

Yeast Expressed Hybrid Peptide CLP Abridged Pro-Inflammatory Cytokine Levels by Endotoxin Neutralization
 , , , and
, , , and 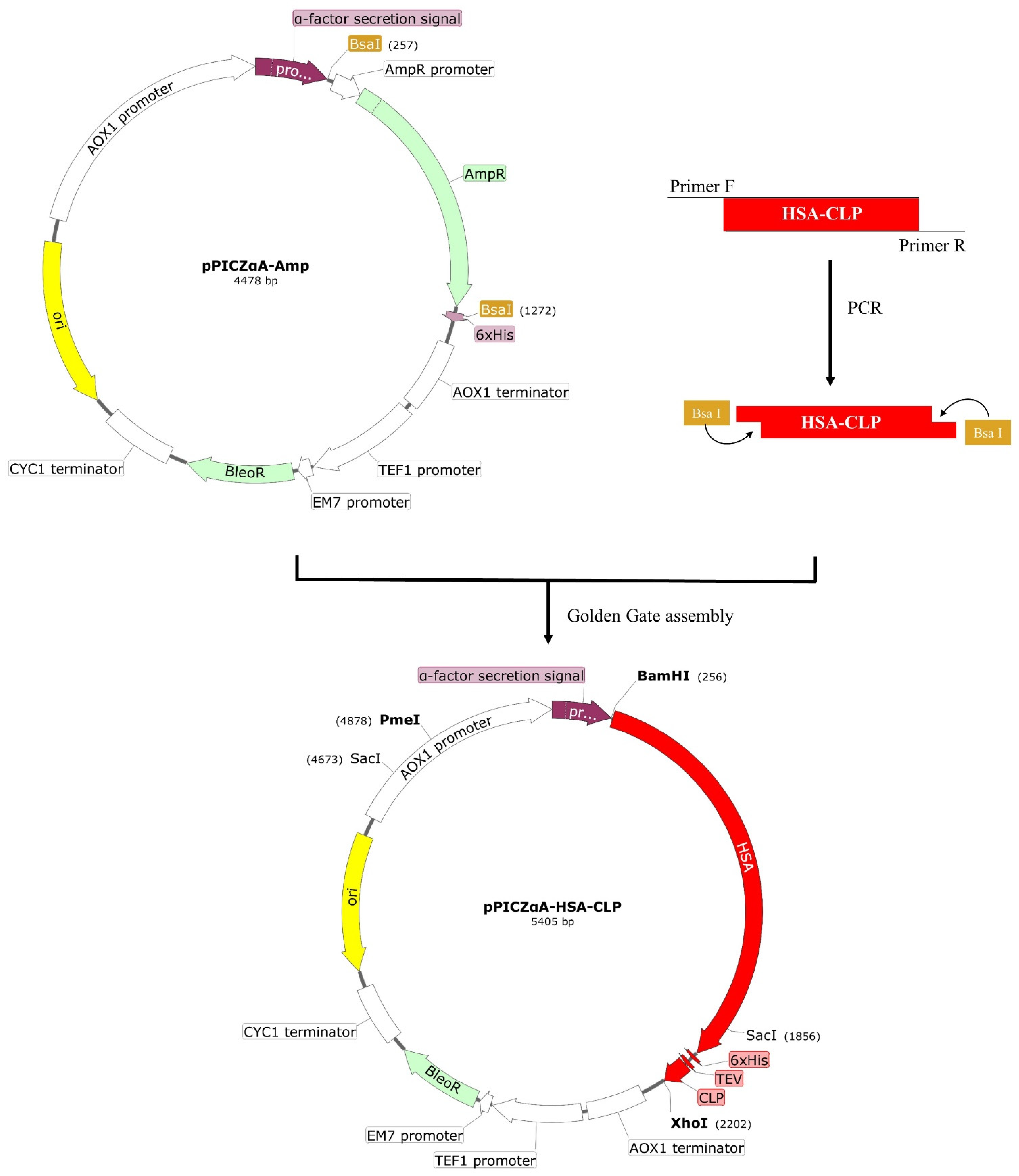{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids, Strains, and Reagents
2.2. Construction of Expression Plasmid pPICZαA-HSA-CLP
2.3. Expression and Purification of Fusion HSA-CLP Peptide
2.4. Cleavage and Purification of Hybrid Peptide CLP
2.5. Activity Assay of Recombinant CLP Peptide
2.5.1. LPS Neutralization
2.5.2. Hemolytic Activity
2.6. Cell Culture
2.6.1. Cell Viability Assay
2.6.2. LPS-Induced Secretion of Nitric Oxide (NO) in Mouse RAW264.7 Macrophages
2.6.3. Evaluation of CLP on LPS-Induced Pro-inflammatory Cytokines in Mouse RAW264.7 Macrophages
2.6.4. Effects of CLP on LPS-Induced Apoptosis of Mouse RAW264.7 Macrophages
2.7. Statistical Analysis
3. Results
3.1. Construction of Recombinant Plasmid pPICZαA-HSA-CLP
3.2. Expression and Purification of Fusion HSA-CLP Peptide
3.3. Cleavage and Purification of Hybrid Peptide CLP
3.4. Inhibition of LPS Activity and Pro-Inflammatory Cytokine Induction
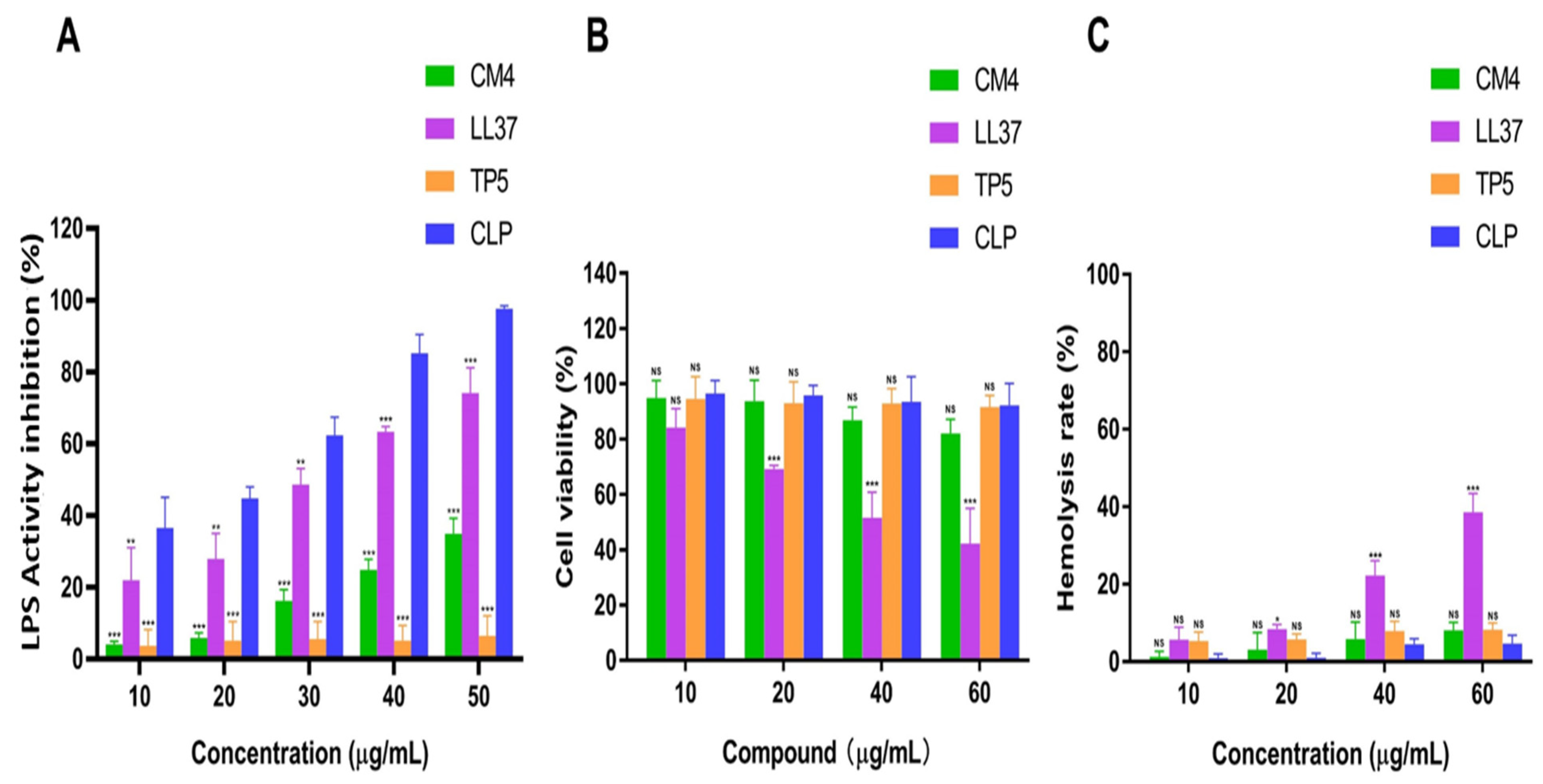
3.4.1. Recombinant Peptide CLP Inhibitory Effect on LPS
3.4.2. Cytotoxicity and Hemolytic Activity of CLP
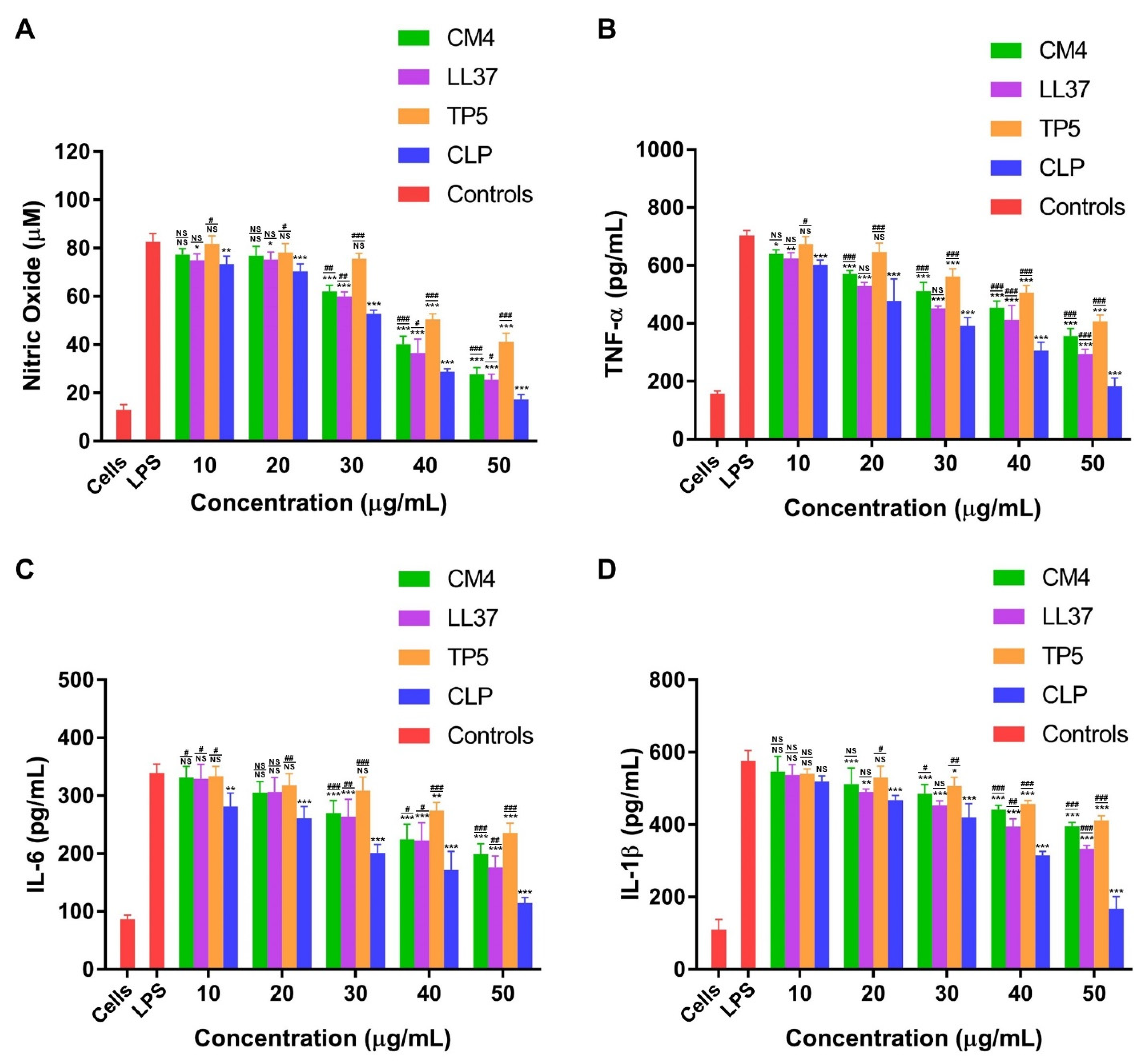
3.4.3. CLP Downregulates LPS-Induced Inflammatory Response in Mouse RAW264.7 Macrophages
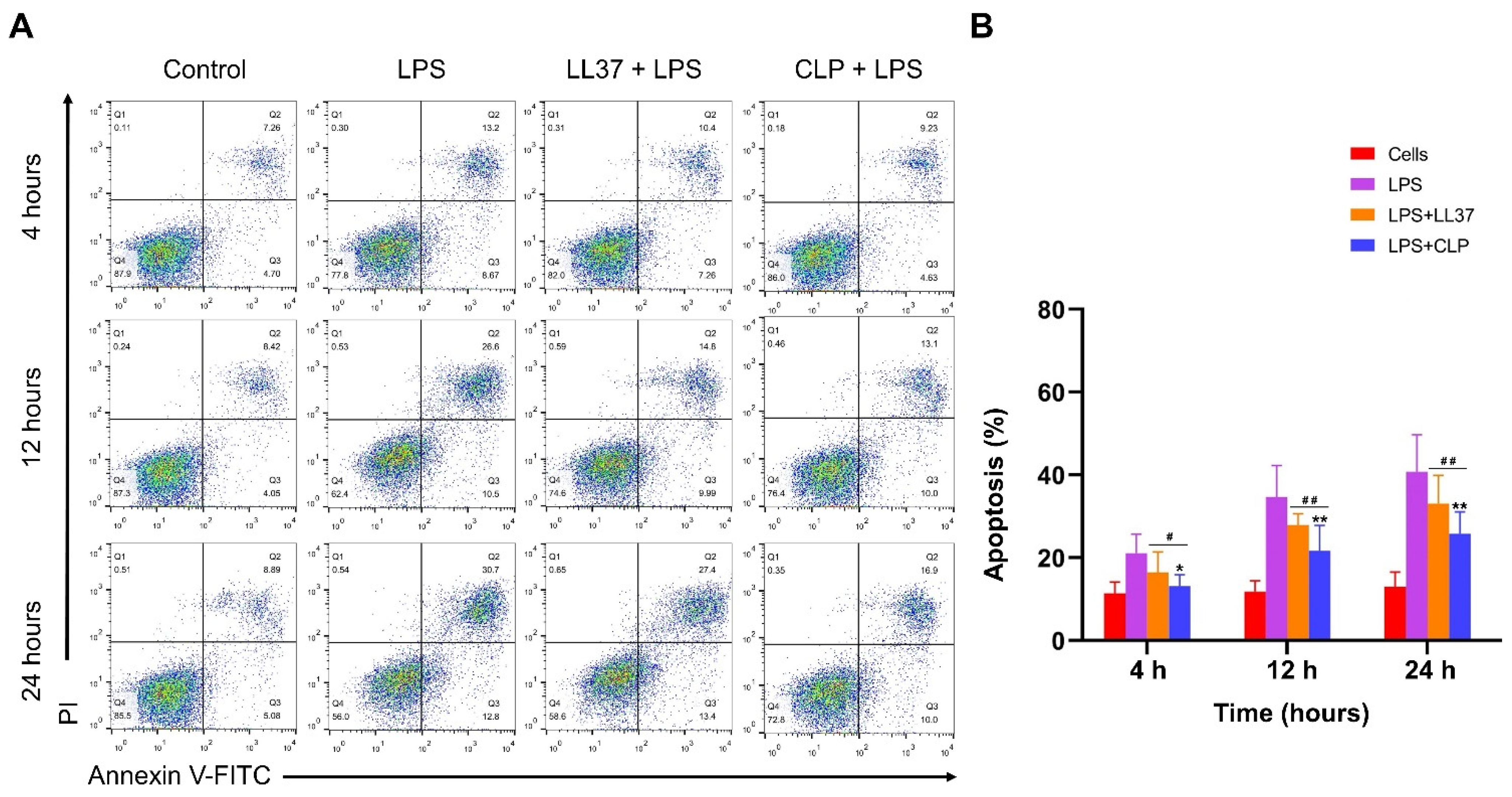
3.4.4. Effects of CLP on LPS-Induced Apoptosis of Mouse RAW264.7 Macrophages
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide Endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [Green Version]
- Wittebole, X.; Castanares-Zapatero, D.; Laterre, P.F. Toll-like Receptor 4 Modulation as a Strategy to Treat Sepsis. Mediat. Inflamm. 2010, 2010, 568396. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Shang, D. Inhibitory Effects of Antimicrobial Peptides on Lipopolysaccharide-Induced Inflammation. Mediat. Inflamm. 2015, 2015, 167572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.J.; Gallo, R.L. Antimicrobial Peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Bahar, A.A.; Ren, D. Antimicrobial Peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Pushpanathan, M.; Gunasekaran, P.; Rajendhran, J. Antimicrobial Peptides: Versatile Biological Properties. Int. J. Pept. 2013, 2013, 675391. [Google Scholar] [CrossRef] [Green Version]
