
Optimization of Submerged Culture Parameters of the Aphid Pathogenic Fungus Fusarium equiseti Based on Sporulation and Mycelial Biomass

Abstract
:1. Introduction
2. Materials and Methods
2.1. Organism, Inoculum Preparation, and Culture Conditions
2.2. Experimental Design
2.2.1. Preliminary Experiments: Screening of the Basic Medium
2.2.2. Single Factor Experimental Design
Incubation Time
Initial pH
Medium-to-Flask Ratio
Primary Inoculum Density
2.2.3. Assessment of Sporulation and Mycelial Production
2.2.4. Box–Behnken Design
2.2.5. Verification Test
2.3. Pathogenicity Test
2.4. Statistical Analysis
3. Results
3.1. Single-Factor Experiments
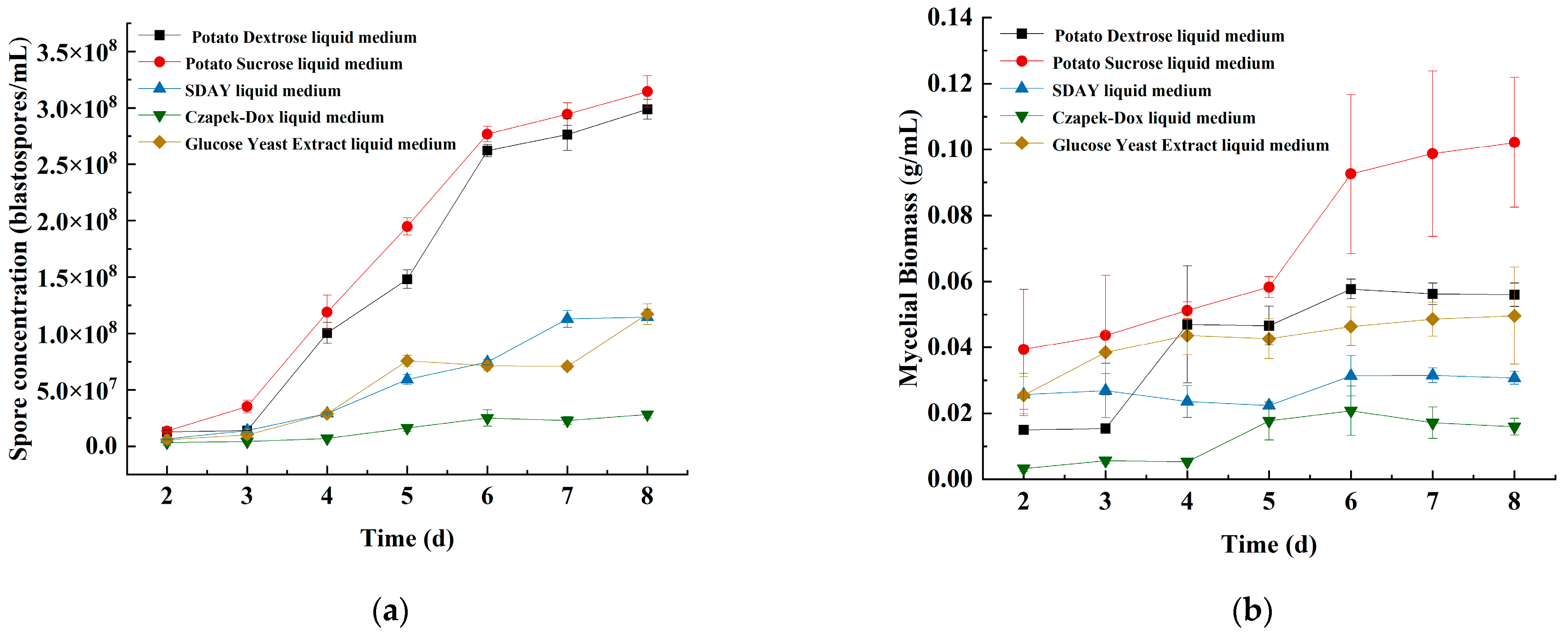
3.1.1. The Effects of the Basic Submerged Medium and Incubation Time on Spore Concentration and Mycelial Biomass
3.1.2. Effects of the Initial pH on Spore Concentration and Mycelial Biomass
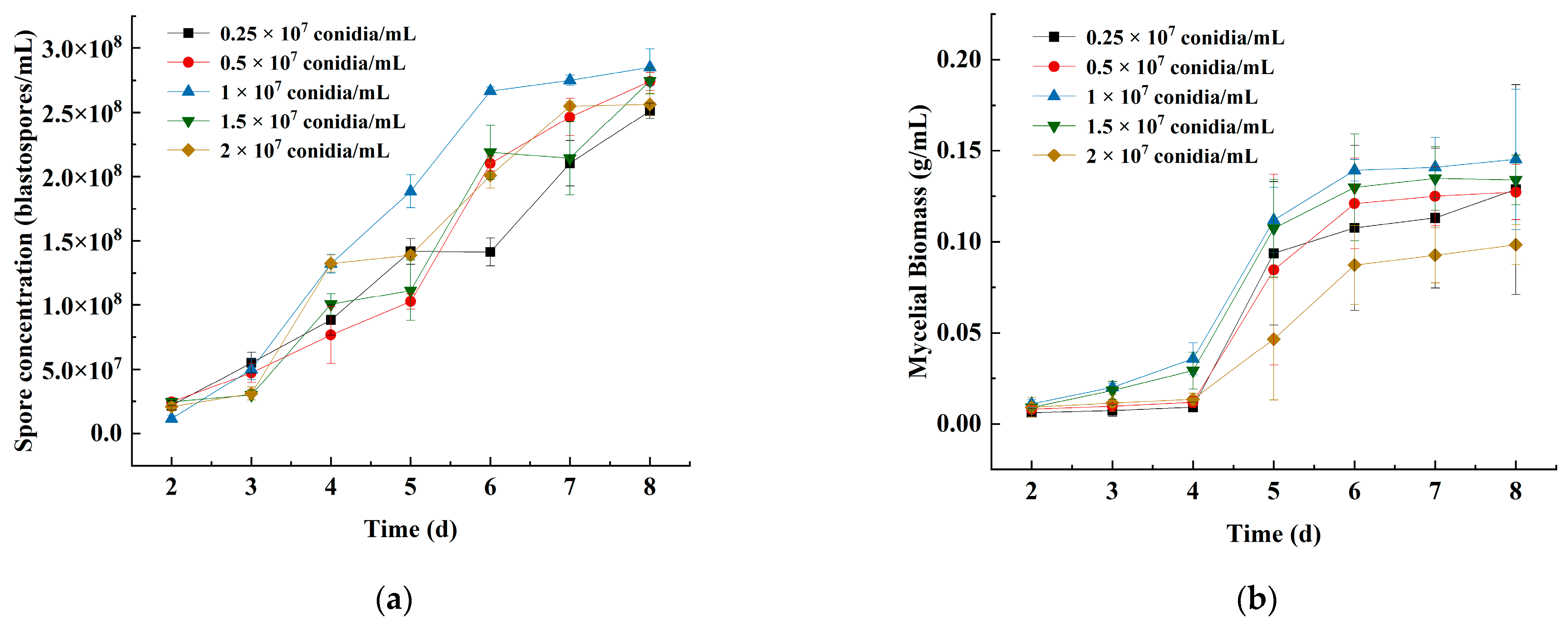
3.1.3. Effects of the Primary Inoculum Density on Spore Concentration and Mycelial Biomass
3.1.4. Effects of the Medium-to-Flask Ratio on Spore Concentration and Mycelial Biomass
3.2. Box–Behnken Design
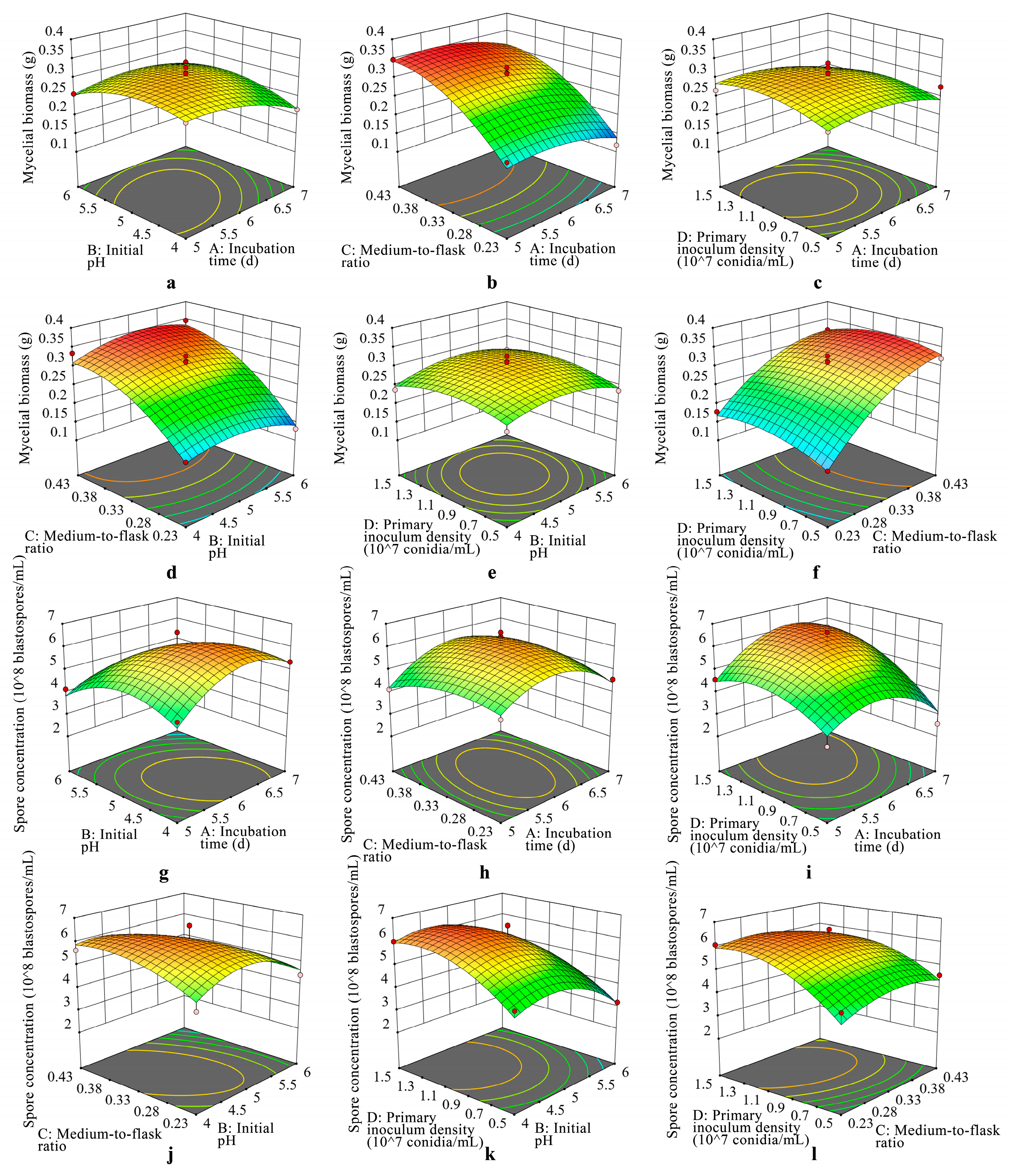
3.3. 3D Response Surfaces and 2D Contour Analysis
3.4. Verification Test
3.5. Pathogenicity Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hajek, A.E.; St Leger, R.J. Interactions Between Fungal Pathogens and Insect Hosts. Annu. Rev. Entomol. 1994, 39, 293–322. [Google Scholar] [CrossRef]
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic fungi: Unconventional applications. Rev. Environ. Sci. Bio/Technol. 2020, 19, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Topuz, E.; Erler, F.; Gumrukcu, E. Survey of indigenous entomopathogenic fungi and evaluation of their pathogenicity against the carmine spider mite, Tetranychus cinnabarinus (Boisd.), and the whitefly, Bemisia tabaci (Genn.) biotype B. Pest Manag. Sci. 2016, 72, 2273–2279. [Google Scholar] [CrossRef] [PubMed]
- Mascarin, G.M.; Lopes, R.B.; Delalibera, Í.; Fernandes, É.K.K.; Luz, C.; Faria, M. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Batta, Y. The first report on entomopathogenic effect of Fusarium avenaceum (Fries) Saccardo (Hypocreales, Ascomycota) against rice weevil (Sitophilus oryzae L.: Curculionidae, Coleoptera). J. Entomol. Acarol. Res. 2012, 44, 11. [Google Scholar] [CrossRef] [Green Version]
- Hami, A.; Rasool, R.S.; Khan, N.A.; Mansoor, S.; Mir, M.A.; Ahmed, N.; Masoodi, K.Z. Morpho-molecular identification and first report of Fusarium equiseti in causing chilli wilt from Kashmir (Northern Himalayas). Sci. Rep. 2021, 11, 3610. [Google Scholar] [CrossRef]
- Ahmad, F.; Anwar, W.; Javed, M.; Basit, R.; Akhter, A.; Ali, S.; Azhar, H.; Khan, A.; Amin, H.; Haider, M. Infection mechanism of Aspergillus and Fusarium species against Bemisia tabaci. Mycopath 2019, 17, 63–72. [Google Scholar]
- Soliman, N.A.; Al-amin, S.M.; Mesbah, A.E.; Ibrahim, A.M.A.; Mahmoud, A.M.A. Pathogenicity of three entomopathogenic fungi against the Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Egypt. J. Biol. Pest Control 2020, 30, 49. [Google Scholar] [CrossRef]
