
Human Fecal Bile Acid Analysis after Investigational Microbiota-Based Live Biotherapeutic Delivery for Recurrent Clostridioides difficile Infection
Abstract
:1. Introduction
2. Methods
2.1. Preparation of BA Standards
2.2. Human Fecal Sample Preparation
2.3. Sample Processing
2.4. LC-MS/MS Analytical Method
- –
- Ultra-performance liquid chromatography (UPLC) Column: Waters ethylene bridge hybrid (BEH) C18 1.7 µm, 2.1 mm × 150 mm Column
- –
- Mobile Phase A: 20 mM Ammonium Acetate 0.1% Formic Acid in Water
- –
- Mobile Phase B: 20 mM Ammonium Acetate 0.1% Formic Acid in 1:3 Methanol:Acetonitrile
- –
- Gradient: 35% B for 5 min, 35−100% B in 20 min, 5 min re-equilibration
- –
- Flow Rate: 0.2 mL/min
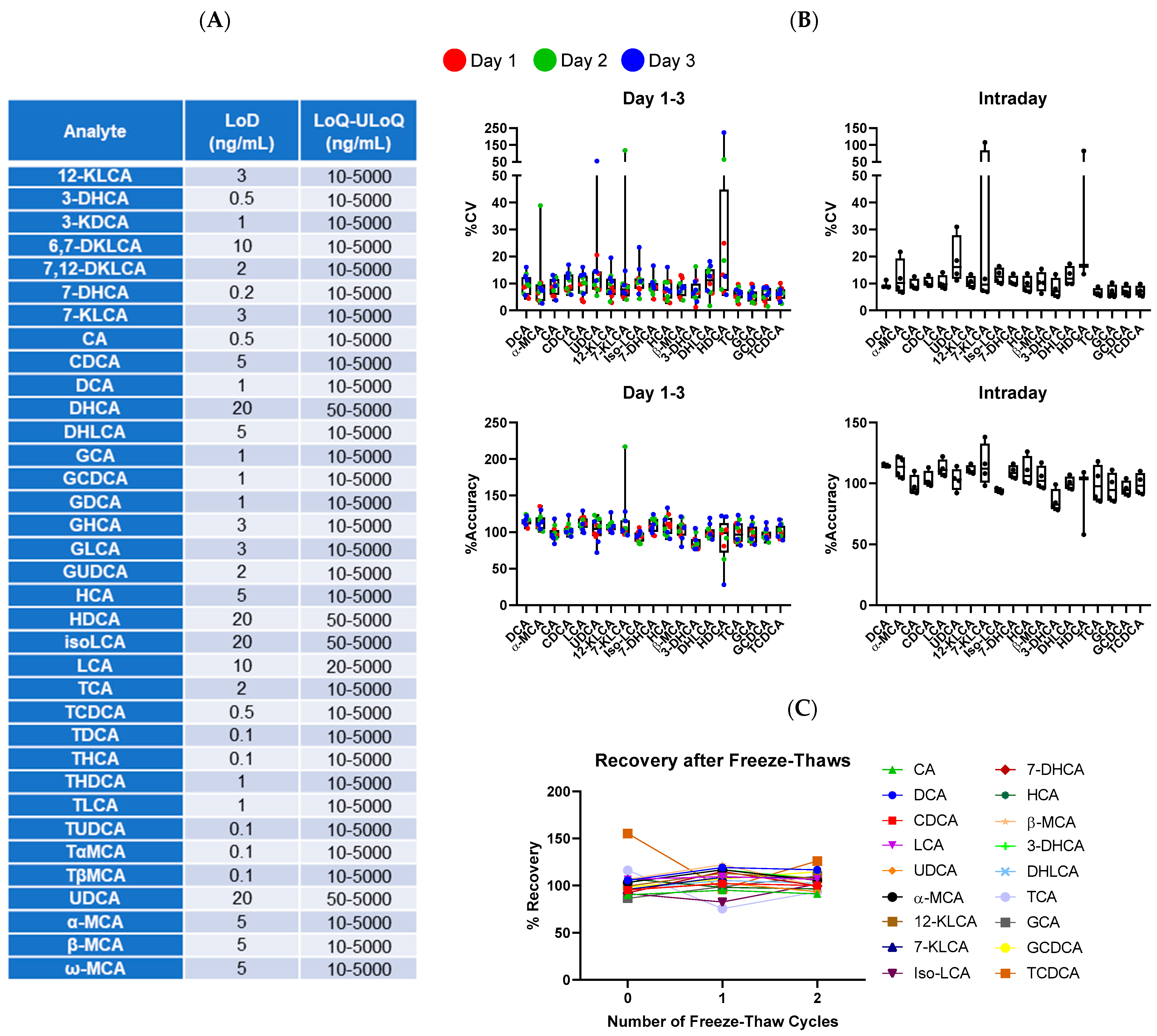
2.5. Assay Characterization
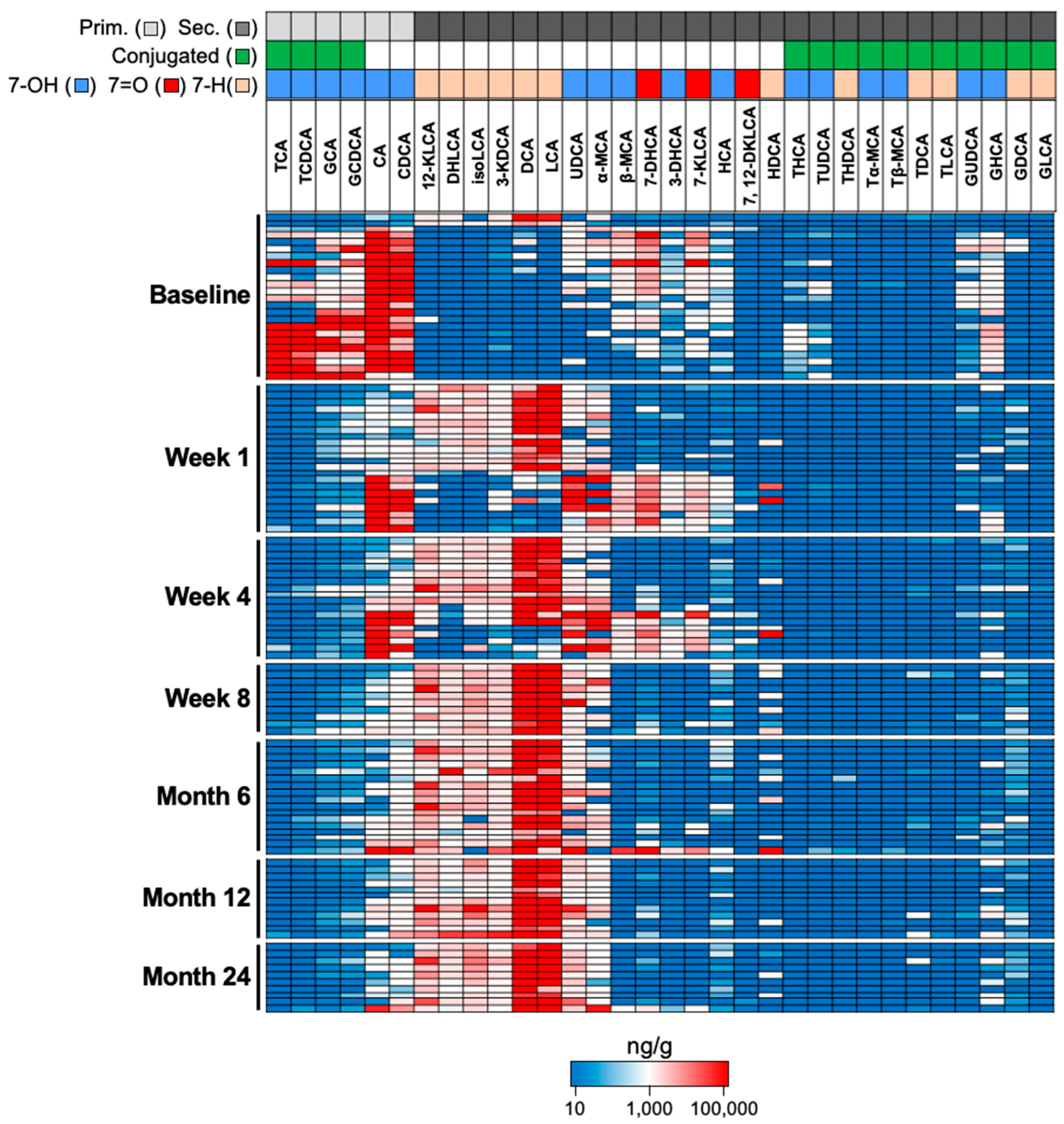
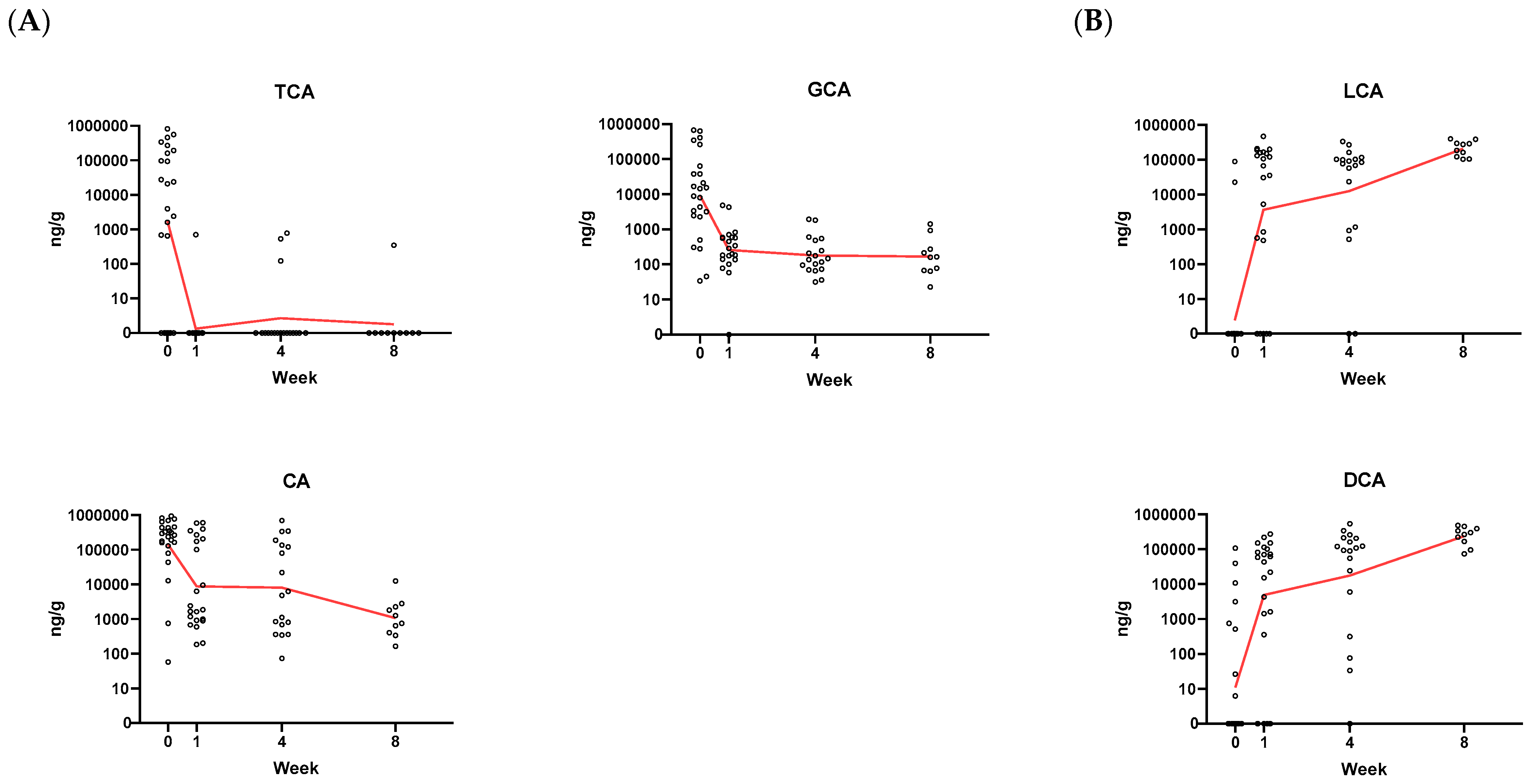
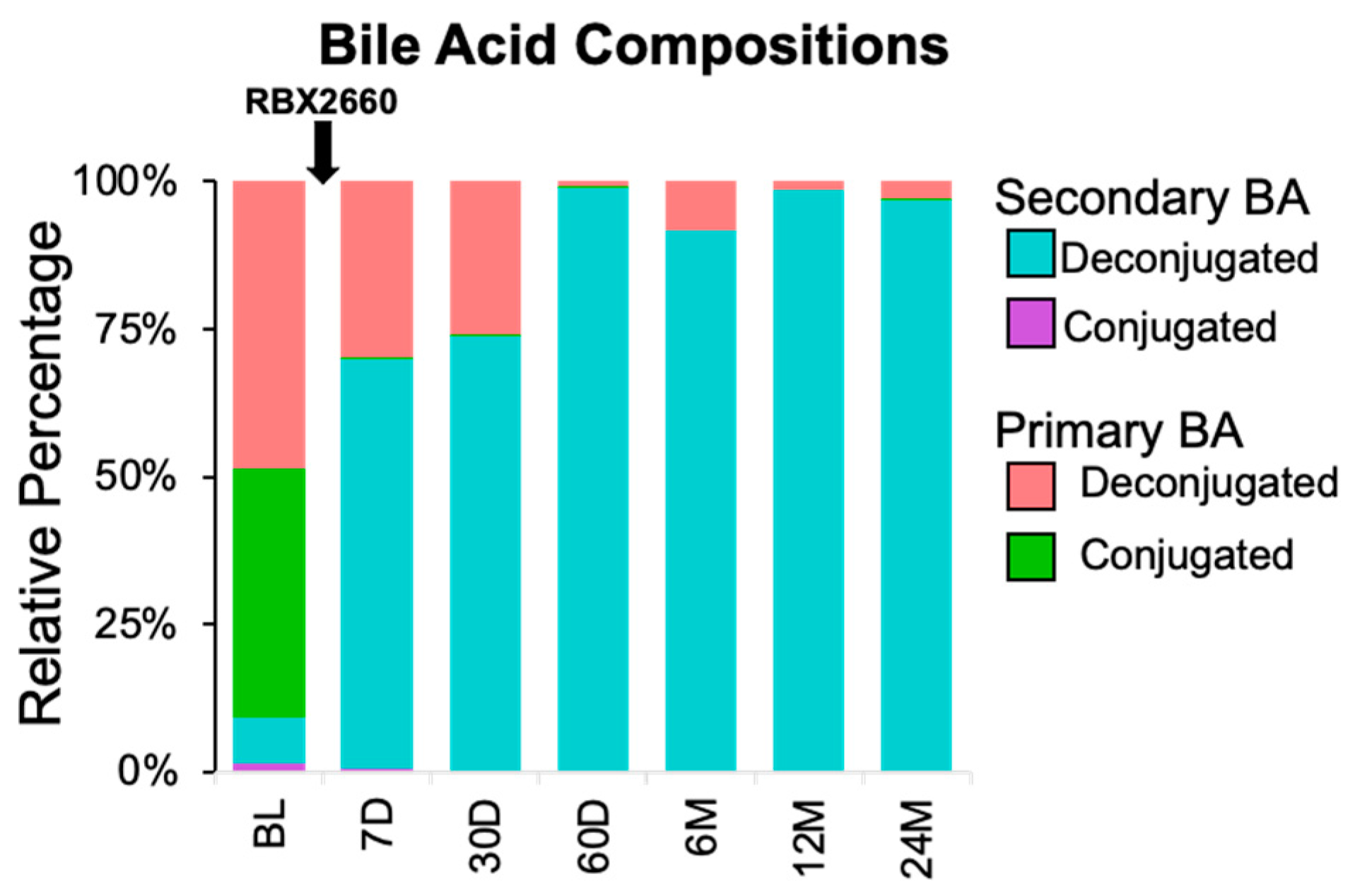
2.6. Application of Assay to Assess BA Composition in Samples from rCDI Patients Treated with RBX2660
2.7. Statistics
3. Results
3.1. Assay Principle and Development
3.2. Assay Properties and Characterization
3.3. Proof-of-Concept of BA Metabolic Assessment in Clinical Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheitoyan-Pesant, C.; Abou Chakra, C.N.; Pépin, J.; Marcil-Héguy, A.; Nault, V.; Valiquette, L. Clinical and Healthcare Burden of Multiple Recurrences of Clostridium difficile Infection. Clin. Infect. Dis. 2016, 62, 574–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullish, B.H.; Allegretti, J.R. The contribution of bile acid metabolism to the pathogenesis of Clostridioides difficile infection. Ther. Adv. Gastroenterol. 2021, 14, 17562848211017724. [Google Scholar] [CrossRef] [PubMed]
