
Lignocellulolytic Biocatalysts: The Main Players Involved in Multiple Biotechnological Processes for Biomass Valorization

Abstract
:1. Introduction
2. Microorganisms as Enzymes Sources
3. Lignocellulolytic Enzymes
3.1. Cellulases
3.2. Hemicellulases
3.3. Ligninases
3.4. Accessory Proteins and Enzymes
4. Lignocellulolytic Microorganisms
4.1. Archaea and Eubacteria
4.2. Fungi
4.3. Natural Habitat from Lignocellulolytic Microorganisms
5. Lignocellulolytic Enzymes Production
5.1. Methods for Enzymatic Production
5.2. Types of Biomasses Used for Enzymatic Production
6. Application of Lignocellulolytic Enzymes
6.1. Food and Beverage Industry
6.2. Textile Industry
6.3. Pulp and Paper Industry
6.4. Biofuels Production
6.5. Bioremediation
7. Recent Advances
7.1. Mixed Cultures
7.2. Genetic Engineering
7.3. Bioprospecting
8. Future Perspectives
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Feistner, A.T.C. Biodiversity: An Introduction, 2nd ed.; Gaston, K.J., Spicer, J.I., Eds.; Blackwell Publishing: Oxford, UK, 2004; Volume 38, pp. 465–465. [Google Scholar] [CrossRef] [Green Version]
- Mora, C.; Tittensor, D.P.; Adl, S.; Simpson, A.G.B.; Worm, B. How Many Species Are There on Earth and in the Ocean? PLoS Biol. 2011, 9, e1001127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodhi, N.S.; Ehrlich, P.R. Conservation Biology for All; Oxford University Press: Oxford, UK, 2010; ISBN 9780199554232. [Google Scholar]
- Steffen, W.; Broadgate, W.; Deutsch, L.; Gaffney, O.; Ludwig, C. The Trajectory of the Anthropocene: The Great Acceleration. Anthr. Rev. 2015, 2, 81–98. [Google Scholar] [CrossRef]
- Magurran, A.E. Biological Diversity. Curr. Biol. 2005, 15, R116–R118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magurran, A.E. Primer Measuring Biological Diversity. Curr. Biol. 2021, 31, R1141–R1224. [Google Scholar] [CrossRef] [PubMed]
- Muthusaravanan, S.; Sivarajasekar, N.; Vivek, J.S.; Paramasivan, T.; Naushad, M.; Prakashmaran, J.; Gayathri, V.; Al-Duaij, O.K. Phytoremediation of Heavy Metals: Mechanisms, Methods and Enhancements. Env. Chem Lett 2018, 16, 1339–1359. [Google Scholar] [CrossRef]
- Ram, R.M.; Keswani, C.; Bisen, K.; Tripathi, R.; Singh, S.P.; Singh, H.B. Biocontrol Technology. In Omics Technologies and Bio-Engineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 177–190. [Google Scholar]
- Danso, D.; Chow, J.; Streita, W.R. Plastics: Environmental and Biotechnological Perspectives on Microbial Degradation. Appl. Environ. Microbiol. 2019, 85, e01095-19. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Bhardwaj, R.; Jassal, S.; Goyal, T.; Khullar, A.; Gupta, N. Application of Enzymes for an Eco-Friendly Approach to Textile Processing. Environ. Sci. Pollut. Res. 2021. [Google Scholar] [CrossRef]
- Seid, A.; Andualem, B. The Role of Green Biotechnology through Genetic Engineering for Climate Change Mitigation and Adaptation, and for Food Security: Current Challenges and Future Perspectives. J. Adv. Biol. Biotechnol. 2021, 24, 1–11. [Google Scholar] [CrossRef]
- Dahman, Y.; Syed, K.; Begum, S.; Roy, P.; Mohtasebi, B. Biofuels. In Biomass, Biopolymer-Based Materials, and Bioenergy; Elsevier: Amsterdam, The Netherlands, 2019; pp. 277–325. [Google Scholar]
- Evans, G.M.; Furlong, J.C. Environmental Biotechnology; Wiley: Hoboken, NJ, USA, 2010; ISBN 9780470684184. [Google Scholar]
- Demain, A.L. Microbial Biotechnology. Trends Biotechnol. 2000, 18, 26–31. [Google Scholar] [CrossRef]
- Oliveira, V.M.; Andreote, F.D.; Cortelo, P.C.; Castro-Gamboa, I.; Costa-Lotufo, L.V.; Polizeli, M.L.T.M.; Thiemann, O.H.; Setubal, J.C. Microorganisms: The Secret Agents of the Biosphere, and Their Key Roles in Biotechnology. Biota Neotrop. 2022, 22, e20221343. [Google Scholar] [CrossRef]
- Ortíz, O.L.; Arango, S.M.R.; Devia, J.L.G. Aplicaciones Biotecnológicas de Los Microorganismos. Nova 2019, 17, 129–163. [Google Scholar] [CrossRef]
- Sanchez, S.; Demain, A.L. Useful Microbial Enzymes—An Introduction. In Biotechnology of Microbial Enzymes; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–11. [Google Scholar]
- Nealson, K.H.; Stahl, D.A. Microorganisms and biogeochemical cycles: What can we learn from layered microbial communities? In Geomicrobiology; De Gruyter: Berlin, Germany, 1997; pp. 5–34. [Google Scholar] [CrossRef]
- Louca, S.; Mazel, F.; Doebeli, M.; Parfrey, L.W. A Census-Based Estimate of Earth’s Bacterial and Archaeal Diversity. PLoS Biol. 2019, 17, e3000106. [Google Scholar] [CrossRef] [Green Version]
- Baldrian, P.; Větrovský, T.; Lepinay, C.; Kohout, P. High-Throughput Sequencing View on the Magnitude of Global Fungal Diversity. Fungal Divers 2022, 114, 539–547. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Wichern, F. Alive and Kicking: Why Dormant Soil Microorganisms Matter. Soil Biol. Biochem. 2018, 116, 419–430. [Google Scholar] [CrossRef]
- Margesin, R.; Collins, T. Microbial Ecology of the Cryosphere (Glacial and Permafrost Habitats): Current Knowledge. Appl. Microbiol. Biotechnol. 2019, 103, 2537–2549. [Google Scholar] [CrossRef] [Green Version]
- Novak Babič, M.; Gostinčar, C.; Gunde-Cimerman, N. Microorganisms Populating the Water-Related Indoor Biome. Appl. Microbiol. Biotechnol. 2020, 104, 6443–6462. [Google Scholar] [CrossRef]
- Nair, B.M.; Prajapati, J.B. The History of Fermented Foods; Farnworth, E.R., Ed.; CRC Press: Boca Raton, FL, USA, 2003; ISBN 9780429214547. [Google Scholar]
- Nath, A.K.; Gupta, A.; Neopany, B.; Vyas, G.; Maneesri, J.; Thakur, N.; Sharma, N.; Achi, O.K.; Lakhanpal, P.; Schillinger, U. Biotechnology and Traditional Fermented Foods. In Indigenous Fermented Foods of South Asia; Joshi, V.K., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 773–828. ISBN 9780429067402. [Google Scholar]
- Singh, R. Microbial Biotechnology: A Promising Implement for Sustainable Agriculture. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 107–114. [Google Scholar]
- Timmis, K.; de Vos, W.M.; Ramos, J.L.; Vlaeminck, S.E.; Prieto, A.; Danchin, A.; Verstraete, W.; de Lorenzo, V.; Lee, S.Y.; Brüssow, H.; et al. The Contribution of Microbial Biotechnology to Sustainable Development Goals. Microb. Biotechnol. 2017, 10, 984–987. [Google Scholar] [CrossRef]
- Choi, J.-M.; Han, S.-S.; Kim, H.-S. Industrial Applications of Enzyme Biocatalysis: Current Status and Future Aspects. Biotechnol. Adv. 2015, 33, 1443–1454. [Google Scholar] [CrossRef]
- Dewan, S.S. Global Markets for Enzymes in Industrial Applications; BCC Research: Boston, MA, USA, 2021. [Google Scholar]
- Pagliaro, M. An Industry in Transition: The Chemical Industry and the Megatrends Driving Its Forthcoming Transformation. Angew. Chem. Int. Ed. 2019, 58, 11154–11159. [Google Scholar] [CrossRef]
- Gavrilescu, M.; Chisti, Y. Biotechnology—A Sustainable Alternative for Chemical Industry. Biotechnol. Adv. 2005, 23, 471–499. [Google Scholar] [CrossRef]
- Aldridge, S. Industry Backs Biocatalysis for Greener Manufacturing. Nat. Biotechnol. 2013, 31, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial Enzymes: Industrial Progress in 21st Century. 3 Biotech 2016, 6, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.S.; Singh, T.; Pandey, A. Microbial Enzymes—An Overview. In Advances in Enzyme Technology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–40. [Google Scholar]
- Li, S.; Yang, X.; Yang, S.; Zhu, M.; Wang, X. Technology prospecting on enzymes: Application, marketing and engineering. Comput. Struct. Biotechnol. J. 2012, 2, e201209017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of Microbial Enzymes in Food Industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Abada, E.A. Application of Microbial Enzymes in the Dairy Industry. In Enzymes in Food Biotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 61–72. [Google Scholar]
- Du, J.; Li, L.; Zhou, S. Microbial Production of Cyanophycin: From Enzymes to Biopolymers. Biotechnol. Adv. 2019, 37, 107400. [Google Scholar] [CrossRef]
- Sharma, B.; Dangi, A.K.; Shukla, P. Contemporary Enzyme Based Technologies for Bioremediation: A Review. J. Env. Manag. 2018, 210, 10–22. [Google Scholar] [CrossRef]
- Bhandari, S.; Poudel, D.K.; Marahatha, R.; Dawadi, S.; Khadayat, K.; Phuyal, S.; Shrestha, S.; Gaire, S.; Basnet, K.; Khadka, U.; et al. Microbial Enzymes Used in Bioremediation. J. Chem. 2021, 2021, 8849512. [Google Scholar] [CrossRef]
- Haripriyan, U.; Gopinath, K.P.; Arun, J.; Govarthanan, M. Bioremediation of Organic Pollutants: A Mini Review on Current and Critical Strategies for Wastewater Treatment. Arch. Microbiol. 2022, 204, 286. [Google Scholar] [CrossRef]
- Polizeli, M.L.T.M.; Rizzatti, A.C.S.; Monti, R.; Terenzi, H.F.; Jorge, J.A.; Amorim, D.S. Xylanases from Fungi: Properties and Industrial Applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef]
- Guimarães, L.H.S.; Peixoto-Nogueira, S.C.; Michelin, M.; Rizzatti, A.C.S.; Sandrim, V.C.; Zanoelo, F.F.; Aquino, A.C.M.M.; Junior, A.B.; Polizeli, M.L.T.M. Screening of Filamentous Fungi for Production of Enzymes of Biotechnological Interest. Braz. J. Microbiol. 2006, 37, 474–480. [Google Scholar] [CrossRef]
- Saldarriaga-Hernández, S.; Velasco-Ayala, C.; Leal-Isla Flores, P.; de Jesús Rostro-Alanis, M.; Parra-Saldivar, R.; Iqbal, H.M.N.; Carrillo-Nieves, D. Biotransformation of Lignocellulosic Biomass into Industrially Relevant Products with the Aid of Fungi-Derived Lignocellulolytic Enzymes. Int. J. Biol. Macromol. 2020, 161, 1099–1116. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Mishra, J.; Arora, N.; Mishra, P.; Li, H.; O′Hair, J.; Bhatti, S.; Zhou, S. Microbial Cellulolytic Enzymes: Diversity and Biotechnology with Reference to Lignocellulosic Biomass Degradation. Rev. Environ. Sci. Bio/Technol. 2020, 19, 621–648. [Google Scholar] [CrossRef]
- Kumar, A.; Chandra, R. Ligninolytic Enzymes and Its Mechanisms for Degradation of Lignocellulosic Waste in Environment. Heliyon 2020, 6, e03170. [Google Scholar] [CrossRef]
- Chukwuma, O.B.; Rafatullah, M.; Tajarudin, H.A.; Ismail, N. Lignocellulolytic Enzymes in Biotechnological and Industrial Processes: A Review. Sustainability 2020, 12, 7282. [Google Scholar] [CrossRef]
- Ejaz, U.; Sohail, M.; Ghanemi, A. Cellulases: From Bioactivity to a Variety of Industrial Applications. Biomimetics 2021, 6, 44. [Google Scholar] [CrossRef]
- Okeke, E.S.; Ezugwu, A.L.; Anaduaka, E.G.; Mayel, M.H.; Ezike, T.C.; Ossai, E.C. Ligninolytic and Cellulolytic Enzymes—Biocatalysts for Green Agenda. Biomass Convers. Biorefin. 2022. [Google Scholar] [CrossRef]
- Zoghlami, A.; Paës, G. Lignocellulosic Biomass: Understanding Recalcitrance and Predicting Hydrolysis. Front. Chem. 2019, 7, 874. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, N.; Mishra, K.; Srivastava, M.; Srivastava, K.R.; Gupta, V.K.; Ramteke, P.W.; Mishra, P.K. Role of Compositional Analysis of Lignocellulosic Biomass for Efficient Biofuel Production. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 29–43. [Google Scholar]
- Read, S.M.; Bacic, T. Prime Time for Cellulose. Science 2002, 295, 59–60. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, L.; Sun, Y.; Mitchell, G.; Liu, C. Advance of Studies on Structure and Decrystallization of Cellulose. Chem. Ind. For. Prod. 2008, 28, 109–114. [Google Scholar]
- Suhas; Gupta, V.K.; Carrott, P.J.M.; Singh, R.; Chaudhary, M.; Kushwaha, S. Cellulose: A Review as Natural, Modified and Activated Carbon Adsorbent. Bioresour. Technol. 2016, 216, 1066–1076. [Google Scholar] [CrossRef]
- Vassilev, S.V.; Baxter, D.; Andersen, L.K.; Vassileva, C.G.; Morgan, T.J. An Overview of the Organic and Inorganic Phase Composition of Biomass. Fuel 2012, 94, 1–33. [Google Scholar] [CrossRef]
- Pasin, T.M.; de Almeida, P.Z.; de Almeida Scarcella, A.S.; da Conceição Infante, J.; Polizeli, M.L.T.M. Bioconversion of Agro-Industrial Residues to Second-Generation Bioethanol. In Biorefinery of Alternative Resources: Targeting Green Fuels and Platform Chemicals; Springer: Singapore, 2020; pp. 23–47. [Google Scholar]
- Saha, B.C. Hemicellulose Bioconversion. J. Ind. Microbiol. Biotechnol. 2003, 30, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, Z.; Li, X.; Liu, X.; Fan, J.; Clark, J.H.; Hu, C. The Production of Furfural Directly from Hemicellulose in Lignocellulosic Biomass: A Review. Catal. Today 2019, 319, 14–24. [Google Scholar] [CrossRef]
