
Co-Infection of Potential Tick-Borne Pathogens of the Order Rickettsiales and Borrelia burgdorferi s. l. and Their Link to Season and Area in Germany
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statements
2.2. Nucleic Acid Extracts of Ticks and Pool Composition
2.3. Protein-Coding Gene-Based PCR to Screen for Bacteria of the Order Rickettsiales
2.4. gltA Gene-Based Sequence Analyses
2.5. Statistics
3. Results
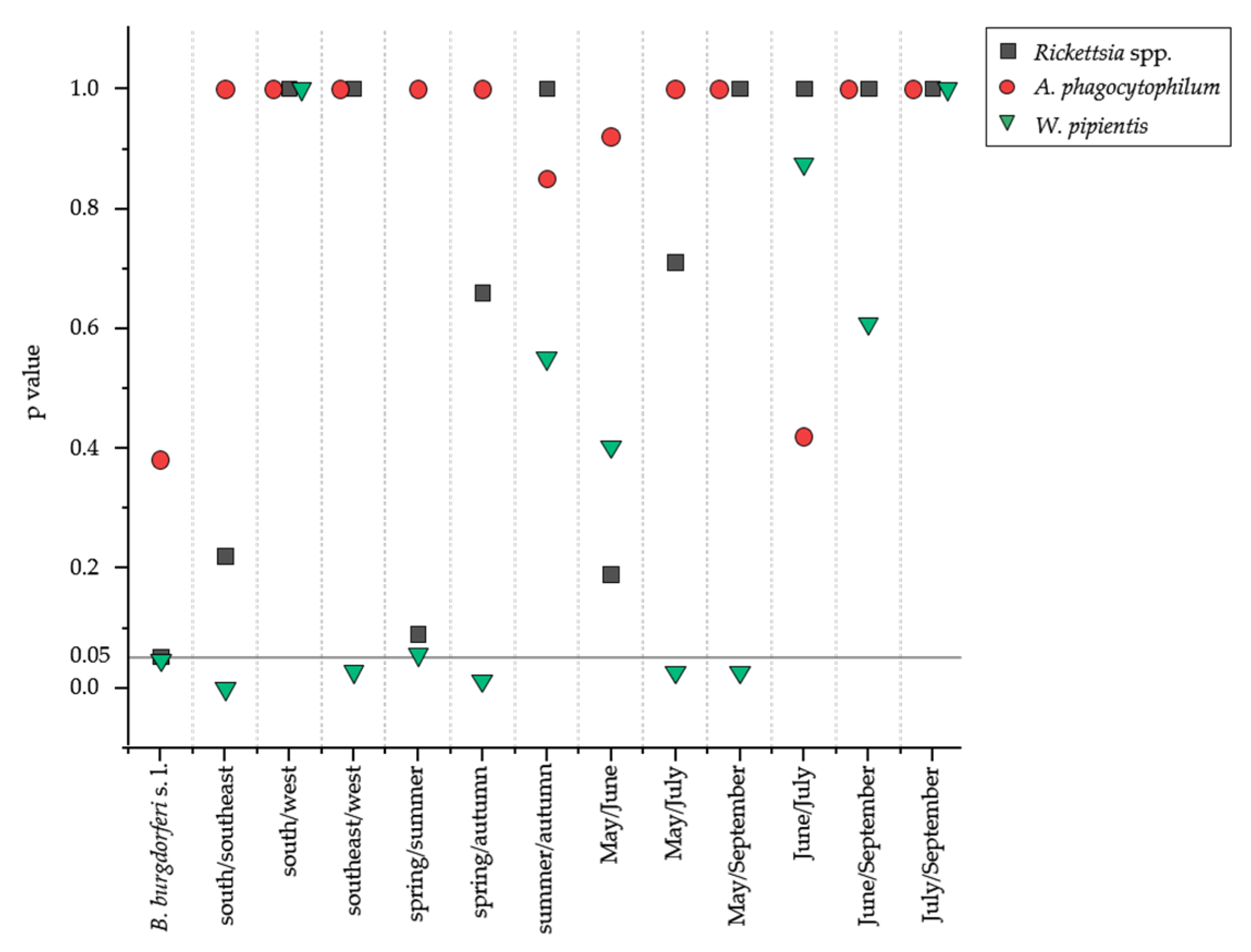
3.1. Rickettsia spp. and W. pipientis Showed Highest Prevalence in Tick Extracts
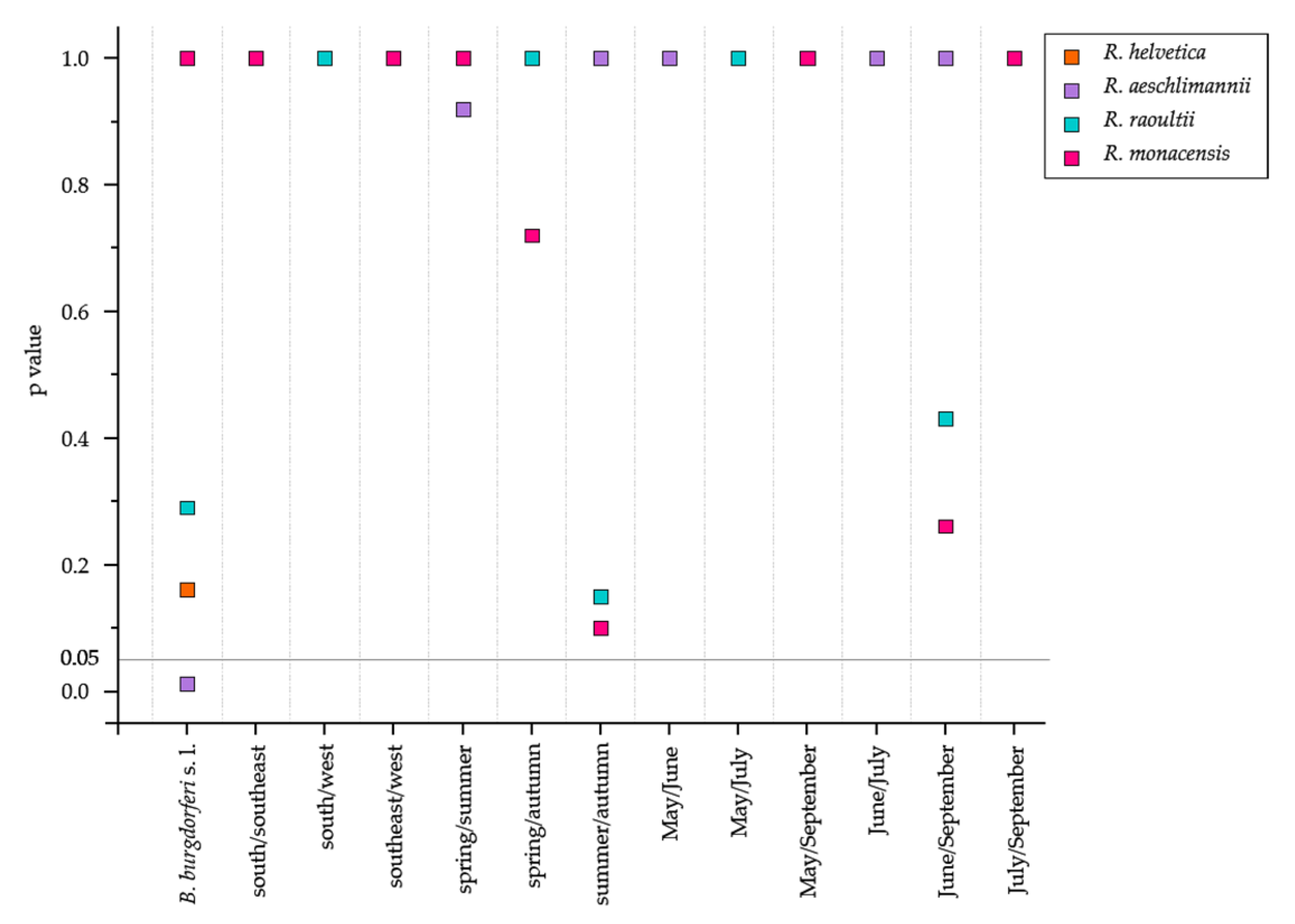
3.2. Rickettsia-Positive Ticks were Dominated by R. helvetica
3.3. Prevalence of Rickettsia Species was Affected by B. burgdorferi s. l. Co-Infection
4. Discussion
4.1. Co-Infection of Potential Tick-Borne Pathogens Are Season Dependent
4.2. Prevalence of R. helvetica to Environmental Variables
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boehnke, D.; Gebhardt, R.; Petney, T.; Norra, S. On the complexity of measuring forests microclimate and interpreting its relevance in habitat ecology: The example of Ixodes ricinus ticks. Parasites Vectors 2017, 10, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobler, G.; Fingerle, V.; Hagedorn, P.; Pfeffer, M.; Silaghi, C.; Tomaso, H.; Henning, K.; Niedrig, M. Gefahren der Übertragung von Krankheitserregern durch Schildzecken in Deutschland. Bundesgesundheitsblatt Gesundh. Gesundh. 2014, 57, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Rubel, F.; Brugger, K.; Chitimia-Dobler, L.; Dautel, H.; Meyer-Kayser, E.; Kahl, O. Atlas of ticks (Acari: Argasidae, Ixodidae) in Germany. Exp. Appl. Acarol. 2021, 84, 183–214. [Google Scholar] [CrossRef] [PubMed]
- Kowalec, M.; Szewczyk, T.; Welc-Falęciak, R.; Siński, E.; Karbowiak, G.; Bajer, A. Rickettsiales occurrence and co-occurrence in Ixodes ricinus ticks in natural and urban areas. Microb. Ecol. 2019, 77, 890–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welc-Falęciak, R.; Kowalec, M.; Karbowiak, G.; Bajer, A.; Behnke, J.M.; Siński, E. Rickettsiaceae and Anaplasmataceae infections in Ixodes ricinus ticks from urban and natural forested areas of Poland. Parasites Vectors 2014, 7, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbowiak, G.; Biernat, B.; Stańczak, J.; Szewczyk, T.; Werszko, J. The role of particular tick developmental stages in the circulation of tick-borne pathogens affecting humans in Central Europe. 