- Li, H. Study on Entomogenous Fungi—Fusarium. Hunan Agric. Sci. 1989, 36–37. [Google Scholar] [CrossRef]
- Teetor-Barsch, G.H.; Roberts, D.W. Entomogenous Fusarium species. Mycopathologia 1983, 84, 3–16. [Google Scholar] [CrossRef]
- da Silva Santos, A.C.; Diniz, A.G.; Tiago, P.V.; de Oliveira, N.T. Entomopathogenic Fusarium species: A review of their potential for the biological control of insects, implications and prospects. Fungal Biol. Rev. 2020, 34, 41–57. [Google Scholar] [CrossRef]
- Chehri, K. Molecular identification of entomopathogenic Fusarium species associated with Tribolium species in stored grains. J. Invertebr. Pathol. 2017, 144, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Anwar, W.; Haider, M.; Shahid, A.; Mushtaq, H.; Hameed, U.; Zia-ur-rehman, M.; Iqbal, M.j. Genetic diversity of Fusarium Isolated from Members of Sternorrhyncha (Hemiptera): Entomopathogens against Bemisia tabaci. Pak. J. Zool. 2017, 49, 639–645. [Google Scholar] [CrossRef]
- Addario, E.; Turchetti, T. Parasitic fungi on Dryocosmus kuriphilus in Castanea sativa necrotic galls. Bull. Insectology 2011, 64, 269–273. [Google Scholar]
- Liu, W.; Xie, Y.; Dong, J.; Xue, J.; Zhang, Y.; Lu, Y.; Wu, J. Pathogenicity of three entomopathogenic fungi to Matsucoccus matsumurae. PLoS One 2014, 9, e103350. [Google Scholar] [CrossRef]
- Carneiro-Leão, M.P.; Tiago, P.V.; Medeiros, L.V.; da Costa, A.F.; de Oliveira, N.T. Dactylopius opuntiae: Control by the Fusarium incarnatum–equiseti species complex and confirmation of mortality by DNA fingerprinting. J. Pest Sci. 2017, 90, 925–933. [Google Scholar] [CrossRef]
- Fan, J.-H.; Xie, Y.-P.; Xue, J.-L.; Xiong, Q.; Jiang, W.-J.; Zhang, Y.-J.; Ren, Z.-M. The strain HEB01 of Fusarium sp., a new pathogen that infects brown soft scale. Ann. Microbiol. 2014, 64, 333–341. [Google Scholar] [CrossRef]
- Sharma, A.; Chandla, V.K.; Thakur, D.R. Biodiversity and Pathogenicity Potential of Mycoflora Associated with Brahmina coriacea in Potato Fields of North-Western Indian Hills. J. Entomol. 2012, 9, 319–331. [Google Scholar] [CrossRef] [Green Version]
- da Silva Santos, A.C.; Oliveira, R.L.S.; da Costa, A.F.; Tiago, P.V.; de Oliveira, N.T. Controlling Dactylopius opuntiae with Fusarium incarnatum–equiseti species complex and extracts of Ricinus communis and Poincianella pyramidalis. J. Pest Sci. 2016, 89, 539–547. [Google Scholar] [CrossRef]
- Gupta, S.; Krasnoff, S.B.; Underwood, N.L.; Renwick, J.A.A.; Roberts, D.W. Isolation of beauvericin as an insect toxin from Fusarium semitectum and Fusarium moniliforme var. subglutinans. Mycopathologia 1991, 115, 185–189. [Google Scholar] [CrossRef]
- Logrieco, A.; Moretti, A.; Castella, G.; Kostecki, M.; Golinski, P.; Ritieni, A.; Chelkowski, J. Beauvericin Production by Fusarium Species. Appl. Environ. Microbiol. 1998, 64, 3084–3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitao, J.K.; Meyer, S.L.F.; Schmidt, W.F.; Fettinger, J.C.; Chitwood, D.J. Nematode-Antagonistic Trichothecenes from Fusarium equiseti. J. Chem. Ecol. 2001, 27, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Akbar, M.F.; Rana, H.; Perveen, F.K. Management of cauliflower aphid (Myzus persicae (Sulzer) Aphididae: Hemiptera) through environment friendly bioinsecticides. Pak. Entomol 2014, 36, 25–30. [Google Scholar]
