
Concept about the Virulence Factor of Legionella

Abstract
:1. Introduction
2. Legionella Pathogenicity Evolution during Long-Term Co-Evolution with Protozoa
3. A Supplement to the Concept of Virulence Factors in Legionella

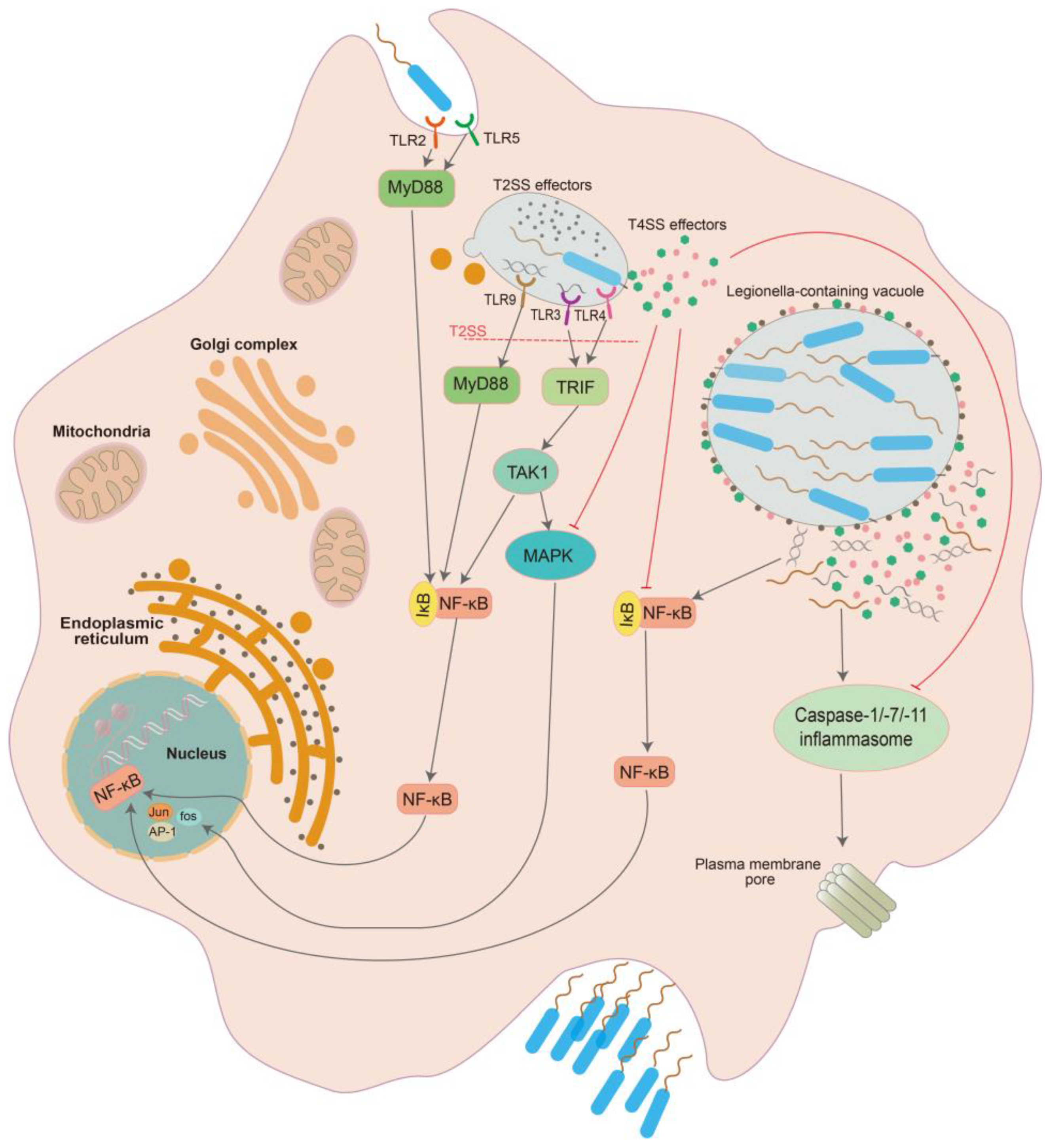{kind=link}
| Classification | Molecule | Coding Gene | Function in Infection of Protozoa | Function in Infection of Humans | Regulate Host Immune Response | Virulence Factor Specific to Protozoa | Ref. |
|---|---|---|---|---|---|---|---|
| Bacterial surface structure | |||||||
| 1 | EnhC | enhC | Absence has no influence in Legionella growth within the amoeba | Adhesive molecule for host cells; trafficking of the Legionella-containing vacuole; decreasing NF-κB activation | Yes | Yes | [93,96,100] |
| 2 | Lcl | lcl | Inducing Legionella autoaggregation promotes the infection of protozoa | A polymorphic adhesin of L. pneumophila that binds to immunogenic GAG and human lung epithelial cells | No, it is antigenic in human infection. | No | [101,102,103] |
| 3 | Hsp60 | htpB | N/A | Promoting attachment and invasion. | No | No | [104,105] |
| 4 | MOMP | mompS | N/A | Mediating phagocytosis via the macrophage integrin receptors CR1 and CR3 | No | No | [106] |
| 5 | type IV pili | pilB-E | Adherence of host cells | Adherence of host cells | No | No | [95,107] |
| 6 | Rtx | rtx | Adherence; intracellular survival and trafficking | Adherence of host cells; helping entry to host cells; affecting cytotoxicity and pore formation, is required for optimal replication of Legionella | No | No | [87,90] |
| 7 | LadC | ladC | Adherence; assisting bacterial replication | Adherence of host cells; assisting bacterial replication | No | No | [108] |
| 8 | flagellin | flaA | Invasion; multiplication | Facilitating the encounter of the host cell; enhancing the invasion capacity; and inducing caspase-1-mediated macrophage death | No | No | [109,110,111] |
| 9 | Mip | mip | No effect on adherence and initial interactions; involved in bacterial resistance to intracellular killing and/or intracellular multiplication. | Bacterial penetration of the lung epithelial barrier; infection of the host cell; contributing to the intracellular survival of Legionellae; necessary for full virulence | No | Yes, having eukaryotic homologous structure for breaking lung epithelial barrier. | [112,113,114] |
| T4SS effectors | |||||||
| 1 | LpdA | lpdA | A phospholipase-D; intracellular multiplication | Modulating the lipid composition of the LCV, affecting the distribution of diacylglycerol and phosphatidic acid; disruptiing the Golgi apparatus; optimal survival of Legionella | No | No | [115,116] |
| 2 | LecE | lecE | A phospholipase-D; intracellular multiplication | Affecting distribution of diacylglycerol and phosphatidic acid | No | No | [115] |
| 3 | LegC7, LegC2 and LegC3 | ylfA, ylfB and legC3 | N/A | Assembling a complex on the LCV to initiate membrane fusion with ER-derived vesicles | No | No | [83] |
| 4 | MavC | mavC | N/A | Inhibiting host immunity by mediating the ubiquitination of E2 enzyme UBE2N in the initial phase of bacterial infection | Yes, directly | Yes | [117] |
| 5 | MavE | mavE | Essential for intracellular growth of Legionella; fusing LCV with endoplasmic reticulum (ER)-derived vesicles. | Essential for intracellular growth in human monocyte-derived macrophages and intrapulmonary proliferation in mice; fusing LCV with ER-derived vesicles; phagosome biogenesis and lysosomal evasion | No | No | [83] |
| 6 | Lgt family | lgt1-3 | A cytotoxic glucosyltransferase. | Glycosylates the mammalian elongation factor eEF1A and inhibits its activity; affecting the protein synthesis of host cells. | Yes, indirectly | No | [118] |
| 7 | LegK1 | legK1 | N/A | An activator of NF-κB | Yes, directly | Yes | [119] |
| 8 | LegK2-4 | legK2-4 | Influence the intracellular replication and growth rate of bacteria; LegK2 plays role in the cytotoxicity of Legionella toward amoeba | Kinases inhibit actin nucleation on phagosomes to interrupt late endosome/lysosome trafficking to the LCV | No | No | [120] |
| 9 | LegG1 | legG1 | A Ran GTPase activator; inhibition migration and chemotaxis of protozoa | A Ran GTPase activator localizing to the LCV membrane; enhancing LCV motility by stabilization or increased polymerization of microtubules and promoting intracellular bacterial replication in phagocytes; modulating phagocyte migration | Yes, indirectly. | No | [121,122] |
| 10 | LegAS4 | legAS4 | Promoting rDNA transcription | Promoting rDNA transcription | Yes, indirectly | No | [123] |
| 11 | LnaB | lnaB | N/A | An activator of NF-κB | Yes, directly | Yes | [119] |
| 12 | SidC | sidC | N/A | subversion of host vesicular transport; participation in the recruitment of Arf1 and ubiquitin to the LCV; utilization of the ubiquitin and phosphoinositide pathways; functions in LCV establishment and is required for optimal growth | No | No | [47,124] |
| 13 | SdcA | sdcA | N/A | Functions in LCV establishment and is required for optimal growth; recruiting Arf1 and ubiquitin to the LCV | No | No | [124] |
| 14 | SidD | sidD | A deAMPylase modifying Rab1; reversing the effect of SidM | A deAMPylase modifying Rab1; reversing the effect of SidM | No | No | [125] |
| 15 | SdeA, SdeB, SdeC and SidE | sdeA, sdeB, sdeC and sidE | SidE family, phosphoribosyl ubiquitination ligases ubiquitinate multiple ER membrane and Golgi resident proteins such as Rab small GTPases to rearrange tubular ER and induce Golgi fragmentation | SidE family members. Phosphoribosyl ubiquitination ligases; ubiquitinate multiple ER membranes and Golgi resident proteins such as Rab small GTPases to rearrange tubular ER and induce Golgi fragmentation. | No | No | [126,127] |
| 16 | SidK | sidK | Inhibiting host vacuolar H+-ATPase to maintain a neutral pH in the phagosome | Inhibiting host vacuolar H+-ATPase to maintain a neutral pH in the phagosome | No | No | [128] |
| 17 | SidJ | sidJ | CaM-activated glutamylase for SidE | CaM-activated glutamylase for SidE | No | No | [129] |
| 18 | SidP | sidP | Phosphoinositide-3-phosphatase; subverting host cell phosphoinositide (PI) metabolism | Phosphoinositide-3-phosphatase; subverting host cell phosphoinositide (PI) metabolism | No | No | [130] |
| 19 | SidM/ DrrA | drrA | Recruiting and activating small GTPase Rab1; no influence in intracellular growth of Legionella | Recruiting small GTPase Rab1 and locking its activation of by guanine nucleotide exchange factor (GEF) and AMPylation activity; no influence in intracellular growth of Legionella | No | No | [131] |
| 20 | AnkB | ankB | Polyubiquitination of proteins; decoration of the LCV with polyubiquitinated proteins; promoting intracellular proliferation | Polyubiquitination of proteins; decoration of the LCV with polyubiquitinated proteins; promoting intracellular proliferation | No | No | [59] |
| 21 | VirD4 | virD4 | A coupling protein for the Lvh T4ASS | A coupling protein for the Lvh T4ASS | No | No | [132] |
| 22 | LpnE | lpnE | Required for efficient infection of amoeba | Helping bacterial entry; influencing host trafficking of Legionella vacuole | No | No | [94,133] |
| 23 | NudA | nudA | A Nudix hydrolase; promoting growth | A Nudix hydrolase; promoting growth | No | No | [134] |
| 24 | PsrA | psrA | Helping bacterial intracellular growth in protozoa | Dispensable for growth in human macrophages. | No | No | [135] |
| 25 | Lem10 | Lem10 | Non-catalytic HD domains-containing protein | Non-catalytic HD domain-containing protein | N/A | N/A | [136] |
| 26 | RavY | ravY | N/A | Essential for promoting intracellular replication but not survival. | No | No | [79] |
| 27 | MavE | mavE | Intracellular growth | Intracellular growth; remodeling the LCV with ER-derived vesicles; preventing LCV from fusing with lysosome | No | No | [83] |
| 28 | MavL | mavL | N/A | As a signaling protein that binds ADP-ribose; interacting with mammalian ubiquitin-conjugating enzyme UBE2Q1 | N/A | N/A | [137] |
| 29 | MavQ | mavQ | A phosphatidylinositol (PI) 3-kinase targeting to ER; driving rapid PI3-phosphate turnover on the ER and remodeling of host ER membrane | A phosphatidylinositol (PI) 3-kinase targeting to ER; driving rapid PI3-phosphate turnover on the ER and remodeling of host ER membrane | No | No | [138] |
| 30 | LtpD | ltpD | N/A | Bound directly to phosphatidylinositol 3-phosphate and inositol (myo)-1 (or 4)-monophosphatase 1; involved in bacterial replication; modulating endocytic vesicle traffic | N/A | N/A | [139] |
| 31 | LtpM | ltpM | A glycosyltransferase; dispensable for replication | A glycosyltransferase; dispensable for replication | N/A | N/A | [140] |
| 32 | SetA | setA | O-glucosyltransferase | O-glucosyltransferase; causes a robust nuclear translocation of transcription factor EB | N/A | N/A | [141,142] |
| 33 | RidL | ridL | Hijacking the host scaffold protein VPS29 critical for endosomal cargo recycling; inhibiting retrograde trafficking; promoting intracellular bacterial replication | Hijacking the host scaffold protein VPS29 critical for endosomal cargo recycling; inhibiting retrograde trafficking; promoting intracellular bacterial replication | No | No | [143] |
| 34 | SidI | sidI | N/A | Interacting with the host translation factor eEF1A and eEF1Bgamma; inhibiting eukaryotic protein translation; inducing host stress response | Yes, indirectly | No | [144] |
| 35 | MesI | mesI | N/A | Inhibiting the activity of SidI; required for optimal intracellular bacterial replication. | Yes, indirectly. | No | [145] |
| 36 | Sde family | sdes | Phosphoribosyl-linked ubiquitination | Phosphoribosyl-linked ubiquitination; resulting in tubule rearrangements of ER | N/A | N/A | [126,146] |
| 37 | PieE | pieE | N/A | PieE localized to the endoplasmic reticulum (ER) and induced the formation of organized smooth ER | N/A | N/A | [147] |
