemann, G.; Kniehl, E.; Gebhardt, K.; Matejczyk, M.M.; Hummers-Pradier, E. The diagnosis of urinary tract infection: A systematic review. Dtsch. Ärzteblatt Int. 2010, 107, 361. [Google Scholar]
- Ranjan, A.; Sridhar, S.T.K.; Matta, N.; Chokkakula, S.; Ansari, R.K. Prevalence of UTI among pregnant women and its complications in newborns. Indian J. Pharm. Pract. 2017, 10, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Wagenlehner, F.M.; Umeh, O.; Steenbergen, J.; Yuan, G.; Darouiche, R.O. Ceftolozane-tazobactam compared with levofloxacin in the treatment of complicated urinary-tract infections, including pyelonephritis: A randomised, double-blind, phase 3 trial (ASPECT-cUTI). Lancet 2015, 385, 1949–1956. [Google Scholar] [CrossRef]
- Foxman, B. Urinary tract infection syndromes: Occurrence, recurrence, bacteriology, risk factors, and disease burden. Infect. Dis. Clin. 2014, 28, 1–13. [Google Scholar] [CrossRef]
- Nielsen, K.L.; Stegger, M.; Kiil, K.; Lilje, B.; Ejrnæs, K.; Leihof, R.F.; Skjøt-Rasmussen, L.; Godfrey, P.; Monsen, T.; Ferry, S. Escherichia coli causing recurrent urinary tract infections: Comparison to non-recurrent isolates and genomic adaptation in recurrent infections. Microorganisms 2021, 9, 1416. [Google Scholar] [CrossRef] [PubMed]
- Ejrnæs, K. Bacterial characteristics of importance for recurrent urinary tract infections caused by Escherichia coli. Dan Med. Bull 2011, 58, B4187. [Google Scholar]
- Ikähelmo, R.; Siitonen, A.; Heiskanen, T.; Kärkkäinen, U.; Kuosmanen, P.; Lipponen, P.; Mäkelä, P.H. Recurrence of urinary tract infection in a primary care setting: Analysis of a I-year follow-up of 179 women. Clin. Infect. Dis. 1996, 22, 91–99. [Google Scholar] [CrossRef]
- O’Connell Motherway, M.; Houston, A.; O’Callaghan, G.; Reunanen, J.; O’Brien, F.; O’Driscoll, T.; Casey, P.G.; de Vos, W.M.; van Sinderen, D.; Shanahan, F. A Bifidobacterial pilus-associated protein promotes colonic epithelial proliferation. Mol. Microbiol. 2019, 111, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Welch, R.A.; Burland, V.; Plunkett, G., III; Redford, P.; Roesch, P.; Rasko, D.; Buckles, E.; Liou, S.-R.; Boutin, A.; Hackett, J. Extensive mosaic structure revealed by the complete genome sequence of uropathogenic Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 17020–17024. [Google Scholar]
- Zhou, G.; Mo, W.-J.; Sebbel, P.; Min, G.; Neubert, T.A.; Glockshuber, R.; Wu, X.-R.; Sun, T.-T.; Kong, X.-P. Uroplakin Ia is the urothelial receptor for uropathogenic Escherichia coli: Evidence from in vitro FimH binding. J. Cell Sci. 2001, 114, 4095–4103. [Google Scholar] [CrossRef] [PubMed]
- Kisiela, D.I.; Avagyan, H.; Friend, D.; Jalan, A.; Gupta, S.; Interlandi, G.; Liu, Y.; Tchesnokova, V.; Rodriguez, V.B.; Sumida, J.P. Inhibition and reversal of microbial attachment by an antibody with parasteric activity against the FimH adhesin of uropathogenic E. coli. PLoS Pathog. 2015, 11, e1004857. [Google Scholar] [CrossRef] [Green Version]
- Ashkar, A.A.; Mossman, K.L.; Coombes, B.K.; Gyles, C.L.; Mackenzie, R. FimH adhesin of type 1 fimbriae is a potent inducer of innate antimicrobial responses which requires TLR4 and type 1 interferon signalling. PLoS Pathog. 2008, 4, e1000233. [Google Scholar] [CrossRef]
- Bjarke Olsen, P.; Klemm, P. Localization of promoters in the fim gene cluster and the effect of H-NS on the transcription of fimB and fimE. FEMS Microbiol. Lett. 1994, 116, 95–100. [Google Scholar] [CrossRef]
- Abraham, J.M.; Freitag, C.S.; Clements, J.R.; Eisenstein, B.I. An invertible element of DNA controls phase variation of type 1 fimbriae of Escherichia coli. Proc. Natl. Acad. Sci. USA 1985, 82, 5724–5727. [Google Scholar] [CrossRef] [Green Version]
- Wullt, B.; Bergsten, G.; Connell, H.; Röllano, P.; Gebretsadik, N.; Hull, R.; Svanborg, C. P fimbriae enhance the early establishment of Escherichia coli in the human urinary tract. Mol. Microbiol. 2000, 38, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.; Mobley, H. Role of P-fimbrial-mediated adherence in pyelonephritis and persistence of uropathogenic Escherichia coli (UPEC) in the mammalian kidney. Kidney Int. 2007, 72, 19–25. [Google Scholar] [CrossRef]
- Lund, B.; Lindberg, F.; Marklund, B.-I.; Normark, S. The PapG protein is the alpha-D-galactopyranosyl-(1----4)-beta-D-galactopyranose-binding adhesin of uropathogenic Escherichia coli. Proc. Natl. Acad. Sci. USA 1987, 84, 5898–5902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.J.; Mulvey, M.A.; Schilling, J.D.; Pinkner, J.S.; Hultgren, S.J. Type 1 pilus-mediated bacterial invasion of bladder epithelial cells. EMBO J. 2000, 19, 2803–2812. [Google Scholar] [CrossRef]
- Song, J.; Bishop, B.L.; Li, G.; Grady, R.; Stapleton, A.; Abraham, S.N. TLR4-mediated expulsion of bacteria from infected bladder epithelial cells. Proc. Natl. Acad. Sci. USA 2009, 106, 14966–14971. [Google Scholar] [CrossRef] [Green Version]
- Justice, S.S.; Hung, C.; Theriot, J.A.; Fletcher, D.A.; Anderson, G.G.; Footer, M.J.; Hultgren, S.J. Differentiation and developmental pathways of uropathogenic Escherichia coli in urinary tract pathogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 1333–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mysorekar, I.U.; Hultgren, S.J. Mechanisms of uropathogenic Escherichia coli persistence and eradication from the urinary tract. Proc. Natl. Acad. Sci. USA 2006, 103, 14170–14175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilling, J.D.; Mulvey, M.A.; Vincent, C.D.; Lorenz, R.G.; Hultgren, S.J. Bacterial invasion augments epithelial cytokine responses to Escherichia coli through a lipopolysaccharide-dependent mechanism. J. Immunol. 2001, 166, 1148–1155. [Google Scholar] [CrossRef] [Green Version]
- Wiles, T.J.; Kulesus, R.R.; Mulvey, M.A. Origins and virulence mechanisms of uropathogenic Escherichia coli. Exp. Mol. Pathol. 2008, 85, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Bien, J.; Sokolova, O.; Bozko, P. Role of uropathogenic Escherichia coli virulence factors in development of urinary tract infection and kidney damage. Int. J. Nephrol. 2012, 2012, 681473. [Google Scholar] [CrossRef] [Green Version]
- Visvikis, O.; Boyer, L.; Torrino, S.; Doye, A.; Lemonnier, M.; Lorès, P.; Rolando, M.; Flatau, G.; Mettouchi, A.; Bouvard, D. Escherichia coli producing CNF1 toxin hijacks Tollip to trigger Rac1-dependent cell invasion. Traffic 2011, 12, 579–590. [Google Scholar] [CrossRef]
- Mills, M.; Meysick, K.C.; O’Brien, A.D. Cytotoxic necrotizing factor type 1 of uropathogenic Escherichia coli kills cultured human uroepithelial 5637 cells by an apoptotic mechanism. Infect. Immun. 2000, 68, 5869–5880. [Google Scholar] [CrossRef]
- Miraglia, A.G.; Travaglione, S.; Meschini, S.; Falzano, L.; Matarrese, P.; Quaranta, M.G.; Viora, M.; Fiorentini, C.; Fabbri, A. Cytotoxic necrotizing factor 1 prevents apoptosis via the Akt/IκB kinase pathway: Role of nuclear factor-κB and Bcl-2. Mol. Biol. Cell 2007, 18, 2735–2744. [Google Scholar]
- Guyer, D.M.; Henderson, I.R.; Nataro, J.P.; Mobley, H.L. Identification of sat, an autotransporter toxin produced by uropathogenic Escherichia coli. Mol. Microbiol. 2000, 38, 53–66. [Google Scholar] [CrossRef]
- Guyer, D.M.; Radulovic, S.; Jones, F.-E.; Mobley, H.L. Sat, the secreted autotransporter toxin of uropathogenic Escherichia coli, is a vacuolating cytotoxin for bladder and kidney epithelial cells. Infect. Immun. 2002, 70, 4539–4546. [Google Scholar] [CrossRef] [Green Version]
- Liévin-Le Moal, V.; Comenge, Y.; Ruby, V.; Amsellem, R.; Nicolas, V.; Servin, A.L. Secreted autotransporter toxin (Sat) triggers autophagy in epithelial cells that relies on cell detachment. Cell Microbiol. 2011, 13, 992–1013. [Google Scholar] [CrossRef] [PubMed]
- Henderson, I.R.; Czeczulin, J.; Eslava, C.; Noriega, F.; Nataro, J.P. Characterization of Pic, a secreted protease of Shigella flexneri and enteroaggregative Escherichia coli. Infect. Immun. 1999, 67, 5587–5596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, K.B.; Totsika, M.; Moriel, D.G.; Lo, A.W.; Yang, J.; Wurpel, D.J.; Rossiter, A.E.; Strugnell, R.A.; Henderson, I.R.; Ulett, G.C. Molecular characterization of the vacuolating autotransporter toxin in uropathogenic Escherichia coli. J. Bacteriol. 2016, 198, 1487–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz, J.M.; Dozois, C.M.; Avelar-González, F.J.; Hernández-Cuellar, E.; Pokharel, P.; de Santiago, A.S.; Guerrero-Barrera, A.L. The vacuolating autotransporter toxin (vat) of Escherichia coli causes cell cytoskeleton changes and produces non-lysosomal vacuole formation in bladder epithelial cells. Front. Cell Infect. Microbiol. 2020, 10, 299. [Google Scholar] [CrossRef]
- Jones, R.L.; Peterson, C.M.; Grady, R.W.; Cerami, A. Low molecular weight iron-binding factor from mammalian tissue that potentiates bacterial growth. J. Exp. Med. 1980, 151, 418–428. [Google Scholar] [CrossRef]
- Stojiljkovic, I.; Perkins-Balding, D. Processing of heme and heme-containing proteins by bacteria. DNA Cell Biol. 2002, 21, 281–295. [Google Scholar]
- Hagan, E.C.; Mobley, H.L. Haem acquisition is facilitated by a novel receptor Hma and required by uropathogenic Escherichia coli for kidney infection. Mol. Microbiol. 2009, 71, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Henderson, J.P.; Crowley, J.R.; Pinkner, J.S.; Walker, J.N.; Tsukayama, P.; Stamm, W.E.; Hooton, T.M.; Hultgren, S.J. Quantitative metabolomics reveals an epigenetic blueprint for iron acquisition in uropathogenic Escherichia coli. PLoS Pathog. 2009, 5, e1000305. [Google Scholar] [CrossRef]