- Chen, B.; Li, Z.Y.; Feng, M.G. Occurrence of entomopathogenic fungi in migratory alate aphids in Yunnan Province of China. BioControl 2008, 53, 317–326. [Google Scholar] [CrossRef]
- Pham, T.A.; Kim, J.J.; Kim, K. Optimization of Solid-State Fermentation for Improved Conidia Production of Beauveria bassiana as a Mycoinsecticide. Mycobiology 2010, 38, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.A. Optimizing nutritional conditions for the liquid culture production of effective fungal biological control agents. J. Ind. Microbiol. Biotechnol. 1997, 19, 180–187. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Kobori, N.N.; Jackson, M.A.; Dunlap, C.A.; Delalibera, I. Nitrogen sources affect productivity, desiccation tolerance and storage stability of Beauveria bassiana blastospores. J. Appl. Microbiol. 2018, 124, 810–820. [Google Scholar] [CrossRef]
- Puri, S.; Beg, Q.K.; Gupta, R. Optimization of Alkaline Protease Production from Bacillus sp. by Response Surface Methodology. Curr. Microbiol. 2002, 44, 286–290. [Google Scholar] [CrossRef]
- Kim, J.S.; Je, Y.H.; Skinner, M.; Parker, B.L. An oil-based formulation of Isaria fumosorosea blastospores for management of greenhouse whitefly Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Pest Manag. Sci. 2013, 69, 576–581. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Jackson, M.A.; Wood, B.W. Virulence of Hypocreales fungi to pecan aphids (Hemiptera: Aphididae) in the laboratory. J. Invertebr. Pathol. 2008, 99, 312–317. [Google Scholar] [CrossRef]
- Alkhaibari, A.M.; Carolino, A.T.; Yavaşoğlu, S.İ.; Maffeis, T.; Mattoso, T.C.; Bull, J.C.; Samuels, R.I.; Butt, T.M. Metarhizium brunneum Blastospore Pathogenesis in Aedes aegypti Larvae: Attack on Several Fronts Accelerates Mortality. PLoS Pathog. 2016, 12, e1005715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.B.; Yang, Z.; Yu, J.; Zhang, Y.; Xue, J.; Li, Z.; Li, J.-j.; Wang, C.-y.; Wang, Z.; Hou, J.; et al. Comparison between conidia and blastospores of Esteya vermicola, an endoparasitic fungus of the pinewood nematode, Bursaphelenchus xylophilus. World J. Microbiol. Biotechnol. 2013, 29, 2429–2436. [Google Scholar] [CrossRef] [PubMed]
- Alkhaibari, A.M.; Lord, A.M.; Maffeis, T.; Bull, J.C.; Olivares, F.L.; Samuels, R.I.; Butt, T.M. Highly specific host-pathogen interactions influence Metarhizium brunneum blastospore virulence against Culex quinquefasciatus larvae. Virulence 2018, 9, 1449–1467. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, P.; Hu, Y.; Zhao, G.; Liu, H.; Li, Z.; Wu, H.; Wang, L.; Zheng, Z. Optimised fermentation conditions and improved collection efficiency using dual cyclone equipment to enhance fungal conidia production. Biocontrol Sci. Technol. 2015, 25, 1011–1023. [Google Scholar] [CrossRef]
- Akbar, W.; Lord, J.C.; Nechols, J.R.; Howard, R.W. Diatomaceous Earth Increases the Efficacy of Beauveria bassiana Against Tribolium castaneum Larvae and Increases Conidia Attachment. J. Econ. Entomol. 2004, 97, 273–280. [Google Scholar] [CrossRef]