| 38 | SdhA | sdhA | Maintaining LCV integrity | Maintaining LCV integrity | Yes, indirectly. | No | [148] |
| 39 | Lart1 | lart1 | Directly targets NAD+-dependent glutamate dehydrogenase (GDH) enzymes | N/A | N/A | N/A | [149] |
| 40 | RavD | ravD | No effects on intracellular growth; suppressing endolysosomal maturation | No effects on intracellular growth; suppressing endolysosomal maturation; inhibiting the NF-κB pathway via deubiquitinase activity exquisitely specific for linear Ub chains | Yes, directly. | Yes | [150,151] |
| 41 | LegG1 | legG1 | A Ran GTPase activator; inhibiting D. discoideum migration and stimulating cell motility | A Ran GTPase activator; inhibiting phagocyte migration and chemotaxis | Yes, indirectly. | No | [122] |
| 42 | RavK | ravK | N/A | Cleaving actin and disrupting host cytoskeletal structure | N/A | N/A | [152] |
| 43 | PlcA-C | plcA-C | Zn (2+)-dependent PLC family; essential for intracellular replication | Zn (2+)-dependent PLC family; essential for intracellular replication | N/A | N/A | [153] |
| 44 | Ceg3 | ceg3 | A mono-ADP-ribosyltransferase that localizes to the host mitochondria, regulating host energy metabolism; not affect the intracellular replication. | A mono-ADP-ribosyltransferase that localizes to the host mitochondria, regulating host energy metabolism; not affect the intracellular replication | N/A | N/A | [154] |
| 45 | Ceg4 | ceg4 | N/A | A phosphotyrosine phosphatase decreasing activation of eukaryotic MAPK pathways | Yes, directly. | Yes | [155] |
| 46 | VipA | vipA | An actin nucleator that enhancesactin polymerization; altering host cell organelle trafficking; not essential for bacterial entry or replication | An actin nucleator that enhances actin polymerization; altering host cell organelle trafficking; not essential for bacterial entry or replication | No | No | [156] |
| 47 | SdbA | sdbA | N/A | Be important for bacterial replication; continuous activation of NF-κB | Yes, directly. | Yes | [157] |
| 48 | SdcA | sdcA | Bacterial E3 ubiquitin ligases as a paralog of SidC; anchoring on the LCV by binding to PI(4)P; recruiting Rab10 to the LCV | Bacterial E3 ubiquitin ligases as a paralog of SidC; anchoring on the LCV by binding to PI(4)P; recruiting Rab10 to the LCV | No | No | [158] |
| 49 | LubX | lubX | N/A | Continuous activating host NF-κB signaling. | Yes, directly | Yes | [159] |
| 50 | LamA | lamA | N/A | An amylase that can rapidly degrade glycogen to generate cytosolic hyper-glucose, directly triggering an M1-like pro-inflammatory differentiation and directly triggers an M1-like pro-inflammatory differentiation. | Yes, indirectly. | No | [160] |
| 51 | Lem27 | lem27 | Dispensable for bacterial intracellular replication | A deubiquitinases regulating LCV Rab10 ubiquitination in concert with SidC and SdcA; dispensable for bacterial intracellular replication in macrophages | No | No | [161] |
| 52 | PmrA | pmrA | Intracellular proliferation in the ciliate; having a global effect on gene expression with PmrB. | Having a global effect on gene expression with PmrB. | N/A | N/A | [162] |
| 53 | PmrB | pmrB | Intracellular proliferation; having a global effect on gene expression with PmrA. | Intracellular proliferation; having a global effect on gene expression with PmrA | N/A | N/A | [162] |
| 54 | RalF | ralF | A guanine nucleotide exchange factor is activated in the presence of the membrane | A guanine nucleotide exchange factor is activated in the presence of the membrane | No | No | [163] |
| 55 | LidA | lidA | Binding to Rab1 to stabilize the Rab1-guanosine nucleotide complex and activity of the host GTPase Rab1; interfering with the covalent modification of Rab1 by SidD and Lem3 | Binding to Rab1 to stabilize the Rab1-guanosine nucleotide complex and activity of the host GTPase Rab1; and interfering with the covalent modification of Rab1 by SidD and Lem3 | No | No | [164] |
| 56 | LepA and LepB | lepA and lepB | Playing a role in the non-lytic release of Legionella from protozoa | N/A | N/A | N/A | [165] |
| 57 | PieE | pieE | N/A | Interacting with multiple Rab GTPases; inducing stacked ER membranes; forming complexes with multiple host proteins | N/A | N/A | [147] |
| 58 | LppA | lppA | A phytate phosphatase (phytase) for efficient replication; promoting intracellular replication in phytate-loaded amoebae | A phytate phosphatase (phytase) | N/A | N/A | [166] |
| 59 | Ceg4 | ceg4 | A phosphotyrosine phosphatase; dephosphorylating a broad range of phosphotyrosine-containing peptides | A phosphotyrosine phosphatase; dephosphorylating a broad range of phosphotyrosine-containing peptides; attenuating the activation of MAPK-controlled pathways | Yes, directly. | Yes | [155] |
| T2SS effectors | |||||||
| 1 | LapA | lapA | An aminopeptidase; playing a role in intracellular replication; generating amino acids for nutrition | An aminopeptidase | N/A | N/A | [73] |
| 2 | LapB | lapB | Lysine and arginine aminopeptidase | Lysine and arginine aminopeptidase | N/A | N/A | [167] |
| 3 | PlaC | plaC | A protein with acyltransferase, phospholipase A, and lysophospholipase A activities; playing a dispensable role in intracellular replication | A protein with acyltransferase, phospholipase A, and lysophospholipase A activities; playing a dispensable role in intracellular replication | N/A | N/A | [168] |
| 4 | NttA | nttA | Contributing to intracellular multiplication in a part of protozoa, such as Acanthamoeba castellanii | N/A | N/A | N/A | [169] |
| 5 | NttD | nttD | Promoting infection of protozoa | N/A | N/A | N/A | [73] |
| 6 | NttE | nttE | Required for optimal infection of Acanthamoeba castellanii and Hartmannella vermiformis amoeba | N/A | N/A | N/A | [170] |
| 7 | ProA | proA | Required for infection of Hartmannella vermiformis but not infection of Acanthamoeba castellanii | Promoting infection of human lung tissue explants and increasing the alveolar septal thickness; directly degrading immunogenic FlaA monomers; a modulator of flagellin-mediated TLR5 stimulation and the NF-κB pathway | Yes, directly | Yes | [169,171,172] |
| 8 | ChiA | chiA | A chitinase; not required for intracellular growth; | A chitinase; not required for intracellular growth; directly or indirectly required for optimal survival in the lung tissue | N/A | N/A | [173] |
| 9 | SrnA | srnA | Required for intracellular infection of specific protozoa, such as Hartmannella vermiformis | N/A | N/A | N/A | [169] |
| T1SS effectors | |||||||
| 1 | RtxA | rtxA | Adherence and entry into host cells, affecting intracellular survival by regulating trafficking | Adherence and entry into host cells, enhancing replication, cytotoxicity, and pore formation | No | No | [90,92] |
| Others | |||||||
| 1 | PlaB | s | A cell-associated phospholipase A/lysophospholipase A, not essential for intracellular replication | A cell-associated phospholipase A/lysophospholipase A; not essential for intracellular replication; helping bacterial colonization in the lung and enlarging inflammation | N/A | N/A | [174] |
| 2 | RsmY | rsmY | N/A | Binding to the UTR of ddx58 (RIG-I encoding gene) and cRel, downregulating the expression of Rig-I like miRNA and decreasing the IFN-β response | Yes, directly | Yes | [175] |
| 3 | tRNA-Phe | lppt29 | N/A | Collectively reducing expression of RIG-I, IRAK1 and cRel and downregulating IFN-β | Yes, directly | Yes | [175] |
4. Innate Immune Response to Legionella
5. Regulation of Host Immune Response by Legionella
6. Legionella Virulence Is Related to the Early Immunosuppressive and Later Inflammatory Levels in the Host
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fields, B.S.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ disease: 25 years of investigation. Clin. Microbiol. Rev. 2002, 15, 506–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, W.T.; Hsu, B.M.; Chang, T.Y.; Hsu, T.K.; Kao, P.M.; Huang, K.H.; Tsai, S.F.; Huang, Y.L.; Fan, C.W. Surveillance and evaluation of the infection risk of free-living amoebae and Legionella in different aquatic environments. Sci. Total Environ. 2014, 499, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Magnet, A.; Peralta, R.H.; Gomes, T.S.; Izquierdo, F.; Fernandez-Vadillo, C.; Galvan, A.L.; Pozuelo, M.J.; Pelaz, C.; Fenoy, S.; Del Águila, C. Vectorial role of Acanthamoeba in Legionella propagation in water for human use. Sci. Total Environ. 2015, 505, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Phin, N.; Parry-Ford, F.; Harrison, T.; Stagg, H.R.; Zhang, N.; Kumar, K.; Lortholary, O.; Zumla, A.; Abubakar, I. Epidemiology and clinical management of Legionnaires’ disease. Lancet. Infect. Dis. 2014, 14, 1011–1021. [Google Scholar] [CrossRef]
- Iliadi, V.; Staykova, J.; Iliadis, S.; Konstantinidou, I.; Sivykh, P.; Romanidou, G.; Vardikov, D.F.; Cassimos, D.; Konstantinidis, T.G. Legionella pneumophila: The Journey from the Environment to the Blood. J. Clin. Med. 2022, 11, 6126. [Google Scholar] [CrossRef]
- Marston, B.J.; Lipman, H.B.; Breiman, R.F. Surveillance for Legionnaires’ disease. Risk factors for morbidity and mortality. Arch. Intern. Med. 1994, 154, 2417–2422. [Google Scholar] [CrossRef]
- Borges, V.; Nunes, A.; Sampaio, D.A.; Vieira, L.; Machado, J.; Simoes, M.J.; Goncalves, P.; Gomes, J.P. Legionella pneumophila strain associated with the first evidence of person-to-person transmission of Legionnaires’ disease: A unique mosaic genetic backbone. Sci. Rep. 2016, 6, 26261. [Google Scholar] [CrossRef] [Green Version]
- Whiley, H.; Bentham, R. Legionella longbeachae and legionellosis. Emerg. Infect. Dis. 2011, 17, 579–583. [Google Scholar] [CrossRef]
- McDade, J.E.; Shepard, C.C.; Fraser, D.W.; Tsai, T.R.; Redus, M.A.; Dowdle, W.R. Legionnaires’ disease: Isolation of a bacterium and demonstration of its role in other respiratory disease. N. Engl. J. Med. 1977, 297, 1197–1203. [Google Scholar] [CrossRef]