- Raffatellu, M.; George, M.D.; Akiyama, Y.; Hornsby, M.J.; Nuccio, S.-P.; Paixao, T.A.; Butler, B.P.; Chu, H.; Santos, R.L.; Berger, T. Lipocalin-2 resistance confers an advantage to Salmonella enterica serotype Typhimurium for growth and survival in the inflamed intestine. Cell Host Microbe 2009, 5, 476–486. [Google Scholar] [CrossRef]
- Valdebenito, M.; Crumbliss, A.L.; Winkelmann, G.; Hantke, K. Environmental factors influence the production of enterobactin, salmochelin, aerobactin, and yersiniabactin in Escherichia coli strain Nissle 1917. Int. J. Med. Microbiol. 2006, 296, 513–520. [Google Scholar] [CrossRef]
- Chaturvedi, K.S.; Hung, C.S.; Crowley, J.R.; Stapleton, A.E.; Henderson, J.P. The siderophore yersiniabactin binds copper to protect pathogens during infection. Nat. Chem. Biol. 2012, 8, 731–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabri, M.; Houle, S.; Dozois, C.M. Roles of the extraintestinal pathogenic Escherichia coli ZnuACB and ZupT zinc transporters during urinary tract infection. Infect. Immun. 2009, 77, 1155–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, A.S.; Kim, K.S.; Wright, D.C.; Sadoff, J.C.; Gemski, P. Role of lipopolysaccharide and capsule in the serum resistance of bacteremic strains of Escherichia coli. J. Infect. Dis. 1986, 154, 497–503. [Google Scholar] [CrossRef]
- O’brien, V.P.; Hannan, T.J.; Nielsen, H.V.; Hultgren, S.J. Drug and vaccine development for the treatment and prevention of urinary tract infections. Urin. Tract Infect. Mol. Pathog. Clin. Manag. 2017, 4, 589–646. [Google Scholar]
- Mortazavi-Tabatabaei, S.A.R.; Ghaderkhani, J.; Nazari, A.; Sayehmiri, K.; Sayehmiri, F.; Pakzad, I. Pattern of antibacterial resistance in urinary tract infections: A systematic review and meta-analysis. Int. J. Prev. Med. 2019, 10, 169. [Google Scholar] [CrossRef]
- Kot, B. Antibiotic Resistance Among Uropathogenic. Pol. J. Microbiol. 2019, 68, 403–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Target Product Profiles for Oral Therapy of Urinary Tract Infections; World Health Organization: Geneva, Switzerland, 2020.
- Aarestrup, F.M.; Wiuff, C.; Mølbak, K.r.; Threlfall, E.J. Is it time to change fluoroquinolone breakpoints for Salmonella spp.? Antimicrob. Agents Chemother. 2003, 47, 827–829. [Google Scholar] [CrossRef] [Green Version]
- Karam, M.R.A.; Habibi, M.; Bouzari, S. Urinary tract infection: Pathogenicity, antibiotic resistance and development of effective vaccines against Uropathogenic Escherichia coli. Mol. Immunol. 2019, 108, 56–67. [Google Scholar] [CrossRef]
- Hickling, D.R.; Nitti, V.W. Management of recurrent urinary tract infections in healthy adult women. Rev. Urol. 2013, 15, 41. [Google Scholar]
- Prát, V.; Matousovic, K.; Horcicková, M.; Hatala, M.; Milotová, Z. Prevention of recurrent urinary infections using Solco Urovac, a polymicrobial vaccine. Cas. Lek. Ceskych 1989, 128, 1106–1109. [Google Scholar]
- Kochiashvili, D.; Khuskivadze, A.; Kochiashvili, G.; Koberidze, G.; Kvakhajelidze, V. Role of the bacterial vaccine Solco-Urovac® in treatment and prevention of recurrent urinary tract infections of bacterial origin. Georgian Med. News 2014, 231, 11–16. [Google Scholar]
- Hopkins, W.J.; Elkahwaji, J.; Beierle, L.M.; Leverson, G.E.; Uehling, D.T. Vaginal mucosal vaccine for recurrent urinary tract infections in women: Results of a phase 2 clinical trial. J. Urol. 2007, 177, 1349–1353. [Google Scholar] [CrossRef] [PubMed]
- Wade, D.; Cooper, J.; Derry, F.; Taylor, J. Uro-Vaxom® versus placebo for the prevention of recurrent symptomatic urinary tract infections in participants with chronic neurogenic bladder dysfunction: A randomised controlled feasibility study. Trials 2019, 20, 223. [Google Scholar] [CrossRef] [PubMed]
- Trial, M.D.-B. Uro-Vaxom@ and the Management of Recurrent Urinary Tract Infection in Adults: A Randomized Multicenter Double-Blind Trial. Eur. Urol. 1994, 26, 137–140. [Google Scholar]
- Cruz, F.; Dambros, M.; Naber, K.G.; Bauer, H.W.; Cozma, G. Recurrent urinary tract infections: Uro-Vaxom®, a new alternative. Eur. Urol. Suppl. 2009, 8, 762–768. [Google Scholar] [CrossRef]
- Kim, K.S.; Kim, J.-Y.; Jeong, I.G.; Paick, J.-S.; Son, H.; Lim, D.J.; Shim, H.B.; Park, W.H.; Jung, H.C.; Choo, M.-S. A prospective multi-center trial of Escherichia coli extract for the prophylactic treatment of patients with chronically recurrent cystitis. J. Korean Med. Sci. 2010, 25, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Magistro, G.; Stief, C.G. Vaccine development for urinary tract infections: Where do we stand? Eur. Urol. Focus 2019, 5, 39–41. [Google Scholar] [CrossRef]
- Marinova, S.; Nenkov, P.; Markova, R.; Nikolaeva, S.; Kostadinova, R.; Mitov, I.; Vretenarska, M. Cellular and humoral systemic and mucosal immune responses stimulated by an oral polybacterial immunomodulator in patients with chronic urinary tract infections. Int. J. Immunopathol. Pharmacol. 2005, 18, 457–473. [Google Scholar] [CrossRef] [Green Version]
- Del Bino, L.; Østerlid, K.E.; Wu, D.-Y.; Nonne, F.; Romano, M.R.; Codée, J.; Adamo, R. Synthetic Glycans to Improve Current Glycoconjugate Vaccines and Fight Antimicrobial Resistance. Chem. Rev. 2022, 122, 15672–15716. [Google Scholar] [CrossRef]
- Billips, B.K.; Yaggie, R.E.; Cashy, J.P.; Schaeffer, A.J.; Klumpp, D.J. A live-attenuated vaccine for the treatment of urinary tract infection by uropathogenic Escherichia coli. J. Infect. Dis. 2009, 200, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.A.; Beanan, J.M.; Olson, R.; Genagon, S.A.; MacDonald, U.; Cope, J.J.; Davidson, B.A.; Johnston, B.; Johnson, J.R. A killed, genetically engineered derivative of a wild-type extraintestinal pathogenic E. coli strain is a vaccine candidate. Vaccine 2007, 25, 3859–3870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Dobbelsteen, G.P.; Faé, K.C.; Serroyen, J.; van den Nieuwenhof, I.M.; Braun, M.; Haeuptle, M.A.; Sirena, D.; Schneider, J.; Alaimo, C.; Lipowsky, G. Immunogenicity and safety of a tetravalent E. coli O-antigen bioconjugate vaccine in animal models. Vaccine 2016, 34, 4152–4160. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, V.S.; Himpsl, S.D.; Smith, S.N.; Sarkissian, C.A.; Mike, L.A.; Stocki, J.A.; Sintsova, A.; Alteri, C.J.; Mobley, H.L. Optimization of an experimental vaccine to prevent Escherichia coli urinary tract infection. MBio 2020, 11, e00555-20. [Google Scholar] [CrossRef] [PubMed]
- Huttner, A.; Hatz, C.; van den Dobbelsteen, G.; Abbanat, D.; Hornacek, A.; Frölich, R.; Dreyer, A.M.; Martin, P.; Davies, T.; Fae, K. Safety, immunogenicity, and preliminary clinical efficacy of a vaccine against extraintestinal pathogenic Escherichia coli in women with a history of recurrent urinary tract infection: A randomised, single-blind, placebo-controlled phase 1b trial. Lancet Infect. Dis. 2017, 17, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.B.; Abbanat, D.; Spiessens, B.; Go, O.; Haazen, W.; Rosa, T.d.; Fae, K.; Poolman, J.; Thoelen, S.; Palacios, P.I.d. 2712. Safety and Immunogenicity of two Doses of ExPEC4V Vaccine Against Extraintestinal Pathogenic Escherichia coli Disease in Healthy Adult Participants. Open Forum Infect. Dis. 2019, 6, S954. [Google Scholar] [CrossRef] [Green Version]
- Frenck, R.W., Jr.; Ervin, J.; Chu, L.; Abbanat, D.; Spiessens, B.; Go, O.; Haazen, W.; van den Dobbelsteen, G.; Poolman, J.; Thoelen, S. Safety and immunogenicity of a vaccine for extra-intestinal pathogenic Escherichia coli (ESTELLA): A phase 2 randomised controlled trial. Lancet Infect. Dis. 2019, 19, 631–640. [Google Scholar] [CrossRef]
- Hasanzadeh, S.; Habibi, M.; Shokrgozar, M.A.; Ahangari Cohan, R.; Ahmadi, K.; Asadi Karam, M.R.; Bouzari, S. In silico analysis and in vivo assessment of a novel epitope-based vaccine candidate against uropathogenic Escherichia coli. Sci. Rep. 2020, 10, 16258. [Google Scholar] [CrossRef]
- Karam, M.R.A.; Oloomi, M.; Mahdavi, M.; Habibi, M.; Bouzari, S. Assessment of immune responses of the flagellin (FliC) fused to FimH adhesin of uropathogenic Escherichia coli. Mol. Immunol. 2013, 54, 32–39. [Google Scholar] [CrossRef]
- Karam, M.R.A.; Oloomi, M.; Mahdavi, M.; Habibi, M.; Bouzari, S. Vaccination with recombinant FimH fused with flagellin enhances cellular and humoral immunity against urinary tract infection in mice. Vaccine 2013, 31, 1210–1216. [Google Scholar] [CrossRef]
- Savar, N.S.; Jahanian-Najafabadi, A.; Mahdavi, M.; Shokrgozar, M.A.; Jafari, A.; Bouzari, S. In silico and in vivo studies of truncated forms of flagellin (FliC) of enteroaggregative Escherichia coli fused to FimH from uropathogenic Escherichia coli as a vaccine candidate against urinary tract infections. J. Biotechnol. 2014, 175, 31–37. [Google Scholar] [CrossRef]