- Brogden, K.A. Antimicrobial Peptides: Pore Formers or Metabolic Inhibitors in Bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Payoungkiattikun, W.; Joompang, A.; Thongchot, S.; Nowichai, B.; Jangpromma, N.; Klaynongsruang, S. Evidence of Multi-Functional Peptide Activity: Potential Role of Kt2 and Rt2 for Anti-Inflammatory, Anti-Oxidative Stress, and Anti-Apoptosis Properties. Appl. Biol. Chem. 2020, 63, 1–13. [Google Scholar] [CrossRef]
- Jiang, S.J.; Tsai, P.I.; Peng, S.Y.; Chang, C.C.; Chung, Y.; Tsao, H.H.; Huang, H.T.; Chen, S.Y.; Hsu, H.J. A Potential Peptide Derived from Cytokine Receptors Can Bind Proinflammatory Cytokines as a Therapeutic Strategy for Anti-Inflammation. Sci. Rep. 2019, 9, 2317. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wei, X.; Zhang, R.; Koci, M.; Si, D.; Ahmad, B.; Guo, H.; Hou, Y. C-Terminal Amination of a Cationic Anti-Inflammatory Peptide Improves Bioavailability and Inhibitory Activity against Lps-Induced Inflammation. Front. Immunol. 2020, 11, 618312. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, X.; Zhang, R.; Petitte, J.N.; Si, D.; Li, Z.; Cheng, J.; Du, M. Design and Development of a Novel Peptide for Treating Intestinal Inflammation. Front. Immunol. 2019, 10, 1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, B.; Li, Z.; Hanif, Q.; Hu, Q.; Wei, X.; Zhang, L.; Khan, S.A.; Aihemaiti, M.; Gulzar, H.; Shahid, M.; et al. A Hybrid Peptide Defb-Tp5 Expressed in Methylotrophic Yeast Neutralizes Lps with Potent Anti-Inflammatory Activities. Front. Pharm. 2020, 11, 461. [Google Scholar] [CrossRef] [PubMed]
- Durr, U.H.; Sudheendra, U.S.; Ramamoorthy, A. Ll-37, the Only Human Member of the Cathelicidin Family of Antimicrobial Peptides. Biochim. Biophys. Acta 2006, 1758, 1408–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Sun, C.; Wang, S.; Su, F.; Zhang, S. Lipopolysaccharide Neutralization by a Novel Peptide Derived from Phosvitin. Int. J. Biochem. Cell Biol. 2013, 45, 2622–2631. [Google Scholar] [CrossRef] [PubMed]
- Pulido, D.; Nogues, M.V.; Boix, E.; Torrent, M. Lipopolysaccharide Neutralization by Antimicrobial Peptides: A Gambit in the Innate Host Defense Strategy. J. Innate Immun. 2012, 4, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Hanif, Q.; Wei, X.; Zhang, L.; Sabir, N.; Li, Z.; Cheng, J.; Khan, S.A.; Basit, A.; Shahid, M.; et al. In Vitro Impact of Yeast Expressed Hybrid Peptide Cath-2tp5 as a Prophylactic Measure toward Sepsis and Inflammation. Front. Bioeng. Biotechnol. 2020, 8, 454. [Google Scholar] [CrossRef]