- Reed, A.D.; Theriot, C.M. Contribution of Inhibitory Metabolites and Competition for Nutrients to Colonization Resistance against Clostridioides difficile by Commensal Clostridium. Microorganisms 2021, 9, 371. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid. Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buffie, C.G.; Bucci, V.; Stein, R.R.; McKenney, P.T.; Ling, L.; Gobourne, A.; No, D.; Liu, H.; Kinnebrew, M.; Viale, A.; et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 2015, 517, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Weingarden, A.R.; Dosa, P.I.; DeWinter, E.; Steer, C.J.; Shaughnessy, M.K.; Johnson, J.R.; Khoruts, A.; Sadowsky, M.J. Changes in Colonic Bile Acid Composition following Fecal Microbiota Transplantation Are Sufficient to Control Clostridium difficile Germination and Growth. PLoS ONE 2016, 11, e0147210. [Google Scholar] [CrossRef] [Green Version]
- Orenstein, R.; Dubberke, E.R.; Khanna, S.; Lee, C.H.; Yoho, D.; Johnson, S.; Hecht, G.; DuPont, H.L.; Gerding, D.N.; Blount, K.F.; et al. Durable reduction of Clostridioides difficile infection recurrence and microbiome restoration after treatment with RBX2660: Results from an open-label phase 2 clinical trial. BMC Infect. Dis. 2022, 22, 245. [Google Scholar] [CrossRef]
- Blount, K.F.; Shannon, W.D.; Deych, E.; Jones, C. Restoration of Bacterial Microbiome Composition and Diversity Among Treatment Responders in a Phase 2 Trial of RBX2660: An Investigational Microbiome Restoration Therapeutic. Open Forum Infect. Dis. 2019, 6, ofz095. [Google Scholar] [CrossRef] [Green Version]
