
A Chloroplast-Localised Fluorescent Protein Enhances the Photosynthetic Action Spectrum in Green Algae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chlamydomonas Strains and Media
2.2. Nucleic Acid Manipulations
2.3. Synthesis of the CpKat Gene
2.4. Construction of the pB10 Chloroplast Transformation Vector
2.5. Chloroplast Transformation
2.6. PCR Analyses of Microalgae
2.7. Protein Blot Analyses
2.8. Measurement of Dissolved Oxygen
2.9. Algal Growth Conditions
2.10. Fluorescence and Spectral Analysis
3. Results
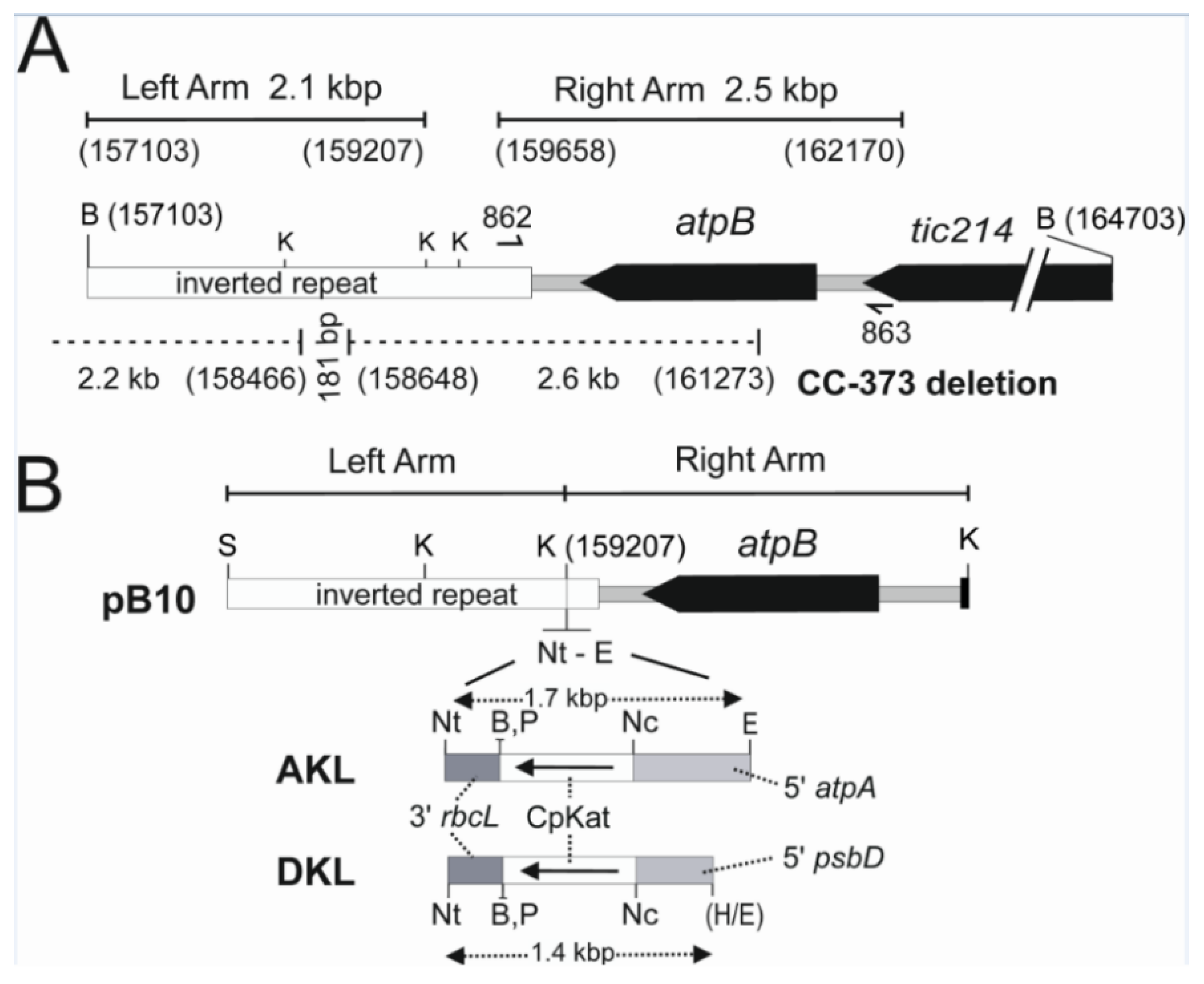
3.1. Design of Chloroplast Transformation Vectors
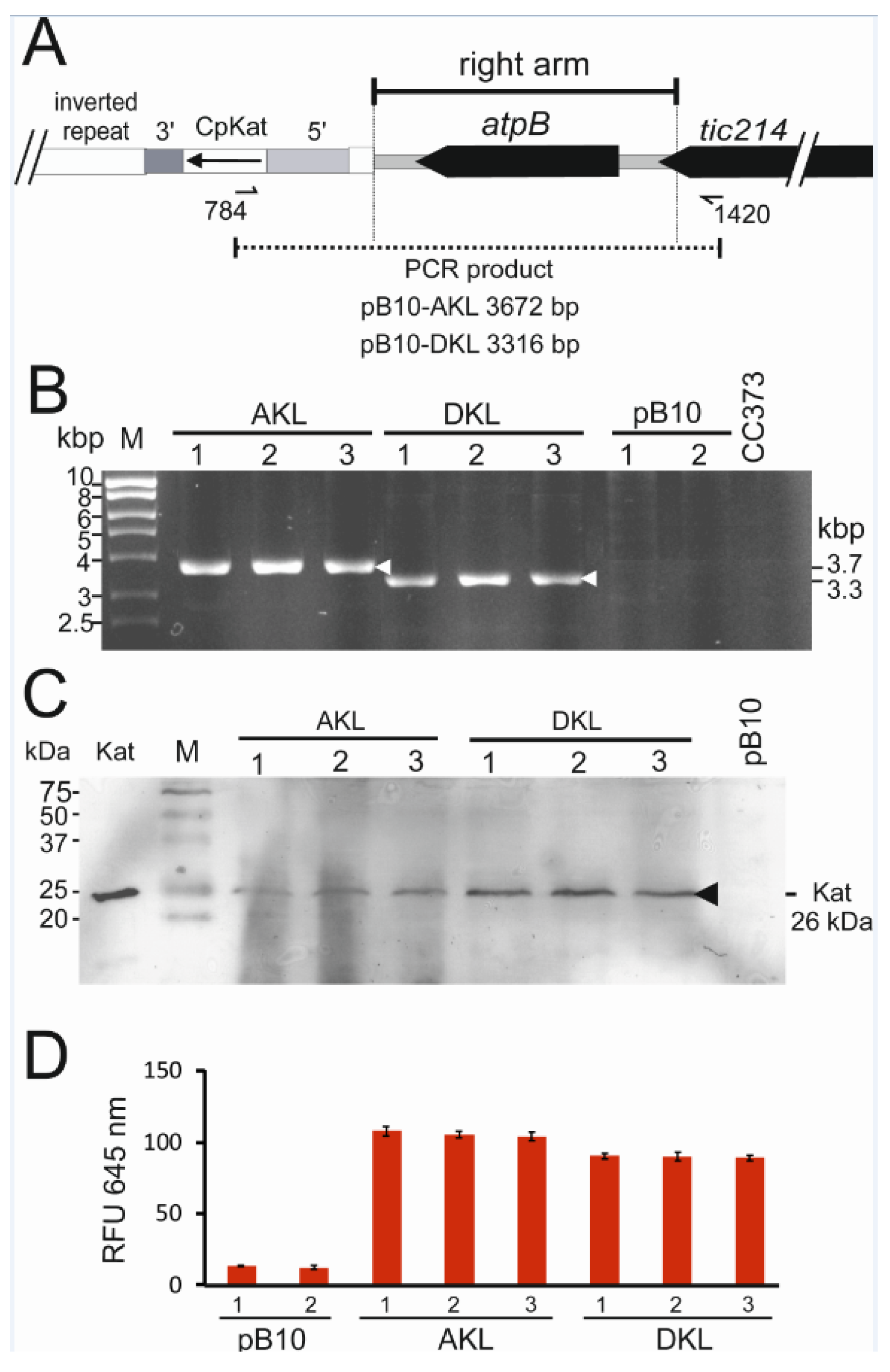
3.2. Isolation of CpKat Chloroplast Transformants Expressing Katushka