- Gírio, F.M.; Fonseca, C.; Carvalheiro, F.; Duarte, L.C.; Marques, S.; Bogel-Łukasik, R. Hemicelluloses for Fuel Ethanol: A Review. Bioresour. Technol. 2010, 101, 4775–4800. [Google Scholar] [CrossRef]
- Polizeli, M.L.T.M.; Somera, A.F.; de Lucas, R.C.; Nozawa, M.S.F.; Michelin, M. Enzymes Involved in the Biodegradation of Sugarcane Biomass: Challenges and Perspectives. In Advances of Basic Science for Second Generation Bioethanol from Sugarcane; Springer International Publishing: Cham, Switzerland, 2017; pp. 55–79. [Google Scholar]
- Qaseem, M.F.; Shaheen, H.; Wu, A.-M. Cell Wall Hemicellulose for Sustainable Industrial Utilization. Renew. Sustain. Energy Rev. 2021, 144, 110996. [Google Scholar] [CrossRef]
- Rennie, E.A.; Scheller, H.V. Xylan Biosynthesis. Curr. Opin. Biotechnol. 2014, 26, 100–107. [Google Scholar] [CrossRef]
- Polizeli, M.L.T.M.; Vici, A.C.; Scarcella, A.S.A.; Cereia, M.; Pereira, M.G. Enzyme System from Aspergillus in Current Industrial Uses and Future Applications in the Production of Second-Generation Ethanol. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2016; pp. 127–140. [Google Scholar]
- Dimarogona, M.; Topakas, E.; Christakopoulos, P. Cellulose degradation by oxidative enzymes. Comput. Struct. Biotechnol. J. 2012, 2, e201209015. [Google Scholar] [CrossRef] [Green Version]
- Katahira, R.; Elder, T.J.; Beckham, G.T. A Brief Introduction to Lignin Structure. In Lignin Valorization: Emerging Approaches; The Royal Society of Chemistry: Cambridge, UK, 2018; Chapter 1; pp. 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ponnusamy, V.K.; Nguyen, D.D.; Dharmaraja, J.; Shobana, S.; Banu, J.R.; Saratale, R.G.; Chang, S.W.; Kumar, G. A Review on Lignin Structure, Pretreatments, Fermentation Reactions and Biorefinery Potential. Bioresour. Technol. 2019, 271, 462–472. [Google Scholar] [CrossRef]
- Himmel, M.E.; Ding, S.-Y.; Johnson, D.K.; Adney, W.S.; Nimlos, M.R.; Brady, J.W.; Foust, T.D. Biomass Recalcitrance: Engineering Plants and Enzymes for Biofuels Production. Science 2007, 315, 804–807. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Pu, Y.; Ragauskas, A.J. Current Understanding of the Correlation of Lignin Structure with Biomass Recalcitrance. Front. Chem. 2016, 4, 45. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.M.; Mota, T.R.; Grandis, A.; de Morais, G.R.; de Lucas, R.C.; Polizeli, M.L.T.M.; Marchiosi, R.; Buckeridge, M.S.; Ferrarese-Filho, O.; dos Santos, W.D. Lignin Plays a Key Role in Determining Biomass Recalcitrance in Forage Grasses. Renew. Energy 2020, 147, 2206–2217. [Google Scholar] [CrossRef]
- Sharma, S.; Yazdani, S.S. Diversity of Microbial Cellulase System. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2016; pp. 49–64. [Google Scholar]
- Leo, V.V.; Zothanpuia; Lalthafala; Ramesh, N.; Singh, B.P. Microorganisms as an Efficient Tool for Cellulase Production: Availability, Diversity, and Efficiency. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 45–61. [Google Scholar]
- Rabinovich, M.L.; Melnick, M.S.; Bolobova, A.V. The Structure and Mechanism of Action of Cellulolytic Enzymes. Biochemistry 2002, 67, 850–871. [Google Scholar] [CrossRef]
- Sharma, A.; Tewari, R.; Rana, S.S.; Soni, R.; Soni, S.K. Cellulases: Classification, Methods of Determination and Industrial Applications. Appl. Biochem. Biotechnol. 2016, 179, 1346–1380. [Google Scholar] [CrossRef]
- Horn, S.J.; Vaaje-Kolstad, G.; Westereng, B.; Eijsink, V. Novel Enzymes for the Degradation of Cellulose. Biotechnol. Biofuels 2012, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Kostylev, M.; Wilson, D. Synergistic Interactions in Cellulose Hydrolysis. Biofuels 2012, 3, 61–70. [Google Scholar] [CrossRef]
- Sharma, A.; Choudhary, J.; Singh, S.; Singh, B.; Kuhad, R.C.; Kumar, A.; Nain, L. Cellulose as Potential Feedstock for Cellulase Enzyme Production: Versatility and Properties of Various Cellulosic Biomasses. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 11–27. [Google Scholar]
- Kuhad, R.C.; Deswal, D.; Sharma, S.; Bhattacharya, A.; Jain, K.K.; Kaur, A.; Pletschke, B.I.; Singh, A.; Karp, M. Revisiting Cellulase Production and Redefining Current Strategies Based on Major Challenges. Renew. Sustain. Energy Rev. 2016, 55, 249–272. [Google Scholar] [CrossRef]
- Bayer, E.A.; Belaich, J.-P.; Shoham, Y.; Lamed, R. The Cellulosomes: Multienzyme Machines for Degradation of Plant Cell Wall Polysaccharides. Annu. Rev. Microbiol. 2004, 58, 521–554. [Google Scholar] [CrossRef]
- Alves, V.D.; Fontes, C.M.G.A.; Bule, P. Cellulosomes: Highly Efficient Cellulolytic Complexes. In Macromolecular Protein Complexes III: Structure and Function; Europe PMC: Hinxtone, UK, 2021; pp. 323–354. [Google Scholar] [CrossRef]
- Yamamoto, K.; Tamaru, Y. Important Roles of the Cellulosome on Degradation of Plant Biomass. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2016; pp. 3–8. [Google Scholar]
- Bayer, E.A.; Setter, E.; Lamed, R. Organization and Distribution of the Cellulosome in Clostridium thermocellum. J. Bacteriol. 1985, 163, 552–559. [Google Scholar] [CrossRef] [Green Version]
- Lamed, R.; Bayer, E.A. The Cellulosome of Clostridium thermocellum. Adv. Appl. Microbiol. 1988, 33, 1–46. [Google Scholar] [CrossRef]
- Maki, M.; Leung, K.T.; Qin, W. The Prospects of Cellulase-Producing Bacteria for the Bioconversion of Lignocellulosic Biomass. Int. J. Biol. Sci. 2009, 5, 500–516. [Google Scholar] [CrossRef]
- Artzi, L.; Bayer, E.A.; Moraïs, S. Cellulosomes: Bacterial Nanomachines for Dismantling Plant Polysaccharides. Nat. Rev. Microbiol. 2017, 15, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.Z.; Timmers, L.F.S.M.; Granada, C.E. Cellulase Production by Bacteria Is a Strain-Specific Characteristic with a High Biotechnological Potential. A Review of Cellulosome of Highly Studied Strains. Cellulose 2022, 29, 8065–8083. [Google Scholar] [CrossRef]
- Mukkala, S.; Bramhachari, P.V.; Reddy, Y.H.K. The Cellulosome: A Fiber-Degrading Strategist of the Rumen Microbiome. In Understanding the Microbiome Interactions in Agriculture and the Environment; Springer Nature Singapore: Singapore, 2022; pp. 215–241. [Google Scholar]
- Koeck, D.E.; Pechtl, A.; Zverlov, V.V.; Schwarz, W.H. Genomics of Cellulolytic Bacteria. Curr. Opin. Biotechnol. 2014, 29, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Moraïs, S.; Stern, J.; Kahn, A.; Galanopoulou, A.P.; Yoav, S.; Shamshoum, M.; Smith, M.A.; Hatzinikolaou, D.G.; Arnold, F.H.; Bayer, E.A. Enhancement of Cellulosome-Mediated Deconstruction of Cellulose by Improving Enzyme Thermostability. Biotechnol. Biofuels 2016, 9, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houfani, A.A.; Anders, N.; Spiess, A.C.; Baldrian, P.; Benallaoua, S. Insights from Enzymatic Degradation of Cellulose and Hemicellulose to Fermentable Sugars—A Review. Biomass Bioenergy 2020, 134, 105481. [Google Scholar] [CrossRef]
- Shallom, D.; Shoham, Y. Microbial Hemicellulases. Curr. Opin. Microbiol. 2003, 6, 219–228. [Google Scholar] [CrossRef]
- Brigham, J.S.; Adney, W.S.; Himmel, M.E. Hemicellulases: Diversity and Applications. In Handbook on Bioethanol; Routledge: Abingdon-on-Thames, UK; Taylor & Francis Group: London, UK, 2018; pp. 119–141. [Google Scholar] [CrossRef]
- Rodrigues, A.G. Endo-β-1,4-Xylanase: An Overview of Recent Developments; Gupata, V.K., Ed.; Springer Science+Business Media, LLC: New York, NY, USA, 2016; pp. 125–149. [Google Scholar] [CrossRef]
- Jordan, D.B.; Wagschal, K. Properties and Applications of Microbial β-D-Xylosidases Featuring the Catalytically Efficient Enzyme from Selenomonas ruminantium. Appl. Microbiol. Biotechnol. 2010, 86, 1647–1658. [Google Scholar] [CrossRef]
- Jin, X.; Song, J.; Ma, J.; Liu, G.-Q. Thermostable β-Xylosidase from Aspergillus fumigatus: Purification, Characterization and Potential Application in Lignocellulose Bioethanol Production. Renew. Energy 2020, 155, 1425–1431. [Google Scholar] [CrossRef]
- Chong, S.-L.; Derba-Maceluch, M.; Koutaniemi, S.; Gómez, L.D.; McQueen-Mason, S.J.; Tenkanen, M.; Mellerowicz, E.J. Active Fungal GH115 α-Glucuronidase Produced in Arabidopsis thaliana Affects Only the UX1-Reactive Glucuronate Decorations on Native Glucuronoxylans. BMC Biotechnol. 2015, 15, 56. [Google Scholar] [CrossRef] [Green Version]
- Contesini, F.J.; Liberato, M.V.; Rubio, M.V.; Calzado, F.; Zubieta, M.P.; Riaño-Pachón, D.M.; Squina, F.M.; Bracht, F.; Skaf, M.S.; Damasio, A.R. Structural and Functional Characterization of a Highly Secreted α-l-Arabinofuranosidase (GH62) from Aspergillus nidulans Grown on Sugarcane Bagasse. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2017, 1865, 1758–1769. [Google Scholar] [CrossRef]
- Dodd, D.; Cann, I.K.O. Enzymatic Deconstruction of Xylan for Biofuel Production. GCB Bioenergy 2009, 1, 2–17. [Google Scholar] [CrossRef] [Green Version]
- Mandal, A. Review on Microbial Xylanases and Their Applications. Int. J. Life Sci. 2015, 4, 178–187. [Google Scholar]
- Lee, S.H.; Lim, V.; Lee, C.K. Newly Isolate Highly Potential Xylanase Producer Strain from Various Environmental Sources. Biocatal. Agric. Biotechnol. 2018, 16, 669–676. [Google Scholar] [CrossRef]
- Qeshmi, F.I.; Homaei, A.; Fernandes, P.; Hemmati, R.; Dijkstra, B.W.; Khajeh, K. Xylanases from Marine Microorganisms: A Brief Overview on Scope, Sources, Features and Potential Applications. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2020, 1868, 140312. [Google Scholar] [CrossRef]
- Denisenko, Y.A.; Gusakov, A.V.; Rozhkova, A.M.; Zorov, I.N.; Bashirova, A.V.; Matys, V.Y.; Nemashkalov, V.A.; Sinitsyn, A.P. Protein Engineering of GH10 Family Xylanases for Gaining a Resistance to Cereal Proteinaceous Inhibitors. Biocatal. Agric. Biotechnol. 2019, 17, 690–695. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Insight into Microbial Hemicellulases Other than Xylanases: A Review. J. Chem. Technol. Biotechnol. 2013, 88, 353–363. [Google Scholar] [CrossRef]
- Abdel-Hamid, A.M.; Solbiati, J.O.; Cann, I.K.O. Insights into Lignin Degradation and Its Potential Industrial Applications. Adv. Appl. Microbiol. 2013, 82, 1–28. [Google Scholar] [CrossRef]
- Plácido, J.; Capareda, S. Ligninolytic Enzymes: A Biotechnological Alternative for Bioethanol Production. Bioresour. Bioprocess. 2015, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Pollegioni, L.; Tonin, F.; Rosini, E. Lignin-Degrading Enzymes. FEBS J. 2015, 282, 1190–1213. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkołazka, A.; Paszczyński, A. Lignin Degradation: Microorganisms, Enzymes Involved, Genomes Analysis and Evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [Green Version]
- Wongwilaiwalin, S.; Rattanachomsri, U.; Laothanachareon, T.; Eurwilaichitr, L.; Igarashi, Y.; Champreda, V. Analysis of a Thermophilic Lignocellulose Degrading Microbial Consortium and Multi-Species Lignocellulolytic Enzyme System. Enzym. Microb. Technol. 2010, 47, 283–290. [Google Scholar] [CrossRef]
- Claus, H. Laccases: Structure, Reactions, Distribution. Micron 2004, 35, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, U.N.; Singh, P.; Pandey, V.P.; Kumar, A. Structure–Function Relationship among Bacterial, Fungal and Plant Laccases. J. Mol. Catal. B Enzym. 2011, 68, 117–128. [Google Scholar] [CrossRef]
- Guimarães, L.R.C.; Woiciechowski, A.L.; Karp, S.G.; Coral, J.D.; Zandoná Filho, A.; Soccol, C.R. Laccases. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2017; pp. 199–216. [Google Scholar]
- de França Passos, D.; Pereir, N., Jr.; de Castro, A.M. A Comparative Review of Recent Advances in Cellulases Production by Aspergillus, Penicillium and Trichoderma Strains and Their Use for Lignocellulose Deconstruction. Curr. Opin. Green Sustain. Chem. 2018, 14, 60–66. [Google Scholar] [CrossRef]
- Ezeilo, U.R.; Zakaria, I.I.; Huyop, F.; Wahab, R.A. Enzymatic Breakdown of Lignocellulosic Biomass: The Role of Glycosyl Hydrolases and Lytic Polysaccharide Monooxygenases. Biotechnol. Biotechnol. Equip. 2017, 31, 647–662. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.J.; Lee, H.J.; Choi, I.-G.; Kim, K.H. Synergistic Proteins for the Enhanced Enzymatic Hydrolysis of Cellulose by Cellulase. Appl. Microbiol. Biotechnol. 2014, 98, 8469–8480. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Microbial Expansins. Annu. Rev. Microbiol. 2017, 71, 479–497. [Google Scholar] [CrossRef]