- Langdon, A.; Schwartz, D.J.; Bulow, C.; Sun, X.; Hink, T.; Reske, K.A.; Jones, C.; Burnham, C.A.D.; Dubberke, E.R.; Dantas, G. Microbiota restoration reduces antibiotic-resistant bacteria gut colonization in patients with recurrent Clostridioides difficile infection from the open-label PUNCH CD study. Genome Med. 2021, 13, 28. [Google Scholar] [CrossRef]
- Kwak, S.; Choi, J.; Hink, T.; Reske, K.A.; Blount, K.; Jones, C.; Bost, M.H.; Sun, X.; Burnham, C.A.D.; Dubberke, E.R.; et al. Impact of investigational microbiota therapeutic RBX2660 on the gut microbiome and resistome revealed by a placebo-controlled clinical trial. Microbiome 2020, 8, 125. [Google Scholar] [CrossRef]
- Blount, K.; Jones, C.; Walsh, D.; Gonzalez, C.; Shannon, W.D. Development and Validation of a Novel Microbiome-Based Biomarker of Post-antibiotic Dysbiosis and Subsequent Restoration. Front. Microbiol. 2021, 12, 781275. [Google Scholar] [CrossRef]
- Shafaei, A.; Rees, J.; Christophersen, C.T.; Devine, A.; Broadhurst, D.; Boyce, M.C. Extraction and quantitative determination of bile acids in feces. Anal. Chim. Acta. 2021, 1150, 338224. [Google Scholar] [CrossRef]
- Wegner, K.; Just, S.; Gau, L.; Mueller, H.; Gérard, P.; Lepage, P.; Clavel, T.; Rohn, S. Rapid analysis of bile acids in different biological matrices using LC-ESI-MS/MS for the investigation of bile acid transformation by mammalian gut bacteria. Anal. Bioanal. Chem. 2017, 409, 1231–1245. [Google Scholar] [CrossRef]
- Perwaiz, S.; Mignault, D.; Tuchweber, B.; Yousef, I.M. Rapid and improved method for the determination of bile acids in human feces using MS. Lipids 2002, 37, 1093–1100. [Google Scholar] [CrossRef]
- Liu, Y.; Rong, Z.; Xiang, D.; Zhang, C.; Liu, D. Detection technologies and metabolic profiling of bile acids: A comprehensive review. Lipids Health Dis. 2018, 17, 121. [Google Scholar] [CrossRef] [Green Version]