- Awan, U.A.; Meng, L.; Xia, S.; Raza, M.F.; Zhang, Z.; Zhang, H. Isolation, fermentation, and formulation of entomopathogenic fungi virulent against adults of Diaphorina citri. Pest Manag. Sci. 2021, 77, 4040–4053. [Google Scholar] [CrossRef]
- Li, S.; Myung, K.; Guse, D.; Donkin, B.; Proctor, R.H.; Grayburn, W.S.; Calvo, A.M. FvVE1 regulates filamentous growth, the ratio of microconidia to macroconidia and cell wall formation in Fusarium verticillioides. Mol. Microbiol. 2006, 62, 1418–1432. [Google Scholar] [CrossRef]
- Smith, S.N. An overview of ecological and habitat aspects in the genus Fusarium with special emphasis on the soil-borne pathogenic forms. Plant Pathol. Bull. 2007, 16, 97–120. [Google Scholar]
- van Wyk, P.S.; Wingfield, M.J.; Marasas, W.F.O.; Bosman, J.L. Development of microconidia in Fusarium section Sporotrichiella. Mycol. Res. 1991, 95, 284–289. [Google Scholar] [CrossRef]
- Qiu, J.; Song, F.; Qiu, Y.; Li, X.; Guan, X. Optimization of the medium composition of a biphasic production system for mycelial growth and spore production of Aschersonia placenta using response surface methodology. J. Invertebr. Pathol. 2013, 112, 108–115. [Google Scholar] [CrossRef]
- Patel, S.; Duggirala, S. Improved Cellulase Production through RSM by using Aspergillus tubingenesis MN239975 in Solid State Fermentation. Res. J. Biotechnol. 2020, 15, 1–10. [Google Scholar]
- Ahmad, S.; Pathak, V.V.; Kothari, R.; Kumar, A.; Naidu Krishna, S.B. Optimization of nutrient stress using C. pyrenoidosa for lipid and biodiesel production in integration with remediation in dairy industry wastewater using response surface methodology. 3 Biotech 2018, 8, 326. [Google Scholar] [CrossRef] [PubMed]
- Sawiphak, S.; Wongjiratthiti, A.; Saengprasan, C. Dioscorea alata as Alternative Culture Media for Fungal Cultivation and Biomass Production. Pertanika J. Trop. Agric. Sci. 2021, 44, 317–336. [Google Scholar] [CrossRef]
- Petlamul, W.; Boukaew, S. Optimisation and Stabilisation of Cellulase and Xylanase Production by Beauveria bassiana. EnvironmentAsia 2019, 12, 11–19. [Google Scholar]
- Amobonye, A.; Bhagwat, P.; Singh, S.; Pillai, S. Beauveria bassiana xylanase: Characterization and wastepaper deinking potential of a novel glycosyl hydrolase from an endophytic fungal entomopathogen. J. Fungi 2021, 7, 668. [Google Scholar] [CrossRef]
- Yin, L.; Chen, M.; Zeng, T.; Liu, X.; Zhu, F.; Huang, R. Improving probiotic spore yield using rice straw hydrolysate. Lett. Appl. Microbiol. 2021, 72, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lan, X.; Jia, R.; Hu, L.; Wang, Y. Response Surface Methodology (RSM) Mediated Optimization of Medium Components for Mycelial Growth and Metabolites Production of Streptomyces alfalfae XN-04. Microorganisms 2022, 10, 1854. [Google Scholar] [CrossRef]
- Vlajkov, V.; Andelic, S.; Pajcin, I.; Grahovac, M.; Budakov, D.; Jokic, A.; Grahovac, J. Medium for the Production of Bacillus-Based Biocontrol Agent Effective against Aflatoxigenic Aspergillus flavus: Dual Approach for Modelling and Optimization. Microorganisms 2022, 10, 1165. [Google Scholar] [CrossRef]