- Karam, M.R.A.; Habibi, M.; Bouzari, S. Use of flagellin and cholera toxin as adjuvants in intranasal vaccination of mice to enhance protective immune responses against uropathogenic Escherichia coli antigens. Biologicals 2016, 44, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, C.N.; Hultgren, S.J. Adhesive pili in UTI pathogenesis and drug development. Pathogens 2016, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.A.; McFadden, C.D.; Carlino-MacDonald, U.B.; Beanan, J.M.; Olson, R.; Wilding, G.E. TheSiderophore Receptor IroN of Extraintestinal Pathogenic Escherichia coli Is a Potential VaccineCandidate. Infect. Immun. 2003, 71, 7164–7169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habibi, M.; Karam, M.R.A.; Bouzari, S. Evaluation of prevalence, immunogenicity and efficacy of FyuA iron receptor in uropathogenic Escherichia coli isolates as a vaccine target against urinary tract infection. Microb. Pathog. 2017, 110, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Brumbaugh, A.R.; Smith, S.N.; Mobley, H.L. Immunization with the yersiniabactin receptor, FyuA, protects against pyelonephritis in a murine model of urinary tract infection. Infect. Immun. 2013, 81, 3309–3316. [Google Scholar] [CrossRef] [Green Version]
- Wieser, A.; Romann, E.; Magistro, G.; Hoffmann, C.; Nörenberg, D.; Weinert, K.; Schubert, S.r. A multiepitope subunit vaccine conveys protection against extraintestinal pathogenic Escherichia coli in mice. Infect. Immun. 2010, 78, 3432–3442. [Google Scholar] [CrossRef] [Green Version]
- Wieser, A.; Magistro, G.; Nörenberg, D.; Hoffmann, C.; Schubert, S. First multi-epitope subunit vaccine against extraintestinal pathogenic Escherichia coli delivered by a bacterial type-3 secretion system (T3SS). Int. J. Med. Microbiol. 2012, 302, 10–18. [Google Scholar] [CrossRef]
- Durant, L.; Metais, A.; Soulama-Mouze, C.; Genevard, J.-M.; Nassif, X.; Escaich, S. Identification of candidates for a subunit vaccine against extraintestinal pathogenic Escherichia coli. Infect. Immun. 2007, 75, 1916–1925. [Google Scholar] [CrossRef] [Green Version]
- Dietzman, D.E.; Fischer, G.W.; Schoenknecht, F.D. Neonatal Escherichia coli septicemia—Bacterial counts in blood. J. Pediatr. 1974, 85, 128–130. [Google Scholar] [CrossRef]
- Kim, K.S.; Itabashi, H.; Gemski, P.; Sadoff, J.; Warren, R.L.; Cross, A.S. The K1 capsule is the critical determinant in the development of Escherichia coli meningitis in the rat. J. Clin. Investig. 1992, 90, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Robbins, J.B.; McCracken, G.H., Jr.; Gotschlich, E.C.; Ørskov, F.; Ørskov, I.; Hanson, L.A. Escherichia coli K1 capsular polysaccharide associated with neonatal meningitis. N. Engl. J. Med. 1974, 290, 1216–1220. [Google Scholar] [CrossRef] [PubMed]
- Bonacorsi, S.; Clermont, O.; Houdouin, V.; Cordevant, C.; Brahimi, N.; Marecat, A.; Tinsley, C.; Nassif, X.; Lange, M.; Bingen, E. Molecular analysis and experimental virulence of French and North American Escherichia coli neonatal meningitis isolates: Identification of a new virulent clone. J. Infect. Dis. 2003, 187, 1895–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasadarao, N.V.; Wass, C.A.; Weiser, J.N.; Stins, M.F.; Huang, S.-H.; Kim, K.S. Outer membrane protein A of Escherichia coli contributes to invasion of brain microvascular endothelial cells. Infect. Immun. 1996, 64, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, C.-H.; Cai, M.; Shin, S.; Xie, Y.; Kim, K.-J.; Khan, N.A.; Di Cello, F.; Kim, K.S. Escherichia coli K1 RS218 interacts with human brain microvascular endothelial cells via type 1 fimbria bacteria in the fimbriated state. Infect. Immun. 2005, 73, 2923–2931. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.-H.; Tseng, Y.-T.; Maruvada, R.; Pearce, D.; Xie, Y.; Paul-Satyaseela, M.; Kim, K.S. NlpI contributes to Escherichia coli K1 strain RS218 interaction with human brain microvascular endothelial cells. Infect. Immun. 2010, 78, 3090–3096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.A.; Wang, Y.; Kim, K.J.; Chung, J.W.; Wass, C.A.; Kim, K.S. Cytotoxic necrotizing factor-1 contributes to Escherichia coli K1 invasion of the central nervous system. J. Biol. Chem. 2002, 277, 15607–15612. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.H.; Jong, A.Y. Cellular mechanisms of microbial proteins contributing to invasion of the blood–brain barrier: MicroReview. Cell Microbiol. 2001, 3, 277–287. [Google Scholar] [CrossRef]
- Cortes, M.A.; Gibon, J.; Chanteloup, N.K.; Moulin-Schouleur, M.; Gilot, P.; Germon, P. Inactivation of ibeA and ibeT results in decreased expression of type 1 fimbriae in extraintestinal pathogenic Escherichia coli strain BEN2908. Infect. Immun. 2008, 76, 4129–4136. [Google Scholar] [CrossRef] [Green Version]
- Fléchard, M.; Cortes, M.A.; Répérant, M.; Germon, P. New role for the ibeA gene in H2O2 stress resistance of Escherichia coli. J. Bacteriol. 2012, 194, 4550–4560. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhang, J.; Chen, Y.; Zhong, H.; Wang, H.; Li, J.; Zhu, G.; Xia, P.; Cui, L.; Li, J. ClbG in Avian Pathogenic Escherichia coli Contributes to Meningitis Development in a Mouse Model. Toxins 2021, 13, 546. [Google Scholar] [CrossRef]
- Zhu, N.; Liu, W.; Prakash, A.; Zhang, C.; Kim, K.S. Targeting E. coli invasion of the blood–brain barrier for investigating the pathogenesis and therapeutic development of E. coli meningitis. Cell Microbiol. 2020, 22, e13231. [Google Scholar] [CrossRef]
- Kim, K.S. Current concepts on the pathogenesis of Escherichia coli meningitis: Implications for therapy and prevention. Curr. Opin. Infect. Dis. 2012, 25, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Jiang, P.; Frøkiær, H.; Heegaard, P.M.; Thymann, T.; Sangild, P.T. Delayed development of systemic immunity in preterm pigs as a model for preterm infants. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Cross, A.; Zollinger, W.; Sadoff, J. Prevention and therapy of experimental Escherichia coli infection with monoclonal antibody. Infect. Immun. 1985, 50, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Finne, J.; Leinonen, M.; Mäkelä, P.H. Antigenic similarities between brain components and bacteria causing meningitis: Implications for vaccine development and pathogenesis. Lancet 1983, 322, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.M.; Maiden, M.C. A world without bacterial meningitis: How genomic epidemiology can inform vaccination strategy. F1000Research 2018, 7, 401. [Google Scholar] [CrossRef]
- Saif, Y. Diseases of Poultry; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Wei, S.; Morrison, M.; Yu, Z. Bacterial census of poultry intestinal microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef]
- Olsen, R.H.; Chadfield, M.; Christensen, J.P.; Scheutz, F.; Christensen, H.; Bisgaard, M. Clonality and virulence traits of Escherichia coli associated with haemorrhagic septicaemia in turkeys. Avian Pathol. 2011, 40, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Emery, D.; Nagaraja, K.V.; Shaw, D.; Newman, J.; White, D. Virulence factors of Escherichia coli associated with colisepticemia in chickens and turkeys. Avian Dis. 1992, 36, 504–511. [Google Scholar] [CrossRef]
- Norton, R.; Macklin, K.; McMurtrey, B. The association of various isolates of Escherichia coli from the United States with induced cellulitis and colibacillosis in young broiler chickens. Avian Pathol. 2000, 29, 571–574. [Google Scholar] [CrossRef]
- Ewers, C.; Janßen, T.; Kießling, S.; Philipp, H.-C.; Wieler, L.H. Molecular epidemiology of avian pathogenic Escherichia coli (APEC) isolated from colisepticemia in poultry. Vet. Microbiol. 2004, 104, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Feng, A.; Akter, S.; Leigh, S.A.; Wang, H.; Pharr, G.T.; Evans, J.; Branton, S.L.; Landinez, M.P.; Pace, L.; Wan, X.-F. Genomic diversity, pathogenicity and antimicrobial resistance of Escherichia coli isolated from poultry in the southern United States. BMC Microbiol. 2023, 23, 15. [Google Scholar] [CrossRef] [PubMed]
- La Ragione, R.; Sayers, A.; Woodward, M. The role of fimbriae and flagella in the colonization, invasion and persistence of Escherichia coli O78 [ratio] K80 in the day-old-chick model. Epidemiol. Infect. 2000, 124, 351–363. [Google Scholar] [CrossRef]
- Johnson, T.J.; Nolan, L.K. Pathogenomics of the virulence plasmids of Escherichia coli. Microbiol. Mol. Biol. Rev. 2009, 73, 750–774. [Google Scholar] [CrossRef] [PubMed]
- Lymberopoulos, M.H.; Houle, S.; Daigle, F.; Léveillé, S.; Brée, A.; Moulin-Schouleur, M.; Johnson, J.R.; Dozois, C.M. Characterization of Stg fimbriae from an avian pathogenic Escherichia coli O78: K80 strain and assessment of their contribution to colonization of the chicken respiratory tract. J. Bacteriol. 2006, 188, 6449–6459. [Google Scholar] [CrossRef] [Green Version]
- Aleksandrowicz, A.; Khan, M.M.; Sidorczuk, K.; Noszka, M.; Kolenda, R. Whatever makes them stick–Adhesins of avian pathogenic Escherichia coli. Vet. Microbiol. 2021, 257, 109095. [Google Scholar] [CrossRef]
- Nolan, M.F.; Malleret, G.; Lee, K.H.; Gibbs, E.; Dudman, J.T.; Santoro, B.; Yin, D.; Thompson, R.F.; Siegelbaum, S.A.; Kandel, E.R. The hyperpolarization-activated HCN1 channel is important for motor learning and neuronal integration by cerebellar Purkinje cells. Cell 2003, 115, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Foley, S.L.; Horne, S.M.; Giddings, C.W.; Robinson, M.; Nolan, L.K. Iss from a virulent avian Escherichia coli. Avian Dis. 2000, 44, 185–191. [Google Scholar] [CrossRef]
- Gao, Q.; Ye, Z.; Wang, X.; Mu, X.; Gao, S.; Liu, X. RstA is required for the virulence of an avian pathogenic Escherichia coli O2 strain E058. Infect. Genet. Evol. 2015, 29, 180–188. [Google Scholar] [CrossRef]