- Dubberke, E.R.; Lee, C.H.; Orenstein, R.; Khanna, S.; Hecht, G.; Gerding, D.N. Results From a Randomized, Placebo-Controlled Clinical Trial of a RBX2660-A Microbiota-Based Drug for the Prevention of Recurrent Clostridium difficile Infection. Clin. Infect. Dis. 2018, 67, 1198–1204. [Google Scholar] [CrossRef] [Green Version]
- Theriot, C.M.; Koenigsknecht, M.J.; Carlson, P.E.; Hatton, G.E.; Nelson, A.M.; Li, B.; Huffnagle, G.B.; ZLi, J.; Young, V.B. Antibiotic-induced shifts in the mouse gut microbiome and metabolome increase susceptibility to Clostridium difficile infection. Nat. Commun. 2014, 5, 3114. [Google Scholar] [CrossRef] [Green Version]
- Winston, J.A.; Theriot, C.M. Impact of microbial derived secondary bile acids on colonization resistance against Clostridium difficile in the gastrointestinal tract. Anaerobe 2016, 41, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Guzior, D.V.; Quinn, R.A. Review: Microbial transformations of human bile acids. Microbiome 2021, 9, 140. [Google Scholar] [CrossRef]
- Sorg, J.A.; Sonenshein, A.L. Bile salts and glycine as cogerminants for Clostridium difficile spores. J. Bacteriol. 2008, 190, 2505–2512. [Google Scholar] [CrossRef]
- Sorg, J.A.; Sonenshein, A.L. Chenodeoxycholate is an inhibitor of Clostridium difficile spore germination. J. Bacteriol. 2009, 191, 1115–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, K.H. Efficiency of various bile salt preparations for stimulation of Clostridium difficile spore germination. J. Clin. Microbiol. 