

Exploiting Non-Conventional Yeasts for Low-Alcohol Beer Production

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Yeast Strains and Media
2.2. Initial Screening for NABLAB Production
2.3. EBC-Tubes Beer Fermentation
2.4. Wort and Beer Analyses
2.4.1. HPLC Analysis
2.4.2. Determination of Alcohols, Total Esters, and Total Aldehydes by GC-FID
2.4.3. Alcolyzer Plus
2.4.4. Sensory Analysis
2.4.5. Statistical Analysis
3. Results and Discussion
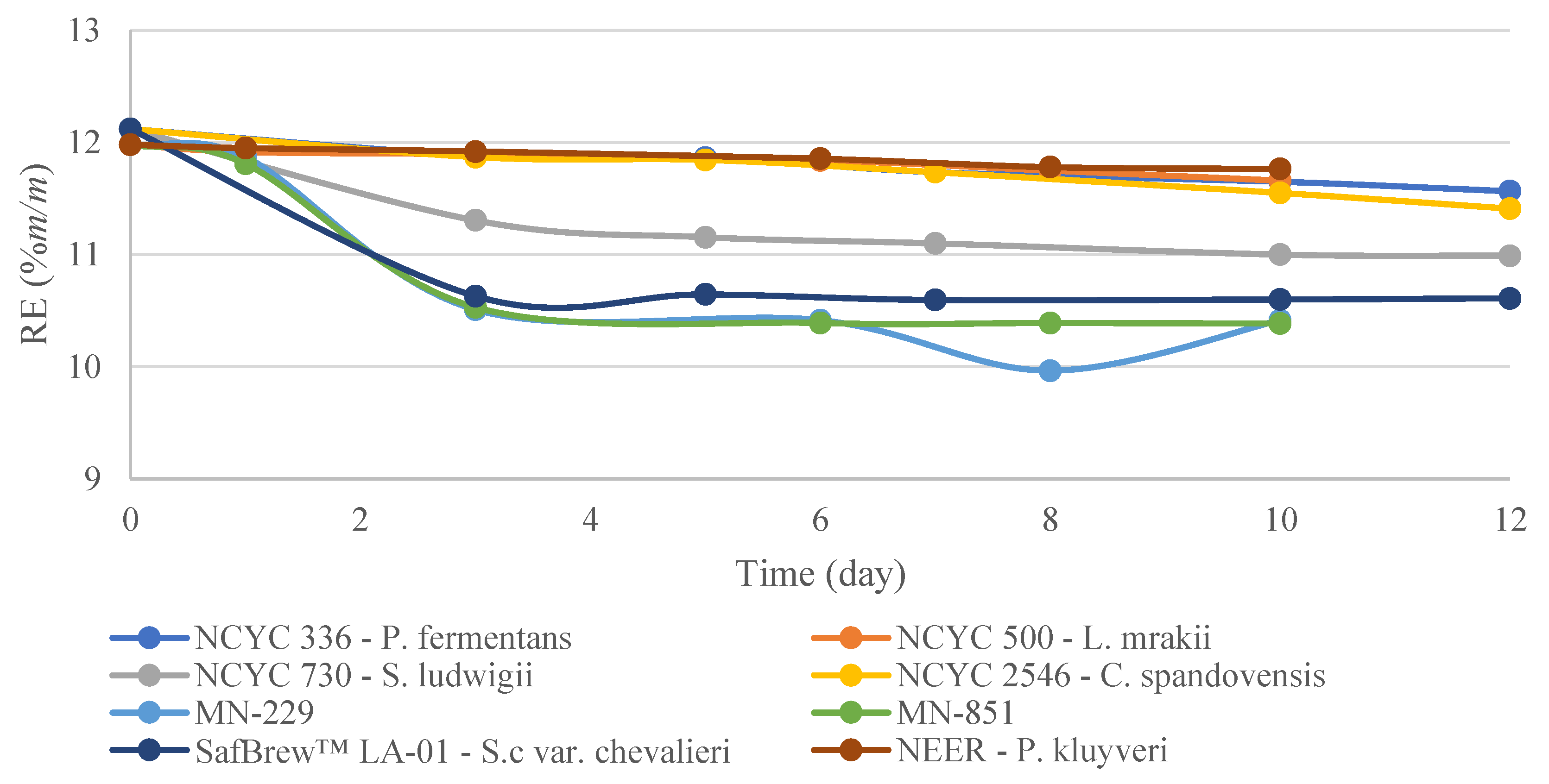
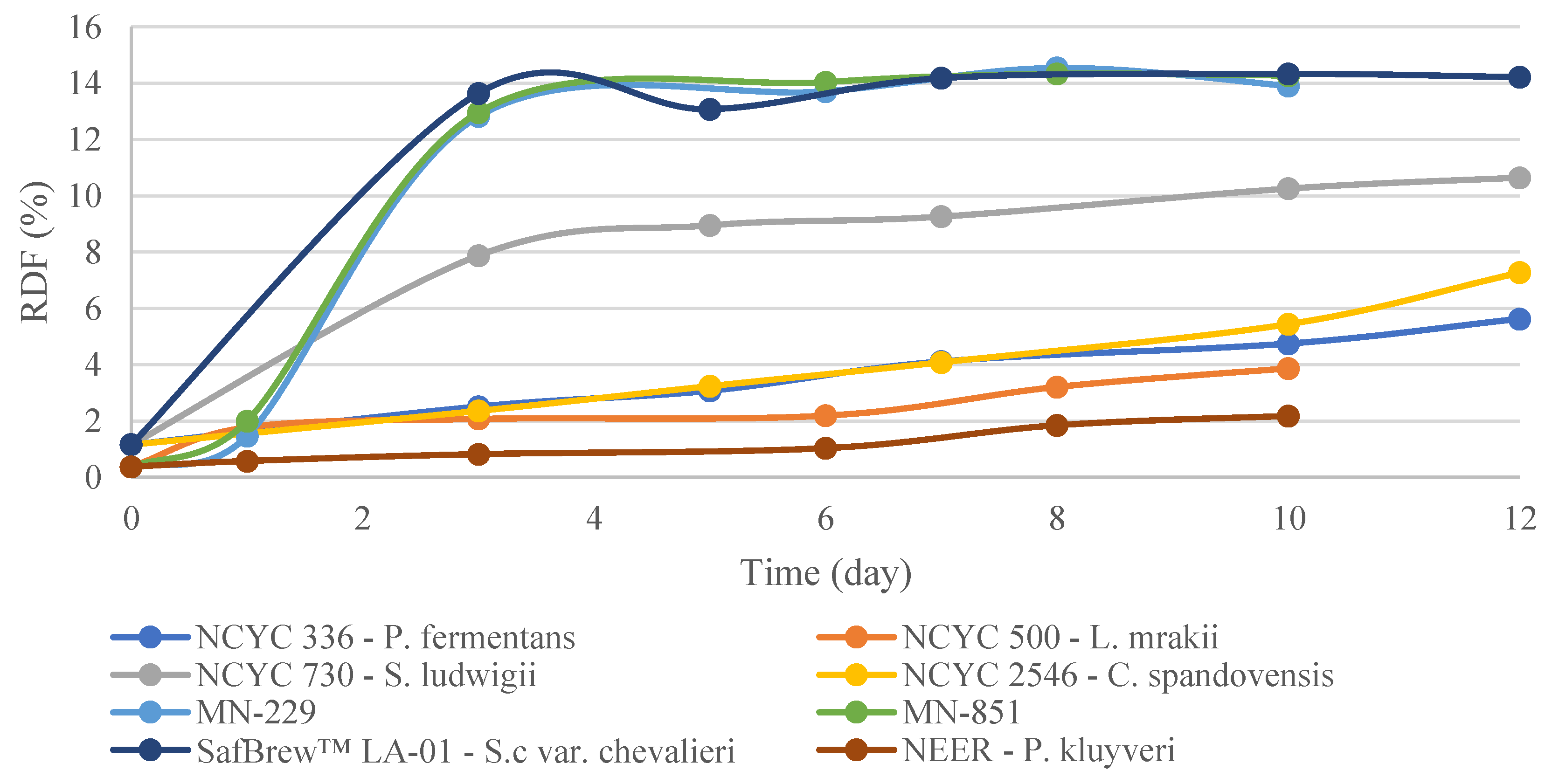
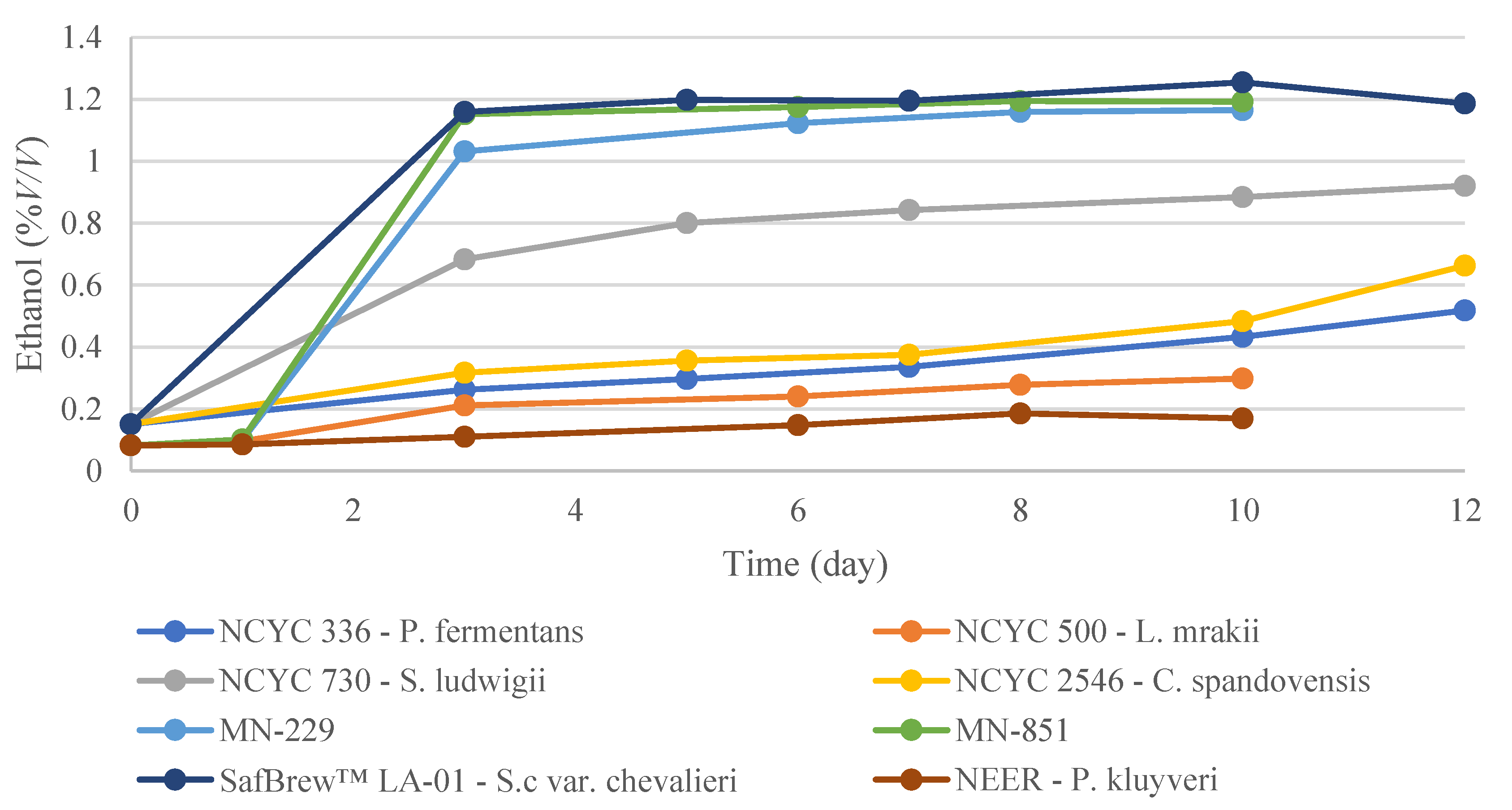
3.1. EBC Tall Tubes
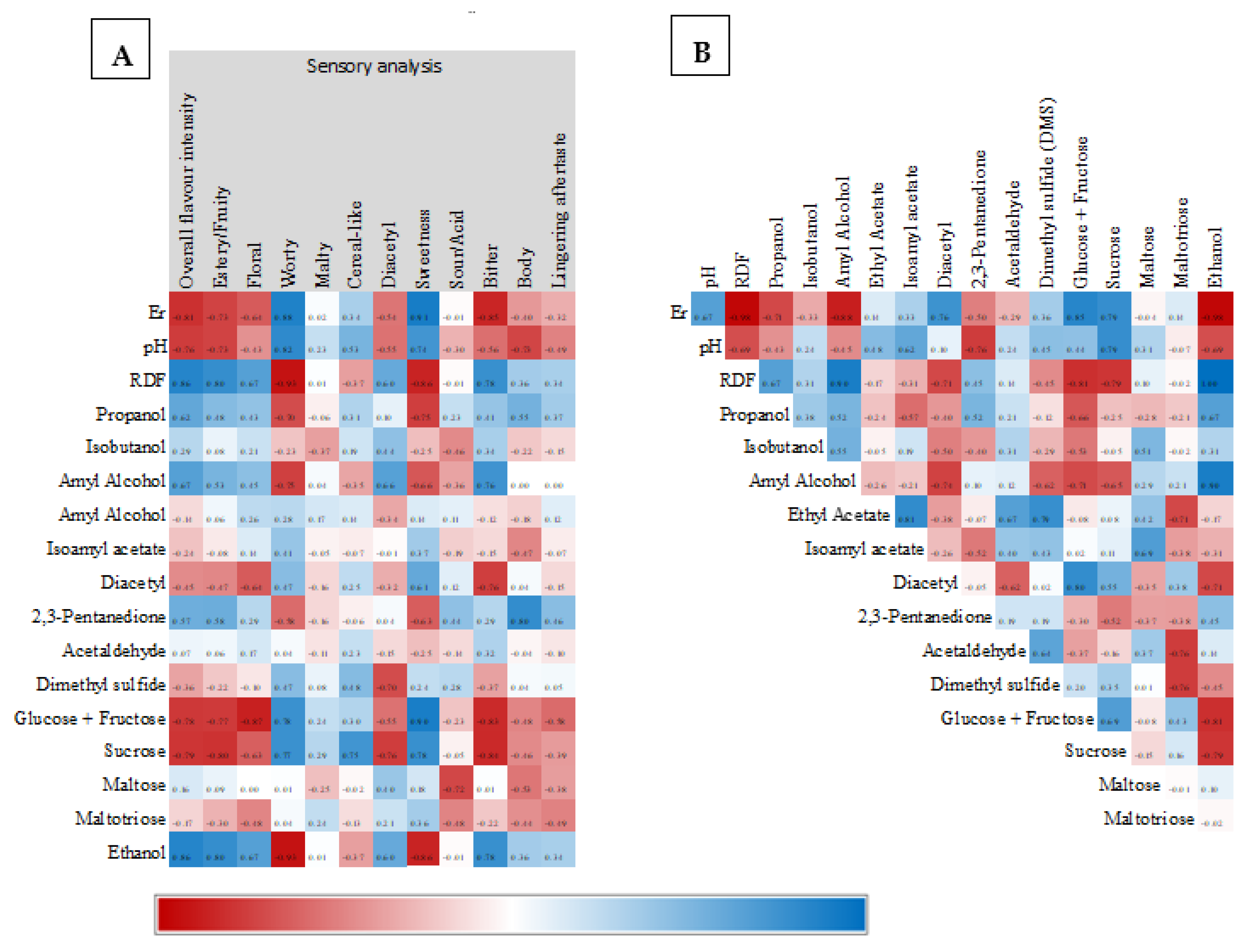
3.2. Volatile Compounds
3.3. Organoleptic Test
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blanco, C.A.; Andrés-Iglesias, C.; Montero, O. Low-alcohol Beers: Flavor Compounds, Defects, and Improvement Strategies. Crit. Rev. Food Sci. Nutr. 2013, 56, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Sohrabvandi, S.; Mousavi, S.; Razavi, S.; Mortazavian, A.; Rezaei, K. Alcohol-free Beer: Methods of Production, Sensorial Defects, and Healthful Effects. Food Rev. Int. 2010, 26, 335–352. [Google Scholar] [CrossRef]
- Bellut, K.; Arendt, E.K. Chance and Challenge: Non-Saccharomyces Yeasts in Nonalcoholic and Low Alcohol Beer Brewing—A Review. J. Am. Soc. Brew. Chem. 2019, 77, 77–91. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Conventional and Non-Conventional Yeasts in Beer Production. Fermentation 2018, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, K.; Petelkov, I.; Shopska, V.; Denkova, R.; Gochev, V.; Kostov, G. Investigation of mashing regimes for low-alcohol beer production. J. Inst. Brew. 2016, 122, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Brányik, T.; Silva, D.P.; Baszczyňski, M.; Lehnert, R.; e Silva, J.B.A. A review of methods of low alcohol and alcohol-free beer production. J. Food Eng. 2012, 108, 493–506. [Google Scholar] [CrossRef]
- Bellut, K.; Michel, M.; Zarnkow, M.; Hutzler, M.; Jacob, F.; De Schutter, D.P.; Daenen, L.; Lynch, K.M.; Zannini, E.; Arendt, E.K. Application of Non-Saccharomyces Yeasts Isolated from Kombucha in the Production of Alcohol-Free Beer. Fermentation 2018, 4, 66. [Google Scholar] [CrossRef] [Green Version]
- Karaoglan, S.Y.; Jung, R.; Gauthier, M.; Dost, P. Maltose-Negative Yeast in Non-Alcoholic and Low-Alcoholic Beer Production. Fermentation 2022, 8, 273. [Google Scholar] [CrossRef]