- Agger, J.W.; Isaksen, T.; Várnai, A.; Vidal-Melgosa, S.; Willats, W.G.T.; Ludwig, R.; Horn, S.J.; Eijsink, V.G.H.; Westereng, B. Discovery of LPMO Activity on Hemicelluloses Shows the Importance of Oxidative Processes in Plant Cell Wall Degradation. Proc. Natl. Acad. Sci. USA 2014, 111, 6287–6292. [Google Scholar] [CrossRef] [Green Version]
- Suleiman, M.; Krüger, A.; Antranikian, G. Biomass-Degrading Glycoside Hydrolases of Archaeal Origin. Biotechnol. Biofuels 2020, 13, 153. [Google Scholar] [CrossRef]
- Kim, H.-W.; Ishikawa, K. Structure of Hyperthermophilic Endocellulase from Pyrococcus Horikoshii. Proteins Struct. Funct. Bioinform. 2010, 78, 496–500. [Google Scholar] [CrossRef]
- Graham, J.E.; Clark, M.E.; Nadler, D.C.; Huffer, S.; Chokhawala, H.A.; Rowland, S.E.; Blanch, H.W.; Clark, D.S.; Robb, F.T. Identification and Characterization of a Multidomain Hyperthermophilic Cellulase from an Archaeal Enrichment. Nat. Commun. 2011, 2, 375. [Google Scholar] [CrossRef] [Green Version]
- Boyce, A.; Walsh, G. Expression and Characterisation of a Thermophilic Endo-1,4-β-Glucanase from Sulfolobus shibatae of Potential Industrial Application. Mol. Biol Rep. 2018, 45, 2201–2211. [Google Scholar] [CrossRef]
- Sakai, H.D.; Kurosawa, N. Saccharolobus caldissimus Gen. Nov., sp. Nov., a Facultatively Anaerobic Iron-Reducing Hyperthermophilic Archaeon Isolated from an Acidic Terrestrial Hot Spring, and Reclassification of Sulfolobus solfataricus as Saccharolobus solfataricus Comb. Nov. and Sulfolobus shibatae as Saccharolobus shibatae Comb. Nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 1271–1278. [Google Scholar] [CrossRef]
- Jones, J.A.D.; Kerr, R.G.; Haltli, B.A.; Tinto, W.F. Temperature and PH Effect on Glucose Production from Pretreated Bagasse by a Novel Species of Citrobacter and Other Bacteria. Heliyon 2018, 4, e00657. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Algora, C.; Baldrian, P. Lignocellulolytic Systems of Soil Bacteria: A Vast and Diverse Toolbox for Biotechnological Conversion Processes. Biotechnol. Adv. 2019, 37, 107374. [Google Scholar] [CrossRef]
- Chukwuma, O.B.; Rafatullah, M.; Tajarudin, H.A.; Ismail, N. A Review on Bacterial Contribution to Lignocellulose Breakdown into Useful Bio-Products. Int J. Environ. Res. Public Health 2021, 18, 6001. [Google Scholar] [CrossRef]
- Sadhu, S. Cellulase Production by Bacteria: A Review. Br. Microbiol. Res. J. 2013, 3, 235–258. [Google Scholar] [CrossRef] [Green Version]
- Saini, A.; Aggarwal, N.K.; Sharma, A.; Yadav, A. Actinomycetes: A Source of Lignocellulolytic Enzymes. Enzym. Res. 2015, 2015, 279381. [Google Scholar] [CrossRef] [Green Version]
- Saini, A.; Aggarwal, N.K.; Yadav, A. Cellulolytic Potential of Actinomycetes Isolated from Different Habitats. Bioeng. Biosci. 2016, 4, 88–94. [Google Scholar] [CrossRef]
- Lee, S.; Kang, M.; Bae, J.-H.; Sohn, J.-H.; Sung, B.H. Bacterial Valorization of Lignin: Strains, Enzymes, Conversion Pathways, Biosensors, and Perspectives. Front. Bioeng. Biotechnol. 2019, 7, 209. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Zhang, K.; Liu, P.; Han, H.; Zhao, S.; Kakade, A.; Khan, A.; Du, D.; Li, X. Lignin Depolymerization and Utilization by Bacteria. Bioresour. Technol. 2018, 269, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Singh, T.; Pal, D.B.; Khan, S.; Ahmad, S.; Jandrajupalli, S.B.; Haque, S.; Singh, R.; Srivastava, N. Enhanced Production of Bacterial Hydrolytic Endoglucanase Enzyme Using Waste Leaves of Water Hyacinth and Its Thermal Stability under the Influence of TiO2 Nanoparticles. Biomass Convers. Biorefin. 2022. [Google Scholar] [CrossRef]
- Ejaz, U.; Moin, S.F.; Sohail, M.; Mersal, G.A.M.; Ibrahim, M.M.; El-Bahy, S.M. Characterization of a Novel End Product Tolerant and Thermostable Cellulase from Neobacillus sedimentimangrovi UE25. Enzym. Microb. Technol. 2023, 162, 110133. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Ali, F.; Rahman, M. Cellulase Activity of a Novel Bacterial Strain Arthrobacter woluwensis TDS9: Its Application on Bioconversion of Paper Mill Sludge. J. Genet. Eng. Biotechnol. 2022, 20, 87. [Google Scholar] [CrossRef] [PubMed]
- Akram, F.; ul Haq, I.; Shah, F.I.; Aqeel, A.; Ahmed, Z.; Mir, A.S.; Qureshi, S.S.; Raja, S.I. Genus Thermotoga: A Valuable Home of Multifunctional Glycoside Hydrolases (GHs) for Industrial Sustainability. Bioorg. Chem. 2022, 127, 105942. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Liu, S.; Dong, S.; Li, R.; Feng, Y.; Cui, Q. Determination of the Native Features of the Exoglucanase Cel48S from Clostridium thermocellum. Biotechnol. Biofuels 2018, 11, 6. [Google Scholar] [CrossRef]
- Yang, J.; Ma, T.; Shang-guan, F.; Han, Z. Improving the Catalytic Activity of Thermostable Xylanase from Thermotoga maritima via Mutagenesis of Non-Catalytic Residues at Glycone Subsites. Enzym. Microb. Technol 2020, 139, 109579. [Google Scholar] [CrossRef]
- Xue, D.; Zeng, X.; Lin, D.; Yao, S. Thermostable Ethanol Tolerant Xylanase from a Cold-Adapted Marine Species Acinetobacter johnsonii. Chin. J. Chem. Eng. 2019, 27, 1166–1170. [Google Scholar] [CrossRef]
- Bakry, M.M.; Salem, S.S.; Atta, H.M.; El-Gamal, M.S.; Fouda, A. Xylanase from Thermotolerant Bacillus haynesii Strain, Synthesis, Characterization, Optimization Using Box-Behnken Design, and Biobleaching Activity. Biomass Convers. Biorefin. 2022. [Google Scholar] [CrossRef]
- Mhiri, S.; Bouanane-Darenfed, A.; Jemli, S.; Neifar, S.; Ameri, R.; Mezghani, M.; Bouacem, K.; Jaouadi, B.; Bejar, S. A Thermophilic and Thermostable Xylanase from Caldicoprobacter algeriensis: Recombinant Expression, Characterization and Application in Paper Biobleaching. Int. J. Biol. Macromol. 2020, 164, 808–817. [Google Scholar] [CrossRef]
- Mehandia, S.; Sharma, S.C.; Arya, S.K. Isolation and Characterization of an Alkali and Thermostable Laccase from a Novel Alcaligenes faecalis and Its Application in Decolorization of Synthetic Dyes. Biotechnol. Rep. 2020, 25, e00413. [Google Scholar] [CrossRef]
- Wang, J.; Chang, F.; Tang, X.; Li, W.; Yin, Q.; Yang, Y.; Hu, Y. Bacterial Laccase of Anoxybacillus ayderensis SK3-4 from Hot Springs Showing Potential for Industrial Dye Decolorization. Ann. Microbiol. 2020, 70, 51. [Google Scholar] [CrossRef]
- Abdelgalil, S.A.; Soliman, N.A.; Abo-Zaid, G.A.; Abdel-Fattah, Y.R. Bioprocessing Strategies for Cost-Effective Large-Scale Production of Bacterial Laccase from Lysinibacillus macroides LSO Using Bio-Waste. Int. J. Environ. Sci. Technol. 2022, 19, 1633–1652. [Google Scholar] [CrossRef]
- Khaled, J.M.; Alyahya, S.A.; Govindan, R.; Chelliah, C.K.; Maruthupandy, M.; Alharbi, N.S.; Kadaikunnan, S.; Issac, R.; Murugan, S.; Li, W.-J. Laccase Producing Bacteria Influenced the High Decolorization of Textile Azo Dyes with Advanced Study. Environ. Res. 2022, 207, 112211. [Google Scholar] [CrossRef]
- Payne, C.M.; Knott, B.C.; Mayes, H.B.; Hansson, H.; Himmel, M.E.; Sandgren, M.; Ståhlberg, J.; Beckham, G.T. Fungal Cellulases. Chem. Rev. 2015, 115, 1308–1448. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, N.; Srivastava, M.; Mishra, P.K.; Gupta, V.K.; Molina, G.; Rodriguez-Couto, S.; Manikanta, A.; Ramteke, P.W. Applications of Fungal Cellulases in Biofuel Production: Advances and Limitations. Renew. Sustain. Energy Rev. 2018, 82, 2379–2386. [Google Scholar] [CrossRef]
- Muaaz-Us-Salam, S.; Cleall, P.J.; Harbottle, M.J. Application of Enzymatic and Bacterial Biodelignification Systems for Enhanced Breakdown of Model Lignocellulosic Wastes. Sci. Total Environ. 2020, 728, 138741. [Google Scholar] [CrossRef]
- Punt, P.J.; van Biezen, N.; Conesa, A.; Albers, A.; Mangnus, J.; van den Hondel, C. Filamentous Fungi as Cell Factories for Heterologous Protein Production. Trends Biotechnol. 2002, 20, 200–206. [Google Scholar] [CrossRef]
- Shoji, J.; Kikuma, T.; Kitamoto, K. Vesicle Trafficking, Organelle Functions, and Unconventional Secretion in Fungal Physiology and Pathogenicity. Curr. Opin. Microbiol. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Shoji, J.; Arioka, M.; Kitamoto, K. Dissecting Cellular Components of the Secretory Pathway in Filamentous Fungi: Insights into Their Application for Protein Production. Biotechnol. Lett. 2007, 30, 7–14. [Google Scholar] [CrossRef]
- Bischof, R.H.; Ramoni, J.; Seiboth, B. Cellulases and beyond: The First 70 Years of the Enzyme Producer Trichoderma reesei. Microb. Cell Factories 2016, 15, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A. Aspergillus nidulans: A Potential Resource of the Production of the Native and Heterologous Enzymes for Industrial Applications. Int. J. Microbiol. 2020, 2020, 8894215. [Google Scholar] [CrossRef]
- Bhati, N.; Shreya; Sharma, A.K. Cost-effective Cellulase Production, Improvement Strategies, and Future Challenges. J. Food Process Eng. 2021, 44, e13623. [Google Scholar] [CrossRef]
- Fang, H.; Li, C.; Zhao, J.; Zhao, C. Biotechnological Advances and Trends in Engineering Trichoderma reesei towards Cellulase Hyperproducer. Biotechnol. Bioprocess. Eng. 2021, 26, 517–528. [Google Scholar] [CrossRef]
- Liu, L.; Huang, W.-C.; Liu, Y.; Li, M. Diversity of Cellulolytic Microorganisms and Microbial Cellulases. Int. Biodeterior. Biodegrad. 2021, 163, 105277. [Google Scholar] [CrossRef]
- Borin, G.P.; Sanchez, C.C.; de Souza, A.P.; de Santana, E.S.; de Souza, A.T.; Leme, A.F.P.; Squina, F.M.; Buckeridge, M.; Goldman, G.H.; Oliveira, J.V.D.C. Comparative Secretome Analysis of Trichoderma reesei and Aspergillus niger during Growth on Sugarcane Biomass. PLoS ONE 2015, 10, e0129275. [Google Scholar] [CrossRef]
- Rastegari, A.A. Molecular Mechanism of Cellulase Production Systems in Penicillium. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 153–166. [Google Scholar]
- Vaishnav, N.; Singh, A.; Adsul, M.; Dixit, P.; Sandhu, S.K.; Mathur, A.; Puri, S.K.; Singhania, R.R. Penicillium: The next Emerging Champion for Cellulase Production. Bioresour. Technol. Rep. 2018, 2, 131–140. [Google Scholar] [CrossRef]
- Méndez-Líter, J.A.; de Eugenio, L.I.; Nieto-Domínguez, M.; Prieto, A.; Martínez, M.J. Hemicellulases from Penicillium and Talaromyces for Lignocellulosic Biomass Valorization: A Review. Bioresour. Technol. 2021, 324, 124623. [Google Scholar] [CrossRef]
- Mäkelä, M.R.; Hildén, K.; Kowalczyk, J.E.; Hatakka, A. Progress and Research Needs of Plant Biomass Degradation by Basidiomycete Fungi. In Grand Challenges in Fungal Biotechnology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 405–438. [Google Scholar]
- Bentil, J.A. Biocatalytic Potential of Basidiomycetes: Relevance, Challenges and Research Interventions in Industrial Processes. Sci. Afr. 2021, 11, e00717. [Google Scholar] [CrossRef]
- Liu, J.; Yang, J.; Wang, R.; Liu, L.; Zhang, Y.; Bao, H.; Jang, J.M.; Wang, E.; Yuan, H. Comparative Characterization of Extracellular Enzymes Secreted by Phanerochaete chrysosporium during Solid-State and Submerged Fermentation. Int. J. Biol. Macromol. 2020, 152, 288–294. [Google Scholar] [CrossRef]
- Okal, E.J.; Aslam, M.M.; Karanja, J.K.; Nyimbo, W.J. Mini Review: Advances in Understanding Regulation of Cellulase Enzyme in White-Rot Basidiomycetes. Microb. Pathog. 2020, 147, 104410. [Google Scholar] [CrossRef]
- Pavlenko, A.A.; Kopilov, E.P.; Nadkernychna, O.V.; Tsekhmister, H.V.; Kyslynska, A.S. Cellulase and Polygalacturonase Activity by Trichoderma viride. Agric. Conspec. Sci. 2022, 87, 35–43. [Google Scholar]
- Srivastava, N.; Elgorban, A.M.; Mishra, P.K.; Marraiki, N.; Alharbi, A.M.; Ahmad, I.; Gupta, V.K. Enhance Production of Fungal Cellulase Cocktail Using Cellulosic Waste. Environ. Technol. Innov. 2020, 19, 100949. [Google Scholar] [CrossRef]
- Naik, A.N.K.; Anuradha, C.M.; Cheemanapalli, S.; Kumar, C.S. Comparison of Production of Cellulolytic Enzymes by Fusarium sp. Under Optimized and Non-Optimized Conditions. J. Environ. Bio-Sci. 2020, 24, 153–168. [Google Scholar]
- Santos, G.B.; de Sousa Francisco Filho, Á.; Rêgo da Silva Rodrigues, J.; Rodrigues de Souza, R. Cellulase Production by Aspergillus niger Using Urban Lignocellulosic Waste as Substrate: Evaluation of Different Cultivation Strategies. J. Environ. Manag. 2022, 305, 114431. [Google Scholar] [CrossRef]