- Dong, C.; Xie, X.Q.; Wang, X.; Zhan, Y.; Yao, Y.J. Application of Box-Behnken design in optimisation for polysaccharides extraction from cultured mycelium of Cordyceps sinensis. Food Bioprod. Process. 2009, 87, 139–144. [Google Scholar] [CrossRef]
- Shih, L.; Tsai, K.-L.; Hsieh, C. Effects of culture conditions on the mycelial growth and bioactive metabolite production in submerged culture of Cordyceps militaris. Biochem. Eng. J. 2007, 33, 193–201. [Google Scholar] [CrossRef]
- Zhao, X.; Chai, J.; Zhang, X.; Hong, B.; Jia, Y. Identification of Fusarium equiseti and Its Pathogenicity to Myzus persicae. Chin. J. Biol. Control 2022, 1–12. [Google Scholar] [CrossRef]
- Amobonye, A.; Bhagwat, P.; Singh, S.; Pillai, S. Enhanced xylanase and endoglucanase production from Beauveria bassiana SAN01, an entomopathogenic fungal endophyte. Fungal Biol. 2021, 125, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Dhar, S.; Jindal, V.; Gupta, V.K. Optimization of growth conditions and medium composition for improved conidiation of newly isolated Beauveria bassiana strains. Indian J. Exp. Biol. 2016, 54, 634–643. [Google Scholar] [PubMed]
- Elhakim, E.; Mohamed, O.; Elazouni, I. Virulence and proteolytic activity of entomopathogenic fungi against the two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). Egypt. J. Biol. Pest Control 2020, 30, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hammami, A.; Bayoudh, A.; Hadrich, B.; Abdelhedi, O.; Jridi, M.; Nasri, M. Response-surface methodology for the production and the purification of a new H2O2-tolerant alkaline protease from Bacillus invictae AH1 strain. Biotechnol. Prog. 2020, 36, e2965. [Google Scholar] [CrossRef]
- Bezerra, M.A.; Santelli, R.E.; Oliveira, E.P.; Villar, L.S.; Escaleira, L.A. Response surface methodology (RSM) as a tool for optimization in analytical chemistry. Talanta 2008, 76, 965–977. [Google Scholar] [CrossRef]
- Pradeep, F.S.; Pradeep, B. Optimization of pigment and biomass production from Fusarium moniliforme under submerged fermentation conditions. culture 2013, 5, 10. [Google Scholar]
- Tang, P.; Zhang, Z.; Niu, L.; Gu, C.; Zheng, W.; Cui, H.-C.; Yuan, X.-H. Fusarium solani G6, a novel vitexin-producing endophytic fungus: Characterization, yield improvement and osteoblastic proliferation activity. Biotechnol. Lett. 2021, 43, 1371–1383. [Google Scholar] [CrossRef]
- Anellis, A.; Berkowitz, D.; Kemper, D.; Rowley, D. Production of types A and B spores of Clostridium botulinum by the biphasic method: Effect on spore population, radiation resistance, and toxigenicity. Appl. Microbiol. 1972, 23, 734–739. [Google Scholar] [CrossRef]
- Das, M.M.; Haridas, M.; Sabu, A. Process development for the enhanced production of bio-nematicide Purpureocillium lilacinum KU8 under solid-state fermentation. Bioresour. Technol. 2020, 308, 123328. [Google Scholar] [CrossRef]
- Arabi, M.I.E.; Bakri, Y.; Jawhar, M. Extracellular xylanase production by Fusarium species in solid state fermentation. Pol. J. Microbiol 2011, 60, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Khilare, V.; Ahmed, R. Effect of different media, pH and temperature on the growth of Fusarium oxysporum f. sp. ciceri causing chickpea wilt. Int. J. Adv. Biol. Res. 2012, 2, 99–102. [Google Scholar]