- Cheng, J.; Ahmat, M.; Guo, H.; Wei, X.; Zhang, L.; Cheng, Q.; Zhang, J.; Wang, J.; Si, D.; Zhang, Y.; et al. Expression, Purification and Characterization of a Novel Hybrid Peptide Clp with Excellent Antibacterial Activity. Molecules 2021, 26, 7142. [Google Scholar] [CrossRef]
- Klubthawee, N.; Adisakwattana, P.; Hanpithakpong, W.; Somsri, S.; Aunpad, R. A Novel, Rationally Designed, Hybrid Antimicrobial Peptide, Inspired by Cathelicidin and Aurein, Exhibits Membrane-Active Mechanisms against Pseudomonas aeruginosa. Sci. Rep. 2020, 10, 9117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, X.; Zhang, S.Q. Antifungal Mechanism of Antibacterial Peptide, Abp-Cm4, from Bombyx Mori against Aspergillus niger. Biotechnol. Lett. 2008, 30, 2157–2163. [Google Scholar] [CrossRef]
- Li, J.F.; Zhang, J.X.; Li, G.; Xu, Y.Y.; Lu, K.; Wang, Z.G.; Liu, J.P. Antimicrobial Activity and Mechanism of Peptide Cm4 against Pseudomonas aeruginosa. Food Funct. 2020, 11, 7245–7254. [Google Scholar] [CrossRef]
- Lin, Q.P.; Zhou, L.F.; Li, N.N.; Chen, Y.Q.; Li, B.C.; Cai, Y.F.; Zhang, S.Q. Lipopolysaccharide Neutralization by the Antibacterial Peptide Cm4. Eur. J. Pharmacol. 2008, 596, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Kahlenberg, J.M.; Kaplan, M.J. Little Peptide, Big Effects: The Role of Ll-37 in Inflammation and Autoimmune Disease. J. Immunol. 2013, 191, 4895–4901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tjabringa, G.S.; Rabe, K.F.; Hiemstra, P.S. The Human Cathelicidin Ll-37: A Multifunctional Peptide Involved in Infection and Inflammation in the Lung. Pulm. Pharm. 2005, 18, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Murakami, T.; Suzuki, K.; Tamura, H.; Kuwahara-Arai, K.; Iba, T.; Nagaoka, I. Antimicrobial Cathelicidin Peptide Ll-37 Inhibits the Lps/Atp-Induced Pyroptosis of Macrophages by Dual Mechanism. PLoS ONE 2014, 9, e85765. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.C.; Liu, D.; Han, Z.X.; Liang, F.; Hao, C.Y.; Lei, Y.T.; Guo, C.R.; Wang, W.H.; Li, X.H.; Yang, X.N.; et al. Thymopentin-Mediated Inhibition of Cancer Stem Cell Stemness Enhances the Cytotoxic Effect of Oxaliplatin on Colon Cancer Cells. Front. Pharm. 2022, 13, 779715. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, Y.; Meng, Y.; You, Z.; Liu, X.; Liu, Z. The Novel Role of Thymopentin in Induction of Maturation of Bone Marrow Dendritic Cells (Bmdcs). Int. Immunopharmacol. 2014, 21, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Mascartlemone, F.; Huygen, K.; Clumeck, N.; Brenez, D.; Bolla, K.; Duchateau, J. Stimulation of Cellular Function by Thymopentin (Tp-5) in 3 Aids Patients. Lancet 1983, 2, 735–736. [Google Scholar] [CrossRef]