- Khodr, A.; Kay, E.; Gomez-Valero, L.; Ginevra, C.; Doublet, P.; Buchrieser, C.; Jarraud, S. Molecular epidemiology, phylogeny and evolution of Legionella. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 43, 108–122. [Google Scholar] [CrossRef]
- Talapko, J.; Frauenheim, E.; Juzbasic, M.; Tomas, M.; Matic, S.; Jukic, M.; Samardzic, M.; Skrlec, I. Legionella pneumophila-Virulence Factors and the Possibility of Infection in Dental Practice. Microorganisms 2022, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Yu, V.L.; Plouffe, J.F.; Pastoris, M.C.; Stout, J.E.; Schousboe, M.; Widmer, A.; Summersgill, J.; File, T.; Heath, C.M.; Paterson, D.L.; et al. Distribution of Legionella species and serogroups isolated by culture in patients with sporadic community-acquired legionellosis: An international collaborative survey. J. Infect. Dis. 2002, 186, 127–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, S.T.; Slow, S.; Scott-Thomas, A.; Murdoch, D.R. Legionellosis Caused by Non-Legionella pneumophila Species, with a Focus on Legionella longbeachae. Microorganisms 2021, 9, 291. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, N.; Higa, F.; Aoki, Y.; Kikuchi, T.; Seki, M.; Tateda, K.; Maki, N.; Uchino, K.; Ogasawara, K.; Kiyota, H.; et al. Distribution of Legionella species and serogroups in patients with culture-confirmed Legionella pneumonia. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2020, 26, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Li, J.S.; O’Brien, E.D.; Guest, C. A review of national legionellosis surveillance in Australia, 1991 to 2000. Commun. Dis. Intell. Q. Rep. 2002, 26, 461–468. [Google Scholar] [PubMed]
- Zhan, X.Y.; Yang, J.L.; Sun, H.; Zhou, X.; Qian, Y.C.; Huang, K.; Leng, Y.; Huang, B.; He, Y. Presence of Viable, Clinically Relevant Legionella Bacteria in Environmental Water and Soil Sources of China. Microbiol. Spectr. 2022, 10, e0114021. [Google Scholar] [CrossRef]
- Phares, C.R.; Wangroongsarb, P.; Chantra, S.; Paveenkitiporn, W.; Tondella, M.L.; Benson, R.F.; Thacker, W.L.; Fields, B.S.; Moore, M.R.; Fischer, J.; et al. Epidemiology of severe pneumonia caused by Legionella longbeachae, Mycoplasma pneumoniae, and Chlamydia pneumoniae: 1-year, population-based surveillance for severe pneumonia in Thailand. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2007, 45, e147–e155. [Google Scholar] [CrossRef] [Green Version]
- Beauté, J.; Plachouras, D.; Sandin, S.; Giesecke, J.; Sparén, P. Healthcare-Associated Legionnaires’ Disease, Europe, 2008–2017. Emerg. Infect. Dis. 2020, 26, 2309–2318. [Google Scholar] [CrossRef]
- Gao, L.Y.; Harb, O.S.; Abu Kwaik, Y. Utilization of similar mechanisms by Legionella pneumophila to parasitize two evolutionarily distant host cells, mammalian macrophages and protozoa. Infect. Immun. 1997, 65, 4738–4746. [Google Scholar] [CrossRef] [Green Version]
- Byrne, B.; Swanson, M.S. Expression of Legionella pneumophila virulence traits in response to growth conditions. Infect. Immun. 1998, 66, 3029–3034. [Google Scholar] [CrossRef]
- Wieland, H.; Faigle, M.; Lang, F.; Northoff, H.; Neumeister, B. Regulation of the Legionella mip-promotor during infection of human monocytes. FEMS Microbiol. Lett. 2002, 212, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Andreozzi, E.; Di Cesare, A.; Sabatini, L.; Chessa, E.; Sisti, D.; Rocchi, M.; Citterio, B. Role of biofilm in protection of the replicative form of Legionella pneumophila. Curr. Microbiol. 2014, 69, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, M.A. Characterization of avirulent mutant Legionella pneumophila that survive but do not multiply within human monocytes. J. Exp. Med. 1987, 166, 1310–1328. [Google Scholar] [CrossRef] [PubMed]
- Fields, B.S.; Barbaree, J.M.; Sanden, G.N.; Morrill, W.E. Virulence of a Legionella anisa strain associated with Pontiac fever: An evaluation using protozoan, cell culture, and guinea pig models. Infect. Immun. 1990, 58, 3139–3142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fields, B.S.; Barbaree, J.M.; Shotts, E.B., Jr.; Feeley, J.C.; Morrill, W.E.; Sanden, G.N.; Dykstra, M.J. Comparison of guinea pig and protozoan models for determining virulence of Legionella species. Infect. Immun. 1986, 53, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Saito, M.; Tanaka, T.; Amako, K.; Yoshida, S. Comparative analysis of virulence traits between a Legionella feeleii strain implicated in Pontiac fever and a strain that caused Legionnaires’ disease. Microb. Pathog. 2015, 89, 79–86. [Google Scholar] [CrossRef]
- Cianciotto, N.P. Pathogenicity of Legionella pneumophila. Int. J. Med. Microbiol. IJMM 2001, 291, 331–343. [Google Scholar] [CrossRef]
- Newton, H.J.; Ang, D.K.; van Driel, I.R.; Hartland, E.L. Molecular pathogenesis of infections caused by Legionella pneumophila. Clin. Microbiol. Rev. 2010, 23, 274–298. [Google Scholar] [CrossRef] [Green Version]
- Best, A.; Price, C.; Ozanic, M.; Santic, M.; Jones, S.; Kwaik, Y.A. A Legionella pneumophila amylase is essential for intracellular replication in human macrophages and amoebae. Sci. Rep. 2018, 8, 6340. [Google Scholar] [CrossRef]
- Miyake, M.; Watanabe, T.; Koike, H.; Molmeret, M.; Imai, Y.; Kwaik, Y.A. Characterization of Legionella pneumophila pmiA, a gene essential for infectivity of protozoa and macrophages. Infect. Immun. 2005, 73, 6272–6282. [Google Scholar] [CrossRef]
- Wang, H.; Lu, J.; Li, K.; Ren, H.; Shi, Y.; Qin, T.; Duan, X.; Fang, M. The virulence of Legionella pneumophila is positively correlated with its ability to stimulate NF-κB activation. Future Microbiol. 2018, 13, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shin, S. Viewing Legionella pneumophila Pathogenesis through an Immunological Lens. J. Mol. Biol. 2019, 431, 4321–4344. [Google Scholar] [CrossRef] [PubMed]
- Rowbotham, T.J. Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae. J. Clin. Pathol. 1980, 33, 1179–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holden, E.P.; Winkler, H.H.; Wood, D.O.; Leinbach, E.D. Intracellular growth of Legionella pneumophila within Acanthamoeba castellanii Neff. Infect. Immun. 1984, 45, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Anand, C.M.; Skinner, A.R.; Malic, A.; Kurtz, J.B. Interaction of L. pneumophilia and a free living amoeba (Acanthamoeba palestinensis). Epidemiol. Infect. 1983, 91, 167–178. [Google Scholar] [CrossRef]
- Vandenesch, F.; Surgot, M.; Bornstein, N.; Paucod, J.C.; Marmet, D.; Isoard, P.; Fleurette, J. Relationship between free amoeba and Legionella: Studies in vitro and in vivo. Zent. Fur Bakteriol. Int. J. Med. Microbiol. 1990, 272, 265–275. [Google Scholar] [CrossRef]
- Tyndall, R.L.; Domingue, E.L. Cocultivation of Legionella pneumophila and free-living amoebae. Appl. Environ. Microbiol. 1982, 44, 954–959. [Google Scholar] [CrossRef] [Green Version]
- Shadrach, W.S.; Rydzewski, K.; Laube, U.; Holland, G.; Ozel, M.; Kiderlen, A.F.; Flieger, A. Balamuthia mandrillaris, free-living ameba and opportunistic agent of encephalitis, is a potential host for Legionella pneumophila bacteria. Appl. Environ. Microbiol. 2005, 71, 2244–2249. [Google Scholar] [CrossRef] [Green Version]
- Lau, H.Y.; Ashbolt, N.J. The role of biofilms and protozoa in Legionella pathogenesis: Implications for drinking water. J. Appl. Microbiol. 2009, 107, 368–378. [Google Scholar] [CrossRef]
- Wadowsky, R.M.; Wilson, T.M.; Kapp, N.J.; West, A.J.; Kuchta, J.M.; States, S.J.; Dowling, J.N.; Yee, R.B. Multiplication of Legionella spp. in tap water containing Hartmannella vermiformis. Appl. Environ. Microbiol. 1991, 57, 1950–1955. [Google Scholar] [CrossRef]
- García, M.T.; Jones, S.; Pelaz, C.; Millar, R.D.; Kwaik, Y.A. Acanthamoeba polyphaga resuscitates viable non-culturable Legionella pneumophila after disinfection. Environ. Microbiol. 2007, 9, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Amaro, F.; Martín-González, A. Microbial warfare in the wild-the impact of protists on the evolution and virulence of bacterial pathogens. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2021, 24, 559–571. [Google Scholar] [CrossRef]
- Albert-Weissenberger, C.; Cazalet, C.; Buchrieser, C. Legionella pneumophila—A human pathogen that co-evolved with fresh water protozoa. Cell. Mol. Life Sci. CMLS 2007, 64, 432–448. [Google Scholar] [CrossRef] [PubMed]
- Khweek, A.A.; Amer, A.O. Factors Mediating Environmental Biofilm Formation by Legionella pneumophila. Front. Cell. Infect. Microbiol. 2018, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, D.; Shames, S.R. Pathogenicity and Virulence of Legionella: Intracellular replication and host response. Virulence 2021, 12, 1122–1144. [Google Scholar] [CrossRef] [PubMed]
- Haneburger, I.; Hilbi, H. Phosphoinositide lipids and the Legionella pathogen vacuole. Curr. Top. Microbiol. Immunol. 2013, 376, 155–173. [Google Scholar] [CrossRef]
- Wasilko, D.J.; Mao, Y. Exploiting the ubiquitin and phosphoinositide pathways by the Legionella pneumophila effector, SidC. Curr. Genet. 2016, 62, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, C.; Harrison, C.F.; Hilbi, H. The natural alternative: Protozoa as cellular models for Legionella infection. Cell. Microbiol. 2014, 16, 15–26. [Google Scholar] [CrossRef]
- Escoll, P.; Rolando, M.; Gomez-Valero, L.; Buchrieser, C. From amoeba to macrophages: Exploring the molecular mechanisms of Legionella pneumophila infection in both hosts. Curr. Top. Microbiol. Immunol. 2013, 376, 1–34. [Google Scholar] [CrossRef]
- Segal, G.; Shuman, H.A. Legionella pneumophila utilizes the same genes to multiply within Acanthamoeba castellanii and human macrophages. Infect. Immun. 1999, 67, 2117–2124. [Google Scholar] [CrossRef]