- Krogerus, K.; Eerikäinen, R.; Aisala, H.; Gibson, B. Repurposing brewery contaminant yeast as production strains for low-alcohol beer fermentation. Yeast 2021, 39, 156–169. [Google Scholar] [CrossRef]
- De Francesco, G.; Turchetti, B.; Sileoni, V.; Marconi, O.; Perretti, G. Screening of new strains of Saccharomycodes ludwigii and Zygosaccharomyces rouxii to produce low-alcohol beer. J. Inst. Brew. 2015, 121, 113–121. [Google Scholar] [CrossRef]
- Matraxia, M.; Alfonzo, A.; Prestianni, R.; Francesca, N.; Gaglio, R.; Todaro, A.; Alfeo, V.; Perretti, G.; Columba, P.; Settanni, L.; et al. Non-conventional yeasts from fermented honey by-products: Focus on Hanseniaspora uvarum strains for craft beer production. Food Microbiol. 2021, 99, 103806. [Google Scholar] [CrossRef] [PubMed]
- Bellut, K.; Krogerus, K.; Arendt, E.K. Lachancea fermentati Strains Isolated From Kombucha: Fundamental Insights, and Practical Application in Low Alcohol Beer Brewing. Front. Microbiol. 2020, 11, 764. [Google Scholar] [CrossRef] [Green Version]
- Van Rijswijck, I.M.H.; Wolkers-Rooijackers, J.C.M.; Abee, T.; Smid, E.J. Performance of non-conventional yeasts in co-culture with brewers’ yeast for steering ethanol and aroma production. Microb. Biotechnol. 2017, 10, 1591–1602. [Google Scholar] [CrossRef]
- Grijalva-Vallejos, N.; Krogerus, K.; Nikulin, J.; Magalhães, F.; Aranda, A.; Matallana, E.; Gibson, B. Potential application of yeasts from Ecuadorian chichas in controlled beer and chicha production. Food Microbiol. 2020, 98, 103644. [Google Scholar] [CrossRef] [PubMed]
- Grijalva-Vallejos, N.; Aranda, A.; Matallana, E. Evaluation of yeasts from Ecuadorian chicha by their performance as starters for alcoholic fermentations in the food industry. Int. J. Food Microbiol. 2019, 317, 108462. [Google Scholar] [CrossRef]
- Johansson, L.; Nikulin, J.; Juvonen, R.; Krogerus, K.; Magalhães, F.; Mikkelson, A.; Nuppunen-Puputti, M.; Sohlberg, E.; de Francesco, G.; Perretti, G.; et al. Sourdough cultures as reservoirs of maltose-negative yeasts for low-alcohol beer brewing. Food Microbiol. 2020, 94, 103629. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-Q.; Quek, A.Y.H. Evaluation of Beer Fermentation with a Novel Yeast Williopsis saturnus. Food Technol. Biotechnol. 2016, 54, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-T.; Hsieh, C.-W.; Lo, Y.-C.; Liou, B.-K.; Lin, H.-W.; Hou, C.-Y.; Cheng, K.-C. Isolation and identification of aroma-producing non-Saccharomyces yeast strains and the enological characteristic comparison in wine making. LWT 2021, 154, 112653. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Zhang, W. Method for Manufacturing Alcohol- Free Beer through Candida Shehatae. Chinese Patent 102220198 B, 6 May 2011. [Google Scholar]