- Sundal, E.; Bertelletti, D. Thymopentin Treatment of Rheumatoid-Arthritis. Arzneimittel-Forsch 1994, 44, 1145–1149. [Google Scholar]
- Clumeck, N.; Cran, S.; Vandeperre, P.; Mascartlemone, F.; Duchateau, J.; Bolla, K. Thymopentin Treatment in Aids and Pre-Aids Patients. Surv. Immunol. Res. 1985, 4, 58–62. [Google Scholar] [CrossRef]
- Cereghino, J.L.; Cregg, J.M. Heterologous Protein Expression in the Methylotrophic Yeast Pichia Pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef]
- Cao, J.; de la Fuente-Nunez, C.; Ou, R.W.; Torres, M.D.; Pande, S.G.; Sinskey, A.J.; Lu, T.K. Yeast-Based Synthetic Biology Platform for Antimicrobial Peptide Production. ACS Synth. Biol. 2018, 7, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, J.; Sun, H.; Li, S.; Chen, T.; Liu, G.; Dyson, P. High-Level Heterologous Production and Functional Secretion by Recombinant Pichia Pastoris of the Shortest Proline-Rich Antibacterial Honeybee Peptide Apidaecin. Sci. Rep. 2017, 7, 14543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Cheng, Q.; Guo, H.; Zhang, R.; Si, D. Expression of Hybrid Peptide Ef-1 in Pichia pastoris, Its Purification, and Antimicrobial Characterization. Molecules 2020, 25, 5538. [Google Scholar] [CrossRef] [PubMed]
- Hong, I.P.; Lee, S.J.; Kim, Y.S.; Choi, S.G. Recombinant Expression of Human Cathelicidin (Hcap18/Ll-37) in Pichia pastoris. Biotechnol. Lett. 2007, 29, 73–78. [Google Scholar] [CrossRef]
- Cheng, J.; Wu, M.; Zhong, R.; Si, D.; Meng, G.; Zhang, R.; Zhang, Y. Rapid Golden Gate Assembly of Exons from Genomic DNA for Protein Expression in Escherichia coli and Pichia pastoris. Biotechniques 2021, 71, 445–450. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial Resistance: A Global Multifaceted Phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Levy, S.B.; Marshall, B. Antibacterial Resistance Worldwide: Causes, Challenges, and Responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef]
- Canton, R.; Morosini, M.I. Emergence and Spread of Antibiotic Resistance Following Exposure to Antibiotics. FEMS Microbiol. Rev. 2011, 35, 977–991. [Google Scholar] [CrossRef] [Green Version]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The Roles of Antimicrobial Peptides in Innate Host Defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef] [Green Version]
- Brandenburg, K.; Heinbockel, L.; Correa, W.; Lohner, K. Peptides with Dual Mode of Action: Killing Bacteria and Preventing Endotoxin-Induced Sepsis. Biochim. Biophys. Acta 2016, 1858, 971–979. [Google Scholar] [CrossRef]