- Chakraborty, S.; Sivaraman, J.; Leung, K.Y.; Mok, Y.-K. Two-component PhoB-PhoR regulatory system and ferric uptake regulator sense phosphate and iron to control virulence genes in type III and VI secretion systems of Edwardsiella tarda. J. Biol. Chem. 2011, 286, 39417–39430. [Google Scholar] [CrossRef] [Green Version]
- Herren, C.D.; Mitra, A.; Palaniyandi, S.K.; Coleman, A.; Elankumaran, S.; Mukhopadhyay, S. The BarA-UvrY two-component system regulates virulence in avian pathogenic Escherichia coli O78: K80: H9. Infect. Immun. 2006, 74, 4900–4909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellata, M.; Dho-Moulin, M.; Dozois, C.M.; Curtiss, R., III; Brown, P.K.; Arné, P.; Brée, A.; Desautels, C.; Fairbrother, J.M. Role of virulence factors in resistance of avian pathogenic Escherichia coli to serum and in pathogenicity. Infect. Immun. 2003, 71, 536–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellata, M.; Ameiss, K.; Mo, H.; Curtiss, R., III. Characterization of the contribution to virulence of three large plasmids of avian pathogenic Escherichia coli χ7122 (O78: K80: H9). Infect. Immun. 2010, 78, 1528–1541. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Jia, X.; Wang, X.; Xiong, L.; Gao, S.; Liu, X. The avian pathogenic Escherichia coli O2 strain E058 carrying the defined aerobactin-defective iucD or iucDiutA mutation is less virulent in the chicken. Infect. Genet. Evol. 2015, 30, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Caza, M.; Lépine, F.; Milot, S.; Dozois, C.M. Specific roles of the iroBCDEN genes in virulence of an avian pathogenic Escherichia coli O78 strain and in production of salmochelins. Infect. Immun. 2008, 76, 3539–3549. [Google Scholar] [CrossRef] [Green Version]
- Perry, R.D.; Fetherston, J.D. Yersiniabactin iron uptake: Mechanisms and role in Yersinia pestis pathogenesis. Microbes Infect. 2011, 13, 808–817. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Wang, X.; Xu, H.; Xu, Y.; Ling, J.; Zhang, D.; Gao, S.; Liu, X. Roles of iron acquisition systems in virulence of extraintestinal pathogenic Escherichia coli: Salmochelin and aerobactin contribute more to virulence than heme in a chicken infection model. BMC Microbiol. 2012, 12, 143. [Google Scholar] [CrossRef] [Green Version]
- Sabri, M.; Caza, M.; Proulx, J.; Lymberopoulos, M.H.; Brée, A.; Moulin-Schouleur, M.; Curtiss, R., III; Dozois, C.M. Contribution of the SitABCD, MntH, and FeoB metal transporters to the virulence of avian pathogenic Escherichia coli O78 strain χ7122. Infect. Immun. 2008, 76, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Feng, Y.; Kariyawasam, S.; Tivendale, K.A.; Wannemuehler, Y.; Zhou, F.; Logue, C.M.; Miller, C.L.; Nolan, L.K. AatA is a novel autotransporter and virulence factor of avian pathogenic Escherichia coli. Infect. Immun. 2010, 78, 898–906. [Google Scholar] [CrossRef] [Green Version]
- Pokharel, P.; Díaz, J.M.; Bessaiah, H.; Houle, S.; Guerrero-Barrera, A.L.; Dozois, C.M. The serine protease autotransporters tagb, tagc, and sha from extraintestinal pathogenic Escherichia coli are internalized by human bladder epithelial cells and cause actin cytoskeletal disruption. Int. J. Mol. Sci. 2020, 21, 3047. [Google Scholar] [CrossRef]
- Dozois, C.M.; Dho-Moulin, M.; Brée, A.; Fairbrother, J.M.; Desautels, C.; Curtiss, R., III. Relationship between the Tsh autotransporter and pathogenicity of avian Escherichia coli and localization and analysis of the Tsh genetic region. Infect. Immun. 2000, 68, 4145–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parreira, V.; Gyles, C. A novel pathogenicity island integrated adjacent to the thrW tRNA gene of avian pathogenic Escherichia coli encodes a vacuolating autotransporter toxin. Infect. Immun. 2003, 71, 5087–5096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamarche, M.G.; Dozois, C.M.; Daigle, F.; Caza, M.; Curtiss, R., III; Dubreuil, J.D.; Harel, J. Inactivation of the pst system reduces the virulence of an avian pathogenic Escherichia coli O78 strain. Infect. Immun. 2005, 73, 4138–4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chouikha, I.; Germon, P.; Brée, A.; Gilot, P.; Moulin-Schouleur, M.; Schouler, C. A selC-associated genomic island of the extraintestinal avian pathogenic Escherichia coli strain BEN2908 is involved in carbohydrate uptake and virulence. J. Bacteriol. 2006, 188, 977–987. [Google Scholar] [CrossRef]
- de Paiva, J.B.; Leite, J.L.; da Silva, L.P.M.; Rojas, T.C.G.; de Pace, F.; Conceição, R.A.; Sperandio, V.; da Silveira, W.D. Influence of the major nitrite transporter NirC on the virulence of a Swollen Head Syndrome avian pathogenic E. coli (APEC) strain. Vet. Microbiol. 2015, 175, 123–131. [Google Scholar] [CrossRef]
- Johnson, T.J.; Siek, K.E.; Johnson, S.J.; Nolan, L.K. DNA sequence of a ColV plasmid and prevalence of selected plasmid-encoded virulence genes among avian Escherichia coli strains. J. Bacteriol. 2006, 188, 745–758. [Google Scholar] [CrossRef] [Green Version]
- Germon, P.; Chen, Y.-H.; He, L.; Blanco, J.E.; Bree, A.; Schouler, C.; Huang, S.-H.; Moulin-Schouleur, M. ibeA, a virulence factor of avian pathogenic Escherichia coli. Microbiology 2005, 151, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.; Drum, D.; Dai, Y.; Kim, J.; Sanchez, S.; Maurer, J.; Hofacre, C.; Lee, M. Impact of antimicrobial usage on antimicrobial resistance in commensal Escherichia coli strains colonizing broiler chickens. Appl. Environ. Microbiol. 2007, 73, 1404–1414. [Google Scholar] [CrossRef] [Green Version]
- Dho-Moulin, M.; Fairbrother, J.M. Avian pathogenic Escherichia coli (APEC). Vet. Res. 1999, 30, 299–316. [Google Scholar]
- Saif, Y.; Barnes, H.; Glisson, J.; Fadly, A.; McDougald, L.; Swayne, D. Diseases of Poultry, 11th ed.; Iowa State Press: Ames, IA, USA, 2003. [Google Scholar]
- McPeake, S.; Smyth, J.; Ball, H. Characterisation of avian pathogenic Escherichia coli (APEC) associated with colisepticaemia compared to faecal isolates from healthy birds. Vet. Microbiol. 2005, 110, 245–253. [Google Scholar] [CrossRef]
- Mombarg, M.; Bouzoubaa, K.; Andrews, S.; Vanimisetti, H.B.; Rodenberg, J.; Karaca, K. Safety and efficacy of an aroA-deleted live vaccine against avian colibacillosis in a multicentre field trial in broilers in Morocco. Avian Pathol. 2014, 43, 276–281. [Google Scholar] [CrossRef]
- Filho, T.F.; Fávaro, C., Jr.; Ingberman, M.; Beirão, B.C.; Inoue, A.; Gomes, L.; Caron, L.F. Effect of spray Escherichia coli vaccine on the immunity of poultry. Avian Dis. 2013, 57, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Tavakkoli, H.; Golchin, M.; Ghanbarpour, R.; Amanollahi, S. Efficacy and safety of Poulvac E. coli vaccine in broiler chickens challenged with E. coli serotype O78 and an acute field isolate. Comp. Clin. Pathol. 2018, 27, 1629–1636. [Google Scholar] [CrossRef]
- Koutsianos, D.; Gantelet, H.; Franzo, G.; Lecoupeur, M.; Thibault, E.; Cecchinato, M.; Koutoulis, K.C. An assessment of the level of protection against colibacillosis conferred by several autogenous and/or commercial vaccination programs in conventional pullets upon experimental challenge. Vet. Sci. 2020, 7, 80. [Google Scholar] [CrossRef] [PubMed]
- Lozica, L.; Kabalin, A.E.; Dolenčić, N.; Vlahek, M.; Gottstein, Ž. Phylogenetic characterization of avian pathogenic Escherichia coli strains longitudinally isolated from broiler breeder flocks vaccinated with autogenous vaccine. Poult. Sci. 2021, 100, 101079. [Google Scholar] [CrossRef]
- Chen, H.; Ji, H.; Kong, X.; Lei, P.; Yang, Q.; Wu, W.; Jin, L.; Sun, D. Bacterial Ghosts-Based Vaccine and Drug Delivery Systems. Pharmaceutics 2021, 13, 1892. [Google Scholar] [CrossRef]
- Kathayat, D.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Avian pathogenic Escherichia coli (APEC): An overview of virulence and pathogenesis factors, zoonotic potential, and control strategies. Pathogens 2021, 10, 467. [Google Scholar] [CrossRef]
- Ebrahimi-Nik, H.; Bassami, M.R.; Mohri, M.; Rad, M.; Khan, M.I. Bacterial ghost of avian pathogenic E. coli (APEC) serotype O78: K80 as a homologous vaccine against avian colibacillosis. PLoS ONE 2018, 13, e0194888. [Google Scholar] [CrossRef] [Green Version]
- Soleymani, S.; Tavassoli, A.; Hashemi Tabar, G.; Kalidari, G.A.; Dehghani, H. Design, development, and evaluation of the efficacy of a nucleic acid-free version of a bacterial ghost candidate vaccine against avian pathogenic E. coli (APEC) O78: K80 serotype. Vet. Res. 2020, 51, 144. [Google Scholar] [CrossRef]
- Yaguchi, K.; Ohgitani, T.; Noro, T.; Kaneshige, T.; Shimizu, Y. Vaccination of chickens with liposomal inactivated avian pathogenic Escherichia coli (APEC) vaccine by eye drop or coarse spray administration. Avian Dis. 2009, 53, 245–249. [Google Scholar] [CrossRef]
- Stromberg, Z.R.; Van Goor, A.; Redweik, G.A.; Mellata, M. Characterization of spleen transcriptome and immunity against avian colibacillosis after immunization with recombinant attenuated Salmonella vaccine strains. Front. Vet. Sci. 2018, 5, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhari, A.A.; Matsuda, K.; Lee, J.H. Construction of an attenuated Salmonella delivery system harboring genes encoding various virulence factors of avian pathogenic Escherichia coli and its potential as a candidate vaccine for chicken colibacillosis. Avian Dis. 2013, 57, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Redweik, G.A.; Stromberg, Z.R.; Van Goor, A.; Mellata, M. Protection against avian pathogenic Escherichia coli and Salmonella Kentucky exhibited in chickens given both probiotics and live Salmonella vaccine. Poult. Sci. 2020, 99, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Li, J.; Zhao, Y.; Lin, H.; Liang, L.; Wang, M.; Liu, H.; Min, Y.; Gao, Y.; Yang, M. Exploiting bacterial outer membrane vesicles as a cross-protective vaccine candidate against avian pathogenic Escherichia coli (APEC). Microb. Cell Factories 2020, 19, 119. [Google Scholar] [CrossRef] [PubMed]