- Rautio, J.; Londesborough, J. Maltose Transport by Brewer’s Yeasts in Brewer’s Wort. J. Inst. Brew. 2003, 109, 251–261. [Google Scholar] [CrossRef]
- Badotti, F.; Dário, M.G.; Alves, S.L., Jr.; Cordioli, M.L.A.; Miletti, L.C.; De Araujo, P.S.; Stambuk, B.U. Switching the mode of sucrose utilization by Saccharomyces cerevisiae. Microb. Cell Factories 2008, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Naumova, E.S.; Sadykova, A.Z.; Martynenko, N.N.; Naumov, G.I. Molecular polymorphism of β-fructosidase SUC genes in the Saccharomyces yeasts. Mol. Biol. 2014, 48, 573–582. [Google Scholar] [CrossRef]
- Sohrabvandi, S.; Malganji, S.H.; Razavi, S.H.; Mousavi, S.M. The Effect of Saccharomyces Strains and Fermentation Condi-tion on the PH, Foam Property and CO2 Concentration of Non-Alcoholic Beer (Ma-al-Shaeer). Appl. Food Biotechnol. 2014, 1, 53–57. [Google Scholar]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-active volatile compounds in beer: Production, regulation and control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [Green Version]
- Postigo, V.; Sánchez, A.; Cabellos, J.M.; Arroyo, T. New Approaches for the Fermentation of Beer: Non-Saccharomyces Yeasts from Wine. Fermentation 2022, 8, 280. [Google Scholar] [CrossRef]
- Willaert, R.; Nedovic, V.A. Primary beer fermentation by immobilised yeast—A review on flavour formation and control strategies. J. Chem. Technol. Biotechnol. 2006, 81, 1353–1367. [Google Scholar] [CrossRef]
- Engan, S. Organoleptic threshold values of some alcohols and esters in beer. J. Inst. Brew. 1972, 78, 33–36. [Google Scholar] [CrossRef]
- Renger, B.R.S.; van Hateren, S.H.; Luyben, K.C.A.M. The Formation of Esters and Higher Alcohols during Brewery Fermen-tation; the Effect of Carbon Dioxide Pressure. J. Insl. Brew. 1992, 98, 509–513. [Google Scholar] [CrossRef]
- Engan, S. Beer Composition: Volatile Substances. In Brewing Science; Academic Press: London, UK, 1981; pp. 93–165. [Google Scholar]