3. Rickettsiae. Ann. Parasitol. 2016, 62, 89–100. [Google Scholar] [CrossRef]
- Karbowiak, G.; Biernat, B.; Stańczak, J.; Werszko, J.; Wróblewski, P.; Szewczyk, T.; Sytykiewicz, H. The role of particular ticks developmental stages in the circulation of tick-borne pathogens in Central Europe. 4. Anaplasmataceae. Ann. Parasitol. 2016, 62, 267–284. [Google Scholar] [CrossRef]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef]
- Knoll, S.; Springer, A.; Hauck, D.; Schunack, B.; Pachnicke, S.; Strube, C. Regional, seasonal, biennial and landscape-associated distribution of Anaplasma phagocytophilum and Rickettsia spp. infections in Ixodes ticks in northern Germany and implications for risk assessment at larger spatial scales. Ticks Tick Borne Dis. 2021, 12, 101657. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Raoult, D. Tick-borne rickettsioses around the world: Emerging diseases challenging old concepts. Clin. Microbiol. Rev. 2005, 18, 719–756. [Google Scholar] [CrossRef]
- Sprong, H.; Wielinga, P.R.; Fonville, M.; Reusken, C.; Brandenburg, A.H.; Borgsteede, F.; Gaasenbeek, C.; van der Giessen, J.W. Ixodes ricinus ticks are reservoir hosts for Rickettsia helvetica and potentially carry flea-borne Rickettsia species. Parasites Vectors 2009, 2, 41. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Tian, Z.; Liu, Z.; Niu, Q.; Han, R.; Li, Y.; Guan, G.; Liu, J.; Liu, G.; Luo, J.; et al. Novel spotted fever group rickettsiae in Haemaphysalis qinghaiensis ticks from Gansu, northwest China. Parasites Vectors 2016, 9, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piotrowski, M.; Rymaszewska, A. Expansion of tick-borne rickettsioses in the world. Microorganisms 2020, 8, 1906. [Google Scholar] [CrossRef]