1983, 18, 1017–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reijnders, D.; Goossens, G.H.; Hermes, G.D.; Neis, E.P.; van der Beek, C.M.; Most, J.; Holst, J.J.; Lenaerts, K.; Kootte, R.S.; Nieuwdorp, M.; et al. Effects of Gut Microbiota Manipulation by Antibiotics on Host Metabolism in Obese Humans: A Randomized Double-Blind Placebo-Controlled Trial. Cell Metab. 2016, 24, 63–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrieze, A.; Out, C.; Fuentes, S.; Jonker, L.; Reuling, I.; Kootte, R.S.; van Nood, E.; Holleman, F.; Knaapen, M.; Romijn, J.A.; et al. Impact of oral vancomycin on gut microbiota, bile acid metabolism, and insulin sensitivity. J. Hepatol. 2014, 60, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Allegretti, J.R.; Kearney, S.; Li, N.; Bogart, E.; Bullock, K.; Gerber, G.K.; Bry, L.; Clish, C.B.; Alm, E.; Korzenik, J.R. Recurrent Clostridium difficile infection associates with distinct bile acid and microbiome profiles. Aliment. Pharmacol. Ther. 2016, 43, 1142–1153. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.R.M.; Flemer, B.; Joyce, S.A.; Zulquernain, A.; Sheehan, D.; Shanahan, F.; O’Toole, P.W. Changes in microbiota composition, bile and fatty acid metabolism, in successful faecal microbiota transplantation for Clostridioides difficile infection. BMC Gastroenterol. 2018, 18, 131. [Google Scholar] [CrossRef] [Green Version]
- Seekatz, A.M.; Theriot, C.M.; Rao, K.; Chang, Y.M.; Freeman, A.E.; Kao, J.Y.; Young, V.B. Restoration of short chain fatty acid and bile acid metabolism following fecal microbiota transplantation in patients with recurrent Clostridium difficile infection. Anaerobe 2018, 53, 64–73. [Google Scholar] [CrossRef]
- Piccoli, S.P.; Garofolo, F. Biomarker assay validation. Bioanalysis 2018, 10, 889–891. [Google Scholar] [CrossRef] [Green Version]