- Francis, D.H. Colibacillosis in pigs and its diagnosis. J. Swine Health Prod. 1999, 7, 241–244. [Google Scholar]
- Fairbrother, J.M.; Nadeau, É. Colibacillosis. Dis. Swine 2019, 807–834. [Google Scholar] [CrossRef]
- Kongsted, H.; Pedersen, K.; Hjulsager, C.K.; Larsen, L.E.; Pedersen, K.S.; Jorsal, S.E.; Bækbo, P. Diarrhoea in neonatal piglets: A case control study on microbiological findings. Porc. Health Manag. 2018, 4, 17. [Google Scholar] [CrossRef]
- Vidal, A.; Aguirre, L.; Seminati, C.; Tello, M.; Redondo, N.; Martín, M.; Darwich, L. Antimicrobial resistance profiles and characterization of Escherichia coli strains from cases of neonatal diarrhea in Spanish pig farms. Vet. Sci. 2020, 7, 48. [Google Scholar] [CrossRef]
- Luppi, A.; Gibellini, M.; Gin, T.; Vangroenweghe, F.; Vandenbroucke, V.; Bauerfeind, R.; Bonilauri, P.; Labarque, G.; Hidalgo, Á. Prevalence of virulence factors in enterotoxigenic Escherichia coli isolated from pigs with post-weaning diarrhoea in Europe. Porc. Health Manag. 2016, 2, 20. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, M.; Sugimoto, C.; Isayama, Y.; Kashiwazaki, M. Virulence factors in Escherichia coli isolated from piglets with neonatal and post-weaning diarrhea in Japan. Vet. Microbiol. 1987, 13, 291–300. [Google Scholar] [CrossRef]
- Schulz, L.L.; Tonsor, G.T. Assessment of the economic impacts of porcine epidemic diarrhea virus in the United States. J. Anim. Sci. 2015, 93, 5111–5118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairbrother, J.M.; Nadeau, É.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.-Y.; Chapman, T.; Trott, D.J.; Bettelheim, K.; Do, T.N.; Driesen, S.; Walker, M.J.; Chin, J. Comparative analysis of virulence genes, genetic diversity, and phylogeny of commensal and enterotoxigenic Escherichia coli isolates from weaned pigs. Appl. Environ. Microbiol. 2007, 73, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Noamani, B.N.; Fairbrother, J.M.; Gyles, C.L. Virulence genes of O149 enterotoxigenic Escherichia coli from outbreaks of postweaning diarrhea in pigs. Vet. Microbiol. 2003, 97, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.S.; Tan, L.K.; Ooi, P.T.; Yeo, C.C.; Thong, K.L. Prevalence and characterization of verotoxigenic-Escherichia coli isolates from pigs in Malaysia. BMC Vet. Res. 2013, 9, 109. [Google Scholar] [CrossRef] [Green Version]
- Gannon, V.; Gyles, C.L.; Friendship, R.W. Characteristics of verotoxigenic Escherichia coli from pigs. Can. J. Vet. Res. 1988, 52, 331. [Google Scholar] [PubMed]
- Dubreuil, J.D.; Isaacson, R.E.; Schifferli, D.M. Animal enterotoxigenic Escherichia coli. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Hartadi, E.B.; Effendi, M.H.; Plumeriastuti, H.; Sofiana, E.D.; Wibisono, F.M.; Hidayatullah, A.R. A review of enterotoxigenic Escherichia coli infection in piglets: Public health importance. Syst. Rev. Pharm. 2020, 11, 687–698. [Google Scholar]
- Osek, J. Clonal analysis of Escherichia coli strains isolated from pigs with post-weaning diarrhea by pulsed-field gel electrophoresis. FEMS Microbiol. Lett. 2000, 186, 327–331. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, J.H.; Hur, J.; Lee, J.H. Isolation of Escherichia coli from piglets in South Korea with diarrhea and characteristics of the virulence genes. Can. J. Vet. Res. 2010, 74, 59–64. [Google Scholar]
- Peng, Z.; Hu, Z.; Li, Z.; Zhang, X.; Jia, C.; Li, T.; Dai, M.; Tan, C.; Xu, Z.; Wu, B. Antimicrobial resistance and population genomics of multidrug-resistant Escherichia coli in pig farms in mainland China. Nat. Commun. 2022, 13, 1116. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, Q.; Cheng, Y.; Liu, R.; Zhao, R.; Wang, J.; Wang, Y.; Yang, S.; Chen, A. Effect of Bacterial Resistance of Escherichia coli From Swine in Large-Scale Pig Farms in Beijing. Front. Microbiol. 2022, 13, 820833. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, S.E.; Abia, A.L.K.; Amoako, D.G.; Perrett, K.; Bester, L.A.; Essack, S.Y. From farm-to-fork: E. coli from an intensive pig production system in South Africa shows high resistance to critically important antibiotics for human and animal use. Antibiotics 2021, 10, 178. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Paruch, L.; Chen, X.; Van Eerde, A.; Skomedal, H.; Wang, Y.; Liu, D.; Liu Clarke, J. Antibiotic application and resistance in swine production in China: Current situation and future perspectives. Front. Vet. Sci. 2019, 6, 136. [Google Scholar] [CrossRef] [Green Version]
- Schoenmakers, K. How China is getting its farmers to kick their antibiotics habit. Nature 2020, 586, S60. [Google Scholar] [CrossRef]
- Wang, Y.; Liao, J.; Mehmood, K.; Chang, Y.-F.; Tang, Z.; Zhang, H. Escherichia coli isolated in pigs, Guangdong, China: Emergence of extreme drug resistance (XDR) bacteria. J. Infect. 2020, 81, 318–356. [Google Scholar] [CrossRef]
- Abreu, R.; Rodríguez-Álvarez, C.; Lecuona, M.; Castro, B.; González, J.C.; Aguirre-Jaime, A.; Arias, Á. Increased antimicrobial resistance of MRSA strains isolated from pigs in Spain between 2009 and 2018. Vet. Sci. 2019, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Österberg, J.; Wingstrand, A.; Nygaard Jensen, A.; Kerouanton, A.; Cibin, V.; Barco, L.; Denis, M.; Aabo, S.; Bengtsson, B. Antibiotic resistance in Escherichia coli from pigs in organic and conventional farming in four European countries. PLoS ONE 2016, 11, e0157049. [Google Scholar] [CrossRef]
- Melkebeek, V.; Goddeeris, B.M.; Cox, E. ETEC vaccination in pigs. Vet. Immunol. Immunopathol. 2013, 152, 37–42. [Google Scholar] [CrossRef]
- Nadeau, É.; Fairbrother, J.; Zentek, J.; Bélanger, L.; Tremblay, D.; Tremblay, C.-L.; Röhe, I.; Vahjen, W.; Brunelle, M.; Hellmann, K. Efficacy of a single oral dose of a live bivalent E. coli vaccine against post-weaning diarrhea due to F4 and F18-positive enterotoxigenic E. coli. Vet. J. 2017, 226, 32–39. [Google Scholar] [CrossRef]
- Hur, J.; Stein, B.D.; Lee, J.H. A vaccine candidate for post-weaning diarrhea in swine constructed with a live attenuated Salmonella delivering Escherichia coli K88ab, K88ac, FedA, and FedF fimbrial antigens and its immune responses in a murine model. Can. J. Vet. Res. 2012, 76, 186–194. [Google Scholar]
- Ruan, X.; Zhang, W. Oral immunization of a live attenuated Escherichia coli strain expressing a holotoxin-structured adhesin–toxoid fusion (1FaeG-FedF-LTA2: 5LTB) protected young pigs against enterotoxigenic E. coli (ETEC) infection. Vaccine 2013, 31, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, Y.; Zhang, Z.; You, J.; Yang, Y.; Li, X. Protective immunity of a multivalent vaccine candidate against piglet diarrhea caused by enterotoxigenic Escherichia coli (ETEC) in a pig model. Vaccine 2018, 36, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Liu, M.; Casey, T.A.; Zhang, W. A tripartite fusion, FaeG-FedF-LT192A2: B, of enterotoxigenic Escherichia coli (ETEC) elicits antibodies that neutralize cholera toxin, inhibit adherence of K88 (F4) and F18 fimbriae, and protect pigs against K88ac/heat-labile toxin infection. Clin. Vaccine Immunol. 2011, 18, 1593–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubreuil, J.D. Pig vaccination strategies based on enterotoxigenic Escherichia coli toxins. Braz. J. Microbiol. 2021, 52, 2499–2509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, C.; Francis, D.H.; Fang, Y.; Knudsen, D.; Nataro, J.P.; Robertson, D.C. Genetic fusions of heat-labile (LT) and heat-stable (ST) toxoids of porcine enterotoxigenic Escherichia coli elicit neutralizing anti-LT and anti-STa antibodies. Infect. Immun. 2010, 78, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Matías, J.; Berzosa, M.; Pastor, Y.; Irache, J.M.; Gamazo, C. Maternal vaccination. immunization of sows during pregnancy against ETEC Infections. Vaccines 2017, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.; Lu, T.; Nandre, R.M.; Duan, Q.; Zhang, W. Immunogenicity characterization of genetically fused or chemically conjugated heat-stable toxin toxoids of enterotoxigenic Escherichia coli in mice and pigs. FEMS Microbiol. Lett. 2019, 366, fnz037. [Google Scholar] [CrossRef]
- Arimizu, Y.; Kirino, Y.; Sato, M.P.; Uno, K.; Sato, T.; Gotoh, Y.; Auvray, F.; Brugere, H.; Oswald, E.; Mainil, J.G. Large-scale genome analysis of bovine commensal Escherichia coli reveals that bovine-adapted E. coli lineages are serving as evolutionary sources of the emergence of human intestinal pathogenic strains. Genome Res. 2019, 29, 1495–1505. [Google Scholar] [CrossRef]
- Durso, L.M.; Smith, D.; Hutkins, R.W. Measurements of fitness and competition in commensal Escherichia coli and E. coli O157: H7 strains. Appl. Environ. Microbiol. 2004, 70, 6466–6472. [Google Scholar] [CrossRef] [Green Version]