- Dobler, G.; Wölfel, R. Typhus and other rickettsioses: Emerging infections in Germany. Dtsch. Arztebl. Int. 2009, 106, 348–354. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borşan, S.-D.; Ionică, A.M.; Galon, C.; Toma-Naic, A.; Peştean, C.; Sándor, A.D.; Moutailler, S.; Mihalca, A.D. High diversity, prevalence, and co-infection rates of tick-borne pathogens in ticks and wildlife hosts in an urban area in Romania. Front. Microbiol. 2021, 12, 645002. [Google Scholar] [CrossRef]
- Blazejak, K.; Janecek, E.; Strube, C. A 10-year surveillance of Rickettsiales (Rickettsia spp. and Anaplasma phagocytophilum) in the city of Hanover, Germany, reveals Rickettsia spp. as emerging pathogens in ticks. Parasites Vectors 2017, 10, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- André, M.R. Diversity of Anaplasma and Ehrlichia/Neoehrlichia agents in terrestrial wild carnivores worldwide: Implications for human and domestic animal health and wildlife conservation. Front. Vet. Sci. 2018, 5, 293. [Google Scholar] [CrossRef]
- Matei, I.A.; Estrada-Peña, A.; Cutler, S.J.; Vayssier-Taussat, M.; Varela-Castro, L.; Potkonjak, A.; Zeller, H.; Mihalca, A.D. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasites Vectors 2019, 12, 599. [Google Scholar] [CrossRef]
- Wass, L.; Grankvist, A.; Bell-Sakyi, L.; Bergström, M.; Ulfhammer, E.; Lingblom, C.; Wennerås, C. Cultivation of the causative agent of human neoehrlichiosis from clinical isolates identifies vascular endothelium as a target of infection. Emerg. Microbes Infect. 2019, 8, 413–425. [Google Scholar] [CrossRef]
- Pedersen, B.N.; Jenkins, A.; Paulsen, K.M.; Okbaldet, Y.B.; Edgar, K.S.; Lamsal, A.; Soleng, A.; Andreassen, Å.K. Distribution of Neoehrlichia mikurensis in Ixodes ricinus ticks along the coast of Norway: The western seaboard is a low-prevalence region. Zoonoses Public Health 2020, 67, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Silaghi, C.; Woll, D.; Mahling, M.; Pfister, K.; Pfeffer, M. Candidatus Neoehrlichia mikurensis in rodents in an area with sympatric existence of the hard ticks Ixodes ricinus and Dermacentor reticulatus, Germany. Parasites Vectors 2012, 5, 285. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, G.; Sekeyova, Z.; Raoult, D.; Mediannikov, O. Multiple tick-associated bacteria in Ixodes ricinus from Slovakia. Ticks Tick Borne Dis. 2012, 3, 406–410. [Google Scholar] [CrossRef]