- Brányik, T.; Vicente, A.A.; Dostálek, P.; Teixeira, J.A.; Brew, J.I. A Review of Flavour Formation in Continuous Beer Fer-mentations. J. Inst. Brew. 2008, 114, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Šmogrovičová, D.; Dömény, Z. Beer volatile by-product formation at different fermentation temperature using immobilised yeasts. Process. Biochem. 1999, 34, 785–794. [Google Scholar] [CrossRef]
- Kobayashi, M.; Shimizu, H.; Shioya, S. Beer Volatile Compounds and Their Application to Low-Malt Beer Fermentation. J. Biosci. Bioeng. 2008, 106, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Lehnert, R.; Kuřec, M.; Brányik, T.; Teixeira, J. Effect of Oxygen Supply on Flavor Formation during Continuous Alcohol-Free Beer Production: A Model Study. J. Am. Soc. Brew. Chem. 2008, 66, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.I.; Garcia, L.; Díaz, M. Fusel Alcohols Production in Beer Fermentation Processes. Process. Biochem. 1994, 29, 303–309. [Google Scholar] [CrossRef]
- Ferreira, I.M.; Guido, L.F. Impact of Wort Amino Acids on Beer Flavour: A Review. Fermentation 2018, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Meilgaard, M.C. Flavour Chemistry of Beer. Part II: Flavour and Threshold of 239 Aroma Volatiles. Master Brew. Assoc. Am. Tech. Q. 1975, 28, 132–141. [Google Scholar]
- Rodrigues, F.; Caldeira, M.; Câmara, J. Development of a dynamic headspace solid-phase microextraction procedure coupled to GC–qMSD for evaluation the chemical profile in alcoholic beverages. Anal. Chim. Acta 2008, 609, 82–104. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavor-active esters: Adding fruitiness to beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Hu, Z.; Gamez, G.; Law, W.S.; Chen, H.; Yang, S.; Chingin, K.; Balabin, R.M.; Wang, R.; Zhang, T.; et al. Simultaneous sampling of volatile and non-volatile analytes in beer for fast fingerprinting by extractive electrospray ionization mass spectrometry. Anal. Bioanal. Chem. 2010, 398, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Pavsier, A.; Bulatti, S. Lager Beer. In Beer in Health and Disease Prevention; Preedy, V., Ed.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 31–43. [Google Scholar]
- Guido, L.; Rajendram, R.; Barros, A.A. Pitching Yeast and Beer Flavour. In Beer in Health and Disease Prevention; Preedy, V.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 23–32. [Google Scholar]
- Bamforth, C.; Kanauchi, M. Enzymology of Vicinal Diketone Reduction in Brewer’s Yeast. J. Inst. Brew. 2004, 110, 83–93. [Google Scholar] [CrossRef]