- Mahmood, R.T.; Masood, M.; Zia, N.; Safder, A.; Asad, M.J.; Nasreen, S.; Ahmed, D.; Iqbal, R. Production and Characterization of Endoglucanase from Phaeolus spadiceus. Pak. J. Biochem. Biotechnol. 2021, 2, 27–37. [Google Scholar] [CrossRef]
- Alves, T.P.; Triques, C.C.; da Silva, E.A.; Fagundes-Klen, M.R.; Hasan, S.D.M. Multi-enzymatic Recovery of Fungal Cellulases (Aspergillus niger) through Solid-state Fermentation of Sugarcane Bagasse. Can. J. Chem. Eng. 2022, 100, 1930–1940. [Google Scholar] [CrossRef]
- Gooruee, R.; Hojjati, M.; Behbahani, B.A.; Shahbazi, S.; Askari, H. Extracellular Enzyme Production by Different Species of Trichoderma Fungus for Lemon Peel Waste Bioconversion. Biomass Convers. Biorefin. 2022. [Google Scholar] [CrossRef]
- Dhaver, P.; Pletschke, B.; Sithole, B.; Govinden, R. Isolation, Screening, Preliminary Optimisation and Characterisation of Thermostable Xylanase Production under Submerged Fermentation by Fungi in Durban, South Africa. Mycology 2022, 13, 271–292. [Google Scholar] [CrossRef]
- Intasit, R.; Cheirsilp, B.; Suyotha, W.; Boonsawang, P. Purification and Characterization of a Highly-Stable Fungal Xylanase from Aspergillus tubingensis Cultivated on Palm Wastes through Combined Solid-State and Submerged Fermentation. Prep. Biochem. Biotechnol. 2022, 52, 311–317. [Google Scholar] [CrossRef]
- Barbieri, G.S.; Bento, H.B.S.; de Oliveira, F.; Picheli, F.P.; Dias, L.M.; Masarin, F.; Santos-Ebinuma, V.C. Xylanase Production by Talaromyces amestolkiae Valuing Agroindustrial Byproducts. BioTech 2022, 11, 15. [Google Scholar] [CrossRef]
- Ambatkar, N.; Jadhav, D.D.; Nandi, S.; Kumbhar, P.; Kommoju, P.-R. Optimized Process for the Production of Fungal Peroxidases and Efficient Saccharification of Pre-Treated Rice Straw. Bioresour. Technol. Rep. 2022, 17, 100913. [Google Scholar] [CrossRef]
- Illuri, R.; Kumar, M.; Eyini, M.; Veeramanikandan, V.; Almaary, K.S.; Elbadawi, Y.B.; Biraqdar, M.A.; Balaji, P. Production, Partial Purification and Characterization of Ligninolytic Enzymes from Selected Basidiomycetes Mushroom Fungi. Saudi J. Biol. Sci. 2021, 28, 7207–7218. [Google Scholar] [CrossRef]
- Bouacem, K.; Allala, F.; Zaraî Jaouadi, N.; Hamdi, S.; Mechri, S.; Ighilahriz, K.; Rekik, H.; Hacene, H.; Bouanane-Darenfed, A.; Jaouadi, B. A Novel Peroxidase from White-Rot Agaricomycetes Fungus Phlebia Radiata Strain KB-DZ15: Its Purification, Characterisation, and Potential Application for Dye-Decolorisation and Lignin-Biodegradation. Biocatal. Biotransform. 2022, 40, 365–377. [Google Scholar] [CrossRef]
- Ezike, T.C.; Ezugwu, A.L.; Udeh, J.O.; Eze, S.O.O.; Chilaka, F.C. Purification and Characterisation of New Laccase from Trametes polyzona WRF03. Biotechnol. Rep. 2020, 28, e00566. [Google Scholar] [CrossRef]
- Backes, E.; Kato, C.G.; da Silva, T.B.V.; Uber, T.M.; Pasquarelli, D.L.; Bracht, A.; Peralta, R.M. Production of Fungal Laccase on Pineapple Waste and Application in Detoxification of Malachite Green. J. Environ. Sci. Health Part B 2022, 57, 90–101. [Google Scholar] [CrossRef]
- Cen, Q.; Wu, X.; Cao, L.; Lu, Y.; Lu, X.; Chen, J.; Fu, G.; Liu, Y.; Ruan, R. Green Production of a Yellow Laccase by Coriolopsis gallica for Phenolic Pollutants Removal. AMB Express 2022, 12, 96. [Google Scholar] [CrossRef]
- Tian, Q.; Dou, X.; Huang, L.; Wang, L.; Meng, D.; Zhai, L.; Shen, Y.; You, C.; Guan, Z.; Liao, X. Characterization of a Robust Cold-Adapted and Thermostable Laccase from Pycnoporus sp. SYBC-L10 with a Strong Ability for the Degradation of Tetracycline and Oxytetracycline by Laccase-Mediated Oxidation. J. Hazard. Mater. 2020, 382, 121084. [Google Scholar] [CrossRef]
- Muñoz-Arenas, L.C.; Fusaro, C.; Hernández-Guzmán, M.; Dendooven, L.; Estrada-Torres, A.; Navarro-Noya, Y.E. Soil Microbial Diversity Drops with Land-use Change in a High Mountain Temperate Forest: A Metagenomics Survey. Environ. Microbiol. Rep. 2020, 12, 185–194. [Google Scholar] [CrossRef]
- Hermosilla, E.; Rubilar, O.; Schalchli, H.; da Silva, A.S.; Ferreira-Leitao, V.; Diez, M.C. Sequential White-Rot and Brown-Rot Fungal Pretreatment of Wheat Straw as a Promising Alternative for Complementary Mild Treatments. Waste Manag. 2018, 79, 240–250. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; Singh, R.; Eltis, L.D.; Mohn, W.W. Bacterial Contributions to Delignification and Lignocellulose Degradation in Forest Soils with Metagenomic and Quantitative Stable Isotope Probing. ISME J. 2019, 13, 413–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-García, L.; Bugg, T.D.H.; Jiménez, D.J. Exploring the Lignin Catabolism Potential of Soil-Derived Lignocellulolytic Microbial Consortia by a Gene-Centric Metagenomic Approach. Microb. Ecol. 2020, 80, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Paixão, D.A.A.; Tomazetto, G.; Sodré, V.R.; Gonçalves, T.A.; Uchima, C.A.; Büchli, F.; Alvarez, T.M.; Persinoti, G.F.; da Silva, M.J.; Bragatto, J.; et al. Microbial Enrichment and Meta-Omics Analysis Identify CAZymes from Mangrove Sediments with Unique Properties. Enzym. Microb. Technol. 2021, 148, 109820. [Google Scholar] [CrossRef]
- Shinde, R.; Shahi, D.K.; Mahapatra, P.; Naik, S.K.; Singh, C.S.; Verma, S.; Singh, A.K. Isolation of Lignocelluloses Degrading Microbes from Soil and Their Screening Based on Qualitative Analysis and Enzymatic Assays. Ann. Plant Soil Res. 2022, 24, 347–354. [Google Scholar] [CrossRef]
- Balabanova, L.; Slepchenko, L.; Son, O.; Tekutyeva, L. Biotechnology Potential of Marine Fungi Degrading Plant and Algae Polymeric Substrates. Front. Microbiol. 2018, 9, 1527. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.-Y.; Li, X. Alkali-Stable Cellulase from a Halophilic Isolate, Gracilibacillus Sp. SK1 and Its Application in Lignocellulosic Saccharification for Ethanol Production. Biomass Bioenergy 2015, 81, 19–25. [Google Scholar] [CrossRef]
- Barzkar, N.; Sohail, M. An Overview on Marine Cellulolytic Enzymes and Their Potential Applications. Appl. Microbiol. Biotechnol. 2020, 104, 6873–6892. [Google Scholar] [CrossRef]
- Sethi, R.; Padmavathi, T.; Sullia, S.B. Lignocellulose Biomass Degradation by Marine Microorganisms. Eur. J. Exp. Biol. 2013, 3, 129–138. [Google Scholar]
- Xiong, X.Q.; Liao, H.D.; Ma, J.S.; Liu, X.M.; Zhang, L.Y.; Shi, X.W.; Yang, X.L.; Lu, X.N.; Zhu, Y.H. Isolation of a Rice Endophytic Bacterium, Pantoea sp. Sd-1, with Ligninolytic Activity and Characterization of Its Rice Straw Degradation Ability. Lett. Appl. Microbiol. 2014, 58, 123–129. [Google Scholar] [CrossRef]
- Corrêa, R.C.G.; Rhoden, S.A.; Mota, T.R.; Azevedo, J.L.; Pamphile, J.A.; de Souza, C.G.M.; Polizeli, M.L.T.M.; Bracht, A.; Peralta, R.M. Endophytic Fungi: Expanding the Arsenal of Industrial Enzyme Producers. J. Ind. Microbiol. Biotechnol. 2014, 41, 1467–1478. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, K.; Liao, H.; Hector, S.B.; Shi, X.; Li, J.; Liu, B.; Xu, T.; Tong, C.; Liu, X.; et al. Genomic and Secretomic Insight into Lignocellulolytic System of an Endophytic Bacterium Pantoea ananatis Sd-1. Biotechnol. Biofuels 2016, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Robl, D.; Mergel, C.M.; Costa, P.D.S.; Pradella, J.G.D.C.; Padilla, G. Endophytic Actinomycetes as Potential Producers of Hemicellulases and Related Enzymes for Plant Biomass Degradation. Braz. Arch. Biol. Technol. 2019, 62, e19180337. [Google Scholar] [CrossRef] [Green Version]
- Satyanarayana, T.; Raghukumar, C.; Shivaji, S. Extremophilic Microbes: Diversity and Perspectives. Curr. Sci. 2005, 89, 78–90. [Google Scholar]
- Pikuta, E.V.; Hoover, R.B.; Tang, J. Microbial Extremophiles at the Limits of Life. Crit. Rev. Microbiol. 2007, 33, 183–209. [Google Scholar] [CrossRef]
- Elleuche, S.; Schröder, C.; Sahm, K.; Antranikian, G. Extremozymes—Biocatalysts with Unique Properties from Extremophilic Microorganisms. Curr. Opin. Biotechnol. 2014, 29, 116–123. [Google Scholar] [CrossRef]
- Arora, N.K.; Panosyan, H. Extremophiles: Applications and Roles in Environmental Sustainability. Environ. Sustain. 2019, 2, 217–218. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Vaid, S.; Bhat, B.; Singh, S.; Bajaj, B.K. Thermostable Enzymes for Industrial Biotechnology. In Advances in Enzyme Technology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 469–495. [Google Scholar]
- Chettri, D.; Verma, A.K.; Sarkar, L.; Verma, A.K. Role of Extremophiles and Their Extremozymes in Biorefinery Process of Lignocellulose Degradation. Extremophiles 2021, 25, 203–219. [Google Scholar] [CrossRef]
- Da-qing, W.; Wen-ran, J.; Tai-peng, S.; Yu-tian, M.; Wei, Z.; Hong-yan, W. Screening Psychrophilic Fungi of Cellulose Degradation and Characteristic of Enzyme Production. J. Northeast Agric. Univ. 2016, 23, 20–27. [Google Scholar] [CrossRef]
- Lamilla, C.; Pavez, M.; Santos, A.; Hermosilla, A.; Llanquinao, V.; Barrientos, L. Bioprospecting for Extracellular Enzymes from Culturable Actinobacteria from the South Shetland Islands, Antarctica. Polar Biol. 2017, 40, 719–726. [Google Scholar] [CrossRef]
- Brück, S.A.; Contato, A.G.; Gamboa-Trujillo, P.; Oliveira, T.B.; Cereia, M.; Polizeli, M.L.T.M. Prospection of Psychrotrophic Filamentous Fungi Isolated from the High Andean Paramo Region of Northern Ecuador: Enzymatic Activity and Molecular Identification. Microorganisms 2022, 10, 282. [Google Scholar] [CrossRef]
- Médigue, C.; Krin, E.; Pascal, G.; Barbe, V.; Bernsel, A.; Bertin, P.N.; Cheung, F.; Cruveiller, S.; D’Amico, S.; Duilio, A.; et al. Coping with Cold: The Genome of the Versatile Marine Antarctica Bacterium Pseudoalteromonas haloplanktis TAC125. Genome Res. 2005, 15, 1325–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, L.M.; Braña, V.; Franco Fraguas, L.; Castro-Sowinski, S. Characterization of the Cellulase-Secretome Produced by the Antarctic Bacterium Flavobacterium sp. AUG42. Microbiol. Res. 2019, 223–225, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, J.; Zhang, W.; Huang, H.; Shi, P.; Luo, H.; Liu, B.; Zhang, Y.; Zhang, Z.; Fan, Y.; et al. A Neutral Thermostable β-1,4-Glucanase from Humicola insolens Y1 with Potential for Applications in Various Industries. PLoS ONE 2015, 10, e0124925. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, C.; Basotra, N.; Singh, S.; di Falco, M.; Tsang, A.; Chadha, B.S. Malbranchea cinnamomea: A Thermophilic Fungal Source of Catalytically Efficient Lignocellulolytic Glycosyl Hydrolases and Metal Dependent Enzymes. Bioresour. Technol. 2016, 200, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Ahirwar, S.; Soni, H.; Prajapati, B.P.; Kango, N. Isolation and Screening of Thermophilic and Thermotolerant Fungi for Production of Hemicellulases from Heated Environments. Mycology 2017, 8, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Kvesitadze, G. Cellulases from Extremophiles. Curr. Trends Biomed. Eng. Biosci. 2017, 4, 44–49. [Google Scholar] [CrossRef]
- Pechtl, A.; Rückert, C.; Maus, I.; Koeck, D.E.; Trushina, N.; Kornberger, P.; Schwarz, W.H.; Schlüter, A.; Liebl, W.; Zverlov, V.V. Complete Genome Sequence of the Novel Cellulolytic, Anaerobic, Thermophilic Bacterium Herbivorax saccincola Type Strain GGR1, Isolated from a Lab Scale Biogas Reactor as Established by Illumina and Nanopore MinION Sequencing. Genome Announc. 2018, 6, e01493-17. [Google Scholar] [CrossRef] [Green Version]
- Leo, V.V.; Asem, D.; Zothanpuia; Singh, B.P. Actinobacteria. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 191–205. [Google Scholar]
- Williams-Rhaesa, A.M.; Rubinstein, G.M.; Scott, I.M.; Lipscomb, G.L.; Poole, I.F.L.; Kelly, R.M.; Adams, M.W.W. Engineering Redox-Balanced Ethanol Production in the Cellulolytic and Extremely Thermophilic Bacterium, Caldicellulosiruptor bescii. Metab. Eng. Commun. 2018, 7, e00073. [Google Scholar] [CrossRef]
- Xu, H.; Han, D.; Xu, Z. Expression of Heterologous Cellulases in Thermotoga sp. Strain RQ2. Biomed. Res. Int. 2015, 2015, 304523. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Chen, Y.; Peng, W.; Wang, P.; Zhao, Y.; Li, X.; Wang, F.; Li, X. Fusion Endoglucanase Cel12B from Thermotoga maritima with Cellulose Binding Domain. Bioresources 2018, 13, 4497–4508. [Google Scholar] [CrossRef]