- Center for Drug Evaluation, Research. Bioanalytical Method Validation Guidance for Industry. U.S. Food and Drug Administration, 2020. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/bioanalytical-method-validation-guidance-industry (accessed on 9 May 2022).
- Rose, C.; Parker, A.; Jefferson, B.; Cartmell, E. The Characterization of Feces and Urine: A Review of the Literature to Inform Advanced Treatment Technology. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1827–1879. [Google Scholar] [CrossRef] [Green Version]
- Caroff, D.A.; Edelstein, P.H.; Hamilton, K.; Pegues, D.A.; CDC Prevention Epicenters Program. The Bristol stool scale and its relationship to Clostridium difficile infection. J. Clin. Microbiol. 2014, 52, 3437–3439. [Google Scholar] [CrossRef]
- Qian, X.; Yanagi, K.; Kane, A.V.; Alden, N.; Lei, M.; Snydman, D.R.; Vickers, R.J.; Lee, K.; Thorpe, C.M. Ridinilazole, a narrow spectrum antibiotic for treatment of Clostridioides difficile infection, enhances preservation of microbiota-dependent bile acids. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G227–G237. [Google Scholar] [CrossRef]
- Garey, K.W.; McPherson, J.; Dinh, A.Q.; Hu, C.; Jo, J.; Wang, W.; Lancaster, C.K.; Gonzales-Luna, A.J.; Loveall, C.; Begum, K.; et al. Efficacy, Safety, Pharmacokinetics, and Microbiome Changes of Ibezapolstat in Adults with Clostridioides difficile Infection: A Phase 2a Multicenter Clinical Trial. Clin. Infect. Dis. 2022, 75, 1164–1170. [Google Scholar] [CrossRef]
- Batta, A.K.; Salen, G.; Arora, R.; Shefer, S.; Batta, M.; Person, A. Side chain conjugation prevents bacterial 7-dehydroxylation of bile acids. J. Biol. Chem. 1990, 265, 10925–10928. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Bile Acid Name | Vendor | Cat# |