- Narziss, L. Abriss Der Bierbrauwerei; Wiley-VCH Verlag: Weinheim, Germany, 2005. [Google Scholar]
- Kucharczyk, K.; Żyła, K.; Tuszyński, T. Simultaneous Optimization of Acetaldehyde and DMS Concentrations for Better Sensory Quality of Beer Fermented on an Industrial Scale. Foods 2020, 9, 1043. [Google Scholar] [CrossRef]
- MacGregor, A.W. Malting and brewing science: Challenges and opportunities. J. Inst. Brew. 1996, 102, 97–102. [Google Scholar] [CrossRef]
- Petkova, N.; Jonkova, G. Effect of Some Technological Factors on the Content of Acetaldehyde in Beer. St. Cerc. St. CICBIA 2010, 11, 359–364. [Google Scholar]
- Kucharczyk, K.; Tuszyński, T. The effect of pitching rate on fermentation, maturation and flavour compounds of beer produced on an industrial scale. J. Inst. Brew. 2015, 121, 349–355. [Google Scholar] [CrossRef]
- Co, K.B.; Chemistry, N.; Sciences, A.L. 3 Testing for Taste and Flavour of Beer. Available online: https://link.springer.com/chapter/10.1007/978-3-662-04857-3_3 (accessed on 23 January 2002).
- Jan, B.; Pieter, A.D.; Trousdale, E.K.; Singleton, V.L. The Reactions between Polyphenols and Aldehydes and the Influence of Acetaldehyde on Haze Formation in Beer. J. Inst. Brew. 1982, 88, 234–243. [Google Scholar]
- Anderson, R.J.; Clapperton, J.F.; Crabb, D.; Hudson, J.R. Dimethyl sulphide as a feature of lager flavour. J. Inst. Brew. 1975, 81, 208–213. [Google Scholar] [CrossRef]
- James, N.; Stahl, U. Amino Acid Permeases and Their Influence on Flavour Compounds in Beer Amino Acids Are the Chief Source of Assimilable Nitrogen in Wort. Beer Flavour Is Highly Influenced by Wort. Brew. Sci. 2014, 67, 120–127. [Google Scholar]
- Donalies, U.E.B.; Stahl, U. Increasing sulphite formation in Saccharomyces cerevisiae by overexpression ofMET14 and SSU1. Yeast 2002, 19, 475–484. [Google Scholar] [CrossRef]
- Boekhout, T.; Robert, V. Yeasts in Food Beneficial and Detrimental Aspects; Woodhead Publishing: Cambridge, UK, 2003; ISBN 0849319269. [Google Scholar]
- Hough, J.S.; Briggs, D.E.; Stevens, R.; Young, T.W. Metabolism of Wort by Yeast. In Malting and Brewing Science: Hopped Wort and Beer; Hall, C., Ed.; Chapman and Hall: London, UK, 1982; pp. 566–612. [Google Scholar]
- Briggs, D.E.; Boulton, C.A.; Brookes, P.A. Malts, Adjuncts and Supplementary Enzymes. In Brewing Science and Practice; CRC Press: Boca Raton, FL, USA, 2004; p. 2004. [Google Scholar]