- Tamariz-Angeles, C.; Lázaro-Palomino, J.; Olivera-Gonzales, P.; Castañeda-Barreto, A.; Villena, G.K. Isolation of Thermotolerant Bacillus subtilis DCH4 from Chancos Hot Spring (Carhuaz, Peru) with Potential to Degrade Lignocellulosic Agriculture Wastes. Rev. Peru. Biol. 2020, 27, 067–078. [Google Scholar] [CrossRef] [Green Version]
- Cilmeli, S.; Doruk, T.; Könen-Adıgüzel, S.; Adıgüzel, A.O. A Thermostable and Acidophilic Mannanase from Bacillus mojavensis: Its Sustainable Production Using Spent Coffee Grounds, Characterization, and Application in Grape Juice Processing. Biomass Convers. Biorefin. 2022. [Google Scholar] [CrossRef]
- Raj, A.; Kumar, S.; Singh, S.K.; Prakash, J. Production and Purification of Xylanase from Alkaliphilic Bacillus licheniformis and Its Pretreatment of Eucalyptus Kraft Pulp. Biocatal. Agric. Biotechnol. 2018, 15, 199–209. [Google Scholar] [CrossRef]
- Lee, J.P.; Seo, G.-W.; An, S.-D.; Kim, H. A Cold-Active Acidophilic Endoglucanase of Paenibacillus sp. Y2 Isolated from Soil in an Alpine Region. J. Appl. Biol. Chem. 2017, 60, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ning, C.; Yuan, M.; Yang, S.; Wei, X.; Xiao, M.; Fu, X.; Zhu, C.; Mou, H. High-Level Expression of a Thermophilic and Acidophilic β-Mannanase from Aspergillus kawachii IFO 4308 with Significant Potential in Mannooligosaccharide Preparation. Bioresour. Technol. 2020, 295, 122257. [Google Scholar] [CrossRef]
- Saroj, P.; Narasimhulu, K. Assessment and Evaluation of Cellulase Production Using Ragi (Eleusine coracana) Husk as a Substrate from Thermo-Acidophilic Aspergillus fumigatus JCM 10253. Bioprocess Biosyst. Eng. 2021, 44, 113–126. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, J.; Li, C.; Wang, T.-F.; Qin, H.-M. Biochemical and Structural Characterization of a Novel Thermophilic and Acidophilic β-Mannanase from Aspergillus calidoustus. Enzym. Microb. Technol. 2021, 150, 109891. [Google Scholar] [CrossRef]
- Olopoda, I.A.; Lawal, O.T.; Omotoyinbo, O.V.; Kolawole, A.N.; Sanni, D.M. Biochemical Characterization of a Thermally Stable, Acidophilic and Surfactant-Tolerant Xylanase from Aspergillus awamori AFE1 and Hydrolytic Efficiency of Its Immobilized Form. Process Biochem. 2022, 121, 45–55. [Google Scholar] [CrossRef]
- Vega, K.; Villena, G.K.; Sarmiento, V.H.; Ludeña, Y.; Vera, N.; Gutiérrez-Correa, M. Production of Alkaline Cellulase by Fungi Isolated from an Undisturbed Rain Forest of Peru. Biotechnol. Res. Int. 2012, 2012, 934325. [Google Scholar] [CrossRef] [Green Version]
- Turner, P.; Mamo, G.; Karlsson, E.N. Potential and Utilization of Thermophiles and Thermostable Enzymes in Biorefining. Microb. Cell Factories 2007, 6, 9. [Google Scholar] [CrossRef]
- Akram, F.; ul Haq, I.; Aqeel, A.; Ahmed, Z.; Shah, F.I. Thermostable Cellulases: Structure, Catalytic Mechanisms, Directed Evolution and Industrial Implementations. Renew. Sustain. Energy Rev. 2021, 151, 111597. [Google Scholar] [CrossRef]
- Maurice, N. Role of Solid-State Fermentation to Enhance Cellulase Production. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 127–153. [Google Scholar]
- Singh, A.; Bajar, S.; Devi, A.; Pant, D. An Overview on the Recent Developments in Fungal Cellulase Production and Their Industrial Applications. Bioresour. Technol. Rep. 2021, 14, 100652. [Google Scholar] [CrossRef]
- Hansen, G.H.; Lübeck, M.; Frisvad, J.C.; Lübeck, P.S.; Andersen, B. Production of Cellulolytic Enzymes from Ascomycetes: Comparison of Solid State and Submerged Fermentation. Process Biochem. 2015, 50, 1327–1341. [Google Scholar] [CrossRef]
- Prévot, V.; Lopez, M.; Copinet, E.; Duchiron, F. Comparative Performance of Commercial and Laboratory Enzymatic Complexes from Submerged or Solid-State Fermentation in Lignocellulosic Biomass Hydrolysis. Bioresour. Technol. 2013, 129, 690–693. [Google Scholar] [CrossRef] [PubMed]
- Karimi, F.; Mazaheri, D.; Saei Moghaddam, M.; Mataei Moghaddam, A.; Sanati, A.L.; Orooji, Y. Solid-State Fermentation as an Alternative Technology for Cost-Effective Production of Bioethanol as Useful Renewable Energy: A Review. Biomass Convers. Biorefin. 2021. [Google Scholar] [CrossRef]
- Ramesh, D.; Muniraj, I.K.; Thangavelu, K.; Karthikeyan, S. Knowledge Update on Bioreactor Technology for Cellulase Production. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 181–193. [Google Scholar]
- Leite, P.; Sousa, D.; Fernandes, H.; Ferreira, M.; Costa, A.R.; Filipe, D.; Gonçalves, M.; Peres, H.; Belo, I.; Salgado, J.M. Recent Advances in Production of Lignocellulolytic Enzymes by Solid-State Fermentation of Agro-Industrial Wastes. Curr. Opin. Green Sustain. Chem. 2021, 27, 100407. [Google Scholar] [CrossRef]
- Pino, M.S.; Rodríguez-Jasso, R.M.; Michelin, M.; Flores-Gallegos, A.C.; Morales-Rodriguez, R.; Teixeira, J.A.; Ruiz, H.A. Bioreactor Design for Enzymatic Hydrolysis of Biomass under the Biorefinery Concept. Chem. Eng. J. 2018, 347, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, V.E.; Michelin, M.; Vici, A.C.; de Almeida, P.Z.; Polizeli, M.L.T.M. Trametes versicolor Laccase Production Using Agricultural Wastes: A Comparative Study in Erlenmeyer Flasks, Bioreactor and Tray. Bioprocess Biosyst. Eng. 2020, 43, 507–514. [Google Scholar] [CrossRef]
- Areeshi, M.Y. Microbial Cellulase Production Using Fruit Wastes and Its Applications in Biofuels Production. Int. J. Food Microbiol. 2022, 378, 109814. [Google Scholar] [CrossRef]
- Contato, A.G.; Oliveira, T.B.; Aranha, G.M.; de Freitas, E.N.; Vici, A.C.; Nogueira, K.M.V.; de Lucas, R.C.; de Almeida Scarcella, A.S.; Buckeridge, M.S.; Silva, R.N.; et al. Prospection of Fungal Lignocellulolytic Enzymes Produced from Jatoba (Hymenaea courbaril) and Tamarind (Tamarindus indica) Seeds: Scaling for Bioreactor and Saccharification Profile of Sugarcane Bagasse. Microorganisms 2021, 9, 533. [Google Scholar] [CrossRef]
- Kumla, J.; Suwannarach, N.; Sujarit, K.; Penkhrue, W.; Kakumyan, P.; Jatuwong, K.; Vadthanarat, S.; Lumyong, S. Cultivation of Mushrooms and Their Lignocellulolytic Enzyme Production Through the Utilization of Agro-Industrial Waste. Molecules 2020, 25, 2811. [Google Scholar] [CrossRef]
- Rana, P.; Inbaraj, B.S.; Gurumayum, S.; Sridhar, K. Sustainable Production of Lignocellulolytic Enzymes in Solid-State Fermentation of Agro-Industrial Waste: Application in Pumpkin (Cucurbita maxima) Juice Clarification. Agronomy 2021, 11, 2379. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Shetti, N.P.; Reddy, K.R.; Kwon, E.E.; Nadagouda, M.N.; Aminabhavi, T.M. Biomass Utilization and Production of Biofuels from Carbon Neutral Materials. Environ. Pollut. 2021, 276, 116731. [Google Scholar] [CrossRef]
- Zahra, T.; Irfan, M.; Nadeem, M.; Ghazanfar, M.; Ahmad, Q.; Ali, S.; Siddique, F.; Yasmeen, Z.; Franco, M. Cellulase Production by Trichoderma viride in Submerged Fermentation Using Response Surface Methodology. Punjab Univ. J. Zool. 2020, 35, 223–228. [Google Scholar] [CrossRef]
- Bušić, A.; Marđetko, N.; Kundas, S.; Morzak, G.; Belskaya, H.; Ivančić Šantek, M.; Komes, D.; Novak, S.; Šantek, B. Bioethanol Production from Renewable Raw Materials and Its Separation and Purification: A Review. Food Technol. Biotechnol. 2018, 56, 289–311. [Google Scholar] [CrossRef]
- Rojas, L.F.; Zapata, P.; Ruiz-Tirado, L. Agro-Industrial Waste Enzymes: Perspectives in Circular Economy. Curr. Opin. Green Sustain. Chem. 2022, 34, 100585. [Google Scholar] [CrossRef]
- Pasin, T.M.; Scarcella, A.S.A.; Oliveira, T.B.; Lucas, R.C.; Cereia, M.; Betini, J.H.A.; Polizeli, M.L.T.M. Paper Industry Wastes as Carbon Sources for Aspergillus Species Cultivation and Production of an Enzymatic Cocktail for Biotechnological Applications. Ind. Biotechnol. 2020, 16, 56–60. [Google Scholar] [CrossRef]
- Beladhadi, R.V.; Shankar, K.; Jayalakshmi, S.K.; Sreeramulu, K. Production of Cocktail of Lignolytic, Cellulolytic and Hemicellulolytic Enzymes by the Novel Bacterium burkholderia Sp SMB1 Utilizing Rice Bran and Straw: Application in the Saccharification of Untreated Agro-Wastes for Bioethanol Production. Waste Biomass Valorization 2022, 13, 1565–1577. [Google Scholar] [CrossRef]
- de Oliveira Júnior, S.D.; dos Santos Gouvêa, P.R.; dos Santos Vasconcelos, A.; de Brito do Nascimento, L.B.; Pessoa, V.A.; da Silva, G.L.; Sales-Campos, C.; Chevreuil, L.R. Cellulase and Xylanase Production by Ganoderma lucidum in Solid-State Fermentation Using Amazonian Lignocellulosic Wastes. Stud. Eng. Exact Sci. 2022, 3, 455–471. [Google Scholar] [CrossRef]
- Lu, X.; Li, F.; Zhou, X.; Hu, J.; Liu, P. Biomass, Lignocellulolytic Enzyme Production and Lignocellulose Degradation Patterns by Auricularia auricula during Solid State Fermentation of Corn Stalk Residues under Different Pretreatments. Food Chem. 2022, 384, 132622. [Google Scholar] [CrossRef]
- Sánchez-Corzo, L.D.; Álvarez-Gutiérrez, P.E.; Meza-Gordillo, R.; Villalobos-Maldonado, J.J.; Enciso-Pinto, S.; Enciso-Sáenz, S. Lignocellulolytic Enzyme Production from Wood Rot Fungi Collected in Chiapas, Mexico, and Their Growth on Lignocellulosic Material. J. Fungi 2021, 7, 450. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wei, Z.; Gao, X.; Wu, J.; Chen, X.; Zhao, Y.; Jia, L.; Wen, D. Reconstruction of Core Microbes Based on Producing Lignocellulolytic Enzymes Causing by Bacterial Inoculation during Rice Straw Composting. Bioresour. Technol. 2020, 315, 123849. [Google Scholar] [CrossRef]
- Chylenski, P.; Forsberg, Z.; Ståhlberg, J.; Várnai, A.; Lersch, M.; Bengtsson, O.; Sæbø, S.; Horn, S.J.; Eijsink, V.G.H. Development of Minimal Enzyme Cocktails for Hydrolysis of Sulfite-Pulped Lignocellulosic Biomass. J. Biotechnol. 2017, 246, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Ferreira Filho, E.X.; Moreira, L.R.S. An Update on Enzymatic Cocktails for Lignocellulose Breakdown. J. Appl. Microbiol. 2018, 125, 632–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarcella, A.S.A.; Pasin, T.M.; Oliveira, T.B.; Lucas, R.C.; Ferreira-Nozawa, M.S.; Freitas, E.N.; Vici, A.C.; Buckeridge, M.S.; Michelin, M.; Polizeli, M.L.T.M. Saccharification of Different Sugarcane Bagasse Varieties by Enzymatic Cocktails Produced by Mycothermus thermophilus and Trichoderma reesei RP698 Cultures in Agro-Industrial Residues. Energy 2021, 226, 120360. [Google Scholar] [CrossRef]
- Marđetko, N.; Trontel, A.; Novak, M.; Pavlečić, M.; Ljubas, B.D.; Grubišić, M.; Tominac, V.P.; Ludwig, R.; Šantek, B. Screening of Lignocellulolytic Enzyme Activities in Fungal Species and Sequential Solid-State and Submerged Cultivation for the Production of Enzyme Cocktails. Polymer 2021, 13, 3736. [Google Scholar] [CrossRef]
- Pinheiro, V.E.; de Almeida, P.Z.; Polizeli, M.L.T.M. Statistical Optimization of Cornmeal Saccharification Using Various Hydrolases. Biomass Convers. Biorefin. 2021. [Google Scholar] [CrossRef]
- Naitam, M.G.; Kaushik, R.; Tomar, G.S. Optimization and Production of Holocellulosic Enzyme Cocktail from Fungi Aspergillus nidulans under Solid-State Fermentation for the Production of Poly(3-Hydroxybutyrate). Res. Sq. 2022. [Google Scholar] [CrossRef]
- Jiménez, D.J.; de Lima Brossi, M.J.; Schückel, J.; Kračun, S.K.; Willats, W.G.T.; van Elsas, J.D. Characterization of Three Plant Biomass-Degrading Microbial Consortia by Metagenomics- and Metasecretomics-Based Approaches. Appl. Microbiol. Biotechnol. 2016, 100, 10463–10477. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, D.J.; Chaib De Mares, M.; Salles, J.F. Temporal Expression Dynamics of Plant Biomass-Degrading Enzymes by a Synthetic Bacterial Consortium Growing on Sugarcane Bagasse. Front. Microbiol. 2018, 9, 299. [Google Scholar] [CrossRef]
- Magurudeniya, H.D.; Baral, N.R.; Rodriguez, A.; Scown, C.D.; Dahlberg, J.; Putnam, D.; George, A.; Simmons, B.A.; Gladden, J.M. Use of Ensiled Biomass Sorghum Increases Ionic Liquid Pretreatment Efficiency and Reduces Biofuel Production Cost and Carbon Footprint. Green Chem. 2021, 23, 3127–3140. [Google Scholar] [CrossRef]
- Kallioinen, A.; Puranen, T.; Siika-aho, M. Mixtures of Thermostable Enzymes Show High Performance in Biomass Saccharification. Appl. Biochem. Biotechnol. 2014, 173, 1038–1056. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, D.J.; Dini-Andreote, F.; DeAngelis, K.M.; Singer, S.W.; Salles, J.F.; van Elsas, J.D. Ecological Insights into the Dynamics of Plant Biomass-Degrading Microbial Consortia. Trends Microbiol. 2017, 25, 788–796. [Google Scholar] [CrossRef]