- Cazalet, C.; Rusniok, C.; Bruggemann, H.; Zidane, N.; Magnier, A.; Ma, L.; Tichit, M.; Jarraud, S.; Bouchier, C.; Vandenesch, F.; et al. Evidence in the Legionella pneumophila genome for exploitation of host cell functions and high genome plasticity. Nat. Genet. 2004, 36, 1165–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazalet, C.; Gomez-Valero, L.; Rusniok, C.; Lomma, M.; Dervins-Ravault, D.; Newton, H.J.; Sansom, F.M.; Jarraud, S.; Zidane, N.; Ma, L.; et al. Analysis of the Legionella longbeachae genome and transcriptome uncovers unique strategies to cause Legionnaires’ disease. PLoS Genet. 2010, 6, e1000851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Valero, L.; Rusniok, C.; Carson, D.; Mondino, S.; Perez-Cobas, A.E.; Rolando, M.; Pasricha, S.; Reuter, S.; Demirtas, J.; Crumbach, J.; et al. More than 18,000 effectors in the Legionella genus genome provide multiple, independent combinations for replication in human cells. Proc. Natl. Acad. Sci. USA 2019, 116, 2265–2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burstein, D.; Amaro, F.; Zusman, T.; Lifshitz, Z.; Cohen, O.; Gilbert, J.A.; Pupko, T.; Shuman, H.A.; Segal, G. Genomic analysis of 38 Legionella species identifies large and diverse effector repertoires. Nat. Genet. 2016, 48, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Cazalet, C.; Jarraud, S.; Ghavi-Helm, Y.; Kunst, F.; Glaser, P.; Etienne, J.; Buchrieser, C. Multigenome analysis identifies a worldwide distributed epidemic Legionella pneumophila clone that emerged within a highly diverse species. Genome Res. 2008, 18, 431–441. [Google Scholar] [CrossRef] [Green Version]
- de Felipe, K.S.; Pampou, S.; Jovanovic, O.S.; Pericone, C.D.; Ye, S.F.; Kalachikov, S.; Shuman, H.A. Evidence for acquisition of Legionella type IV secretion substrates via interdomain horizontal gene transfer. J. Bacteriol. 2005, 187, 7716–7726. [Google Scholar] [CrossRef] [Green Version]
- Price, C.T.D.; Kwaik, Y.A. Evolution and Adaptation of Legionella pneumophila to Manipulate the Ubiquitination Machinery of Its Amoebae and Mammalian Hosts. Biomolecules 2021, 11, 112. [Google Scholar] [CrossRef]
- Hryniewicz-Jankowska, A.; Czogalla, A.; Bok, E.; Sikorsk, A.F. Ankyrins, multifunctional proteins involved in many cellular pathways. Folia Histochem. Et Cytobiol. 2002, 40, 239–249. [Google Scholar]
- Price, C.T.; Al-Khodor, S.; Al-Quadan, T.; Kwaik, Y.A. Indispensable role for the eukaryotic-like ankyrin domains of the ankyrin B effector of Legionella pneumophila within macrophages and amoebae. Infect. Immun. 2010, 78, 2079–2088. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Valero, L.; Rusniok, C.; Jarraud, S.; Vacherie, B.; Rouy, Z.; Barbe, V.; Medigue, C.; Etienne, J.; Buchrieser, C. Extensive recombination events and horizontal gene transfer shaped the Legionella pneumophila genomes. BMC Genom. 2011, 12, 536. [Google Scholar] [CrossRef] [Green Version]
- Habyarimana, F.; Price, C.T.; Santic, M.; Al-Khodor, S.; Kwaik, Y.A. Molecular characterization of the Dot/Icm-translocated AnkH and AnkJ eukaryotic-like effectors of Legionella pneumophila. Infect. Immun. 2010, 78, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Moliner, C.; Raoult, D.; Fournier, P.E. Evidence of horizontal gene transfer between amoeba and bacteria. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2009, 15 (Suppl. 2), 178–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amor, J.C.; Swails, J.; Zhu, X.; Roy, C.R.; Nagai, H.; Ingmundson, A.; Cheng, X.; Kahn, R.A. The structure of RalF, an ADP-ribosylation factor guanine nucleotide exchange factor from Legionella pneumophila, reveals the presence of a cap over the active site. J. Biol. Chem. 2005, 280, 1392–1400. [Google Scholar] [CrossRef] [Green Version]
- Shabardina, V.; Kischka, T.; Kmita, H.; Suzuki, Y.; Makałowski, W. Environmental adaptation of Acanthamoeba castellanii and Entamoeba histolytica at genome level as seen by comparative genomic analysis. Int. J. Biol. Sci. 2018, 14, 306–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degtyar, E.; Zusman, T.; Ehrlich, M.; Segal, G. A Legionella effector acquired from protozoa is involved in sphingolipids metabolism and is targeted to the host cell mitochondria. Cell. Microbiol. 2009, 11, 1219–1235. [Google Scholar] [CrossRef]
- Linsky, M.; Segal, G. A horizontally acquired Legionella genomic island encoding a LuxR type regulator and effector proteins displays variation in gene content and regulation. Mol. Microbiol. 2021, 116, 766–782. [Google Scholar] [CrossRef]
- Gomez-Valero, L.; Buchrieser, C. Intracellular parasitism, the driving force of evolution of Legionella pneumophila and the genus Legionella. Genes Immun. 2019, 20, 394–402. [Google Scholar] [CrossRef]
- Lurie-Weinberger, M.N.; Gomez-Valero, L.; Merault, N.; Glöckner, G.; Buchrieser, C.; Gophna, U. The origins of eukaryotic-like proteins in Legionella pneumophila. Int. J. Med. Microbiol. IJMM 2010, 300, 470–481. [Google Scholar] [CrossRef]
- Gomez-Valero, L.; Chiner-Oms, A.; Comas, I.; Buchrieser, C. Evolutionary Dissection of the Dot/Icm System Based on Comparative Genomics of 58 Legionella Species. Genome Biol. Evol. 2019, 11, 2619–2632. [Google Scholar] [CrossRef]
- Zhan, X.Y.; Yang, J.L.; Zhou, X.; Qian, Y.C.; Huang, K.; Sun, H.; Wang, H.; Leng, Y.; Huang, B.; He, Y. Virulence effector SidJ evolution in Legionella pneumophila is driven by positive selection and intragenic recombination. PeerJ 2021, 9, e12000. [Google Scholar] [CrossRef]
- Dowling, J.N.; Saha, A.K.; Glew, R.H. Virulence factors of the family Legionellaceae. Microbiol. Rev. 1992, 56, 32–60. [Google Scholar] [CrossRef] [PubMed]
- Isaac, D.T.; Isberg, R. Master manipulators: An update on Legionella pneumophila Icm/Dot translocated substrates and their host targets. Future Microbiol. 2014, 9, 343–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, R.C.; Gunderson, F.F.; Tyson, J.Y.; Richardson, K.H.; Portlock, T.J.; Garnett, J.A.; Cianciotto, N.P. Type II Secretion-Dependent Aminopeptidase LapA and Acyltransferase PlaC Are Redundant for Nutrient Acquisition during Legionella pneumophila Intracellular Infection of Amoebas. mBio 2018, 9, e00528-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, P.J.; Lee, J.V. Differences in aerosol survival between pathogenic and non-pathogenic strains of Legionella pneumophila serogroup 1. J. Appl. Bacteriol. 1988, 65, 135–141. [Google Scholar] [CrossRef]
- Bandyopadhyay, P.; Liu, S.; Gabbai, C.B.; Venitelli, Z.; Steinman, H.M. Environmental mimics and the Lvh type IVA secretion system contribute to virulence-related phenotypes of Legionella pneumophila. Infect. Immun. 2007, 75, 723–735. [Google Scholar] [CrossRef] [Green Version]
- Cianciotto, N.P. Type II secretion and Legionella virulence. Curr. Top. Microbiol. Immunol. 2013, 376, 81–102. [Google Scholar] [CrossRef]
- Fuche, F.; Vianney, A.; Andrea, C.; Doublet, P.; Gilbert, C. Functional type 1 secretion system involved in Legionella pneumophila virulence. J. Bacteriol. 2015, 197, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Mondino, S.; Schmidt, S.; Rolando, M.; Escoll, P.; Gomez-Valero, L.; Buchrieser, C. Legionnaires’ Disease: State of the Art Knowledge of Pathogenesis Mechanisms of Legionella. Annu. Rev. Pathol. 2020, 15, 439–466. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Roy, C.R. The Legionella pneumophila Effector RavY Contributes to a Replication-Permissive Vacuolar Environment during Infection. Infect. Immun. 2021, 89, e0026121. [Google Scholar] [CrossRef]
- Shames, S.R.; Liu, L.; Havey, J.C.; Schofield, W.B.; Goodman, A.L.; Roy, C.R. Multiple Legionella pneumophila effector virulence phenotypes revealed through high-throughput analysis of targeted mutant libraries. Proc. Natl. Acad. Sci. USA 2017, 114, E10446–E10454. [Google Scholar] [CrossRef] [Green Version]
- Creasey, E.A.; Isberg, R.R. The protein SdhA maintains the integrity of the Legionella-containing vacuole. Proc. Natl. Acad. Sci. USA 2012, 109, 3481–3486. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.Y.; Kim, S.; Aurass, P.; Huo, W.; Creasey, E.A.; Edwards, M.; Lowe, M.; Isberg, R.R. SdhA blocks disruption of the Legionella-containing vacuole by hijacking the OCRL phosphatase. Cell Rep. 2021, 37, 109894. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, B.; Voth, K.; Price, C.T.; Jones, S.; Ozanic, M.; Santic, M.; Cygler, M.; Kwaik, Y.A. An Indispensable Role for the MavE Effector of Legionella pneumophila in Lysosomal Evasion. mBio 2021, 12, e03458-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Luo, Z.Q. The Legionella pneumophila effector SidJ is required for efficient recruitment of endoplasmic reticulum proteins to the bacterial phagosome. Infect. Immun. 2007, 75, 592–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glueck, N.K.; O’Brien, K.M.; Seguin, D.C.; Starai, V.J. Legionella pneumophila LegC7 effector protein drives aberrant endoplasmic reticulum:endosome contacts in yeast. Traffic 2021, 22, 284–302. [Google Scholar] [CrossRef]
- Arasaki, K.; Toomre, D.K.; Roy, C.R. The Legionella pneumophila effector DrrA is sufficient to stimulate SNARE-dependent membrane fusion. Cell Host Microbe 2012, 11, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Cirillo, S.L.G.; Lum, J.; Cirillo, J.D. Identification of novel loci involved in entry by Legionella pneumophila. Microbiology 2000, 146 Pt 6, 1345–1359. [Google Scholar] [CrossRef] [Green Version]
- Lally, E.T.; Kieba, I.R.; Sato, A.; Green, C.L.; Rosenbloom, J.; Korostoff, J.; Wang, J.F.; Shenker, B.J.; Ortlepp, S.; Robinson, M.K.; et al. RTX toxins recognize a beta2 integrin on the surface of human target cells. J. Biol. Chem. 1997, 272, 30463–30469. [Google Scholar] [CrossRef] [Green Version]
- Payne, N.R.; Horwitz, M.A. Phagocytosis of Legionella pneumophila is mediated by human monocyte complement receptors. J. Exp. Med. 1987, 166, 1377–1389. [Google Scholar] [CrossRef] [Green Version]
- Cirillo, S.L.; Bermudez, L.E.; El-Etr, S.H.; Duhamel, G.E.; Cirillo, J.D. Legionella pneumophila entry gene rtxA is involved in virulence. Infect. Immun. 2001, 69, 508–517. [Google Scholar] [CrossRef] [Green Version]