- Harder, J.; Glaser, R.; Schroder, J.M. Human Antimicrobial Proteins Effectors of Innate Immunity. J. Endotoxin Res. 2007, 13, 317–338. [Google Scholar] [CrossRef] [PubMed]
- Ajish, C.; Yang, S.; Kumar, S.D.; Kim, E.Y.; Min, H.J.; Lee, C.W.; Shin, S.H.; Shin, S.Y. A Novel Hybrid Peptide Composed of Lfcinb6 and Kr-12-A4 with Enhanced Antimicrobial, Anti-Inflammatory, and Anti-Biofilm Activities. Sci. Rep. 2022, 12, 4365. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Leishangthem, G.D.; Gill, K.; Singh, A.K.; Das, S.; Singh, K.; Xess, I.; Dinda, A.; Kapil, A.; Patro, I.K.; et al. A Novel Antimicrobial Peptide Derived from Modified N-Terminal Domain of Bovine Lactoferrin: Design, Synthesis, Activity against Multidrug-Resistant Bacteria and Candida. Biochim. Biophys. Acta 2013, 1828, 677–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Vasil, A.I.; Hale, J.D.; Hancock, R.E.; Vasil, M.L.; Hodges, R.S. Effects of Net Charge and the Number of Positively Charged Residues on the Biological Activity of Amphipathic Alpha-Helical Cationic Antimicrobial Peptides. Biopolymers 2008, 90, 369–383. [Google Scholar] [CrossRef]
- Ahmad, B.; Hanif, Q.; Xubiao, W.; Lulu, Z.; Shahid, M.; Dayong, S.; Rijun, Z. Expression and Purification of Hybrid Ll-37talpha1 Peptide in Pichia Pastoris and Evaluation of Its Immunomodulatory and Anti-Inflammatory Activities by Lps Neutralization. Front. Immunol. 2019, 10, 1365. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, L.; Zhang, R.; Koci, M.; Si, D.; Ahmad, B.; Cheng, J.; Wang, J.; Aihemaiti, M.; Zhang, M. A Novel Cecropin-Ll37 Hybrid Peptide Protects Mice against Ehec Infection-Mediated Changes in Gut Microbiota, Intestinal Inflammation, and Impairment of Mucosal Barrier Functions. Front. Immunol. 2020, 11, 1361. [Google Scholar] [CrossRef]
- Wei, X.B.; Wu, R.J.; Si, D.Y.; Liao, X.D.; Zhang, L.L.; Zhang, R.J. Novel Hybrid Peptide Cecropin a (1-8)-Ll37 (17-30) with Potential Antibacterial Activity. Int. J. Mol. Sci. 2016, 17, 983. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.N.; Tong, Y.C.; Wang, H.L.; Zhang, J.; Li, Z.X.; Abbas, Z.; Yang, T.T.; Liu, M.Y.; Chen, P.Y.; Hua, Z.C.; et al. Novel Hybrid Peptide Cathelicidin 2 (1-13)-Thymopentin (Tp5) and Its Derived Peptides with Effective Antibacterial, Antibiofilm, and Anti-Adhesion Activities. Int. J. Mol. Sci. 2021, 22, 1681. [Google Scholar] [CrossRef]