- Haggard, D.L. Bovine enteric colibacillosis. Vet. Clin. N. Am. Food Anim. Pract. 1985, 1, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Bashahun, G.; Amina, A. Colibacillosis in calves: A review of literature. J. Anim. Sci. Vet. Med. 2017, 2, 62–71. [Google Scholar] [CrossRef]
- Harnett, N.M.; Gyles, C.L. Resistance to drugs and heavy metals, colicin production, and biochemical characteristics of selected bovine and porcine Escherichia coli strains. Appl. Environ. Microbiol. 1984, 48, 930–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigobelo, E.; Gamez, H.; Marin, J.M.; Macedo, C.; Ambrosin, J.; Ávila, F.A.D. Virulence factors of Escherichia coli isolated from diarrheic calves. Arq. Bras. Med. Veterinária E Zootec. 2006, 58, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Bush, L.J.; Staley, T. Absorption of colostral immunoglobulins in newborn calves. J. Dairy Sci. 1980, 63, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Acres, S.D. Enterotoxigenic Escherichia coli infections in newborn calves: A review. J. Dairy Sci. 1985, 68, 229–256. [Google Scholar] [CrossRef]
- Ercan, N.; Tuzcu, N.; Başbug, O.; Tuzcu, M.; Alim, A. Diagnostic value of serum procalcitonin, neopterin, and gamma interferon in neonatal calves with septicemic colibacillosis. J. Vet. Diagn. Investig. 2016, 28, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Besser, T.E.; Gay, C.C. Septicemic colibacillosis and failure of passive transfer of colostral immunoglobulin in calves. Vet. Clin. North America. Food Anim. Pract. 1985, 1, 445–459. [Google Scholar] [CrossRef]
- Bazeley, K. Investigation of diarrhoea in the neonatal calf. Practice 2003, 25, 152–159. [Google Scholar] [CrossRef]
- Moxley, R.A.; Smith, D.R. Attaching-effacing Escherichia coli infections in cattle. Vet. Clin. Food Anim. Pract. 2010, 26, 29–56. [Google Scholar] [CrossRef]
- Picco, N.Y.; Alustiza, F.E.; Bellingeri, R.V.; Grosso, M.C.; Motta, C.E.; Larriestra, A.J.; Vissio, C.; Tiranti, K.I.; Terzolo, H.R.; Moreira, A.R. Molecular screening of pathogenic Escherichia coli strains isolated from dairy neonatal calves in Cordoba province, Argentina. Rev. Argent. Microbiol. 2015, 47, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, C.; Kwon, D.; Chae, C. Prevalence of the enteroaggregative Escherichia coli heat-stable enterotoxin 1 gene and its relationship with fimbrial and enterotoxin genes in E. coli isolated from diarrheic piglets. J. Vet. Diagn. Investig. 2001, 13, 26–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldo, V.; Salogni, C.; Giovannini, S.; D’Incau, M.; Boniotti, M.B.; Birbes, L.; Pitozzi, A.; Formenti, N.; Grassi, A.; Pasquali, P. Pathogenicity of Shiga toxin type 2e Escherichia coli in pig colibacillosis. Front. Vet. Sci. 2020, 7, 545818. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil, J.D. Escherichia coli STb toxin and colibacillosis: Knowing is half the battle. FEMS Microbiol. Lett. 2008, 278, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Salvadori, M.R.; Valadares, G.F.; Leite, D.d.S.; Blanco, J.; Yano, T. Virulence factors of Escherichia coli isolated from calves with diarrhea in Brazil. Braz. J. Microbiol. 2003, 34, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Srivani, M.; Reddy, Y.N.; Subramanyam, K.; Reddy, M.R.; Rao, T.S. Prevalence and antimicrobial resistance pattern of Shiga toxigenic Escherichia coli in diarrheic buffalo calves. Vet. World 2017, 10, 774. [Google Scholar] [CrossRef] [Green Version]
- Algammal, A.M.; Hetta, H.F.; Batiha, G.E.; Hozzein, W.N.; El Kazzaz, W.M.; Hashem, H.R.; Tawfik, A.M.; El-Tarabili, R.M. Virulence-determinants and antibiotic-resistance genes of MDR-E. coli isolated from secondary infections following FMD-outbreak in cattle. Sci. Rep. 2020, 10, 19779. [Google Scholar] [CrossRef]
- Ali, A.; Liaqat, S.; Tariq, H.; Abbas, S.; Arshad, M.; Li, W.-J.; Ahmed, I. Neonatal calf diarrhea: A potent reservoir of multi-drug resistant bacteria, environmental contamination and public health hazard in Pakistan. Sci. Total Environ. 2021, 799, 149450. [Google Scholar] [CrossRef]
- Tadesse, N. Prevalence and Multidrug Resistance Profiles of Escherichia coli in Dairy Farms. Int. J. Vet. Sci. Res. 2020, 6, 142–147. [Google Scholar] [CrossRef]
- Fesseha, H.; Mathewos, M.; Aliye, S.; Mekonnen, E. Isolation and antibiogram of Escherichia coli O157: H7 from diarrhoeic calves in urban and peri-urban dairy farms of Hawassa town. Vet. Med. Sci. 2022, 8, 864–876. [Google Scholar] [CrossRef]
- de Verdier, K.; Nyman, A.; Greko, C.; Bengtsson, B. Antimicrobial resistance and virulence factors in Escherichia coli from Swedish dairy calves. Acta Vet. Scand. 2012, 54, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hariharan, H.; Coles, M.; Poole, D.; Page, R. Antibiotic resistance among enterotoxigenic Escherichia coli from piglets and calves from piglets and calves with diarrhea. Can. Vet. J. 2004, 45, 605. [Google Scholar] [PubMed]
- Bradley, A.J. Bovine mastitis: An evolving disease. Vet. J. 2002, 164, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lacasse, P. Mammary tissue damage during bovine mastitis: Causes and control. J. Anim. Sci. 2008, 86, 57–65. [Google Scholar] [CrossRef]
- Gomes, F.; Henriques, M. Control of bovine mastitis: Old and recent therapeutic approaches. Curr. Microbiol. 2016, 72, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Gerjets, I.; Kemper, N. Coliform mastitis in sows: A review. J. Swine Health Prod. 2009, 17, 97–105. [Google Scholar]
- Goulart, D.B.; Mellata, M. Escherichia coli Mastitis in Dairy Cattle: Etiology, Diagnosis, and Treatment Challenges. Front. Microbiol. 2022, 13, 928346. [Google Scholar] [CrossRef]
- Blum, S.E.; Goldstone, R.J.; Connolly, J.P.; Répérant-Ferter, M.; Germon, P.; Inglis, N.F.; Krifucks, O.; Mathur, S.; Manson, E.; Mclean, K. Postgenomics characterization of an essential genetic determinant of mammary pathogenic Escherichia coli. MBio 2018, 9, e00423-18. [Google Scholar] [CrossRef] [Green Version]
- Blum, S.E.; Heller, E.D.; Sela, S.; Elad, D.; Edery, N.; Leitner, G. Genomic and phenomic study of mammary pathogenic Escherichia coli. PLoS ONE 2015, 10, e0136387. [Google Scholar] [CrossRef] [Green Version]
- Shpigel, N.Y.; Elazar, S.; Rosenshine, I. Mammary pathogenic Escherichia coli. Curr. Opin. Microbiol. 2008, 11, 60–65. [Google Scholar] [CrossRef]
- Ali, T.; ur Rahman, S.; Zhang, L.; Shahid, M.; Han, D.; Gao, J.; Zhang, S.; Ruegg, P.L.; Saddique, U.; Han, B. Characteristics and genetic diversity of multi-drug resistant extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli isolated from bovine mastitis. Oncotarget 2017, 8, 90144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, V.; McClure, J.; Léger, D.; Keefe, G.; Scholl, D.; Morck, D.; Barkema, H. Antimicrobial resistance profiles of common mastitis pathogens on Canadian dairy farms. J. Dairy Sci. 2012, 95, 4319–4332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorović, D.; Velhner, M.; Grego, E.; Vidanović, D.; Milanov, D.; Krnjaić, D.; Kehrenberg, C. Molecular characterization of multidrug-resistant Escherichia coli isolates from bovine clinical mastitis and pigs in the Vojvodina Province, Serbia. Microb. Drug Resist. 2018, 24, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Tahar, S.; Nabil, M.M.; Safia, T.; Ngaiganam, E.P.; Omar, A.; Hafidha, C.; Hanane, Z.; ROLAIN, J.; Diene, S.M. Molecular characterization of multidrug-resistant Escherichia coli isolated from milk of dairy cows with clinical mastitis in Algeria. J. Food Prot. 2020, 83, 2173–2178. [Google Scholar] [CrossRef] [PubMed]
- Hogan, J.; Smith, K.; Todhunter, D.; Schoenberger, P. Field trial to determine efficacy of an Escherichia coli J5 mastitis vaccine. J. Dairy Sci. 1992, 75, 78–84. [Google Scholar] [CrossRef] [PubMed]
- McClure, A.M.; Christopher, E.E.; Wolff, W.; Fales, W.; Krause, G.F.; Miramonti, J. Effect of Re-17 mutant Salmonella typhimurium bacterin toxoid on clinical coliform mastitis. J. Dairy Sci. 1994, 77, 2272–2280. [Google Scholar] [CrossRef]
- Liu, X.; Sun, W.; Wu, N.; Rong, N.; Kang, C.; Jian, S.; Chen, C.; Chen, C.; Zhang, X. Synthesis of Escherichia coli OmpA oral nanoparticles and evaluation of immune functions against the major etiologic agent of cow mastitis. Vaccines 2021, 9, 304. [Google Scholar] [CrossRef]
- Ali, M.S.E.-D.; Mikhail, W.Z.A.; Salama, M.A.M.; Mohamed, Y. Impact of Offspring Sex and Dam’s Pre-partum Vaccination on Colostrum Composition and Blood Hormones in Egyptian Buffaloes. World 2021, 11, 51–59. [Google Scholar]
- Walle, K.V.; Vanrompay, D.; Cox, E. Bovine innate and adaptive immune responses against Escherichia coli O157: H7 and vaccination strategies to reduce faecal shedding in ruminants. Vet. Immunol. Immunopathol. 2013, 152, 109–120. [Google Scholar] [CrossRef]
- Fingermann, M.; Avila, L.; De Marco, M.B.; Vázquez, L.; Di Biase, D.N.; Müller, A.V.; Lescano, M.; Dokmetjian, J.C.; Fernández Castillo, S.; Pérez Quiñoy, J.L. OMV-based vaccine formulations against Shiga toxin producing Escherichia coli strains are both protective in mice and immunogenic in calves. Hum. Vaccines Immunother. 2018, 14, 2208–2213. [Google Scholar] [CrossRef] [Green Version]
- Ingelheim, B. Boehringer Ingelheim Launches Fencovis®, a New Vaccine to Prevent Calf Diarrhea. Available online: https://www.boehringer-ingelheim.com/animal-health/livestock/ruminants/vaccine-prevent-calf-diarrhea (accessed on 15 November 2022).