- D’Auria, G.; Jiménez, N.; Peris-Bondia, F.; Pelaz, C.; Latorre, A.; Moya, A. Virulence factor rtx in Legionella pneumophila, evidence suggesting it is a modular multifunctional protein. BMC Genom. 2008, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, S.L.G.; Yan, L.; Littman, M.; Samrakandi, M.M.; Cirillo, J.D. Role of the Legionella pneumophila rtxA gene in amoebae. Microbiology 2002, 148, 1667–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Conover, G.M.; Isberg, R.R. Legionella pneumophila EnhC is required for efficient replication in tumour necrosis factor alpha-stimulated macrophages. Cell. Microbiol. 2008, 10, 1906–1923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, H.J.; Sansom, F.M.; Dao, J.; McAlister, A.D.; Sloan, J.; Cianciotto, N.P.; Hartland, E.L. Sel1 repeat protein LpnE is a Legionella pneumophila virulence determinant that influences vacuolar trafficking. Infect. Immun. 2007, 75, 5575–5585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, B.J.; Kwaik, Y.A. Expression of multiple pili by Legionella pneumophila: Identification and characterization of a type IV pilin gene and its role in adherence to mammalian and protozoan cells. Infect. Immun. 1998, 66, 1768–1775. [Google Scholar] [CrossRef] [Green Version]
- Krinos, C.; High, A.S.; Rodgers, F.G. Role of the 25 kDa major outer membrane protein of Legionella pneumophila in attachment to U-937 cells and its potential as a virulence factor for chick embryos. J. Appl. Microbiol. 1999, 86, 237–244. [Google Scholar] [CrossRef]
- Wagner, C.; Khan, A.S.; Kamphausen, T.; Schmausser, B.; Unal, C.; Lorenz, U.; Fischer, G.; Hacker, J.; Steinert, M. Collagen binding protein Mip enables Legionella pneumophila to transmigrate through a barrier of NCI-H292 lung epithelial cells and extracellular matrix. Cell. Microbiol. 2007, 9, 450–462. [Google Scholar] [CrossRef]
- Cianciotto, N.P.; Fields, B.S. Legionella pneumophila mip gene potentiates intracellular infection of protozoa and human macrophages. Proc. Natl. Acad. Sci. USA 1992, 89, 5188–5191. [Google Scholar] [CrossRef] [Green Version]
- Köhler, R.; Fanghänel, J.; König, B.; Lüneberg, E.; Frosch, M.; Rahfeld, J.U.; Hilgenfeld, R.; Fischer, G.; Hacker, J.; Steinert, M. Biochemical and functional analyses of the Mip protein: Influence of the N-terminal half and of peptidylprolyl isomerase activity on the virulence of Legionella pneumophila. Infect. Immun. 2003, 71, 4389–4397. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Haenssler, E.; Uehara, T.; Losick, V.P.; Park, J.T.; Isberg, R.R. The Legionella pneumophila EnhC protein interferes with immunostimulatory muramyl peptide production to evade innate immunity. Cell Host Microbe 2012, 12, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Vandersmissen, L.; De Buck, E.; Saels, V.; Coil, D.A.; Anné, J. A Legionella pneumophila collagen-like protein encoded by a gene with a variable number of tandem repeats is involved in the adherence and invasion of host cells. FEMS Microbiol. Lett. 2010, 306, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Duncan, C.; Prashar, A.; So, J.; Tang, P.; Low, D.E.; Terebiznik, M.; Guyard, C. Lcl of Legionella pneumophila is an immunogenic GAG binding adhesin that promotes interactions with lung epithelial cells and plays a crucial role in biofilm formation. Infect. Immun. 2011, 79, 2168–2181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Nour, M.; Duncan, C.; Prashar, A.; Rao, C.; Ginevra, C.; Jarraud, S.; Low, D.E.; Ensminger, A.W.; Terebiznik, M.R.; Guyard, C. The Legionella pneumophila collagen-like protein mediates sedimentation, autoaggregation, and pathogen-phagocyte interactions. Appl. Environ. Microbiol. 2014, 80, 1441–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garduño, R.A.; Garduño, E.; Hoffman, P.S. Surface-associated hsp60 chaperonin of Legionella pneumophila mediates invasion in a HeLa cell model. Infect. Immun. 1998, 66, 4602–4610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, P.S.; Garduno, R.A. Surface-associated heat shock proteins of Legionella pneumophila and Helicobacter pylori: Roles in pathogenesis and immunity. Infect. Dis. Obstet. Gynecol. 1999, 7, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Bellinger-Kawahara, C.; Horwitz, M.A. Complement component C3 fixes selectively to the major outer membrane protein (MOMP) of Legionella pneumophila and mediates phagocytosis of liposome-MOMP complexes by human monocytes. J. Exp. Med. 1990, 172, 1201–1210. [Google Scholar] [CrossRef] [Green Version]
- Liles, M.R.; Viswanathan, V.K.; Cianciotto, N.P. Identification and temperature regulation of Legionella pneumophila genes involved in type IV pilus biogenesis and type II protein secretion. Infect. Immun. 1998, 66, 1776–1782. [Google Scholar] [CrossRef] [Green Version]
- Newton, H.J.; Sansom, F.M.; Dao, J.; Cazalet, C.; Bruggemann, H.; Albert-Weissenberger, C.; Buchrieser, C.; Cianciotto, N.P.; Hartland, E.L. Significant role for ladC in initiation of Legionella pneumophila infection. Infect. Immun. 2008, 76, 3075–3085. [Google Scholar] [CrossRef] [Green Version]
- Ren, T.; Zamboni, D.S.; Roy, C.R.; Dietrich, W.F.; Vance, R.E. Flagellin-deficient Legionella mutants evade caspase-1- and Naip5-mediated macrophage immunity. PLoS Pathog. 2006, 2, e18. [Google Scholar] [CrossRef]
- Pruckler, J.M.; Benson, R.F.; Moyenuddin, M.; Martin, W.T.; Fields, B.S. Association of flagellum expression and intracellular growth of Legionella pneumophila. Infect. Immun. 1995, 63, 4928–4932. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, C.; Heuner, K.; Brand, B.C.; Hacker, J.; Steinert, M. Flagellum of Legionella pneumophila positively affects the early phase of infection of eukaryotic host cells. Infect. Immun. 2001, 69, 2116–2122. [Google Scholar] [CrossRef]
- Cianciotto, N.P.; Eisenstein, B.I.; Mody, C.H.; Engleberg, N.C. A mutation in the mip gene results in an attenuation of Legionella pneumophila virulence. J. Infect. Dis. 1990, 162, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Cianciotto, N.P.; Stamos, J.K.; Kamp, D.W. Infectivity of Legionella pneumophila mip mutant for alveolar epithelial cells. Curr. Microbiol. 1995, 30, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Helbig, J.H.; König, B.; Knospe, H.; Bubert, B.; Yu, C.; Lück, C.P.; Riboldi-Tunnicliffe, A.; Hilgenfeld, R.; Jacobs, E.; Hacker, J.; et al. The PPIase active site of Legionella pneumophila Mip protein is involved in the infection of eukaryotic host cells. Biol. Chem. 2003, 384, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Viner, R.; Chetrit, D.; Ehrlich, M.; Segal, G. Identification of two Legionella pneumophila effectors that manipulate host phospholipids biosynthesis. PLoS Pathog. 2012, 8, e1002988. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, G.N.; Aurass, P.; Oates, C.V.; Tate, E.W.; Hartland, E.L.; Flieger, A.; Frankel, G. Legionella pneumophila Effector LpdA Is a Palmitoylated Phospholipase D Virulence Factor. Infect. Immun. 2015, 83, 3989–4002. [Google Scholar] [CrossRef] [Green Version]
- Gan, N.; Nakayasu, E.S.; Hollenbeck, P.J.; Luo, Z.Q. Legionella pneumophila inhibits immune signalling via MavC-mediated transglutaminase-induced ubiquitination of UBE2N. Nat. Microbiol. 2019, 4, 134–143. [Google Scholar] [CrossRef]
- Belyi, Y.; Tabakova, I.; Stahl, M.; Aktories, K. Lgt: A family of cytotoxic glucosyltransferases produced by Legionella pneumophila. J. Bacteriol. 2008, 190, 3026–3035. [Google Scholar] [CrossRef] [Green Version]
- Losick, V.P.; Haenssler, E.; Moy, M.Y.; Isberg, R.R. LnaB: A Legionella pneumophila activator of NF-kappaB. Cell. Microbiol. 2010, 12, 1083–1097. [Google Scholar] [CrossRef] [Green Version]
- Michard, C.; Sperandio, D.; Baïlo, N.; Pizarro-Cerdá, J.; LeClaire, L.; Chadeau-Argaud, E.; Pombo-Grégoire, I.; Hervet, E.; Vianney, A.; Gilbert, C.; et al. The Legionella Kinase LegK2 Targets the ARP2/3 Complex To Inhibit Actin Nucleation on Phagosomes and Allow Bacterial Evasion of the Late Endocytic Pathway. mBio 2015, 6, e00354-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothmeier, E.; Pfaffinger, G.; Hoffmann, C.; Harrison, C.F.; Grabmayr, H.; Repnik, U.; Hannemann, M.; Wölke, S.; Bausch, A.; Griffiths, G.; et al. Activation of Ran GTPase by a Legionella effector promotes microtubule polymerization, pathogen vacuole motility and infection. PLoS Pathog. 2013, 9, e1003598. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Wagner, M.A.; Rothmeier, E.; Müller-Taubenberger, A.; Hilbi, H. Icm/Dot-dependent inhibition of phagocyte migration by Legionella is antagonized by a translocated Ran GTPase activator. Cell. Microbiol. 2014, 16, 977–992. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Lu, Q.; Wang, G.; Xu, H.; Huang, H.; Cai, T.; Kan, B.; Ge, J.; Shao, F. SET-domain bacterial effectors target heterochromatin protein 1 to activate host rDNA transcription. EMBO Rep. 2013, 14, 733–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horenkamp, F.A.; Mukherjee, S.; Alix, E.; Schauder, C.M.; Hubber, A.M.; Roy, C.R.; Reinisch, K.M. Legionella pneumophila subversion of host vesicular transport by SidC effector proteins. Traffic 2014, 15, 488–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.; Luo, Z.Q. Legionella pneumophila SidD is a deAMPylase that modifies Rab1. Nature 2011, 475, 506–509. [Google Scholar] [CrossRef] [Green Version]
- Kotewicz, K.M.; Ramabhadran, V.; Sjoblom, N.; Vogel, J.P.; Haenssler, E.; Zhang, M.; Behringer, J.; Scheck, R.A.; Isberg, R.R. A Single Legionella Effector Catalyzes a Multistep Ubiquitination Pathway to Rearrange Tubular Endoplasmic Reticulum for Replication. Cell Host Microbe 2017, 21, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.; Masclef, L.; Affar, E.B. Surviving the host-pathogen interaction: Legionella uses its SidE arsenal to mediate phosphoribosyl-linked ubiquitination of Golgi proteins inducing organelle fragmentation. Cell Death Differ. 2021, 28, 2986–2988. [Google Scholar] [CrossRef]