- Jiang, X.; Qian, K.; Liu, G.; Sun, L.; Zhou, G.; Li, J.; Fang, X.; Ge, H.; Lv, Z. Design and Activity Study of a Melittin-Thanatin Hybrid Peptide. AMB Express 2019, 9, 14. [Google Scholar] [CrossRef]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia Pastoris: A Highly Successful Expression System for Optimal Synthesis of Heterologous Proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef]
- Lin-Cereghino, G.P.; Stark, C.M.; Kim, D.; Chang, J.; Shaheen, N.; Poerwanto, H.; Agari, K.; Moua, P.; Low, L.K.; Tran, N.; et al. The Effect of Alpha-Mating Factor Secretion Signal Mutations on Recombinant Protein Expression in Pichia Pastoris. Gene 2013, 519, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Rueda, F.; Gasser, B.; Sánchez-Chardi, A.; Roldán, M.; Villegas, S.; Puxbaum, V.; Ferrer-Miralles, N.; Unzueta, U.; Vázquez, E.; Garcia-Fruitós, E.; et al. Functional Inclusion Bodies Produced in the Yeast Pichia Pastoris. Microb. Cell Fact. 2016, 15, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.; Huang, Y.; Zheng, H.; Tang, L.; He, J.; Xiang, L.; Liu, D.; Jiang, H. Secretion and Activity of Antimicrobial Peptide Cecropin D Expressed in Pichia Pastoris. Exp. Med. 2012, 4, 1063–1068. [Google Scholar] [CrossRef] [Green Version]
- Popa, C.; Shi, X.; Ruiz, T.; Ferrer, P.; Coca, M. Biotechnological Production of the Cell Penetrating Antifungal Paf102 Peptide in Pichia Pastoris. Front. Microbiol. 2019, 10, 1472. [Google Scholar] [CrossRef] [PubMed]
- Mulder, K.C.; de Lima, L.A.; Aguiar, P.S.; Carneiro, F.C.; Franco, O.L.; Dias, S.C.; Parachin, N.S. Production of a Modified Peptide Clavanin in Pichia Pastoris: Cloning, Expression, Purification and in Vitro Activities. AMB Express 2015, 5, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbulu, S.; Jimenez, J.J.; Gutiez, L.; Cintas, L.M.; Herranz, C.; Hernandez, P.E. Cloning and Expression of Synthetic Genes Encoding the Broad Antimicrobial Spectrum Bacteriocins Srcam 602, or-7, E-760, and L-1077, by Recombinant Pichia Pastoris. Biomed. Res. Int. 2015, 2015, 767183. [Google Scholar] [CrossRef] [Green Version]
- Basanta, A.; Gómez-Sala, B.; Sánchez, J.; Diep, D.B.; Herranz, C.; Hernández, P.E.; Cintas, L.M. Use of the Yeast Pichia Pastoris as an Expression Host for Secretion of Enterocin L50, a Leaderless Two-Peptide (L50a and L50b) Bacteriocin from Enterococcus Faecium L50. Appl. Env. Microbiol. 2010, 76, 3314–3324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, S.Q.; Wu, X.; Chen, Y.Q.; Diao, Z.Y. Expression and Characterization of Antimicrobial Peptide Abp-Cm4 in Methylotrophic Yeast Pichia pastoris. Process Biochem. 2006, 41, 251–256. [Google Scholar] [CrossRef]