|---|---|---|---|
| T-α-MCA | Tauro-α-muricholic Acid | Cayman | 20288 |
| CA | Cholic Acid | Cayman | 20250 |
| CDCA | Chenodeoxycholic Acid | Cayman | 10011286 |
| d4-CA | d4-Cholic Acid | Cayman | 20849 |
| d4-GCDCA | d4-Glyco Chenodeoxycholic Acid | Cayman | 21890 |
| d4-TCA | d4-Tauro Cholic Acid | Cayman | 21891 |
| d4-DCA | d4-Deoxycholic Acid | Cayman | 20851 |
| d4-LCA | d4-Lithocholic Acid | Cayman | 20831 |
| d4-UDCA | Ursodeoxycholic Acid | Cayman | 21892 |
| d4-GCA | Glyco Cholic Acid | Cayman | 21889 |
| DCA | Deoxycholic Acid | Cayman | 20756 |
| GCA | Glyco Cholic Acid | Cayman | 20276 |
| GCDCA | Glyco Chenodeoxycholic Acid | Cayman | 16942 |
| GDCA | Glyco Deoxycholic Acid | Cayman | 20274 |
| GHCA | Glyco Hyocholic Acid | Cayman | 22670 |
| GLCA | Glyco Lithocholic Acid | Cayman | 21723 |
| GUDCA | Glyco Ursodeoxycholic Acid | Cayman | 21698 |
| HCA | Hyocholic Acid | Cayman | 20293 |
| HDCA | Hyodeoxycholic Acid | Cayman | 20294 |
| LCA | Lithocholic Acid | Cayman | 20253 |
| TCA | Tauro Cholic Acid | Cayman | 16215 |
| TCDCA | Tauro Chenodeoxycholic Acid | Cayman | 20275 |
| TDCA | Tauro Deoxycholic Acid | Cayman | 15935 |
| THCA | Tauro Hyocholic Acid | Cayman | 22669 |
| THDCA | Tauro Hyodeoxycholic Acid | Cayman | 21956 |
| TLCA | Tauro Lithocholic Acid | Cayman | 17275 |
| TUDCA | Tauro Ursodeoxycholic Acid | Cayman | 20277 |
| T-β-MCA | Tauro-β-muricholic Acid | Cayman | 20289 |
| UDCA | Ursodeoxycholic Acid | Cayman | 15121 |
| α-MCA | α-muricholic Aacid | Cayman | 20291 |
| β-MCA | β-muricholic Acid | Cayman | 20287 |
| ω-MCA | ω-muricholic Acid | Cayman | 20292 |
| 7-KLCA | 7-Ketolithocholic Acid | Steraloids | C1600-000 |