- Xu, L.; Shen, X.; Bryan, A.; Banga, S.; Swanson, M.S.; Luo, Z.Q. Inhibition of host vacuolar H+-ATPase activity by a Legionella pneumophila effector. PLoS Pathog. 2010, 6, e1000822. [Google Scholar] [CrossRef] [Green Version]
- Sulpizio, A.G.; Minelli, M.E.; Mao, Y. Glutamylation of Bacterial Ubiquitin Ligases by a Legionella Pseudokinase. Trends Microbiol. 2019, 27, 967–969. [Google Scholar] [CrossRef]
- Toulabi, L.; Wu, X.; Cheng, Y.; Mao, Y. Identification and structural characterization of a Legionella phosphoinositide phosphatase. J. Biol. Chem. 2013, 288, 24518–24527. [Google Scholar] [CrossRef] [Green Version]
- Murata, T.; Delprato, A.; Ingmundson, A.; Toomre, D.K.; Lambright, D.G.; Roy, C.R. The Legionella pneumophila effector protein DrrA is a Rab1 guanine nucleotide-exchange factor. Nat. Cell Biol. 2006, 8, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, P.; Lang, E.A.; Rasaputra, K.S.; Steinman, H.M. Implication of the VirD4 coupling protein of the Lvh type 4 secretion system in virulence phenotypes of Legionella pneumophila. J. Bacteriol. 2013, 195, 3468–3475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, H.J.; Sansom, F.M.; Bennett-Wood, V.; Hartland, E.L. Identification of Legionella pneumophila-specific genes by genomic subtractive hybridization with Legionella micdadei and identification of lpnE, a gene required for efficient host cell entry. Infect. Immun. 2006, 74, 1683–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelstein, P.H.; Hu, B.; Shinzato, T.; Edelstein, M.A.; Xu, W.; Bessman, M.J. Legionella pneumophila NudA Is a Nudix hydrolase and virulence factor. Infect. Immun. 2005, 73, 6567–6576. [Google Scholar] [CrossRef] [Green Version]
- Graham, C.I.; Patel, P.G.; Tanner, J.R.; Hellinga, J.; MacMartin, T.L.; Hausner, G.; Brassinga, A.K.C. Autorepressor PsrA is required for optimal Legionella pneumophila growth in Acanthamoeba castellanii protozoa. Mol. Microbiol. 2021, 116, 624–647. [Google Scholar] [CrossRef]
- Morar, M.; Evdokimova, E.; Chang, C.; Ensminger, A.W.; Savchenko, A. Crystal structure of the Legionella pneumophila Lem10 effector reveals a new member of the HD protein superfamily. Proteins 2015, 83, 2319–2325. [Google Scholar] [CrossRef] [Green Version]
- Voth, K.; Pasricha, S.; Chung, I.Y.W.; Wibawa, R.R.; Zainudin, E.; Hartland, E.L.; Cygler, M. Structural and Functional Characterization of Legionella pneumophila Effector MavL. Biomolecules 2021, 11, 1802. [Google Scholar] [CrossRef]
- Hsieh, T.S.; Lopez, V.A.; Black, M.H.; Osinski, A.; Pawlowski, K.; Tomchick, D.R.; Liou, J.; Tagliabracci, V.S. Dynamic remodeling of host membranes by self-organizing bacterial effectors. Science 2021, 372, 935–941. [Google Scholar] [CrossRef]
- Harding, C.R.; Mattheis, C.; Mousnier, A.; Oates, C.V.; Hartland, E.L.; Frankel, G.; Schroeder, G.N. LtpD is a novel Legionella pneumophila effector that binds phosphatidylinositol 3-phosphate and inositol monophosphatase IMPA1. Infect. Immun. 2013, 81, 4261–4270. [Google Scholar] [CrossRef] [Green Version]
- Levanova, N.; Mattheis, C.; Carson, D.; To, K.N.; Jank, T.; Frankel, G.; Aktories, K.; Schroeder, G.N. The Legionella effector LtpM is a new type of phosphoinositide-activated glucosyltransferase. J. Biol. Chem. 2019, 294, 2862–2879. [Google Scholar] [CrossRef] [Green Version]
- Beck, W.H.J.; Kim, D.; Das, J.; Yu, H.; Smolka, M.B.; Mao, Y. Glucosylation by the Legionella Effector SetA Promotes the Nuclear Localization of the Transcription Factor TFEB. iScience 2020, 23, 101300. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Song, Q.; Liang, H.; Zhu, Y.; Wei, T.; Dong, N.; Xiao, J.; Shao, F.; Lai, L.; Chen, X. Legionella effector SetA as a general O-glucosyltransferase for eukaryotic proteins. Nat. Chem. Biol. 2019, 15, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Finsel, I.; Ragaz, C.; Hoffmann, C.; Harrison, C.F.; Weber, S.; van Rahden, V.A.; Johannes, L.; Hilbi, H. The Legionella effector RidL inhibits retrograde trafficking to promote intracellular replication. Cell Host Microbe 2013, 14, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Banga, S.; Liu, Y.; Xu, L.; Gao, P.; Shamovsky, I.; Nudler, E.; Luo, Z.Q. Targeting eEF1A by a Legionella pneumophila effector leads to inhibition of protein synthesis and induction of host stress response. Cell. Microbiol. 2009, 11, 911–926. [Google Scholar] [CrossRef] [Green Version]
- McCloskey, A.; Perri, K.; Chen, T.; Han, A.; Luo, Z.Q. The metaeffector MesI regulates the activity of the Legionella effector SidI through direct protein-protein interactions. Microbes Infect. 2021, 23, 104794. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; McEwen, J.M.; Sjoblom, N.M.; Kotewicz, K.M.; Isberg, R.R.; Scheck, R.A. Members of the Legionella pneumophila Sde family target tyrosine residues for phosphoribosyl-linked ubiquitination. RSC Chem. Biol. 2021, 2, 1509–1519. [Google Scholar] [CrossRef]
- Mousnier, A.; Schroeder, G.N.; Stoneham, C.A.; So, E.C.; Garnett, J.A.; Yu, L.; Matthews, S.J.; Choudhary, J.S.; Hartland, E.L.; Frankel, G. A new method to determine in vivo interactomes reveals binding of the Legionella pneumophila effector PieE to multiple rab GTPases. mBio 2014, 5, e01148-14. [Google Scholar] [CrossRef] [Green Version]
- Harding, C.R.; Stoneham, C.A.; Schuelein, R.; Newton, H.; Oates, C.V.; Hartland, E.L.; Schroeder, G.N.; Frankel, G. The Dot/Icm effector SdhA is necessary for virulence of Legionella pneumophila in Galleria mellonella and A/J mice. Infect. Immun. 2013, 81, 2598–2605. [Google Scholar] [CrossRef] [Green Version]
- Black, M.H.; Osinski, A.; Park, G.J.; Gradowski, M.; Servage, K.A.; Pawłowski, K.; Tagliabracci, V.S. A Legionella effector ADP-ribosyltransferase inactivates glutamate dehydrogenase. J. Biol. Chem. 2021, 296, 100301. [Google Scholar] [CrossRef]
- Pike, C.M.; Boyer-Andersen, R.; Kinch, L.N.; Caplan, J.L.; Neunuebel, M.R. The Legionella effector RavD binds phosphatidylinositol-3-phosphate and helps suppress endolysosomal maturation of the Legionella-containing vacuole. J. Biol. Chem. 2019, 294, 6405–6415. [Google Scholar] [CrossRef]
- Wan, M.; Wang, X.; Huang, C.; Xu, D.; Wang, Z.; Zhou, Y.; Zhu, Y. A bacterial effector deubiquitinase specifically hydrolyses linear ubiquitin chains to inhibit host inflammatory signalling. Nat. Microbiol. 2019, 4, 1282–1293. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, W.; Tan, Y.; Nakayasu, E.S.; Staiger, C.J.; Luo, Z.Q. A Legionella Effector Disrupts Host Cytoskeletal Structure by Cleaving Actin. PLoS Pathog. 2017, 13, e1006186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aurass, P.; Schlegel, M.; Metwally, O.; Harding, C.R.; Schroeder, G.N.; Frankel, G.; Flieger, A. The Legionella pneumophila Dot/Icm-secreted effector PlcC/CegC1 together with PlcA and PlcB promotes virulence and belongs to a novel zinc metallophospholipase C family present in bacteria and fungi. J. Biol. Chem. 2013, 288, 11080–11092. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Zhou, M.; Gritsenko, M.A.; Nakayasu, E.S.; Song, L.; Luo, Z.Q. Legionella pneumophila modulates host energy metabolism by ADP-ribosylation of ADP/ATP translocases. eLife 2022, 11, e73611. [Google Scholar] [CrossRef] [PubMed]
- Quaile, A.T.; Stogios, P.J.; Egorova, O.; Evdokimova, E.; Valleau, D.; Nocek, B.; Kompella, P.S.; Peisajovich, S.; Yakunin, A.F.; Ensminger, A.W.; et al. The Legionella pneumophila effector Ceg4 is a phosphotyrosine phosphatase that attenuates activation of eukaryotic MAPK pathways. J. Biol. Chem. 2018, 293, 3307–3320. [Google Scholar] [CrossRef] [Green Version]
- Franco, I.S.; Shohdy, N.; Shuman, H.A. The Legionella pneumophila effector VipA is an actin nucleator that alters host cell organelle trafficking. PLoS Pathog. 2012, 8, e1002546. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.Q.; Isberg, R.R. Multiple substrates of the Legionella pneumophila Dot/Icm system identified by interbacterial protein transfer. Proc. Natl. Acad. Sci. USA 2004, 101, 841–846. [Google Scholar] [CrossRef] [Green Version]
- Jeng, E.E.; Bhadkamkar, V.; Ibe, N.U.; Gause, H.; Jiang, L.; Chan, J.; Jian, R.; Jimenez-Morales, D.; Stevenson, E.; Krogan, N.J.; et al. Systematic Identification of Host Cell Regulators of Legionella pneumophila Pathogenesis Using a Genome-wide CRISPR Screen. Cell Host Microbe 2019, 26, 551–563.e556. [Google Scholar] [CrossRef]
- Bartfeld, S.; Engels, C.; Bauer, B.; Aurass, P.; Flieger, A.; Brüggemann, H.; Meyer, T.F. Temporal resolution of two-tracked NF-kappaB activation by Legionella pneumophila. Cell. Microbiol. 2009, 11, 1638–1651. [Google Scholar] [CrossRef]
- Price, C.; Jones, S.; Mihelcic, M.; Santic, M.; Kwaik, Y.A. Paradoxical Pro-inflammatory Responses by Human Macrophages to an Amoebae Host-Adapted Legionella Effector. Cell Host Microbe 2020, 27, 571–584.e577. [Google Scholar] [CrossRef]
- Liu, S.; Luo, J.; Zhen, X.; Qiu, J.; Ouyang, S.; Luo, Z.Q. Interplay between bacterial deubiquitinase and ubiquitin E3 ligase regulates ubiquitin dynamics on Legionella phagosomes. eLife 2020, 9, e58114. [Google Scholar] [CrossRef] [PubMed]
- Al-Khodor, S.; Kalachikov, S.; Morozova, I.; Price, C.T.; Kwaik, Y.A. The PmrA/PmrB two-component system of Legionella pneumophila is a global regulator required for intracellular replication within macrophages and protozoa. Infect. Immun. 2009, 77, 374–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folly-Klan, M.; Alix, E.; Stalder, D.; Ray, P.; Duarte, L.V.; Delprato, A.; Zeghouf, M.; Antonny, B.; Campanacci, V.; Roy, C.R.; et al. A novel membrane sensor controls the localization and ArfGEF activity of bacterial RalF. PLoS Pathog. 2013, 9, e1003747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neunuebel, M.R.; Mohammadi, S.; Jarnik, M.; Machner, M.P. Legionella pneumophila LidA affects nucleotide binding and activity of the host GTPase Rab1. J. Bacteriol. 2012, 194, 1389–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Reyes, M.; Clarke, M.; Shuman, H.A. Host cell-dependent secretion and translocation of the LepA and LepB effectors of Legionella pneumophila. Cell. Microbiol. 2007, 9, 1660–1671. [Google Scholar] [CrossRef]