- Tarocco, F.; Lecuona, R.E.; Couto, A.S.; Arcas, J.A. Optimization of erythritol and glycerol accumulation in conidia of Beauveria bassiana by solid-state fermentation, using response surface methodology. Appl. Microbiol. Biotechnol. 2005, 68, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.; Wan, A.; Rahman, M.; Goswami, R.; Barasubiye, T.; Seifert, K.; Lévesque, C. Growth, population dynamics, and diversity of Fusarium equiseti in ginseng fields. Eur. J. Plant Pathol. 2008, 121, 173–184. [Google Scholar] [CrossRef]
- Mishra, S.; Malik, A. Comparative evaluation of five Beauveria isolates for housefly (Musca domestica L.) control and growth optimization of selected strain. Parasitol. Res. 2012, 111, 1937–1945. [Google Scholar] [CrossRef]
- Slininger, P.J.; Silman, R.W.; Jackson, M.A. Oxygen delivery requirements of Colletotrichum truncatum during germination, vegetative growth, and sporulation. Appl. Microbiol. Biotechnol. 1993, 39, 744–749. [Google Scholar] [CrossRef]
- Kumar, G.; Kunamneni, A.; Prabhakar, T.; Ellaiah, P. Optimization of Process Parameters for the Production of Inulinase from a Newly Isolated Aspergillus niger AUP19. World J. Microbiol. Biotechnol. 2005, 21, 1359–1361. [Google Scholar] [CrossRef]
- Niaz, M.; Iftikhar, T.; Qureshi, F.F.; Niaz, M. Extracellular lipase production by Aspergillus nidulans (MBL-S-6) under submerged fermentation. Int. J. Agric. Biol. 2014, 16, 536–542. [Google Scholar]
- Jiru, T.M.; Groenewald, M.; Pohl, C.; Steyn, L.; Kiggundu, N.; Abate, D. Optimization of cultivation conditions for biotechnological production of lipid by Rhodotorula kratochvilovae (syn, Rhodosporidium kratochvilovae) SY89 for biodiesel preparation. 3 Biotech 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Reihani, S.F.S.; Khosravi-Darani, K. Influencing factors on single-cell protein production by submerged fermentation: A review. Electron. J. Biotechnol. 2019, 37, 34–40. [Google Scholar] [CrossRef]
- Kamal, M.; Ali, M.; Shishir, M.R.I.; Saifullah, M.; Haque, M.; Mondal, S.C. Optimization of process parameters for improved production of biomass protein from Aspergillus niger using banana peel as a substrate. Food Sci. Biotechnol. 2019, 28, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Santa, H.S.D.; Santa, O.R.D.; Brand, D.; Vandenberghe, L.P.d.S.; Soccol, C.R. Spore production of Beauveria bassiana from agro-industrial residues. Braz. Arch. Biol. Technol. 2005, 48, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Alabdalall, A.H.; Alanazi, N.; AAldakeel, S.; Abdulazeez, S.; Borgio, J.F. Molecular, physiological, and biochemical characterization of extracellular lipase production by Aspergillus niger using submerged fermentation. PeerJ 2020, 8, e9425. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.S.; Sooch, B.S.; Puri, M. Optimization of medium and process parameters for the production of inulinase from a newly isolated Kluyveromyces marxianus YS-1. Bioresour. Technol. 2007, 98, 2518–2525. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Kaur, G. Optimization of different process variables for the production of an indolizidine alkaloid, swainsonine from Metarhizium anisopliae. J. Basic Microbiol. 2012, 52, 590–597. [Google Scholar] [CrossRef]