- Ingelheim, B. J-VAC® by Merial—Protects Dairy Cows from Coliform Mastitis. Available online: https://www.bi-vetmedica.com/species/cattle/products/j-vac (accessed on 13 November 2022).
- Ingelheim, B. Bovine E. coli Vaccine Products of the Company Against Mastitis. Available online: https://www.bi-vetmedica.com/species/cattle/products/Barguard.html (accessed on 13 November 2022).
- Wang, H.; Yuan, L.; Wang, T.; Cao, L.; Liu, F.; Song, J.; Zhang, Y. Construction of the waaF Subunit and DNA Vaccine Against Escherichia coli in Cow Mastitis and Preliminary Study on Their Immunogenicity. Front. Vet. Sci. 2022, 9, 877685. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, J.; Vidal, S.; Siel, D.; Caruffo, M.; Valdés, A.; Cabrera, G.; Lapierre, L.; Sáenz, L. Novel Proteoliposome-Based Vaccine against E. coli: A Potential New Tool for the Control of Bovine Mastitis. Animals 2022, 12, 2533. [Google Scholar] [CrossRef] [PubMed]





| Type of Vaccine | Component of Vaccine | Results/Observations/Outcomes | Animal Model (Year) | References |
|---|---|---|---|---|
| Antigen-Based Vaccine | Recombinant Mycobacterium smegmatis (Smeg) and Mycobacterium bovis BCG to express BfpA or intimin. | Yielded high titer of IgG and IgA antibodies in the serum of immunized mice. Mice immunized with recombinant BfpA showed TNF-α and INF-γ, and TNF-α only with recombinant intimin. | Mice immunized by oral gavage or intraperitoneal injection (2012). | [91] |
| Combination of purified recombinant EspA, Intimin, and Tir. | Showed protection of immunized cattle against O157 challenges. | Male Holstein-Friesian calves immunized orally (2010). | [97] | |
| Lactobacillus casei expressing intimin-β and immune-dominant isotopes of Int280. | Induced cellular and humoral responses in mice. Serum antibodies inhibited EPEC adhesion to epithelial cells in vitro. | Mice immunized intranasally (2008). | [98] | |
| Plant-based vaccine | Transgenic plants expressing intimin, and BfpA. | Proposed edible vaccines under strategic and regulation planning. | Mice immunized orally (2002). | [99] |
| Live-attenuated bacterial vaccine | Live attenuated ΔespFΔushA Citrobacter rodentium strain. | Oral administration in mice yielded efficient systemic and humoral immunity against C. rodentium virulence factors. | Mice immunized by oral gavage (2022). | [100] |
| Adjuvanted whole-cell vaccine | Cholera toxoid (CTB)-adjuvanted formalin-killed whole bacterial cell (EPEC). | 100% survival rate of Balb/C mice when challenged with EPEC. | Mice immunized intraperitoneally (2016). | [101] |
| Type of Vaccine | Component of Vaccine | Results/Observations/Outcomes | Animal Model/Phase (Year) | References |
|---|---|---|---|---|
| Attenuated bacteria-based vaccines | Attenuated ETEC E1392/75-2A ΔaroCΔompR and ETEC E1392/75-2AΔaroCΔompR ΔompC mutations | Significant yield in IgA and IgG and CS1 and CS3 specific antibodies. | Mice immunized intranasally (2001) | [179] |
| ETVAX (attenuated bacteria expressing CS6 in E. coli K-12 and CFA/I, CS3, CS5 in ETEC O78 toxin-negative) with LCTBA hybrid protein and mutated Heat-label (LT) | Currently in clinical trials (NCT02531802). High titers of fecal, jejunal and serum IgA and IgG in orally immunized humans. | Phase II clinical trials (2020) | [186,187] | |
| ACE527 ETEC complex (ACAM2022 (O141:H5, expressing CS5 and CS6), ACAM2025 (O39:H12, expressing CFA/I) and ACAM2027 (O71:H-, expressing CS2, CS3, and CS1) | 33–98% protection in reducing the duration of diarrhea in human clinical trials (NCT00901654). | Phase I clinical trials (2015) | [178,188] | |
| Attenuated ETEC strains expressing CFA/I, CS2 and CS3 and CS1, CS2, and CS3 generating ACAM2010, 2007, 2017 strains respectively | Elevated levels of IgA against CFA in orally immunized human volunteers. | Human volunteers, double-blind trials (2008) | [178] | |
| Attenuated ETEC strains expressing CS5, CS6, LT, ST and EAST1 generating ACAM2025, 2022, 2027 strains. | Double-blind placebo-controlled Phase II challenge trial. | Phase II clinical trials (2019) NCT01739231 | [189,190] | |
| Strains of Vibrio cholerae expressing CFA/I | Significantly yielded high levels of IgA and IgG titers in the serum of immunized mice. | Mice immunized orally (2008) | [191] | |
| Non-toxigenic E. coli expressing CS2, CS4, CS5, or CS6 and CFA/I | Significantly induced IgG+IgM and IgA antibodies CS6 in sera and feces, respectively, in immunized mice. | Oral immunization of mice (2010) | [192,193,194] | |
| Adhesin-based vaccines | Recombinant ETEC expressing two-partner secretion protein A (EtpA) | Less bacterial (ETEC) colonization in the gut of immunized mice. | Intranasal immunization of mice model (2009-2016) | [195] |
| CS21/LngA formulated with cholera toxin | Increased specific IgG and IgA in serum and feces and intestinal lavages, respectively. Reduced shedding in immunized mice. | Intranasal immunization of mice (2017) | [196] | |
| OMV-based vaccines | ETEC OMVs ΔmsbBΔeltA | Detoxified OMVs induced higher titers of IgG1, IgM, and IgA and significantly reduced wild-type ETEC colonization in immunized mice. | Mice immunized intranasally (2015) | [196] |
| Vibrio cholerae OMVs ΔmsbBΔctxABΔflaA expressing ETEC FliC and CFA/I | OMVs yielded higher titers of IgG1, IgM, and IgA and reduced wild-type ETEC colonization and spread in immunized mice. | Mice immunized intranasally (2015) | [197] | |
| Autotransporter-based vaccines | Recombinant Ag43 and pAT (autotransporters) | Significant increase in fecal IgA and partial protection against intestinal colonization of ETEC in immunized mice. | Mice immunized intranasally (2011) | [198] |
| Toxin-based vaccines | Heat-labile (LT) toxin using skin patches. | Yielded significant levels of anti-LT IgG and IgA in 97–100% of human volunteers. Currently, in Phase 2, the clinical trial complete phase (NCT00565461). | Phase II clinical trial (2020) | [199] |
| STaP13F-LTR192G toxoid fusion protein | Induced IgG-specific antibodies for LT and STa in serum and feces and IgA in feces in immunized mice. | Oral immunization of mice (2019) | [179] |
| Type of Vaccine | Component of Vaccine | Results/Observations/Outcomes/Animal Model/Phase (Year) | References |
|---|---|---|---|
| Live-attenuated vaccine | ShigETEC attenuated Shigella strain expressing ETEC antigens (LTB and detoxified version of ST) | Currently, phase I clinical trials yielded high titer IgG and IgA against bacterial lysates and anti-ETEC toxins (2022) | [185,232] |
| WRSs2 attenuated ΔvirG S. sonnei in which enterotoxin genes senA/senB are deleted. | Currently, phase II clinical trials, NCT04242264 (2021) | [233] | |
| WRSs3 attenuated ΔvirG S. sonnei in which senA/senB, and acetyl transferase genes msbB are deleted. | Currently, phase II clinical trials (2021) | [233] | |
| Shigella flexneri 2a, O antigen mutant (Δwzy) combined with E. coli LT mutant. | Provided cross-protective immunity against Shigella and ETEC. Pre-clinical stage (2018) | [234] | |
| The formalin-inactivated trivalent Shigella whole-cell vaccine | Phase II clinical trials (2022) | [224] | |
| Heat-killed multi serotype | Heat-killed cocktail of 6 strains of Shigella inactivated vaccine | Pre-clinical stage (2016) | [223] |
| Subunit vaccines | S4V-EPA, four-valent, O-antigen bioconjugates against S. sonnei, flexneri 3a and flexneri 6, and S. flexneri 2a. | Phase II clinical trials (2022) | [235] Limmatech |
| SF2a-TT15, S. flexneri 2a synthetic O-antigen conjugates against S. flexneri 2a | Phase II clinical trials (2022) | [236] | |
| InvaplexAR-DETOX, artificially detoxified S. flexneri 2a invasin complex with recombinant IpaB/IpaC | Phase I clinical trial (2021) | WRAIR, [233] | |
| ZF0901, Bivalent O-antigen glycoconjugate against S. flexneri 2a and S. sonnei | Phase III clinical trial (2021) | Beijing Zhifei Lvzhu biopharmaceuticals, [237] | |
| altSonflex 1-2-3, a four valent Shigella native outer membrane vesicle (OMVs) against S. flexneri 1b, 2a, 3a and S. sonnei. | Phase II clinical trial (2022) | [224] | |
| Adjuvanted whole-cell vaccine | Alum-adjuvanted and CTB-adjuvanted EIEC whole-cell vaccine | Higher IgG yield and immune response against EIEC and ETEC in orally immunized mice (2016) | [101] |
| Type of Vaccine | Component of Vaccine | Results/Observations/Outcomes | Animal Model (Year) | References |
|---|---|---|---|---|
| Adjuvanted enhanced vaccine | Intranasal immunization of mice using siderophore enterotoxin (Ent) conjugated with CTB. | Increased fecal antibodies against Ent and reduced AIEC colonization in immunized mice. | Orally immunized mice (2021) | [285] |