- Piddocke, M.P. The Effect of High Gravity on the Brewer’s Yeast Metabolism—Physiological Studies and “Omics”—Analyses; Technical University of Denmark: Lyngby, Denmark, 2009; General. [Google Scholar]
- Perpã¨te, P. Influence of beer ethanol content on the wort flavour perception. Food Chem. 2000, 71, 379–385. [Google Scholar] [CrossRef]
- Tsachaki, M.; Gady, A.-L.; Kalopesas, M.; Linforth, R.S.T.; Athès, V.; Marin, M.; Taylor, A.J. Effect of Ethanol, Temperature, and Gas Flow Rate on Volatile Release from Aqueous Solutions under Dynamic Headspace Dilution Conditions. J. Agric. Food Chem. 2008, 56, 5308–5315. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Designation | Species | Origin | Yeast Bank |
|---|---|---|---|
| NCYC 328 | Hanseniaspora valbyensis | Draught beer, UK. | NCYC, National Collection of Yeast Cultures |
| NCYC 336 | Pichia fermentans | Draught beer, UK. | NCYC, National Collection of Yeast Cultures |
| NCYC 500 | Lindnera mrakii | Soil, Dobodura area, Papua New Guinea | NCYC, National Collection of Yeast Cultures |
| NCYC 730 | Saccharomycodes ludwigii | Isolated from grape must, Germany | NCYC, National Collection of Yeast Cultures |
| NCYC 1508 | Zygosaccharomyces rouxii | Isolated from 70% malt extract | NCYC, National Collection of Yeast Cultures |
| NCYC 2546 | Candida spandovensis | Beer, Germany | NCYC, National Collection of Yeast Cultures |
| MN-229 | Saccharomyces cerevisiae | Engineered organism | Renaissance BioScience |
| MN-851 | Saccharomyces cerevisiae | Engineered organism | Renaissance BioScience |
| SafBrew™ LA-01 | Saccharomyces cerevisiae var. Chevalieri | Selected from Saf Colection | Fermentis |
| NEER | Pichia kluyveri | Isolated in New Zealand | Chr. Hansen SmartBevTM |
| Strain Designation | Ethanol (g L ⁻¹) | Glycerol (g L ⁻¹) | Maltose (g L ⁻¹) | Fructose (g L ⁻¹) | Glucose (g L ⁻¹) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Wort | n.d. | n.d. | 65.52 | ± | 0.97 | 1.85 | ± | 0.11 | 11.68 | ± | 0.20 | ||||
| NCYC 328—H. valbyensis | 26.29 | ± | 4.56 | 1.24 | ± | 0.33 | n.d. | 0.50 | ± | 0.51 | 0.36 | ± | 0.41 | ||
| NCYC 336—P. fermentans | 0.40 | ± | 0.56 | n.d. | 66.90 | ± | 1.51 | n.d. | n.d. | ||||||
| NCYC 500—L. mrakii | n.d. | n.d. | 62.83 | ± | 5.73 | n.d. | n.d. | ||||||||
| NCYC 730—S. ludwigii | n.d. | n.d. | 63.87 | ± | 5.27 | 0.29 | ± | 0.04 | n.d. | ||||||
| NCYC 1508 - Z. rouxii | 7.12 | ± | 6.44 | 2.21 | ± | 0.88 | 12.65 | ± | 8.05 | 0.16 | ± | 0.23 | 2.00 | ± | 0.80 |