- Xu, C.P.; Sinha, J.; Bae, J.T.; Kim, S.W.; Yun, J.W. Optimization of physical parameters for exo–biopolymer production in submerged mycelial cultures of two entomopathogenic fungi Paecilomyces japonica and Paecilomyces tenuipes. Lett. Appl. Microbiol. 2006, 42, 501–506. [Google Scholar] [CrossRef]
- Santos, M.C.d.; Mendonça, M.d.L.; Bicas, J.L. Modeling bikaverin production by Fusarium oxysporum CCT7620 in shake flask cultures. Bioresour. Bioprocess. 2020, 7, 1–8. [Google Scholar] [CrossRef]
- Yujie, C.; Hongwei, Y.; Hailong, J. Optimal Conditions for Trichoderma longibrachiatum Strain T05 for Producing Conidia and Chlamydospores. J. Northeast For. Univ. 2016, 44, 110–113. [Google Scholar] [CrossRef]
- Srivastava, S.; Pathak, N.; Srivastava, P. Identification of limiting factors for the optimum growth of Fusarium oxysporum in liquid medium. Toxicol. Int. 2011, 18, 111. [Google Scholar] [CrossRef]
- Kim, H.; Yun, J. A comparative study on the production of exopolysaccharides between two entomopathogenic fungi Cordyceps militaris and Cordyceps sinensis in submerged mycelial cultures. J. Appl. Microbiol. 2005, 99, 728–738. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Name | Components Per 1 L Distilled Water |
|---|---|---|
| 1 | Potato dextrose liquid medium | 200 g peeled potato, 20 g glucose; |
| 2 | Potato sucrose liquid medium | 200 g peeled potato, 20 g sucrose; |
| 3 | Sabouraud Dextrose Agar with yeast extract liquid medium | 40 g glucose, 10 g peptone, 10 g yeast powder; |
| 4 | Czapek–Dox liquid medium | 30 g sucrose, 3 g NaNO3, 0.5 g KCl, 1 g K2HPO3, 0.01 g FeSO4, 0.5 g MgSO4.7H2O; |
| 5 | Glucose yeast extract liquid medium | 40 g glucose, 20 g yeast powder. |
| Factors | Levels | ||
|---|---|---|---|
| −1 | 0 | 1 | |
| A: Incubation time (d) | 5 | 6 | 7 |
| B: Initial pH | 4 | 5 | 6 |
| C: Medium-to-flask ratio | 0.23 | 0.33 | 0.43 |
| D: Primary inoculum density (107 conidia/mL) | 0.5 | 1 | 1.5 |
| Experimental Serial No. | Incubation Time (d) A | Initial pH B | Medium-to-Flask Ratio C | Primary Inoculum Density (107 Conidia/mL) D | Spore Concentration 108 (Blastospores/mL) | Mycelial Biomass (g/mL) |
|---|---|---|---|---|---|---|
| 1 | 6 | 4 | 0.33 | 0.5 | 4.55 | 0.24 |
| 2 | 6 | 6 | 0.43 | 1 | 3.41 | 0.35 |
| 3 | 6 | 5 | 0.33 | 1 | 6.01 | 0.31 |
| 4 | 7 | 5 | 0.23 | 1 | 4.58 | 0.12 |
| 5 | 7 | 6 | 0.33 | 1 | 3.22 | 0.26 |
| 6 | 7 | 4 | 0.33 | 1 | 5.35 | 0.22 |
| 7 | 6 | 5 | 0.33 | 1 | 5.84 | 0.28 |
| 8 | 6 | 5 | 0.33 | 1 | 5.98 | 0.29 |
| 9 | 6 | 5 | 0.33 | 1 | 5.86 | 0.31 |
| 10 | 6 | 4 | 0.33 | 1.5 | 6.01 | 0.24 |
| 11 | 6 | 5 | 0.43 | 1.5 | 5.27 | 0.32 |
| 12 | 5 | 4 | 0.33 | 1 | 4.50 | 0.29 |
| 13 | 6 | 5 | 0.23 | 1.5 | 6.06 | 0.18 |
| 14 | 6 | 4 | 0.43 | 1 | 5.63 | 0.34 |
| 15 | 5 | 6 | 0.33 | 1 | 4.14 | 0.26 |
| 16 | 6 | 6 | 0.23 | 1 | 4.41 | 0.13 |
| 17 | 6 | 5 | 0.43 | 0.5 | 4.55 | 0.32 |
| 18 | 6 | 6 | 0.33 | 0.5 | 3.18 | 0.24 |
| 19 | 6 | 5 | 0.23 | 0.5 | 4.38 | 0.14 |
| 20 | 7 | 5 | 0.33 | 1.5 | 5.58 | 0.26 |