- Weber, S.; Stirnimann, C.U.; Wieser, M.; Frey, D.; Meier, R.; Engelhardt, S.; Li, X.; Capitani, G.; Kammerer, R.A.; Hilbi, H. A type IV translocated Legionella cysteine phytase counteracts intracellular growth restriction by phytate. J. Biol. Chem. 2014, 289, 34175–34188. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Yin, S.; Zhang, W.; Gong, X.; Zhang, N.; Fang, K.; Ge, H. Crystal Structure and Biochemical Characterization of an Aminopeptidase LapB from Legionella pneumophila. J. Agric. Food Chem. 2017, 65, 7569–7578. [Google Scholar] [CrossRef]
- Banerji, S.; Bewersdorff, M.; Hermes, B.; Cianciotto, N.P.; Flieger, A. Characterization of the major secreted zinc metalloprotease- dependent glycerophospholipid:cholesterol acyltransferase, PlaC, of Legionella pneumophila. Infect. Immun. 2005, 73, 2899–2909. [Google Scholar] [CrossRef] [Green Version]
- Tyson, J.Y.; Pearce, M.M.; Vargas, P.; Bagchi, S.; Mulhern, B.J.; Cianciotto, N.P. Multiple Legionella pneumophila Type II secretion substrates, including a novel protein, contribute to differential infection of the amoebae Acanthamoeba castellanii, Hartmannella vermiformis, and Naegleria lovaniensis. Infect. Immun. 2013, 81, 1399–1410. [Google Scholar] [CrossRef] [Green Version]
- Portlock, T.J.; Tyson, J.Y.; Dantu, S.C.; Rehman, S.; White, R.C.; McIntire, I.E.; Sewell, L.; Richardson, K.; Shaw, R.; Pandini, A.; et al. Structure, Dynamics and Cellular Insight Into Novel Substrates of the Legionella pneumophila Type II Secretion System. Front. Mol. Biosci. 2020, 7, 112. [Google Scholar] [CrossRef]
- Scheithauer, L.; Thiem, S.; Schmelz, S.; Dellmann, A.; Büssow, K.; Brouwer, R.; Ünal, C.M.; Blankenfeldt, W.; Steinert, M. Zinc metalloprotease ProA of Legionella pneumophila increases alveolar septal thickness in human lung tissue explants by collagen IV degradation. Cell. Microbiol. 2021, 23, e13313. [Google Scholar] [CrossRef] [PubMed]
- Scheithauer, L.; Thiem, S.; Ünal, C.M.; Dellmann, A.; Steinert, M. Zinc Metalloprotease ProA from Legionella pneumophila Inhibits the Pro-Inflammatory Host Response by Degradation of Bacterial Flagellin. Biomolecules 2022, 12, 624. [Google Scholar] [CrossRef] [PubMed]
- DebRoy, S.; Dao, J.; Söderberg, M.; Rossier, O.; Cianciotto, N.P. Legionella pneumophila type II secretome reveals unique exoproteins and a chitinase that promotes bacterial persistence in the lung. Proc. Natl. Acad. Sci. USA 2006, 103, 19146–19151. [Google Scholar] [CrossRef] [Green Version]
- Schunder, E.; Adam, P.; Higa, F.; Remer, K.A.; Lorenz, U.; Bender, J.; Schulz, T.; Flieger, A.; Steinert, M.; Heuner, K. Phospholipase PlaB is a new virulence factor of Legionella pneumophila. Int. J. Med. Microbiol. 2010, 300, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Sahr, T.; Escoll, P.; Rusniok, C.; Bui, S.; Pehau-Arnaudet, G.; Lavieu, G.; Buchrieser, C. Translocated Legionella pneumophila small RNAs mimic eukaryotic microRNAs targeting the host immune response. Nat. Commun. 2022, 13, 762. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Park, B.; Park, G.; Kim, J.; Lim, S.A.; Lee, K.M. Innate immunity against Legionella pneumophila during pulmonary infections in mice. Arch. Pharmacal. Res. 2017, 40, 131–145. [Google Scholar] [CrossRef]
- Naujoks, J.; Lippmann, J.; Suttorp, N.; Opitz, B. Innate sensing and cell-autonomous resistance pathways in Legionella pneumophila infection. Int. J. Med. Microbiol. 2018, 308, 161–167. [Google Scholar] [CrossRef]
- Ricci, M.L.; Torosantucci, A.; Scaturro, M.; Chiani, P.; Baldassarri, L.; Pastoris, M.C. Induction of protective immunity by Legionella pneumophila flagellum in an A/J mouse model. Vaccine 2005, 23, 4811–4820. [Google Scholar] [CrossRef]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Hawn, T.R.; Verbon, A.; Lettinga, K.D.; Zhao, L.P.; Li, S.S.; Laws, R.J.; Skerrett, S.J.; Beutler, B.; Schroeder, L.; Nachman, A.; et al. A common dominant TLR5 stop codon polymorphism abolishes flagellin signaling and is associated with susceptibility to legionnaires’ disease. J. Exp. Med. 2003, 198, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Akamine, M.; Higa, F.; Arakaki, N.; Kawakami, K.; Takeda, K.; Akira, S.; Saito, A. Differential roles of Toll-like receptors 2 and 4 in in vitro responses of macrophages to Legionella pneumophila. Infect. Immun. 2005, 73, 352–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawn, T.R.; Smith, K.D.; Aderem, A.; Skerrett, S.J. Myeloid differentiation primary response gene (88) and toll-like receptor 2-deficient mice are susceptible to infection with aerosolized Legionella pneumophila. J. Infect. Dis. 2006, 193, 1693–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braedel-Ruoff, S.; Faigle, M.; Hilf, N.; Neumeister, B.; Schild, H. Legionella pneumophila mediated activation of dendritic cells involves CD14 and TLR2. J. Endotoxin Res. 2005, 11, 89–96. [Google Scholar] [CrossRef]
- Girard, R.; Pedron, T.; Uematsu, S.; Balloy, V.; Chignard, M.; Akira, S.; Chaby, R. Lipopolysaccharides from Legionella and Rhizobium stimulate mouse bone marrow granulocytes via Toll-like receptor 2. J. Cell Sci. 2003, 116, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Grigoryeva, L.S.; Cianciotto, N.P. Human macrophages utilize a wide range of pathogen recognition receptors to recognize Legionella pneumophila, including Toll-Like Receptor 4 engaging Legionella lipopolysaccharide and the Toll-like Receptor 3 nucleic-acid sensor. PLoS Pathog. 2021, 17, e1009781. [Google Scholar] [CrossRef]
- Cunha, L.D.; Zamboni, D.S. Recognition of Legionella pneumophila nucleic acids by innate immune receptors. Microbes Infect. 2014, 16, 985–990. [Google Scholar] [CrossRef]
- Hawn, T.R.; Berrington, W.R.; Smith, I.A.; Uematsu, S.; Akira, S.; Aderem, A.; Smith, K.D.; Skerrett, S.J. Altered inflammatory responses in TLR5-deficient mice infected with Legionella pneumophila. J. Immunol. 2007, 179, 6981–6987. [Google Scholar] [CrossRef] [Green Version]
- Newton, C.A.; Perkins, I.; Widen, R.H.; Friedman, H.; Klein, T.W. Role of Toll-like receptor 9 in Legionella pneumophila-induced interleukin-12 p40 production in bone marrow-derived dendritic cells and macrophages from permissive and nonpermissive mice. Infect. Immun. 2007, 75, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Archer, K.A.; Roy, C.R. MyD88-dependent responses involving toll-like receptor 2 are important for protection and clearance of Legionella pneumophila in a mouse model of Legionnaires’ disease. Infect. Immun. 2006, 74, 3325–3333. [Google Scholar] [CrossRef] [Green Version]
- Archer, K.A.; Alexopoulou, L.; Flavell, R.A.; Roy, C.R. Multiple MyD88-dependent responses contribute to pulmonary clearance of Legionella pneumophila. Cell. Microbiol. 2009, 11, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Scharf, S.; Hippenstiel, S.; Flieger, A.; Suttorp, N.; N’Guessan, P.D. Induction of human β-defensin-2 in pulmonary epithelial cells by Legionella pneumophila: Involvement of TLR2 and TLR5, p38 MAPK, JNK, NF-κB, and AP-1. Am. J. Physiology. Lung Cell. Mol. Physiol. 2010, 298, L687–L695. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Case, C.L.; Archer, K.A.; Nogueira, C.V.; Kobayashi, K.S.; Flavell, R.A.; Roy, C.R.; Zamboni, D.S. Type IV secretion-dependent activation of host MAP kinases induces an increased proinflammatory cytokine response to Legionella pneumophila. PLoS Pathog. 2008, 4, e1000220. [Google Scholar] [CrossRef] [PubMed]
- Molofsky, A.B.; Byrne, B.G.; Whitfield, N.N.; Madigan, C.A.; Fuse, E.T.; Tateda, K.; Swanson, M.S. Cytosolic recognition of flagellin by mouse macrophages restricts Legionella pneumophila infection. J. Exp. Med. 2006, 203, 1093–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamboni, D.S.; Kobayashi, K.S.; Kohlsdorf, T.; Ogura, Y.; Long, E.M.; Vance, R.E.; Kuida, K.; Mariathasan, S.; Dixit, V.M.; Flavell, R.A.; et al. The Birc1e cytosolic pattern-recognition receptor contributes to the detection and control of Legionella pneumophila infection. Nat. Immunol. 2006, 7, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Amer, A.; Franchi, L.; Kanneganti, T.D.; Body-Malapel, M.; Ozören, N.; Brady, G.; Meshinchi, S.; Jagirdar, R.; Gewirtz, A.; Akira, S.; et al. Regulation of Legionella phagosome maturation and infection through flagellin and host Ipaf. J. Biol. Chem. 2006, 281, 35217–35223. [Google Scholar] [CrossRef] [Green Version]
- Case, C.L.; Shin, S.; Roy, C.R. Asc and Ipaf Inflammasomes direct distinct pathways for caspase-1 activation in response to Legionella pneumophila. Infect. Immun. 2009, 77, 1981–1991. [Google Scholar] [CrossRef] [Green Version]
- Kanneganti, T.D.; Ozören, N.; Body-Malapel, M.; Amer, A.; Park, J.H.; Franchi, L.; Whitfield, J.; Barchet, W.; Colonna, M.; Vandenabeele, P.; et al. Bacterial RNA and small antiviral compounds activate caspase-1 through cryopyrin/Nalp3. Nature 2006, 440, 233–236. [Google Scholar] [CrossRef] [Green Version]
- Bürckstümmer, T.; Baumann, C.; Blüml, S.; Dixit, E.; Dürnberger, G.; Jahn, H.; Planyavsky, M.; Bilban, M.; Colinge, J.; Bennett, K.L.; et al. An orthogonal proteomic-genomic screen identifies AIM2 as a cytoplasmic DNA sensor for the inflammasome. Nat. Immunol. 2009, 10, 266–272. [Google Scholar] [CrossRef]
- Ge, J.; Gong, Y.N.; Xu, Y.; Shao, F. Preventing bacterial DNA release and absent in melanoma 2 inflammasome activation by a Legionella effector functioning in membrane trafficking. Proc. Natl. Acad. Sci. USA 2012, 109, 6193–6198. [Google Scholar] [CrossRef] [PubMed]