| NCYC 2546—C. spandovensis | 0.46 | ± | 0.64 | 0.14 | ± | 0.20 | 59.76 | ± | 6.21 | n.d. | n.d. | ||||
| MN 229 | n.d. | n.d. | 64.47 | ± | 3.13 | n.d. | n.d. | ||||||||
| MN 851 | n.d. | n.d. | 68.61 | ± | 1.76 | 0.24 | ± | 0.03 | n.d. | ||||||
| SafBrew™ LA-01—S.c var. chevalieri | n.d. | n.d. | 62.07 | ± | 0.62 | n.d. | n.d. | ||||||||
| NEER—P. kluyveri | n.d. | n.d. | 65.46 | ± | 3.68 | n.d. | n.d. | ||||||||
| Strain Designation | Ethanol | Maltotriose | Maltose | Sucrose | Glucose + Fructose | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (%v/v) | (g L ⁻¹) | (g L ⁻¹) | (g L ⁻¹) | (g L ⁻¹) | |||||||||||
| Wort | n.d. | 16.60 | ± | 0.10 | 56.50 | ± | 1.30 | 2.20 | ± | 0.40 | 15.95 | ± | 0.05 | ||
| NCYC 336—P. fermentans | 0.52 | ± | 0.03 | 16.35 | ± | 0.45 | 53.60 | ± | 0.40 | 2.55 | ± | 0.05 | 8.10 | ± | 0.60 |
| NCYC 500—L. mrakii | 0.30 | ± | 0.12 | 15.30 | ± | 0.30 | 54.15 | ± | 0.75 | 1.75 | ± | 0.05 | 8.25 | ± | 0.11 |
| NCYC 730—S. ludwigii | 0.92 | ± | 0.05 | 16.05 | ± | 0.05 | 54.25 | ± | 0.25 | n.d. | 4.00 | ± | 0.80 | ||
| NCYC 2546—C. spandovensis | 0.66 | ± | 0.27 | 15.95 | ± | 0.15 | 53.85 | ± | 0.25 | n.d. | 7.70 | ± | 4.50 | ||
| MN-229 | 1.17 | ± | 0.04 | 14.60 | ± | 0.20 | 52.55 | ± | 0.35 | n.d. | n.d. | ||||
| MN-851 | 1.19 | ± | 0.01 | 15.20 | ± | 0.30 | 55.25 | ± | 1.25 | n.d. | n.d. | ||||
| SafBrew™ LA-01 - S.c var. chevalieri | 1.19 | ± | 0.06 | 16.00 | ± | 0.10 | 53.85 | ± | 1.05 | n.d. | 1.00 | ± | 0.00 | ||
| NEER—P. kluyveri | 0.17 | ± | 0.03 | 15.05 | ± | 0.05 | 54.15 | ± | 0.25 | 1.90 | ± | 0.00 | 7.35 | ± | 0.25 |
| Compound | Odor Description | Detection Threshold (mg L ⁻¹) | NCYC 336—P. fermentans | NCYC 500—L. mrakii | NCYC 730—S. ludwigii | NCYC 2546—C. spandovensis | NEER—P. kluyveri | SafBrew™ LA-01 | MN-229 | MN-851 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (mg L ⁻¹) | (mg L ⁻¹) | (mg L ⁻¹) | (mg L ⁻¹) | (mg L ⁻¹) | (mg L ⁻¹) | (mg L ⁻¹) | (mg L ⁻¹) | ||||||||||||||||||||
| Higher alcohols | |||||||||||||||||||||||||||
| Propanol | Alcohol a | 600 e, 800 f,g | 4.40 | ± | 3.45 | 2.34 | ± | 0.04 | 3.06 | ± | 0.12 | 3.96 | ± | 1.72 | 3.15 | ± | 0.55 | 4.21 | ± | 0.63 | 5.46 | ± | 0.61 | 9.90 | ± | 0.19 | |
| Isobutanol | Alcohol a | 100 e, 200 f,g | 5.87 | ± | 0.41 | 3.16 | ± | 0.06 | 9.55 | ± | 0.03 | 3.81 | ± | 1.34 | 7.22 | ± | 1.94 | 4.40 | ± | 0.07 | 4.88 | ± | 0.94 | 11.32 | ± | 0.06 | |