- Portillo, A.; Palomar, A.M.; de Toro, M.; Santibáñez, S.; Santibáñez, P.; Oteo, J.A. Exploring the bacteriome in anthropophilic ticks: To investigate the vectors for diagnosis. PLoS ONE 2019, 14, e0213384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben, I.; Lozynskyi, I. Prevalence of Anaplasma phagocytophilum in Ixodes ricinus and Dermacentor reticulatus and coinfection with Borrelia burgdorferi and tick-borne encephalitis virus in western Ukraine. Vector Borne Zoonotic Dis. 2019, 19, 793–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sytykiewicz, H.; Karbowiak, G.; Hapunik, J.; Szpechciński, A.; Supergan-Marwicz, M.; Goławska, S.; Sprawka, I.; Czerniewicz, P. Molecular evidence of Anaplasma phagocytophilum and Babesia microti co-infections in Ixodes ricinus ticks in central-eastern region of Poland. Ann. Agric. Environ. Med. 2012, 19, 45–49. [Google Scholar]
- Greay, T.L.; Gofton, A.W.; Paparini, A.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. Recent insights into the tick microbiome gained through next-generation sequencing. Parasites Vectors 2018, 11, 12. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Janecek, E.; Streichan, S.; Strube, C. SNP-based real-time pyrosequencing as a sensitive and specific tool for identification and differentiation of Rickettsia species in Ixodes ricinus ticks. BMC Infect. Dis. 2012, 12, 261. [Google Scholar] [CrossRef] [Green Version]
- Bunikis, J.; Garpmo, U.; Tsao, J.; Berglund, J.; Fish, D.; Barbour, A.G. Sequence typing reveals extensive strain diversity of the Lyme borreliosis agents Borrelia burgdorferi in North America and Borrelia afzelii in Europe. Microbiology 2004, 150, 1741–1755. [Google Scholar] [CrossRef] [Green Version]