- Case, C.L.; Kohler, L.J.; Lima, J.B.; Strowig, T.; de Zoete, M.R.; Flavell, R.A.; Zamboni, D.S.; Roy, C.R. Caspase-11 stimulates rapid flagellin-independent pyroptosis in response to Legionella pneumophila. Proc. Natl. Acad. Sci. USA 2013, 110, 1851–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerqueira, D.M.; Pereira, M.S.; Silva, A.L.; Cunha, L.D.; Zamboni, D.S. Caspase-1 but Not Caspase-11 Is Required for NLRC4-Mediated Pyroptosis and Restriction of Infection by Flagellated Legionella Species in Mouse Macrophages and In Vivo. J. Immunol. 2015, 195, 2303–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mascarenhas, D.P.A.; Cerqueira, D.M.; Pereira, M.S.F.; Castanheira, F.V.S.; Fernandes, T.D.; Manin, G.Z.; Cunha, L.D.; Zamboni, D.S. Inhibition of caspase-1 or gasdermin-D enable caspase-8 activation in the Naip5/NLRC4/ASC inflammasome. PLoS Pathog. 2017, 13, e1006502. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, A.V.; Margolis, S.R.; Quirino, G.F.S.; Mascarenhas, D.P.A.; Rauch, I.; Nichols, R.D.; Ansaldo, E.; Fontana, M.F.; Vance, R.E.; Zamboni, D.S. Gasdermin-D and Caspase-7 are the key Caspase-1/8 substrates downstream of the NAIP5/NLRC4 inflammasome required for restriction of Legionella pneumophila. PLoS Pathog. 2019, 15, e1007886. [Google Scholar] [CrossRef]
- Schiavoni, G.; Mauri, C.; Carlei, D.; Belardelli, F.; Pastoris, M.C.; Proietti, E. Type I IFN protects permissive macrophages from Legionella pneumophila infection through an IFN-gamma-independent pathway. J. Immunol. 2004, 173, 1266–1275. [Google Scholar] [CrossRef] [Green Version]
- Salins, S.; Newton, C.; Widen, R.; Klein, T.W.; Friedman, H. Differential induction of gamma interferon in Legionella pneumophila-infected macrophages from BALB/c and A/J mice. Infect. Immun. 2001, 69, 3605–3610. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.S.; Yang, C.; Fung, K.Y.; Bachem, A.; Bourges, D.; Bedoui, S.; Hartland, E.L.; van Driel, I.R. Cooperation between Monocyte-Derived Cells and Lymphoid Cells in the Acute Response to a Bacterial Lung Pathogen. PLoS Pathog. 2016, 12, e1005691. [Google Scholar] [CrossRef] [Green Version]
- Copenhaver, A.M.; Casson, C.N.; Nguyen, H.T.; Fung, T.C.; Duda, M.M.; Roy, C.R.; Shin, S. Alveolar macrophages and neutrophils are the primary reservoirs for Legionella pneumophila and mediate cytosolic surveillance of type IV secretion. Infect. Immun. 2014, 82, 4325–4336. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; McDermot, D.S.; Pasricha, S.; Brown, A.S.; Bedoui, S.; Lenz, L.L.; van Driel, I.R.; Hartland, E.L. IFNγ receptor down-regulation facilitates Legionella survival in alveolar macrophages. J. Leukoc. Biol. 2020, 107, 273–284. [Google Scholar] [CrossRef]
- Naujoks, J.; Tabeling, C.; Dill, B.D.; Hoffmann, C.; Brown, A.S.; Kunze, M.; Kempa, S.; Peter, A.; Mollenkopf, H.J.; Dorhoi, A.; et al. IFNs Modify the Proteome of Legionella-Containing Vacuoles and Restrict Infection Via IRG1-Derived Itaconic Acid. PLoS Pathog. 2016, 12, e1005408. [Google Scholar] [CrossRef]
- Nathan, C.; Shiloh, M.U. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci. USA 2000, 97, 8841–8848. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.S.; Yang, C.; Hartland, E.L.; van Driel, I.R. The regulation of acute immune responses to the bacterial lung pathogen Legionella pneumophila. J. Leukoc. Biol. 2017, 101, 875–886. [Google Scholar] [CrossRef]
- Mallama, C.A.; McCoy-Simandle, K.; Cianciotto, N.P. The Type II Secretion System of Legionella pneumophila Dampens the MyD88 and Toll-Like Receptor 2 Signaling Pathway in Infected Human Macrophages. Infect. Immun. 2017, 85, e00897-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoy-Simandle, K.; Stewart, C.R.; Dao, J.; DebRoy, S.; Rossier, O.; Bryce, P.J.; Cianciotto, N.P. Legionella pneumophila type II secretion dampens the cytokine response of infected macrophages and epithelia. Infect. Immun. 2011, 79, 1984–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiaden, A.; Spirig, T.; Weber, S.S.; Brüggemann, H.; Bosshard, R.; Buchrieser, C.; Hilbi, H. The Legionella pneumophila response regulator LqsR promotes host cell interactions as an element of the virulence regulatory network controlled by RpoS and LetA. Cell. Microbiol. 2007, 9, 2903–2920. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Luo, J.; Wang, H.; Huang, D.; Tan, Y.; Liu, Y.; Wang, Y.; Yu, K.; Zhang, Y.; Liu, X.; et al. Legionella pneumophila regulates host cell motility by targeting Phldb2 with a 14-3-3ζ-dependent protease effector. eLife 2022, 11, e73220. [Google Scholar] [CrossRef]
- Fontana, M.F.; Banga, S.; Barry, K.C.; Shen, X.; Tan, Y.; Luo, Z.Q.; Vance, R.E. Secreted bacterial effectors that inhibit host protein synthesis are critical for induction of the innate immune response to virulent Legionella pneumophila. PLoS Pathog. 2011, 7, e1001289. [Google Scholar] [CrossRef]
- Losick, V.P.; Isberg, R.R. NF-kappaB translocation prevents host cell death after low-dose challenge by Legionella pneumophila. J. Exp. Med. 2006, 203, 2177–2189. [Google Scholar] [CrossRef] [Green Version]
- Abu-Zant, A.; Jones, S.; Asare, R.; Suttles, J.; Price, C.; Graham, J.; Kwaik, Y.A. Anti-apoptotic signalling by the Dot/Icm secretion system of L. pneumophila. Cell. Microbiol. 2007, 9, 246–264. [Google Scholar] [CrossRef]
- Schmeck, B.; N’Guessan, P.D.; Ollomang, M.; Lorenz, J.; Zahlten, J.; Opitz, B.; Flieger, A.; Suttorp, N.; Hippenstiel, S. Legionella pneumophila-induced NF-kappaB- and MAPK-dependent cytokine release by lung epithelial cells. Eur. Respir. J. 2007, 29, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Xu, H.; Li, T.; Zhou, Y.; Zhang, Z.; Li, S.; Liu, L.; Shao, F. A Legionella type IV effector activates the NF-kappaB pathway by phosphorylating the IkappaB family of inhibitors. Proc. Natl. Acad. Sci. USA 2009, 106, 13725–13730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, N.; Nash, T.W.; Horwitz, M.A. Interferon-gamma-activated human monocytes inhibit the intracellular multiplication of Legionella pneumophila. J. Immunol. 1986, 137, 2662–2669. [Google Scholar]
- Nash, T.W.; Libby, D.M.; Horwitz, M.A. IFN-gamma-activated human alveolar macrophages inhibit the intracellular multiplication of Legionella pneumophila. J. Immunol. 1988, 140, 3978–3981. [Google Scholar] [PubMed]
- Byrd, T.F.; Horwitz, M.A. Interferon gamma-activated human monocytes downregulate transferrin receptors and inhibit the intracellular multiplication of Legionella pneumophila by limiting the availability of iron. J. Clin. Investig. 1989, 83, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Khweek, A.A.; Dávila, N.S.F.; Caution, K.; Akhter, A.; Abdulrahman, B.A.; Tazi, M.; Hassan, H.; Novotny, L.A.; Bakaletz, L.O.; Amer, A.O. Biofilm-derived Legionella pneumophila evades the innate immune response in macrophages. Front. Cell. Infect. Microbiol. 2013, 3, 18. [Google Scholar] [CrossRef] [PubMed]
- Akhter, A.; Gavrilin, M.A.; Frantz, L.; Washington, S.; Ditty, C.; Limoli, D.; Day, C.; Sarkar, A.; Newland, C.; Butchar, J.; et al. Caspase-7 activation by the Nlrc4/Ipaf inflammasome restricts Legionella pneumophila infection. PLoS Pathog. 2009, 5, e1000361. [Google Scholar] [CrossRef] [Green Version]
- Byrne, B.G.; Dubuisson, J.F.; Joshi, A.D.; Persson, J.J.; Swanson, M.S. Inflammasome components coordinate autophagy and pyroptosis as macrophage responses to infection. mBio 2013, 4, e00620-12. [Google Scholar] [CrossRef] [Green Version]
- Mou, Q.; Leung, P.H.M. Differential expression of virulence genes in Legionella pneumophila growing in Acanthamoeba and human monocytes. Virulence 2018, 9, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Massis, L.M.; Assis-Marques, M.A.; Castanheira, F.V.; Capobianco, Y.J.; Balestra, A.C.; Escoll, P.; Wood, R.E.; Manin, G.Z.; Correa, V.M.; Alves-Filho, J.C.; et al. Legionella longbeachae Is Immunologically Silent and Highly Virulent In Vivo. J. Infect. Dis. 2017, 215, 440–451. [Google Scholar] [CrossRef] [Green Version]
- Asare, R.; Santic, M.; Gobin, I.; Doric, M.; Suttles, J.; Graham, J.E.; Price, C.D.; Kwaik, Y.A. Genetic susceptibility and caspase activation in mouse and human macrophages are distinct for Legionella longbeachae and L. pneumophila. Infect. Immun. 2007, 75, 1933–1945. [Google Scholar] [CrossRef] [Green Version]
- McHugh, S.L.; Yamamoto, Y.; Klein, T.W.; Friedman, H. Murine macrophages differentially produce proinflammatory cytokines after infection with virulent vs. avirulent Legionella pneumophila. J. Leukoc. Biol. 2000, 67, 863–868. [Google Scholar] [CrossRef] [PubMed]
- McHugh, S.; Yamamoto, Y.; Klein, T.W.; Friedman, H. Differential expression of IL-1 and TNF receptors in murine macrophages infected with virulent vs. avirulent Legionella pneumophila. Can. J. Microbiol. 2000, 46, 885–891. [Google Scholar] [CrossRef]
- Nakachi, N.; Matsunaga, K.; Klein, T.W.; Friedman, H.; Yamamoto, Y. Differential effects of virulent versus avirulent Legionella pneumophila on chemokine gene expression in murine alveolar macrophages determined by cDNA expression array technique. Infect. Immun. 2000, 68, 6069–6072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemot, J.; Ginevra, C.; Allam, C.; Kay, E.; Gilbert, C.; Doublet, P.; Jarraud, S.; Chapalain, A. TNF-α response in macrophages depends on clinical Legionella pneumophila isolates genotypes. Virulence 2022, 13, 160–173. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.-L.; Li, D.; Zhan, X.-Y. Concept about the Virulence Factor of Legionella. Microorganisms 2023, 11, 74. https://doi.org/10.3390/microorganisms11010074
Yang J-L, Li D, Zhan X-Y. Concept about the Virulence Factor of Legionella. Microorganisms. 2023; 11(1):74. https://doi.org/10.3390/microorganisms11010074
Chicago/Turabian StyleYang, Jin-Lei, Danyang Li, and Xiao-Yong Zhan. 2023. "Concept about the Virulence Factor of Legionella" Microorganisms 11, no. 1: 74. https://doi.org/10.3390/microorganisms11010074