- Fernandes, C.G.; Sawant, S.C.; Mule, T.A.; Khadye, V.S.; Lali, A.M.; Odaneth, A.A. Enhancing Cellulases through Synergistic β-Glucosidases for Intensifying Cellulose Hydrolysis. Process Biochem. 2022, 120, 202–212. [Google Scholar] [CrossRef]
- Toushik, S.H.; Lee, K.-T.; Lee, J.-S.; Kim, K.-S. Functional Applications of Lignocellulolytic Enzymes in the Fruit and Vegetable Processing Industries. J. Food Sci. 2017, 82, 585–593. [Google Scholar] [CrossRef] [Green Version]
- De Souza, T.S.P.; Kawaguti, H.Y. Cellulases, Hemicellulases, and Pectinases: Applications in the Food and Beverage Industry. Food Bioprocess Technol. 2021, 14, 1446–1477. [Google Scholar] [CrossRef]
- Soares, J.F.; Prá, V.D.; Kempka, A.P.; Prestes, R.C.; Tres, M.V.; Kuhn, R.C.; Mazutti, M.A. Cellulases for Food Applications. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2016; pp. 201–208. [Google Scholar]
- Singh, D.; Gupta, N. Microbial Laccase: A Robust Enzyme and Its Industrial Applications. Biologia 2020, 75, 1183–1193. [Google Scholar] [CrossRef]
- Kumar, A.V.; Suresh Chandra Kurup, R.; Snishamol, C.; Nagendra Prabhu, G. Role of Cellulases in Food, Feed, and Beverage Industries; Springer: Singapore, 2019; pp. 323–343. [Google Scholar]
- Vieira, S.L.; Stefanello, C.; Sorbara, J.O.B. Formulating Poultry Diets Based on Their Indigestible Components. Poult. Sci. 2014, 93, 2411–2416. [Google Scholar] [CrossRef]
- Lobedanz, S.; Damhus, T.; Borchert, T.V.; Hansen, T.T.; Lund, H.; Lai, W.; Lin, M.; Leclerc, M.; Kirk, O. Enzymes in Industrial Biotechnology. In Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 1–73. [Google Scholar]
- Bilal, M.; Iqbal, H.M.N. Sustainable Bioconversion of Food Waste into High-Value Products by Immobilized Enzymes to Meet Bio-Economy Challenges and Opportunities—A Review. Food Res. Int. 2019, 123, 226–240. [Google Scholar] [CrossRef]
- Wojdyło, A.; Samoticha, J.; Chmielewska, J. Effect of Different Pre-Treatment Maceration Techniques on the Content of Phenolic Compounds and Color of Dornfelder Wines Elaborated in Cold Climate. Food Chem. 2021, 339, 127888. [Google Scholar] [CrossRef]
- Chakraborty, S.; Gupta, R.; Jain, K.K.; Hemansi; Gautam, S.; Kuhad, R.C. Cellulases: Application in Wine and Brewery Industry. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2016; pp. 193–200. [Google Scholar]
- Espejo, F. Role of Commercial Enzymes in Wine Production: A Critical Review of Recent Research. J. Food Sci. Technol. 2021, 58, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Scott, A. Cutting Out Textile Pollution. Chem. Eng. News 2015, 93, 18–19. [Google Scholar] [CrossRef]
- Madhu, A.; Chakraborty, J.N. Developments in Application of Enzymes for Textile Processing. J. Clean. Prod. 2017, 145, 114–133. [Google Scholar] [CrossRef]
- Rahman, M.; Hack-Polay, D.; Billah, M.M.; Un Nabi, M.N. Bio-Based Textile Processing through the Application of Enzymes for Environmental Sustainability. Int. J. Technol. Manag. Sustain. Dev. 2020, 19, 87–106. [Google Scholar] [CrossRef]
- Rodríguez-Couto, S. A Promising Inert Support for Laccase Production and Decolouration of Textile Wastewater by the White-Rot Fungus Trametes pubescesns. J. Hazard. Mater. 2012, 233–234, 158–162. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, L.; Zhang, H.; Wang, S.; Zhang, X.; Geng, A. A Novel Homodimer Laccase from Cerrena unicolor BBP6: Purification, Characterization, and Potential in Dye Decolorization and Denim Bleaching. PLoS ONE 2018, 13, e0202440. [Google Scholar] [CrossRef] [Green Version]
- Panwar, V.; Sheikh, J.N.; Dutta, T. Sustainable Denim Bleaching by a Novel Thermostable Bacterial Laccase. Appl. Biochem. Biotechnol. 2020, 192, 1238–1254. [Google Scholar] [CrossRef]
- Tülek, A.; Karataş, E.; Çakar, M.M.; Aydın, D.; Yılmazcan, Ö.; Binay, B. Optimisation of the Production and Bleaching Process for a New Laccase from Madurella mycetomatis, Expressed in Pichia pastoris: From Secretion to Yielding Prominent. Mol. Biotechnol. 2021, 63, 24–39. [Google Scholar] [CrossRef]
- Sen, A.; Kapila, R.; Chaudhary, S.; Nigam, A. Biotechnological Applications of Microbial Enzymes to Replace Chemicals in the Textile Industry—A Review. J. Text. Assoc. 2021, 8/22, 68–73. [Google Scholar]
- Bussler, L.; Jacomini, D.; Corrêa, J.M.; Kadowaki, M.K.; Maller, A.; de Simão, R.C.G. Recombinant Cellulase of Caulobacter crescentus: Potential Applications for Biofuels and Textile Industries. Cellulose 2021, 28, 2813–2832. [Google Scholar] [CrossRef]
- Sondhi, S.; Kumar, D.; Angural, S.; Sharma, P.; Gupta, N. Enzymatic Approach for Bioremediation of Effluent from Pulp and Paper Industry by Thermo Alkali Stable Laccase from Bacillus tequilensis SN4. J. Commer. Biotechnol. 2017, 23, 12–21. [Google Scholar] [CrossRef]
- Shindhal, T.; Rakholiya, P.; Varjani, S.; Pandey, A.; Ngo, H.H.; Guo, W.; Ng, H.Y.; Taherzadeh, M.J. A Critical Review on Advances in the Practices and Perspectives for the Treatment of Dye Industry Wastewater. Bioengineered 2021, 12, 70–87. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Moloantoa, K.M.; Khetsha, Z.P. The Biobleaching Potential of Laccase Produced from Mandarin peelings: Impetus for a Circular Bio-Based Economy in Textile Biofinishing. Arab. J. Chem. 2022, 15, 104305. [Google Scholar] [CrossRef]
- Nathan, V.K. Application of Extremozymes in the Paper and Pulp Industries. In Extremozymes and Their Industrial Applications; Elsevier: Amsterdam, The Netherlands, 2022; pp. 231–247. [Google Scholar]
- Bajpai, P. Basic Overview of Pulp and Paper Manufacturing Process. In Green Chemistry and Sustainability in Pulp and Paper Industry; Springer International Publishing: Cham, Switzerland, 2015; pp. 11–39. [Google Scholar]
- Singh, S.; Singh, V.K.; Aamir, M.; Dubey, M.K.; Patel, J.S.; Upadhyay, R.S.; Gupta, V.K. Cellulase in Pulp and Paper Industry. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2016; pp. 152–162. [Google Scholar]
- Bajaj, P.; Mahajan, R. Cellulase and Xylanase Synergism in Industrial Biotechnology. Appl. Microbiol. Biotechnol. 2019, 103, 8711–8724. [Google Scholar] [CrossRef]
- Gupta, G.K.; Dixit, M.; Kapoor, R.K.; Shukla, P. Xylanolytic Enzymes in Pulp and Paper Industry: New Technologies and Perspectives. Mol. Biotechnol. 2022, 64, 130–143. [Google Scholar] [CrossRef]
- Kumar, A.; Prerna; Bilal, M.; Singh, A.K.; Ratna, S.; Talluri Rameshwari, K.R.; Ahmed, I.; Iqbal, H.M.N. Enzyme Cocktail: A Greener Approach for Biobleaching in Paper and Pulp Industry. In Nanotechnology in Paper and Wood Engineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 303–328. [Google Scholar]
- Sharma, A.; Balda, S.; Gupta, N.; Capalash, N.; Sharma, P. Enzyme Cocktail: An Opportunity for Greener Agro-Pulp Biobleaching in Paper Industry. J. Clean. Prod. 2020, 271, 122573. [Google Scholar] [CrossRef]
- Hasanin, M.S.; Hashem, A.H.; Abd El-Sayed, E.S.; El-Saied, H. Green Ecofriendly Bio-Deinking of Mixed Office Waste Paper Using Various Enzymes from Rhizopus microsporus AH3: Efficiency and Characteristics. Cellulose 2020, 27, 4443–4453. [Google Scholar] [CrossRef]
- Kumar, M.; Campbell, L.; Turner, S. Secondary Cell Walls: Biosynthesis and Manipulation. J. Exp. Bot. 2016, 67, 515–531. [Google Scholar] [CrossRef]
- Walia, A.; Guleria, S.; Mehta, P.; Chauhan, A.; Parkash, J. Microbial Xylanases and Their Industrial Application in Pulp and Paper Biobleaching: A Review. 3 Biotech 2017, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Bilal, M.; Rafeeq, H.; Jabeen, Z.; Afsheen, N.; Sher, F.; Kumar, V.; Bharagava, R.N.; Ferreira, L.F.R.; Iqbal, H.M.N. Role of Laccase in the Pulp and Paper Industry. In Nanotechnology in Paper and Wood Engineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 35–60. [Google Scholar]
- Bhagat, D.D.; Dudhagara, P.R.; Desai, P.V. Statistical approach for pectinase production by Bacillus firmus sdb9 and evaluation of pectino-xylanolytic enzymes for pretreatment of kraft pulp. J. Microbiol. Biotechnol. Food Sci. 2016, 5, 396–406. [Google Scholar] [CrossRef] [Green Version]
- Angural, S.; Bala, I.; Kumar, A.; Kumar, D.; Jassal, S.; Gupta, N. Bleach Enhancement of Mixed Wood Pulp by Mixture of Thermo-Alkali-Stable Xylanase and Mannanase Derived through Co-Culturing of Alkalophilic Bacillus sp. NG-27 and Bacillus nealsonii PN-11. Heliyon 2021, 7, e05673. [Google Scholar] [CrossRef] [PubMed]
- Nagar, S.; Gupta, V.K. Hyper Production and Eco-Friendly Bleaching of Kraft Pulp by Xylanase from Bacillus pumilus SV-205 Using Agro Waste Material. Waste Biomass Valorization 2021, 12, 4019–4031. [Google Scholar] [CrossRef]
- Orozco Colonia, B.S.; Lorenci Woiciechowski, A.; Malanski, R.; Junior Letti, L.A.; Soccol, C.R. Pulp Improvement of Oil Palm Empty Fruit Bunches Associated to Solid-State Biopulping and Biobleaching with Xylanase and Lignin Peroxidase Cocktail Produced by Aspergillus sp. LPB-5. Bioresour. Technol. 2019, 285, 121361. [Google Scholar] [CrossRef] [PubMed]
- Hayes, C.J.; Burgess, D.R.; Manion, J.A. Combustion Pathways of Biofuel Model Compounds. In Advances in Physical Organic Chemistry; Elsevier: Amsterdam, The Netherlands, 2015; pp. 103–187. [Google Scholar]
- Saladini, F.; Patrizi, N.; Pulselli, F.M.; Marchettini, N.; Bastianoni, S. Guidelines for Emergy Evaluation of First, Second and Third Generation Biofuels. Renew. Sustain. Energy Rev. 2016, 66, 221–227. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Kim, S.-H.; Yoon, J.-J.; Yang, Y.-H. Current Status and Strategies for Second Generation Biofuel Production Using Microbial Systems. Energy Convers. Manag. 2017, 148, 1142–1156. [Google Scholar] [CrossRef]
- Merklein, K.; Fong, S.S.; Deng, Y. Biomass Utilization. In Biotechnology for Biofuel Production and Optimization; Elsevier: Amsterdam, The Netherlands, 2016; pp. 291–324. [Google Scholar]
- Saravanan, A.; Senthil Kumar, P.; Jeevanantham, S.; Karishma, S.; Vo, D.-V.N. Recent Advances and Sustainable Development of Biofuels Production from Lignocellulosic Biomass. Bioresour. Technol. 2022, 344, 126203. [Google Scholar] [CrossRef]
- Kumar, B.; Bhardwaj, N.; Agrawal, K.; Chaturvedi, V.; Verma, P. Current Perspective on Pretreatment Technologies Using Lignocellulosic Biomass: An Emerging Biorefinery Concept. Fuel Process. Technol. 2020, 199, 106244. [Google Scholar] [CrossRef]
- Chauhan, A.K. Biofuel: Types and Process Overview. In Nanomaterials in Biofuels Research; Srivastava, M., Srivastava, N., Mishra, P., Gupta, V., Eds.; Clean Energy Production Technologies Book Series; Springer: Singapore, 2020; pp. 1–28. [Google Scholar] [CrossRef]
- Bangaru, A.; Saree, K.A.; Kruthiventi, C.; Banala, M.; Shreya, V.; Vineetha, Y.; Shalini, A.; Mishra, B.; Yadavalli, R.; Chandrasekhar, K.; et al. Role of Enzymes in Biofuel Production: Recent Developments and Challenges. In Bio-Clean Energy Technologies; Chowdhary, P., Ed.; Springer Nature Singapore Pte Ltd.: Singapore, 2022; Volume 1, pp. 81–112. [Google Scholar] [CrossRef]
- Mazarei, M.; Baxter, H.L.; Srivastava, A.; Li, G.; Xie, H.; Dumitrache, A.; Rodriguez, M.; Natzke, J.M.; Zhang, J.-Y.; Turner, G.B.; et al. Silencing Folylpolyglutamate Synthetase1 (FPGS1) in Switchgrass (Panicum virgatum L.) Improves Lignocellulosic Biofuel Production. Front. Plant Sci. 2020, 11, 843. [Google Scholar] [CrossRef]
- Lee, J.H.; Won, H.J.; Hoang Nguyen Tran, P.; Lee, S.; Kim, H.; Jung, J.H. Improving Lignocellulosic Biofuel Production by CRISPR/Cas9-mediated Lignin Modification in Barley. GCB Bioenergy 2021, 13, 742–752. [Google Scholar] [CrossRef]
- Bilal, M.; Iqbal, H.M.N.; Hu, H.; Wang, W.; Zhang, X. Metabolic Engineering and Enzyme-Mediated Processing: A Biotechnological Venture towards Biofuel Production—A Review. Renew. Sustain. Energy Rev. 2018, 82, 436–447. [Google Scholar] [CrossRef]
- Choi, K.R.; Jiao, S.; Lee, S.Y. Metabolic Engineering Strategies toward Production of Biofuels. Curr. Opin. Chem. Biol. 2020, 59, 1–14. [Google Scholar] [CrossRef]
- Banu, J.R.; Kumar, G.; Chattopadhyay, I. Management of Microbial Enzymes for Biofuels and Biogas Production by Using Metagenomic and Genome Editing Approaches. 3 Biotech 2021, 11, 429. [Google Scholar] [CrossRef]