- Krebs, H.A.; Johnson, W.A. Metabolism of ketonic acids in animal tissues. Biochem. J. 1937, 31, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Blanda, V.; D’Agostino, R.; Giudice, E.; Randazzo, K.; La Russa, F.; Villari, S.; Vullo, S.; Torina, A. New real-time PCRs to differentiate Rickettsia spp. and Rickettsia conorii. Molecules 2020, 25, 4431. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.; Fingerle, V.; Noll, M. Analysis of tick surface decontamination methods. Microorganisms 2020, 8, 987. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.; Müller, T.; Fingerle, V.; Noll, M. Presence of human pathogens of the Borrelia burgdorferi sensu lato complex shifts the sequence read abundances of tick microbiomes in two german locations. Microorganisms 2021, 9, 1814. [Google Scholar] [CrossRef] [PubMed]
- Courtney, J.W.; Kostelnik, L.M.; Zeidner, N.S.; Massung, R.F. Multiplex real-time PCR for detection of Anaplasma phagocytophilum and Borrelia burgdorferi. J. Clin. Microbiol. 2004, 42, 3164–3168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capelli, G.; Ravagnan, S.; Montarsi, F.; Ciocchetta, S.; Cazzin, S.; Porcellato, E.; Babiker, A.M.; Cassini, R.; Salviato, A.; Cattoli, G.; et al. Occurrence and identification of risk areas of Ixodes ricinus-borne pathogens: A cost-effectiveness analysis in north-eastern Italy. Parasites Vectors 2012, 5, 61. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, A.; Krämer, A.; Sachse, S.; Straube, E. Detection of Rickettsia spp. and Anaplasma phagocytophilum in Ixodes ricinus ticks in a region of Middle Germany (Thuringia). Ticks Tick-Borne Dis. 2010, 1, 52–56. [Google Scholar] [CrossRef]
- Plantard, O.; Bouju-Albert, A.; Malard, M.-A.; Hermouet, A.; Capron, G.; Verheyden, H. Detection of Wolbachia in the tick Ixodes ricinus is due to the presence of the hymenoptera endoparasitoid Ixodiphagus hookeri. PLoS ONE 2012, 7, e30692. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information. National Center for biotechnology information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 27 December 2022).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Hiltemann, S.; Batut, B.; Clements, D. 16S Microbial analysis with Mothur (extended) (galaxy training materials). Available online: https://training.galaxyproject.org/training-material/topics/metagenomics/tutorials/mothur-miseq-sop/tutorial.html (accessed on 13 April 2021).