| CTB-Ent, immunization of mice. | Mucosal IgA against Ent and GlcEnt, protection from systemic infection and decreased AIEC and Crohn’s disease and colitis in mice. | Orally immunized mice (2022) | [286] | |
| Inhibition of FimH adhesin | Thiazolylaminomannosides and n-heptyl α-D-mannose based inhibition of AIEC LF82 adherence to colon tissue by blocking FimH | FimH blocker molecule EB8018/TAK-018 is under phase 2a clinical trial (NCT03943446) | Clinical trial (2020) | [262] |
| Probiotics, prebiotics, and postbiotics | Probiotics containing a portion of S. cerevisiae CNCM I-3856 | Known to prevent colitis induced by AIEC in the mouse model of Crohn’s disease. | Orally immunized mice (2018) | [287] |
| Probiotics Lactobacillus rhamnosus GG and Lactobacillus reuteri. | Known to reduce AIEC survival and growth. | Orally immunized mice (2018) | [288] | |
| Prebiotics containing long-chain arabinoxylans | Known to inhibit the mucin adhesion of AIEC. | Orally immunized mice (2020) | [289] | |
| Prebiotics containing insulin and galacto-oligosaccharides | Limit AIEC survival and growth | Orally immunized mice (2017) | [290] | |
| Postbiotics such as colicins E1 and E9 that are species-specific bacteriocins. | Known to kill intracellular, biofilm-forming and cell-adhering AIEC. | Mice model (2018) | [291] | |
| Fecal microbiota transplantation (FMT) | Restoration of normal intestinal flora to prevent CD and AIEC colonization | Placebo-controlled trials using FMT have shown improvements in patients with active disease. | Pre-clinical trial stage (2022) | [292] |
| Phage Therapy | LF82 bacteriophages that were able to replicate in ileal, colon samples and feces. | An oral dose of bacteriophages has inhibited AIEC strain LF82 colonization and colitis symptoms in the gut. Phase 1/2a clinical trial is ongoing. | Human volunteers (2015) | [293] Mount Sinai hospital |
| Five-phage cocktail against IBD | IBD suppression. Currently in Phase II clinical trial (NCT04737876). | Clinical trial (2022) | [294] |
| Type of Vaccine | Component of Vaccine | Results/Observations/Outcomes | Animal Model (Year) | References |
|---|---|---|---|---|
| Bacterial ghost of APEC+A106:D112 | Design of bacterial ghost of E. coli O78:K80 by making the porous cell wall | Aerosol-vaccinated chickens challenged with APEC O78:K80 had reduced air sac lesions and less death. Vaccinated chickens showed increased levels of IFNγ, IgA and IgY. | Broiler chicken (2018) | [445] |
| Nucleic acid-free bacterial ghost vaccine of E. coli O78:K80 by removing cytoplasmic content and nucleic acids. | Chickens vaccinated by injection or inhalation both showed humoral and cellular immune responses and cytokine responses. Challenge with O78:K80 showed lower lesion scores and bacterial numbers in the vaccinated group. | Broiler chicken (2020) | [446] | |
| Liposomal inactivated APEC vaccine | Liposomal inactivated avian pathogenic Escherichia coli (APEC) strain containing vaccine. | Chickens vaccinated via eye drops or coarse spray produced anti-LPS antibodies (IgG) in serum and IgA in oral mucus. | Broiler chicken (2009) | [447] |
| Recombinant attenuated Salmonella vaccine (RASV) | RASV-producing E. coli common pilus (ECP) and booster dose with a combination of RASV χ8025(pYA3337) and χ8025(pYA4428) or χ8025(pYA3337), RASV χ8025(pYA4428) carrying ecp operon genes. | Chickens were vaccinated orally and challenged with APEC O2 or O78 strain via air sac. Immunized chickens after vaccination showed significantly increased levels of serum (IgY) and intestinal (IgA) antibody. Challenged chicken showed partial protection against APEC. | White Leghorns chicken (2018) | [448] |
| RASV with Δlon, ΔcpxR, and ΔasdA16 and producing P-fimbriae, aerobactin receptor, and CS31A surface antigen of APEC. | Immunized chickens showed increased IgG and IgA antibodies. Chickens were challenged via the air-sac route and showed partial protection against virulent APEC. | Broiler chicken (2013) | [449] | |
| RASV plus commercial probiotics supplements | White Leghorn Chickens are given RASV, and probiotics elicited significant serum and mucosal antibodies. When challenged with APEC virulent strains showed lower bacterial loads and lesions of airsacculitis and pericarditis/perihepatitis. | White leghorn chicken (2020) | [450] | |
| Outer membrane vesicle (OMV) based vaccine | Purified OMVs from O1, O2, and O78 strains to develop a multi-serogroup vaccine (MOMVs). | Vaccinated chickens effectively yielded specific antibody responses against each OMV antigen. They also yielded significant cellular and humoral immune responses. Immunization with MOMVs showed 100%, 90% and 100% cross-protection against challenge by O1, O2 and O78 APEC strains. | Broiler chicken (2020) | [451] |
| Type of Vaccine | Component of Vaccine | Results/Observations/Outcomes/Year | References |
|---|---|---|---|
| ScourGuard® 4K | Cocktail of inactivated bovine rotavirus, coronavirus and E. coli bacterin | Vaccination of healthy pregnant cows prevented diarrheal disease in calves bovine ETEC bearing K99 pili, bovine rotavirus (serotypes G6 and G10), and coronavirus (2021) | [525] (Zoetis, USA) |
| ScourGuard® 4Kc | Cocktail of inactivated bovine rotavirus, coronavirus, Clostridium perfringens type C and E. coli bacterin-toxoid | Vaccination of healthy pregnant cows prevented diarrhea caused by bovine rotavirus (serotypes G6 and G10), coronavirus, ETEC and C. perfringens in calves given colostrum from vaccinated mother (2021) | [525] (Zoetis, USA) |
| Bolus and Dual-force gel | Contains passive antibodies against diarrheal pathogens | A single dose administered after birth protects calves from E. coli and coronavirus infection (2021) | First Defense |
| Tri-Shield First defense | Contains passive antibodies against diarrheal pathogens. Should be administered with maternal colostrum | A single dose administered after birth provides passive immunity against K99+ E. coli, coronavirus and rotavirus (2021) | First Defense |
| First Defense Technology | Hyper-immunized colostrum antibodies | Antibodies that neutralize E. coli and coronavirus and provide instant immunity (2021) | First Defense |
| Bioniche® vaccine | A Type III secretion system-based vaccine | Reduces E. coli O157:H7 (EHEC) growth and colonization in cattle (2013) | [526] Bioniche |
| Epitopix® vaccine | Siderophore receptor and porin protein (SRP) vaccine | Reduces E. coli O157:H7 (EHEC) growth and colonization in cattle (2018) | [527] |
| Fencovis® vaccine | Administered to pregnant cows to provide passive immunity to newborn calves via maternal colostrum | Active immunization of cows stimulates the development of antibodies against E. coli F5, rotavirus and coronavirus and prevents neonatal diarrhea (2022) | [528] |
| J-VAC® vaccine | Broad spectrum adjuvanted bacterin-toxoid | Prevents bovine mastitis caused by E. coli and endotoxemia caused by E. coli and Salmonella Typhimurium (2022) | [529] |
| Bar-Guard-99™ vaccine | Utilizes whole cell antibodies | Provide rapid passive immunity against E. coli K99 and diarrheal diseases (2022) | [530] |
| ENVIRACOR® J-5 vaccine | Bacterin-based vaccine | Controls clinical signs related to bovine mastitis caused by E. coli (2021) | Zoetis, USA |
| BOVILIS® J-5 vaccine | Endotoxin-based vaccine | Prevents milk loss, culling and death related to bovine mastitis caused by E. coli (2021) | MERK animal health |
| DNA and subunit-based vaccine | Lipopolysaccharide-based pcwaaF (DNA vaccine) and rwaaF (recombinant waaF subunit vaccine) | Greater IgG, IL-2, IL-4, and IFN-γ, and fecal sIgA. Mice survive better post-challenge with mastitis, causing E. coli (2022) | [531] |
| Proteo-liposome based vaccine | Proteo-liposome extracted from bovine mastitis clinical isolate (RM5870) | A significant level of IgG, IgG1 and IgG2a, IgA. Improved survival rates of mice post-challenge with E. coli causing mastitis. Reduced bacterial loads, inflammation, and tissue damage in mammary glands (2022) | [532] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pokharel, P.; Dhakal, S.; Dozois, C.M. The Diversity of Escherichia coli Pathotypes and Vaccination Strategies against This Versatile Bacterial Pathogen. Microorganisms 2023, 11, 344. https://doi.org/10.3390/microorganisms11020344
Pokharel P, Dhakal S, Dozois CM. The Diversity of Escherichia coli Pathotypes and Vaccination Strategies against This Versatile Bacterial Pathogen. Microorganisms. 2023; 11(2):344. https://doi.org/10.3390/microorganisms11020344
Chicago/Turabian StylePokharel, Pravil, Sabin Dhakal, and Charles M. Dozois. 2023. "The Diversity of Escherichia coli Pathotypes and Vaccination Strategies against This Versatile Bacterial Pathogen" Microorganisms 11, no. 2: 344. https://doi.org/10.3390/microorganisms11020344