- Lopez-Hidalgo, A.M.; Magaña, G.; Rodriguez, F.; de Leon-Rodriguez, A.; Sanchez, A. Co-Production of Ethanol-Hydrogen by Genetically Engineered Escherichia coli in Sustainable Biorefineries for Lignocellulosic Ethanol Production. Chem. Eng. J. 2021, 406, 126829. [Google Scholar] [CrossRef]
- Singhania, R.R.; Ruiz, H.A.; Awasthi, M.K.; Dong, C.-D.; Chen, C.-W.; Patel, A.K. Challenges in Cellulase Bioprocess for Biofuel Applications. Renew. Sustain. Energy Rev. 2021, 151, 111622. [Google Scholar] [CrossRef]
- Klein-Marcuschamer, D.; Oleskowicz-Popiel, P.; Simmons, B.A.; Blanch, H.W. The Challenge of Enzyme Cost in the Production of Lignocellulosic Biofuels. Biotechnol. Bioeng. 2012, 109, 1083–1087. [Google Scholar] [CrossRef]
- Damásio, A.R.L.; Pessela, B.C.; Segato, F.; Prade, R.A.; Guisan, J.M.; Polizeli, M.L.T.M. Improvement of Fungal Arabinofuranosidase Thermal Stability by Reversible Immobilization. Process Biochem. 2012, 47, 2411–2417. [Google Scholar] [CrossRef]
- Rajnish, K.N.; Samuel, M.S.; John J, A.; Datta, S.; Chandrasekar, N.; Balaji, R.; Jose, S.; Selvarajan, E. Immobilization of Cellulase Enzymes on Nano and Micro-Materials for Breakdown of Cellulose for Biofuel Production-a Narrative Review. Int. J. Biol. Macromol. 2021, 182, 1793–1802. [Google Scholar] [CrossRef]
- Kumar, V.; Shahi, S.K.; Singh, S. Bioremediation: An Eco-Sustainable Approach for Restoration of Contaminated Sites. In Microbial Bioprospecting for Sustainable Development; Springer: Singapore, 2018; pp. 115–136. [Google Scholar]
- Li, Q.; Liu, J.; Gadd, G.M. Fungal Bioremediation of Soil Co-Contaminated with Petroleum Hydrocarbons and Toxic Metals. Appl. Microbiol. Biotechnol. 2020, 104, 8999–9008. [Google Scholar] [CrossRef]
- Gouma, S.; Fragoeiro, S.; Bastos, A.C.; Magan, N. Bacterial and Fungal Bioremediation Strategies. In Microbial Biodegradation and Bioremediation; Elsevier: Amsterdam, The Netherlands, 2014; pp. 301–323. [Google Scholar]
- Bilal, M.; Iqbal, H.M.N.; Barceló, D. Persistence of Pesticides-Based Contaminants in the Environment and Their Effective Degradation Using Laccase-Assisted Biocatalytic Systems. Sci. Total Environ. 2019, 695, 133896. [Google Scholar] [CrossRef]
- Bilal, M.; Iqbal, H.M.N.; Barceló, D. Mitigation of Bisphenol A Using an Array of Laccase-Based Robust Bio-Catalytic Cues—A Review. Sci. Total Environ. 2019, 689, 160–177. [Google Scholar] [CrossRef] [PubMed]
- Steinbuchel, A. Laccases in Bioremediation and Waste Valorisation; Schlosser, D., Ed.; Springer International Publishing: Cham, Switzerland, 2020; Volume 33, ISBN 978-3-030-47905-3. [Google Scholar]
- Zhuo, R.; Fan, F. A Comprehensive Insight into the Application of White Rot Fungi and Their Lignocellulolytic Enzymes in the Removal of Organic Pollutants. Sci. Total Environ. 2021, 778, 146132. [Google Scholar] [CrossRef] [PubMed]
- Beltrán-Flores, E.; Torán, J.; Caminal, G.; Blánquez, P.; Sarrà, M. The Removal of Diuron from Agricultural Wastewaters by Trametes versicolor Immobilized on Pinewood in Simple Channel Reactors. Sci. Total Environ. 2020, 728, 138414. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Peris, A.; Torán, J.; Eljarrat, E.; Sarrà, M.; Blánquez, P.; Caminal, G. Exploring the Degradation Capability of Trametes versicolor on Selected Hydrophobic Pesticides through Setting Sights Simultaneously on Culture Broth and Biological Matrix. Chemosphere 2020, 250, 126293. [Google Scholar] [CrossRef]
- Mir-Tutusaus, J.A.; Caminal, G.; Sarrà, M. Influence of Process Variables in a Continuous Treatment of Non-Sterile Hospital Wastewater by Trametes versicolor and Novel Method for Inoculum Production. J. Environ. Manag. 2018, 212, 415–423. [Google Scholar] [CrossRef]
- Cruz del Álamo, A.; Pariente, M.I.; Martínez, F.; Molina, R. Trametes versicolor Immobilized on Rotating Biological Contactors as Alternative Biological Treatment for the Removal of Emerging Concern Micropollutants. Water Res. 2020, 170, 115313. [Google Scholar] [CrossRef]
- Brugnari, T.; Pereira, M.G.; Bubna, G.A.; de Freitas, E.N.; Contato, A.G.; Corrêa, R.C.G.; Castoldi, R.; de Souza, C.G.M.; Polizeli, M.L.T.M.; Bracht, A.; et al. A Highly Reusable MANAE-Agarose-Immobilized Pleurotus ostreatus Laccase for Degradation of Bisphenol A. Sci. Total Environ. 2018, 634, 1346–1351. [Google Scholar] [CrossRef]
- Hu, C.; Huang, D.; Zeng, G.; Cheng, M.; Gong, X.; Wang, R.; Xue, W.; Hu, Z.; Liu, Y. The Combination of Fenton Process and Phanerochaete chrysosporium for the Removal of Bisphenol A in River Sediments: Mechanism Related to Extracellular Enzyme, Organic Acid and Iron. Chem. Eng. J. 2018, 338, 432–439. [Google Scholar] [CrossRef]
- Jaén-Gil, A.; Castellet-Rovira, F.; Llorca, M.; Villagrasa, M.; Sarrà, M.; Rodríguez-Mozaz, S.; Barceló, D. Fungal Treatment of Metoprolol and Its Recalcitrant Metabolite Metoprolol Acid in Hospital Wastewater: Biotransformation, Sorption and Ecotoxicological Impact. Water Res. 2019, 152, 171–180. [Google Scholar] [CrossRef]
- Mohammadi-Sichani, M.; Mazaheri Assadi, M.; Farazmand, A.; Kianirad, M.; Ahadi, A.M.; Hadian-Ghahderijani, H. Ability of Agaricus bisporus, Pleurotus ostreatus and Ganoderma lucidum Compost in Biodegradation of Petroleum Hydrocarbon-Contaminated Soil. Int. J. Environ. Sci. Technol. 2019, 16, 2313–2320. [Google Scholar] [CrossRef]
- Wang, X.; Qin, X.; Hao, Z.; Luo, H.; Yao, B.; Su, X. Degradation of Four Major Mycotoxins by Eight Manganese Peroxidases in Presence of a Dicarboxylic Acid. Toxins 2019, 11, 566. [Google Scholar] [CrossRef] [Green Version]
- Lothe, A.G.; Kalra, S.S.; Wang, M.; Mack, E.E.; Walecka-Hutchison, C.; Kickhoefer, V.A.; Rome, L.H.; Mahendra, S. Vault Packaged Enzyme Mediated Degradation of Amino-Aromatic Energetic Compounds. Chemosphere 2020, 242, 125117. [Google Scholar] [CrossRef]
- Mateljak, I.; Gomez-Fernandez, B.; Alcalde, M. Laccase Engineering by Directed and Computational Evolution. In Laccases in Bioremediation and Waste Valorisation; Microbiology Monographs; Schlosser, D., Ed.; Springer: Cham, Switzerland, 2020; Volume 33, pp. 191–212. [Google Scholar] [CrossRef]
- Lima, M.S.; de Lucas, R.C. Co-Cultivation, Co-Culture, Mixed Culture, and Microbial Consortium of Fungi: An Understudied Strategy for Biomass Conversion. Front. Microbiol. 2022, 12, 837685. [Google Scholar] [CrossRef]
- Lodha, A.; Pawar, S.; Rathod, V. Optimised Cellulase Production from Fungal Co-Culture of Trichoderma reesei NCIM 1186 and Penicillium citrinum NCIM 768 under Solid State Fermentation. J. Environ. Chem. Eng. 2020, 8, 103958. [Google Scholar] [CrossRef]
- Dasari, P.R.; Ramteke, P.W.; Kesri, S.; Kongala, P.R. Comparative Study of Cellulase Production Using Submerged and Solid-State Fermentation. In Approaches to Enhance Industrial Production of Fungal Cellulases. Fungal Biology; Srivastava, M., Srivastava, N., Ramteke, P., Mishra, P., Eds.; Springer: Cham, Switzerland, 2019; pp. 37–52. [Google Scholar] [CrossRef]
- Sperandio, G.B.; Ferreira Filho, E.X. Fungal Co-Cultures in the Lignocellulosic Biorefinery Context: A Review. Int. Biodeterior. Biodegrad. 2019, 142, 109–123. [Google Scholar] [CrossRef]
- Preda, D.; Popa, D.-G.; Constantinescu-Aruxandei, D.; Oancea, F. Enhancement of Lignolytic Enzyme Activity in Ganoderma lucidum by Co-Cultivation with Bacteria. Chem. Proc. 2021, 7, 7035. [Google Scholar]
- Singh, N.; Gupta, R.P.; Puri, S.K.; Mathur, A.S. Bioethanol Production from Pretreated Whole Slurry Rice Straw by Thermophilic Co-Culture. Fuel 2021, 301, 121074. [Google Scholar] [CrossRef]
- Rabello, G.C.; Pirota, R.D.P.B.; Barros, G.O.F.; Farinas, C.S. Simultaneous Biosynthesis of Biomass-Degrading Enzymes Using Co-Cultivation of Aspergillus niger and Trichoderma reesei. Biocatal. Biotransform. 2014, 32, 236–243. [Google Scholar] [CrossRef]
- Zhao, C.; Deng, L.; Fang, H. Mixed Culture of Recombinant Trichoderma reesei and Aspergillus niger for Cellulase Production to Increase the Cellulose Degrading Capability. Biomass Bioenergy 2018, 112, 93–98. [Google Scholar] [CrossRef]
- Sperandio, G.B.; Filho, E.X.F. An Overview of Trichoderma reesei Co-Cultures for the Production of Lignocellulolytic Enzymes. Appl. Microbiol. Biotechnol. 2021, 105, 3019–3025. [Google Scholar] [CrossRef]
- Campioni, T.S.; de Azevedo Carvalho, A.F.; de Figueiredo, F.C.; da Silva, D.F.; de Oliva Neto, P. Xylanases and Cellulases Biosynthesis by Selected Fungi in a Simple and Economic Bio System Using Sugarcane Straw. Int. J. Environ. Agric. Biotechnol. 2020, 5, 217–230. [Google Scholar] [CrossRef]
- Fatma, S.; Saleem, A.; Tabassum, R. Wheat Straw Hydrolysis by Using Co-Cultures of Trichoderma reesei and Monascus purpureus toward Enhanced Biodegradation of the Lignocellulosic Biomass in Bioethanol Biorefinery. Biomass Convers. Biorefin. 2021, 11, 743–754. [Google Scholar] [CrossRef]
- Silva, Y.H.; Oliveira, T.B.; Lima, M.S.; Pasin, T.M.; de Almeida Scarcella, A.S.; Polizeli, M.L.T.M.; Martinez, C.A.; Buckeridge, M.S.; Santos, W.D.; Lucas, R.C. Co-Culture of Trichoderma reesei, Talaromyces sp. and Aspergillus spp. Produces A Multi-Enzyme Cocktail for the Hydrolysis of Sugarcane Bagasse Pretreated with Piperonilic Acid (PIP) and Methylenedioxycinnamic Acid (MDCA). Res. Sq. 2022. [Google Scholar] [CrossRef]
- Karuppiah, V.; Zhixiang, L.; Liu, H.; Murugappan, V.; Kumaran, S.; Perianaika Anahas, A.M.; Chen, J. Co-Cultivation of T. asperellum GDFS1009 and B. amyloliquefaciens 1841: Strategy to Regulate the Production of Ligno-Cellulolytic Enzymes for the Lignocellulose Biomass Degradation. J. Environ. Manag. 2022, 301, 113833. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Jha, P.K.; Panwar, A. Production of Bioethanol from Wheat Straw via Optimization of Co-Culture Conditions of Bacillus licheniformis and Saccharomyces cerevisiae. Discov. Energy 2021, 1, 5. [Google Scholar] [CrossRef]
- Phillips, E. Genetic Engineering Applications to Improve Cellulase Production and Efficiency: Part I. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 209–225. [Google Scholar]
- Jiang, X.; Du, J.; He, R.; Zhang, Z.; Qi, F.; Huang, J.; Qin, L. Improved Production of Majority Cellulases in Trichoderma reesei by Integration of Cbh1 Gene from Chaetomium thermophilum. Front. Microbiol. 2020, 11, 1633. [Google Scholar] [CrossRef]
- Brunecky, R.; Subramanian, V.; Yarbrough, J.M.; Donohoe, B.S.; Vinzant, T.B.; Vanderwall, T.A.; Knott, B.C.; Chaudhari, Y.B.; Bomble, Y.J.; Himmel, M.E.; et al. Synthetic Fungal Multifunctional Cellulases for Enhanced Biomass Conversion. Green Chem. 2020, 22, 478–489. [Google Scholar] [CrossRef] [Green Version]
- Ajeje, S.B.; Hu, Y.; Song, G.; Peter, S.B.; Afful, R.G.; Sun, F.; Asadollahi, M.A.; Amiri, H.; Abdulkhani, A.; Sun, H. Thermostable Cellulases/Xylanases from Thermophilic and Hyperthermophilic Microorganisms: Current Perspective. Front. Bioeng. Biotechnol. 2021, 9, 794304. [Google Scholar] [CrossRef]
- Xing, H.; Zou, G.; Liu, C.; Chai, S.; Yan, X.; Li, X.; Liu, R.; Yang, Y.; Zhou, Z. Improving the Thermostability of a GH11 Xylanase by Directed Evolution and Rational Design Guided by B-Factor Analysis. Enzym. Microb Technol 2021, 143, 109720. [Google Scholar] [CrossRef]
- Tang, F.; Chen, D.; Yu, B.; Luo, Y.; Zheng, P.; Mao, X.; Yu, J.; He, J. Improving the Thermostability of Trichoderma reesei Xylanase 2 by Introducing Disulfide Bonds. Electron. J. Biotechnol. 2017, 26, 52–59. [Google Scholar] [CrossRef]
- Wang, Q.; Zhong, C.; Xiao, H. Genetic Engineering of Filamentous Fungi for Efficient Protein Expression and Secretion. Front. Bioeng. Biotechnol. 2020, 8, 293. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yao, G.; Wu, R.; Gao, L.; Kan, Q.; Liu, M.; Yang, P.; Liu, G.; Qin, Y.; Song, X.; et al. Synergistic and Dose-Controlled Regulation of Cellulase Gene Expression in Penicillium oxalicum. PLoS Genet. 2015, 11, e1005509. [Google Scholar] [CrossRef]