- Batut, B.; Hiltemann, S.; Bagnacani, A.; Baker, D.; Bhardwaj, V.; Blank, C.; Bretaudeau, A.; Brillet-Guéguen, L.; Čech, M.; Chilton, J.; et al. Community-driven data analysis training for biology. Cell Syst. 2018, 6, 752–758.e1. [Google Scholar] [CrossRef]
- Schicht, S.; Schnieder, T.; Strube, C. Rickettsia spp. and coinfections with other pathogenic microorganisms in hard ticks from northern Germany. J. Med. Entomol. 2012, 49, 766–771. [Google Scholar] [CrossRef] [Green Version]
- Tappe, J.; Jordan, D.; Janecek, E.; Fingerle, V.; Strube, C. Revisited: Borrelia burgdorferi sensu lato infections in hard ticks (Ixodes ricinus) in the city of Hanover (Germany). Parasites Vectors 2014, 7, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tappe, J.; Strube, C. Anaplasma phagocytophilum and Rickettsia spp. infections in hard ticks (Ixodes ricinus) in the city of Hanover (Germany): Revisited. Ticks Tick Borne Dis. 2013, 4, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Zindel, R.; Gottlieb, Y.; Aebi, A. Arthropod symbioses: A neglected parameter in pest- and disease-control programmes. J. Appl. Ecol. 2011, 48, 864–872. [Google Scholar] [CrossRef] [Green Version]
- Collatz, J.; Selzer, P.; Fuhrmann, A.; Oehme, R.M.; Mackenstedt, U.; Kahl, O.; Steidle, J.L.M. A hidden beneficial: Biology of the tick-wasp Ixodiphagus hookeri in Germany. J. Appl. Entomol. 2011, 135, 351–358. [Google Scholar] [CrossRef]
- Svitálková, Z.; Haruštiaková, D.; Mahríková, L.; Berthová, L.; Slovák, M.; Kocianová, E.; Kazimírová, M. Anaplasma phagocytophilum prevalence in ticks and rodents in an urban and natural habitat in south-western Slovakia. Parasites Vectors 2015, 8, 276. [Google Scholar] [CrossRef] [Green Version]
- Klitgaard, K.; Kjær, L.J.; Isbrand, A.; Hansen, M.F.; Bødker, R. Multiple infections in questing nymphs and adult female Ixodes ricinus ticks collected in a recreational forest in Denmark. Ticks Tick-Borne Dis. 2019, 10, 1060–1065. [Google Scholar] [CrossRef]
- Turebekov, N.; Abdiyeva, K.; Yegemberdiyeva, R.; Dmitrovsky, A.; Yeraliyeva, L.; Shapiyeva, Z.; Amirbekov, A.; Oradova, A.; Kachiyeva, Z.; Ziyadina, L.; et al. Prevalence of Rickettsia species in ticks including identification of unknown species in two regions in Kazakhstan. Parasites Vectors 2019, 12, 197. [Google Scholar] [CrossRef]
- Akimov, I.; Nebogatkin, I. Distribution of the tick Haemaphysalis punctata (Acari, Ixodidae) in Ukraine. Vestn. Zool. 2012, 46, e-46–e-51. [Google Scholar] [CrossRef] [Green Version]
- Guccione, C.; Colomba, C.; Tolomeo, M.; Trizzino, M.; Iaria, C.; Cascio, A. Rickettsiales in Italy. Pathogens 2021, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Portillo, A.; Santibáñez, S.; García-Álvarez, L.; Palomar, A.M.; Oteo, J.A. Rickettsioses in Europe. Microbes Infect. 2015, 17, 834–838. [Google Scholar] [CrossRef] [PubMed]
- May, K.; Strube, C. Prevalence of Rickettsiales (Anaplasma phagocytophilum and Rickettsia spp.) in hard ticks (Ixodes ricinus) in the city of Hamburg, Germany. Parasitol. Res. 2014, 113, 2169–2175. [Google Scholar] [CrossRef] [PubMed]