| 7-DHCA/7-KDCA | 7-Dehydrocholic Acid/7-ketodeoxycholic Acid | Steraloids | C1250-000 |
| 3-DHCA | 3-Dehydrocholic Acid | Steraloids | C1272-000 |
| Iso-LCA | Iso-lithocholic Acid | Steraloids | C1475-000 |
| 6-7-DKLCA | 6,7-Diketolithocholic Acid | Steraloids | C1485-000 |
| 7,12 DKLCA | 7,12-Diketolithocholic Acid | Steraloids | C1500-000 |
| 12-KLCA/12-KDCA | 12-Ketolithocholic Acid/12-Ketodeoxhycholic Acid | Steraloids | C1650-000 |
| DHLCA/3-KLCA | Dehydrolithocholic Acid/3-Ketolithocholic Acid | Steraloids | C1750-000 |
| DHCA | Dehydrocholic Acid | Steraloids | C2000-000 |
| 3-KDCA | 3-Ketodeoxycholic Acid | Steraloids | C1725-000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papazyan, R.; Ferdyan, N.; Srinivasan, K.; Gonzalez, C.; Shannon, W.D.; Blount, K.; Fuchs, B.C. Human Fecal Bile Acid Analysis after Investigational Microbiota-Based Live Biotherapeutic Delivery for Recurrent Clostridioides difficile Infection. Microorganisms 2023, 11, 135. https://doi.org/10.3390/microorganisms11010135
Papazyan R, Ferdyan N, Srinivasan K, Gonzalez C, Shannon WD, Blount K, Fuchs BC. Human Fecal Bile Acid Analysis after Investigational Microbiota-Based Live Biotherapeutic Delivery for Recurrent Clostridioides difficile Infection. Microorganisms. 2023; 11(1):135. https://doi.org/10.3390/microorganisms11010135
Chicago/Turabian StylePapazyan, Romeo, Nicky Ferdyan, Karthik Srinivasan, Carlos Gonzalez, William D. Shannon, Ken Blount, and Bryan C. Fuchs. 2023. "Human Fecal Bile Acid Analysis after Investigational Microbiota-Based Live Biotherapeutic Delivery for Recurrent Clostridioides difficile Infection" Microorganisms 11, no. 1: 135. https://doi.org/10.3390/microorganisms11010135