- Meng, D.M.; Li, W.J.; Shi, L.Y.; Lv, Y.J.; Sun, X.Q.; Hu, J.C.; Fan, Z.C. Expression, Purification and Characterization of a Recombinant Antimicrobial Peptide Hispidalin in Pichia Pastoris. Protein Expr. Purif. 2019, 160, 19–27. [Google Scholar] [CrossRef]
- Schaumberger, S.; Ladinig, A.; Reisinger, N.; Ritzmann, M.; Schatzmayr, G. Evaluation of the Endotoxin Binding Efficiency of Clay Minerals Using the Limulus Amebocyte Lysate Test: An in Vitro Study. Amb Express 2014, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Oren, Z.; Lerman, J.C.; Gudmundsson, G.H.; Agerberth, B.; Shai, Y. Structure and Organization of the Human Antimicrobial Peptide Ll-37 in Phospholipid Membranes: Relevance to the Molecular Basis for Its Non-Cell-Selective Activity. Biochem. J. 1999, 341, 501–513. [Google Scholar] [CrossRef]
- Kaconis, Y.; Kowalski, I.; Howe, J.; Brauser, A.; Richter, W.; Razquin-Olazarán, I.; Inigo-Pestana, M.; Garidel, P.; Rössle, M.; de Tejada, G.M.; et al. Biophysical Mechanisms of Endotoxin Neutralization by Cationic Amphiphilic Peptides. Biophys. J. 2011, 100, 2652–2661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.H.; Mann, D.M.; Tsai, C.M. Neutralization of Endotoxin in Vitro and in Vivo by a Human Lactoferrin-Derived Peptide. Infect. Immun. 1999, 67, 1353–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.K.; Seo, C.H.; Luchian, T.; Park, Y. Antimicrobial Peptide Cma3 Derived from the Ca-Ma Hybrid Peptide: Antibacterial and Anti-Inflammatory Activities with Low Cytotoxicity and Mechanism of Action in Escherichia coli. Antimicrob. Agents Chemother. 2016, 60, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Harten, R.M.; van Woudenbergh, E.; van Dijk, A.; Haagsman, H.P. Cathelicidins: Immunomodulatory Antimicrobials. Vaccines 2018, 6, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, G. Gram-Positive and Gram-Negative Bacterial Toxins in Sepsis: A Brief Review. Virulence 2014, 5, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Minasyan, H. Sepsis: Mechanisms of Bacterial Injury to the Patient. Scand. J. Trauma Resusc. Emerg. Med. 2019, 27, 19. [Google Scholar] [CrossRef] [Green Version]
- Kell, D.B.; Pretorius, E. On the Translocation of Bacteria and Their Lipopolysaccharides between Blood and Peripheral Locations in Chronic, Inflammatory Diseases: The Central Roles of Lps and Lps-Induced Cell Death. Integr. Biol. 2015, 7, 1339–1377. [Google Scholar] [CrossRef] [Green Version]