- Biernat, B.; Stańczak, J.; Michalik, J.; Sikora, B.; Cieniuch, S. Rickettsia helvetica and R. monacensis infections in immature Ixodes ricinus ticks derived from sylvatic passerine birds in west-central Poland. Parasitol. Res. 2016, 115, 3469–3477. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.N.; Jenkins, A.; Kjelland, V. Tick-borne pathogens in Ixodes ricinus ticks collected from migratory birds in southern Norway. PLoS ONE 2020, 15, e0230579. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variable | Characteristic |
|---|---|
| B. burgdorferi s. l. | Positive (n = 380), negative (n = 380) |
| Area | South (n = 120), southeast (n = 560), west (n = 80) |
| Month | May (n = 180), June (n = 380), July (n = 140), September (n = 60) |
| Season | Spring (n = 180), summer (n = 520), autumn (n = 60) |
| Bacterial Species/ Genus | Target Gene | Amplicon Length [bp] | Final Primer Concentration (nM) | Primer Set * | References |
|---|---|---|---|---|---|
| A. phagocytophilum | msp2 | 77 | 400 | ApMSP2f: 5′-ATGGAAGGTAGTGTTGGTTATGGTATT-3′ ApMSP2r: 5′-TTGGTCTTGAAGCGCTCGTA-3′ | [35] |
| N. mikurensis | groEL | 1024 | 500 | NM-128f: 5′-AACAGGTGAAACACTAGATAAGTCCAT-3′ NM-1152f: 5′-TTCTACTTTGAACATTTGAAGAATTACTAT-3′ | [36] |
| Rickettsia spp. | gltA | 340 | 400 | RH314f: 5′-AAACAGGTTGCTCATCATTC-3′ RH654r: 5′-AGAGCATTTTTTATTATTGG-3′ | [37] |
| W. pipientis | wsp | 591 | 500 | wsp-81f: 5′-TGGTCCAATAAGTGATGAAGAAAC-3′ wsp-691r: 5′-AAAAATTAAACGCTACTCCA-3′ | [38] |
| Bacterial Species or Genus | B. burgdorferi s. l. Finding | Prevalence of Positive Findings | Total Prevalence of Positive Findings |
|---|---|---|---|
| A. phagocytophilum | negative | 13 (1.7%) | 21 (2.8%) |
| positive | 8 (1.0%) | ||
| N. mikurensis | negative | 1 (0.1%) | 1 (0.1%) |
| positive | 0 (0.0%) | ||
| Rickettsia spp. | negative | 53 (7.0%) | 127 (16.7%) |
| positive | 74 (9.7%) | ||
| W. pipientis | negative | 50 (6.6%) | 121 (15.9%) |
| positive | 71 (9.3%) |
| Frequence of co-Infection | Genus/ Species | Prevalence | Total Prevalence |
|---|---|---|---|
| Double | B. burgdorferi s. l. + A. phagocytophilum | 3/0.4% | 60 (7.9%) |
| B. burgdorferi s. l. + Rickettsia spp. | 57/7.5% | ||
| Triple | B. burgdorferi s. l. + A. Phagocytophilum + Rickettsia spp. | 2/0.3% | 2/0.3% |
| 62/8.1% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffmann, A.; Müller, T.; Fingerle, V.; Silaghi, C.; Noll, M. Co-Infection of Potential Tick-Borne Pathogens of the Order Rickettsiales and Borrelia burgdorferi s. l. and Their Link to Season and Area in Germany. Microorganisms 2023, 11, 157. https://doi.org/10.3390/microorganisms11010157
Hoffmann A, Müller T, Fingerle V, Silaghi C, Noll M. Co-Infection of Potential Tick-Borne Pathogens of the Order Rickettsiales and Borrelia burgdorferi s. l. and Their Link to Season and Area in Germany. Microorganisms. 2023; 11(1):157. https://doi.org/10.3390/microorganisms11010157
Chicago/Turabian StyleHoffmann, Angeline, Thomas Müller, Volker Fingerle, Cornelia Silaghi, and Matthias Noll. 2023. "Co-Infection of Potential Tick-Borne Pathogens of the Order Rickettsiales and Borrelia burgdorferi s. l. and Their Link to Season and Area in Germany" Microorganisms 11, no. 1: 157. https://doi.org/10.3390/microorganisms11010157