| Amyl Alcohol | Alcohol, banana, medicinal, solvent, fruity a | 50–70 h | 9.46 | ± | 0.94 | 5.69 | ± | 0.06 | 20.87 | ± | 0.31 | 8.07 | ± | 3.24 | 5.06 | ± | 1.41 | 20.45 | ± | 0.60 | 15.73 | ± | 3.17 | 36.06 | ± | 0.17 | |
| Esters | |||||||||||||||||||||||||||
| Ethyl Acetate | Solvent, fruity, sweetish b | 25–30 h | 0.67 | ± | 0.13 | 52.18 | ± | 25.54 | 2.27 | ± | 0.19 | 1.59 | ± | 1.01 | 32.59 | ± | 8.27 | 2.79 | ± | 0.03 | 3.65 | ± | 0.87 | 5.56 | ± | 0.26 | |
| Isoamyl acetate | Banana, apple, solvent, estery, pear b | 1.2 f,h,i,2 h | 0.00 | ± | 0.00 | 0.94 | ± | 0.24 | 0.83 | ± | 0.30 | 0.26 | ± | 0.24 | 9.06 | ± | 1.94 | 0.38 | ± | 0.02 | 0.35 | ± | 0.04 | 0.69 | ± | 0.01 | |
| Vicinal diketones | |||||||||||||||||||||||||||
| Diacetyl | Butter a | 0.15 c | 0.14 | ± | 0.02 | 0.11 | ± | 0.00 | 0.07 | ± | 0.01 | 0.31 | ± | 0.03 | 0.11 | ± | 0.03 | 0.08 | ± | 0.01 | 0.06 | ± | 0.02 | 0.04 | ± | 0.00 | |
| 2,3-Pentanedione | Honey, toffee-like c | 1–1.5 j | 0.02 | ± | 0.00 | 0.02 | ± | 0.00 | 0.02 | ± | 0.00 | 0.08 | ± | 0.04 | 0.02 | ± | 0.00 | 0.03 | ± | 0.01 | 0.04 | ± | 0.01 | 0.03 | ± | 0.00 | |
| Aldehydes | |||||||||||||||||||||||||||
| Acetaldehyde | Grassy, green leaves, fruity a | 10–25 k | 10.80 | ± | 2.91 | 20.10 | ± | 3.54 | 13.29 | ± | 3.18 | 12.34 | ± | 10.19 | 15.45 | ± | 3.35 | 1.47 | ± | 0.05 | 25.83 | ± | 20.78 | 30.18 | ± | 20.53 | |
| Sulfur compounds | |||||||||||||||||||||||||||
| Dimethyl sulfide (DMS) | Cabbagy, Cooked-vegetable d | 0.025—0.030 d, 0.05 i | 9.15 | ± | 0.35 | 20.30 | ± | 0.00 | 6.85 | ± | 1.35 | 9.45 | ± | 1.15 | 16.55 | ± | 3.45 | 8.30 | ± | 0.50 | 16.50 | ± | 1.50 | 11.95 | ± | 3.85 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simões, J.; Coelho, E.; Magalhães, P.; Brandão, T.; Rodrigues, P.; Teixeira, J.A.; Domingues, L. Exploiting Non-Conventional Yeasts for Low-Alcohol Beer Production. Microorganisms 2023, 11, 316. https://doi.org/10.3390/microorganisms11020316
Simões J, Coelho E, Magalhães P, Brandão T, Rodrigues P, Teixeira JA, Domingues L. Exploiting Non-Conventional Yeasts for Low-Alcohol Beer Production. Microorganisms. 2023; 11(2):316. https://doi.org/10.3390/microorganisms11020316
Chicago/Turabian StyleSimões, João, Eduardo Coelho, Paulo Magalhães, Tiago Brandão, Pedro Rodrigues, José António Teixeira, and Lucília Domingues. 2023. "Exploiting Non-Conventional Yeasts for Low-Alcohol Beer Production" Microorganisms 11, no. 2: 316. https://doi.org/10.3390/microorganisms11020316