| 21 | 5 | 5 | 0.33 | 1.5 | 4.58 | 0.27 |
| 22 | 6 | 6 | 0.33 | 1.5 | 4.31 | 0.26 |
| 23 | 5 | 5 | 0.33 | 0.5 | 3.52 | 0.26 |
| 24 | 5 | 5 | 0.43 | 1 | 4.13 | 0.35 |
| 25 | 7 | 5 | 0.43 | 1 | 5.13 | 0.28 |
| 26 | 6 | 5 | 0.33 | 1 | 6.63 | 0.33 |
| 27 | 5 | 5 | 0.23 | 1 | 4.60 | 0.19 |
| 28 | 6 | 4 | 0.23 | 1 | 4.52 | 0.16 |
| 29 | 7 | 5 | 0.33 | 0.5 | 2.57 | 0.28 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value | Significance |
|---|---|---|---|---|---|---|
| Model | 27.04 | 14 | 1.93 | 14.49 | <0.0001 | significant |
| A | 0.077 | 1 | 0.077 | 0.57 | 0.4611 | |
| B | 5.19 | 1 | 5.19 | 38.94 | <0.0001 | |
| C | 0.016 | 1 | 0.016 | 0.12 | 0.7370 | |
| D | 6.83 | 1 | 6.83 | 51.21 | <0.0001 | |
| AB | 0.79 | 1 | 0.79 | 5.91 | 0.0291 | |
| AC | 0.25 | 1 | 0.25 | 1.91 | 0.1889 | |
| AD | 0.96 | 1 | 0.96 | 7.19 | 0.0179 | |
| BC | 1.12 | 1 | 1.12 | 8.40 | 0.0117 | |
| BD | 0.028 | 1 | 0.028 | 0.21 | 0.6550 | |
| CD | 0.23 | 1 | 0.23 | 1.72 | 0.2105 | |
| A2 | 7.17 | 1 | 7.17 | 53.79 | <0.0001 | |
| B2 | 5.08 | 1 | 5.08 | 38.09 | <0.0001 | |
| C2 | 1.34 | 1 | 1.34 | 10.05 | 0.0068 | |
| D2 | 3.35 | 1 | 3.35 | 25.16 | 0.0002 | |
| Residual | 1.87 | 14 | 0.13 | |||
| Lack of Fit | 1.45 | 10 | 0.15 | 1.39 | 0.4028 | Not significant |
| Pure Error | 0.42 | 4 | 0.10 | |||
| Cor Total | 28.91 | 28 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value | Significance |
|---|---|---|---|---|---|---|
| Model | 0.11 | 14 | 0.0078 | 17.18 | <0.0001 | significant |
| A | 0.0037 | 1 | 0.0037 | 8.15 | 0.0127 | |
| B | 0 | 1 | 0 | 0.044 | 0.8374 | |
| C | 0.087 | 1 | 0.087 | 192.17 | <0.0001 | |
| D | 0.0002 | 1 | 0.002 | 0.35 | 0.5635 | |
| AB | 0.0012 | 1 | 0.0012 | 2.72 | 0.1211 | |
| AC | 0.0000034 | 1 | 0.0000034 | 0.0076 | 0.9319 | |
| AD | 0.0001 | 1 | 0.0001 | 0.31 | 0.5862 | |
| BC | 0.0005 | 1 | 0.0005 | 1.07 | 0.3175 | |
| BD | 0.0002 | 1 | 0.0002 | 0.34 | 0.5707 | |
| CD | 0.0004 | 1 | 0.0004 | 0.80 | 0.3868 | |
| A2 | 0.0032 | 1 | 0.0032 | 7.18 | 0.0179 | |
| B2 | 0.0053 | 1 | 0.0053 | 11.80 | 0.0040 | |
| C2 | 0.011 | 1 | 0.011 | 23.55 | 0.0003 | |
| D2 | 0.0041 | 1 | 0.0041 | 9.02 | 0.0095 | |
| Residual | 0.0063 | 14 | 0.0005 | |||
| Lack of Fit | 0.0051 | 10 | 0.0005 | 1.61 | 0.3428 | Not significant |
| Pure Error | 0.0013 | 4 | 0.0003 | |||
| Cor Total | 0.12 | 28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Chai, J.; Wang, F.; Jia, Y. Optimization of Submerged Culture Parameters of the Aphid Pathogenic Fungus Fusarium equiseti Based on Sporulation and Mycelial Biomass. Microorganisms 2023, 11, 190. https://doi.org/10.3390/microorganisms11010190
Zhao X, Chai J, Wang F, Jia Y. Optimization of Submerged Culture Parameters of the Aphid Pathogenic Fungus Fusarium equiseti Based on Sporulation and Mycelial Biomass. Microorganisms. 2023; 11(1):190. https://doi.org/10.3390/microorganisms11010190
Chicago/Turabian StyleZhao, Xueyi, Junfa Chai, Fang Wang, and Yanxia Jia. 2023. "Optimization of Submerged Culture Parameters of the Aphid Pathogenic Fungus Fusarium equiseti Based on Sporulation and Mycelial Biomass" Microorganisms 11, no. 1: 190. https://doi.org/10.3390/microorganisms11010190