- Alazi, E.; Niu, J.; Kowalczyk, J.E.; Peng, M.; Aguilar Pontes, M.V.; Kan, J.A.L.; Visser, J.; Vries, R.P.; Ram, A.F.J. The Transcriptional Activator GaaR of Aspergillus niger Is Required for Release and Utilization of D-Galacturonic Acid from Pectin. FEBS Lett. 2016, 590, 1804–1815. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Li, Z.; Xia, C.; Qu, Y.; Liu, M.; Yang, P.; Yu, L.; Song, X. Combining Manipulation of Transcription Factors and Overexpression of the Target Genes to Enhance Lignocellulolytic Enzyme Production in Penicillium oxalicum. Biotechnol. Biofuels 2017, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Xu, Y.; Song, X.; Li, S.; Xia, C.; Xu, J.; Qin, Y.; Liu, G.; Qu, Y. Deletion of the Middle Region of the Transcription Factor ClrB in Penicillium oxalicum Enables Cellulase Production in the Presence of Glucose. J. Biol. Chem. 2019, 294, 18685–18697. [Google Scholar] [CrossRef]
- Peng, M.; Khosravi, C.; Lubbers, R.J.M.; Kun, R.S.; Aguilar Pontes, M.V.; Battaglia, E.; Chen, C.; Dalhuijsen, S.; Daly, P.; Lipzen, A.; et al. CreA-Mediated Repression of Gene Expression Occurs at Low Monosaccharide Levels during Fungal Plant Biomass Conversion in a Time and Substrate Dependent Manner. Cell Surf. 2021, 7, 100050. [Google Scholar] [CrossRef]
- Huberman, L.B.; Liu, J.; Qin, L.; Glass, N.L. Regulation of the Lignocellulolytic Response in Filamentous Fungi. Fungal Biol. Rev. 2016, 30, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, L.M.; Parreiras, L.S.; Murakami, M.T. Rational Engineering of the Trichoderma reesei RUT-C30 Strain into an Industrially Relevant Platform for Cellulase Production. Biotechnol. Biofuels 2020, 13, 93. [Google Scholar] [CrossRef]
- Luo, Y.; Valkonen, M.; Jackson, R.E.; Palmer, J.M.; Bhalla, A.; Nikolaev, I.; Saloheimo, M.; Ward, M. Modification of Transcriptional Factor ACE3 Enhances Protein Production in Trichoderma reesei in the Absence of Cellulase Gene Inducer. Biotechnol. Biofuels 2020, 13, 137. [Google Scholar] [CrossRef]
- Gao, L.; He, X.; Guo, Y.; Wu, Z.; Zhao, J.; Liu, G.; Qu, Y. Combinatorial Engineering of Transcriptional Activators in Penicillium oxalicum for Improved Production of Corn-Fiber-Degrading Enzymes. J. Agric. Food Chem. 2021, 69, 2539–2548. [Google Scholar] [CrossRef]
- Qian, Y.; Zhong, L.; Sun, Y.; Sun, N.; Zhang, L.; Liu, W.; Qu, Y.; Zhong, Y. Enhancement of Cellulase Production in Trichoderma reesei via Disruption of Multiple Protease Genes Identified by Comparative Secretomics. Front. Microbiol. 2019, 10, 2784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derntl, C.; Guzmán-Chávez, F.; Mello-de-Sousa, T.M.; Busse, H.-J.; Driessen, A.J.M.; Mach, R.L.; Mach-Aigner, A.R. In Vivo Study of the Sorbicillinoid Gene Cluster in Trichoderma reesei. Front. Microbiol. 2017, 8, 2037. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Mannan, A.A.; Han, Y.; Oyarzún, D.A.; Zhang, F. Dynamic Metabolic Control: Towards Precision Engineering of Metabolism. J. Ind. Microbiol. Biotechnol. 2018, 45, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Lemmerer, M.; Mairhofer, J.; Lepak, A.; Longus, K.; Hahn, R.; Nidetzky, B. Decoupling of Recombinant Protein Production from Escherichia coli Cell Growth Enhances Functional Expression of Plant Leloir Glycosyltransferases. Biotechnol. Bioeng. 2019, 116, 1259–1268. [Google Scholar] [CrossRef] [Green Version]
- Bodie, E.; Virag, A.; Pratt, R.J.; Leiva, N.; Ward, M.; Dodge, T. Reduced Viscosity Mutants of Trichoderma reesei with Improved Industrial Fermentation Characteristics. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab014. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, Q.; Wang, Q.; Qin, Y.; Zhong, Y.; Gao, L.; Liu, G.; Qu, Y. Disruption of the Trichoderma reesei Gul1 Gene Stimulates Hyphal Branching and Reduces Broth Viscosity in Cellulase Production. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab012. [Google Scholar] [CrossRef]
- Kunamneni, A. Cloning and Recombinant Expression of a Cellulase. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2016; pp. 99–106. [Google Scholar]
- Shahzadi, I.; Al-Ghamdi, M.A.; Nadeem, M.S.; Sajjad, M.; Ali, A.; Khan, J.A.; Kazmi, I. Scale-up Fermentation of Escherichia coli for the Production of Recombinant Endoglucanase from Clostridium thermocellum. Sci. Rep. 2021, 11, 7145. [Google Scholar] [CrossRef]
- Kiribayeva, A.; Mukanov, B.; Silayev, D.; Akishev, Z.; Ramankulov, Y.; Khassenov, B. Cloning, Expression, and Characterization of a Recombinant Xylanase from Bacillus sonorensis T6. PLoS ONE 2022, 17, e0265647. [Google Scholar] [CrossRef]
- Liu, P.; Wu, B.; Chen, M.; Dai, Y.; Song, C.; Sun, L.; Huang, Y.; Li, S.; Hu, G.; He, M. Enhancing Secretion of Endoglucanase in Zymomonas mobilis by Disturbing Peptidoglycan Synthesis. Appl. Environ. Microbiol. 2022, 88, e02161-21. [Google Scholar] [CrossRef]
- Saleem, A.; Waris, S.; Ahmed, T.; Tabassum, R. Biochemical Characterization and Molecular Docking of Cloned Xylanase Gene from Bacillus subtilis RTS Expressed in E. coli. Int. J. Biol. Macromol. 2021, 168, 310–321. [Google Scholar] [CrossRef]
- Vadala, B.S.; Deshpande, S.; Apte-Deshpande, A. Soluble Expression of Recombinant Active Cellulase in E. coli Using B. subtilis (Natto Strain) Cellulase Gene. J. Genet. Eng. Biotechnol. 2021, 19, 7. [Google Scholar] [CrossRef]
- Yan, Z.; Zhang, J.; Bao, J. Increasing Cellulosic Ethanol Production by Enhancing Phenolic Tolerance of Zymomonas mobilis in Adaptive Evolution. Bioresour. Technol. 2021, 329, 124926. [Google Scholar] [CrossRef]
- Chetty, B.J.; Inokuma, K.; Hasunuma, T.; van Zyl, W.H.; den Haan, R. Improvement of Cell-Tethered Cellulase Activity in Recombinant Strains of Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2022, 106, 6347–6361. [Google Scholar] [CrossRef]
- Adegboye, M.F.; Ojuederie, O.B.; Talia, P.M.; Babalola, O.O. Bioprospecting of Microbial Strains for Biofuel Production: Metabolic Engineering, Applications, and Challenges. Biotechnol. Biofuels 2021, 14, 5. [Google Scholar] [CrossRef]
- Sarkar, D.; Hansdah, R.; Kar, A.; Sarkar, A. Microbial Bioprospecting in Development of Integrated Biomass Based Biorefineries. In Bioprospecting of Microbial Diversity; Elsevier: Amsterdam, The Netherlands, 2022; pp. 257–275. [Google Scholar]
- Simon, C.; Daniel, R. Metagenomic Analyses: Past and Future Trends. Appl. Environ. Microbiol. 2011, 77, 1153–1161. [Google Scholar] [CrossRef] [Green Version]
- Wani, A.K.; Rahayu, F.; Kadarwati, F.T.; Suhara, C.; Singh, R.; Dhanjal, D.S.; Akhtar, N.; Mir, T.G.; Chopra, C. Metagenomic Screening Strategies for Bioprospecting Enzymes from Environmental Samples. IOP Conf. Ser. Earth Environ. Sci. 2022, 974, 012003. [Google Scholar] [CrossRef]
- Pabbathi, N.P.P.; Velidandi, A.; Tavarna, T.; Gupta, S.; Raj, R.S.; Gandam, P.K.; Baadhe, R.R. Role of Metagenomics in Prospecting Novel Endoglucanases, Accentuating Functional Metagenomics Approach in Second-Generation Biofuel Production: A Review. Biomass Convers. Biorefin. 2021. [Google Scholar] [CrossRef]
- Batista-García, R.A.; del Rayo Sánchez-Carbente, M.; Talia, P.; Jackson, S.A.; O’Leary, N.D.; Dobson, A.D.W.; Folch-Mallol, J.L. From Lignocellulosic Metagenomes to Lignocellulolytic Genes: Trends, Challenges and Future Prospects. Biofuels Bioprod. Biorefining 2016, 10, 864–882. [Google Scholar] [CrossRef]
- Escuder-Rodríguez, J.-J.; DeCastro, M.-E.; Cerdán, M.-E.; Rodríguez-Belmonte, E.; Becerra, M.; González-Siso, M.-I. Cellulases from Thermophiles Found by Metagenomics. Microorganisms 2018, 6, 66. [Google Scholar] [CrossRef]
- Jain, L.; Agrawal, D. Performance Evaluation of Fungal Cellulases with Dilute Acid Pretreated Sugarcane Bagasse: A Robust Bioprospecting Strategy for Biofuel Enzymes. Renew. Energy 2018, 115, 978–988. [Google Scholar] [CrossRef]
- Mezule, L.; Civzele, A. Bioprospecting White-Rot Basidiomycete Irpex lacteus for Improved Extraction of Lignocellulose-Degrading Enzymes and Their Further Application. J. Fungi 2020, 6, 256. [Google Scholar] [CrossRef] [PubMed]
- Georgiadou, D.N.; Avramidis, P.; Ioannou, E.; Hatzinikolaou, D.G. Microbial Bioprospecting for Lignocellulose Degradation at a Unique Greek Environment. Heliyon 2021, 7, e07122. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, M.; Shrivastava, S.; Shukla, P. Bioprospecting of Xylanase Producing Fungal Strains: Multilocus Phylogenetic Analysis and Enzyme Activity Profiling. J. Basic Microbiol. 2022, 62, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Lemos, L.N.; Pereira, R.V.; Quaggio, R.B.; Martins, L.F.; Moura, L.M.S.; da Silva, A.R.; Antunes, L.P.; da Silva, A.M.; Setubal, J.C. Genome-Centric Analysis of a Thermophilic and Cellulolytic Bacterial Consortium Derived from Composting. Front. Microbiol. 2017, 8, 644. [Google Scholar] [CrossRef] [Green Version]
- Jorquera, M.A.; Graether, S.P.; Maruyama, F. Editorial: Bioprospecting and Biotechnology of Extremophiles. Front. Bioeng. Biotechnol. 2019, 7, 204. [Google Scholar] [CrossRef]
- Deshmukh, R.; Jagtap, S. Bioprospecting of Extremophiles for Industrial Enzymes. In Bioprospecting of Microbial Diversity; Elsevier: Amsterdam, The Netherlands, 2022; pp. 471–482. [Google Scholar]
- Fatani, S.; Saito, Y.; Alarawi, M.; Gojobori, T.; Mineta, K. Genome Sequencing and Identification of Cellulase Genes in Bacillus paralicheniformis Strains from the Red Sea. BMC Microbiol. 2021, 21, 254. [Google Scholar] [CrossRef]
- Alves, K.J.; da Silva, M.C.P.; Cotta, S.R.; Ottoni, J.R.; van Elsas, J.D.; Oliveira, V.M.; Andreote, F.D. Mangrove Soil as a Source for Novel Xylanase and Amylase as Determined by Cultivation-Dependent and Cultivation-Independent Methods. Braz. J. Microbiol. 2020, 51, 217–228. [Google Scholar] [CrossRef]



{kind=link}
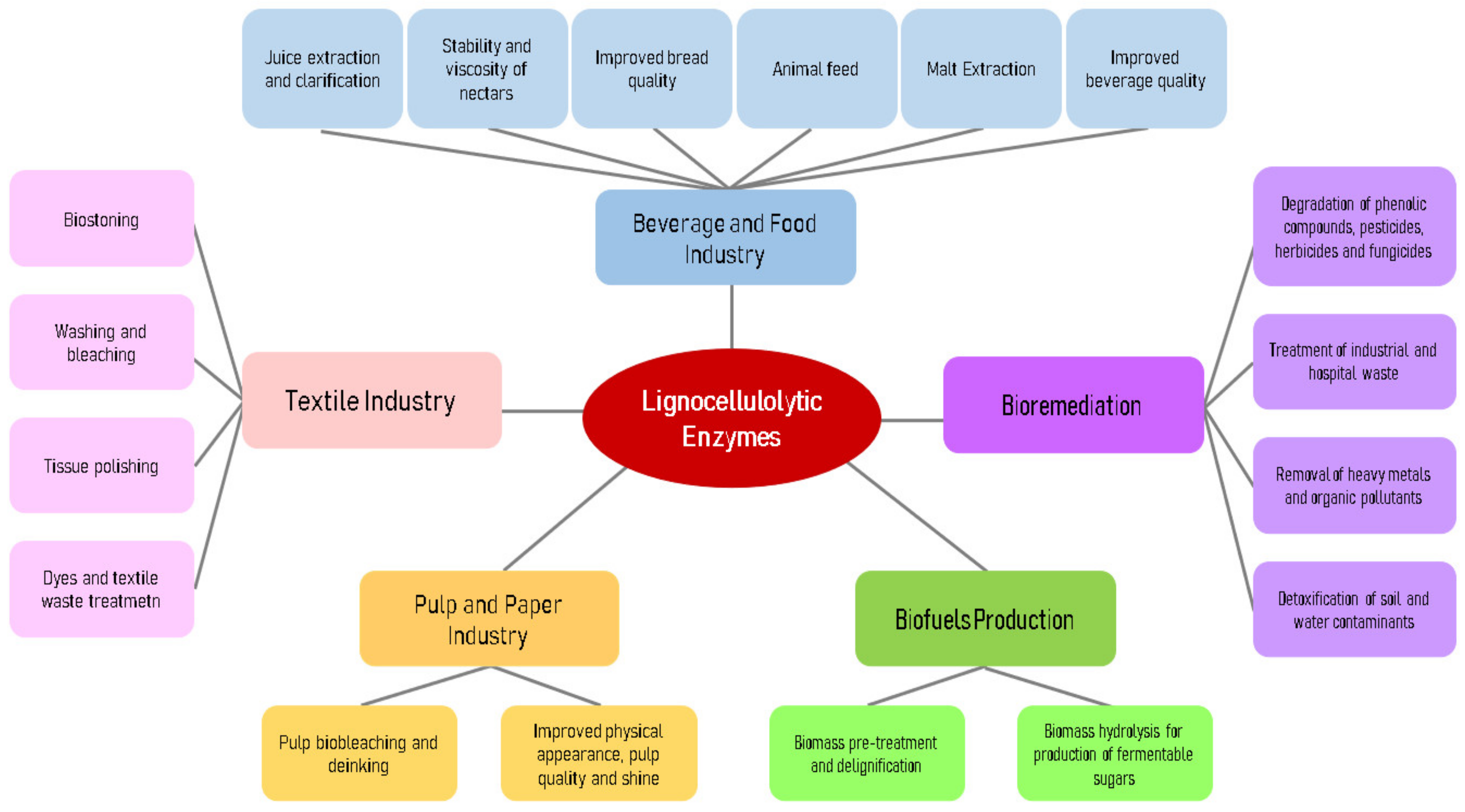{kind=link}
| Enzymes | Microorganism | Optimal pH | Optimal Temperature | Reference |
|---|---|---|---|---|
| Endoglucanases | Bacillus subtilis | 5 | 60 | [129] |
| Neobacillus sedimentimangrovi | 7 | 60 | [130] | |
| Arthrobacter woluwensis | 8 | 50 | [131] | |
| Thermotoga naphtophila | 6 | 90 | [132] | |
| Exoglucanases | Clostridium thermocellum | 5.7 | 70 | [133] |
| Xylanases | Thermotoga marítima TmxB | 5 | 100 | [134] |
| Acinetobacter johnsonii | 6 | 55 | [135] | |
| Bacillus haynesii | 7 | 40 | [136] | |
| Caldicoprobacter algeriensis | 6.5 | 80 | [137] | |
| Peroxidases | Pseudomonas spp. | 3–8 | 20–80 | [138] |
| Bacillus ayderensis SK3-4 | 7 | 75 | [139] | |
| Laccases | Lysinibacillus macroides | 7 | 30 | [140] |
| Pseudomonas parafulva | 8 | 50 | [141] |
| Enzyme | Microorganism | Optimal pH | Optimal Temperature (°C) | Reference |
|---|---|---|---|---|
| Endoglucanases | Trichoderma viride | 5 | 40 | [161] |
| Cladosporium cladosporioides | 4 | 30 | [162] | |
| Fusarium spp. | 5.5 | 30 | [163] | |
| Aspergillus niger | 5.5 | 30 | [164] | |
| Exoglucanases | Trichoderma viride | 5 | 40 | [161] |
| Fusarium spp. | 5.5 | 30 | [163] | |
| Aspergillus niger | 5.5 | 30 | [164] | |
| Phaeolus spadiceus | 4.5 | 25–30 | [165] | |
| β-glycosidases | Trichoderma viride | 5 | 40 | [161] |
| Cladosporium cladosporioides | 4 | 30 | [162] | |
| Aspergillus niger | 5–9 | 25–45 | [166] | |
| Fusarium spp. | 5.5 | 30 | [163] | |
| Xylanases | Trichoderma spp. | 5 | 28 | [167] |
| Trichoderma harzianum | 6 | 70 | [168] | |
| Aspergillus tubingensis | 3–8 | 30–60 | [169] | |
| Talaromyces amestolkiae | 7 | 30 | [170] | |
| Peroxidases | Pleurotus ostreatus | 3.3 | 25 | [171] |
| Hypsizygus ulmarius | 7 | 28 | [172] | |
| Pleurostuus florida | 7 | 28 | [172] | |
| Phlebia radiata | 3 | 80 | [173] | |
| Laccases | Trametes polyzona | 4.5 | 55 | [174] |
| Trametes versicolor | 4–5 | 40–50 | [175] | |
| Coriolopsis gallica | 6–8 | 40–60 | [176] | |
| Pycnoporus spp. | 6 | 0 | [177] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benatti, A.L.T.; Polizeli, M.d.L.T.d.M. Lignocellulolytic Biocatalysts: The Main Players Involved in Multiple Biotechnological Processes for Biomass Valorization. Microorganisms 2023, 11, 162. https://doi.org/10.3390/microorganisms11010162
Benatti ALT, Polizeli MdLTdM. Lignocellulolytic Biocatalysts: The Main Players Involved in Multiple Biotechnological Processes for Biomass Valorization. Microorganisms. 2023; 11(1):162. https://doi.org/10.3390/microorganisms11010162
Chicago/Turabian StyleBenatti, Ana Laura Totti, and Maria de Lourdes Teixeira de Moraes Polizeli. 2023. "Lignocellulolytic Biocatalysts: The Main Players Involved in Multiple Biotechnological Processes for Biomass Valorization" Microorganisms 11, no. 1: 162. https://doi.org/10.3390/microorganisms11010162