- Niedbala, W.; Cai, B.; Liew, F.Y. Role of Nitric Oxide in the Regulation of T Cell Functions. Ann. Rheum. Dis. 2006, 65 (Suppl. 3), iii37–iii40. [Google Scholar] [CrossRef] [Green Version]
- Kolios, G.; Valatas, V.; Ward, S.G. Nitric Oxide in Inflammatory Bowel Disease: A Universal Messenger in an Unsolved Puzzle. Immunology 2004, 113, 427–437. [Google Scholar] [CrossRef]
- Abramson, S.B. Nitric Oxide in Inflammation and Pain Associated with Osteoarthritis. Arthritis Res. Therapy 2008, 10 (Suppl 2), S2. [Google Scholar] [CrossRef] [PubMed]
- Lind, L. Circulating Markers of Inflammation and Atherosclerosis. Atherosclerosis 2003, 169, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Nadia, E.; Yao, Y.; Shi, Z.; Ren, G. Tandem Repeated Expression of Lunasin Gene in Pichia Pastoris and Its Anti-Inflammatory Activity in Vitro. J. Biosci. Bioeng. 2018, 126, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.-B.; Je, J.-Y.; Cho, Y.-S. Antioxidant and Anti-Inflammatory Peptide Fraction from Salmon Byproduct Protein Hydrolysates by Peptic Hydrolysis. Food Res. Int. 2012, 49, 92–98. [Google Scholar] [CrossRef]
- Mookherjee, N.; Hancock, R.E. Cationic Host Defence Peptides: Innate Immune Regulatory Peptides as a Novel Approach for Treating Infections. Cell. Mol. Life Sci. 2007, 64, 922–933. [Google Scholar] [CrossRef]
- Nan, Y.H.; Bang, J.K.; Jacob, B.; Park, I.S.; Shin, S.Y. Prokaryotic Selectivity and Lps-Neutralizing Activity of Short Antimicrobial Peptides Designed from the Human Antimicrobial Peptide Ll-37. Peptides 2012, 35, 239–247. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, J.; Ahmad, B.; Raza, M.A.; Guo, H.; Ahmat, M.; Wei, X.; Zhang, L.; Li, Z.; Cheng, Q.; Zhang, J.; et al. Yeast Expressed Hybrid Peptide CLP Abridged Pro-Inflammatory Cytokine Levels by Endotoxin Neutralization. Microorganisms 2023, 11, 131. https://doi.org/10.3390/microorganisms11010131
Cheng J, Ahmad B, Raza MA, Guo H, Ahmat M, Wei X, Zhang L, Li Z, Cheng Q, Zhang J, et al. Yeast Expressed Hybrid Peptide CLP Abridged Pro-Inflammatory Cytokine Levels by Endotoxin Neutralization. Microorganisms. 2023; 11(1):131. https://doi.org/10.3390/microorganisms11010131
Chicago/Turabian StyleCheng, Junhao, Baseer Ahmad, Muhammad Asif Raza, Henan Guo, Marhaba Ahmat, Xubiao Wei, Lulu Zhang, Zhongxuan Li, Qiang Cheng, Jing Zhang, and et al. 2023. "Yeast Expressed Hybrid Peptide CLP Abridged Pro-Inflammatory Cytokine Levels by Endotoxin Neutralization" Microorganisms 11, no. 1: 131. https://doi.org/10.3390/microorganisms11010131