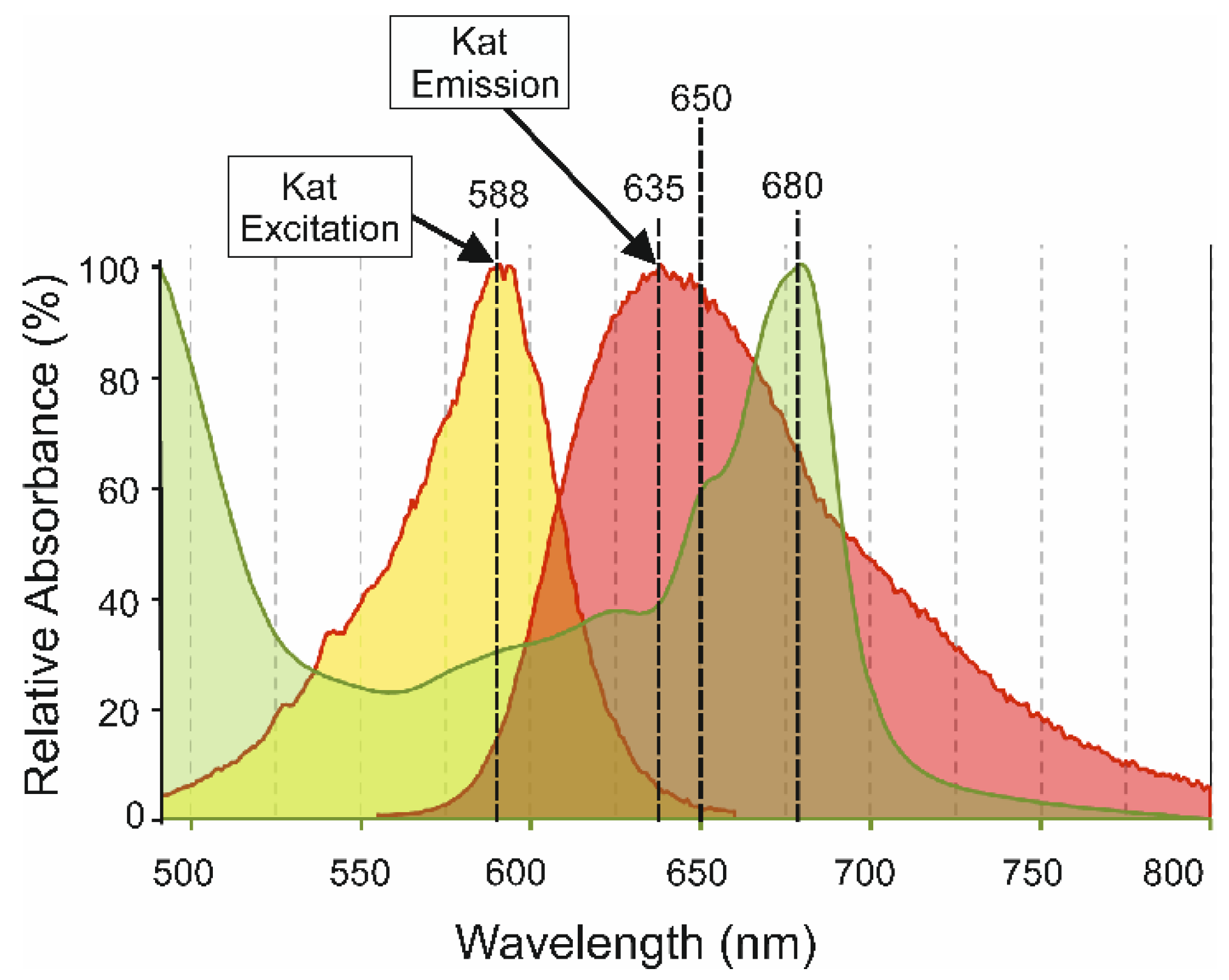
3.3. Chlorophyll Absorption of Katushka Fluorescence
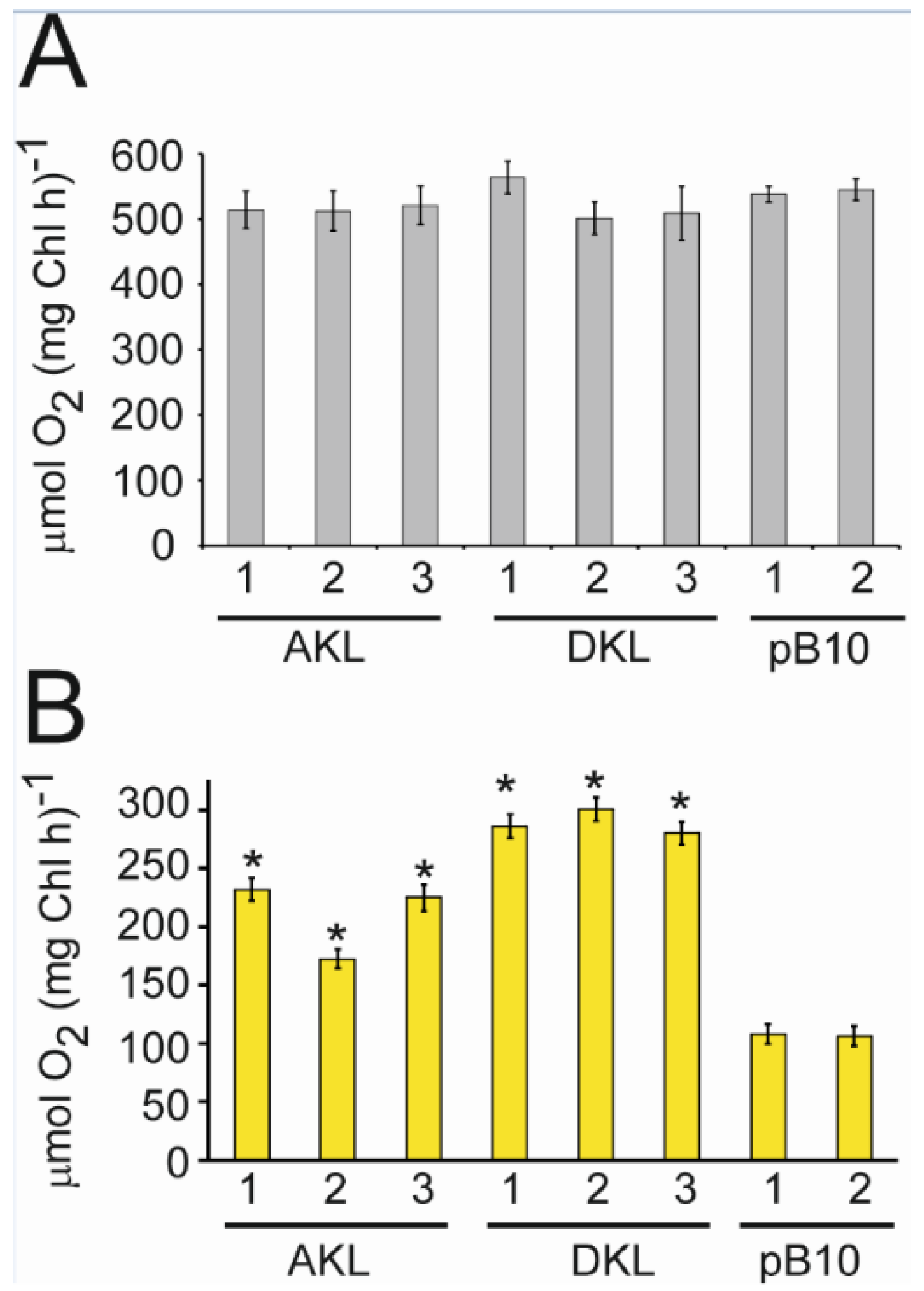
3.4. CpKat Chloroplast Transformants Use Yellow Light for Photosynthesis
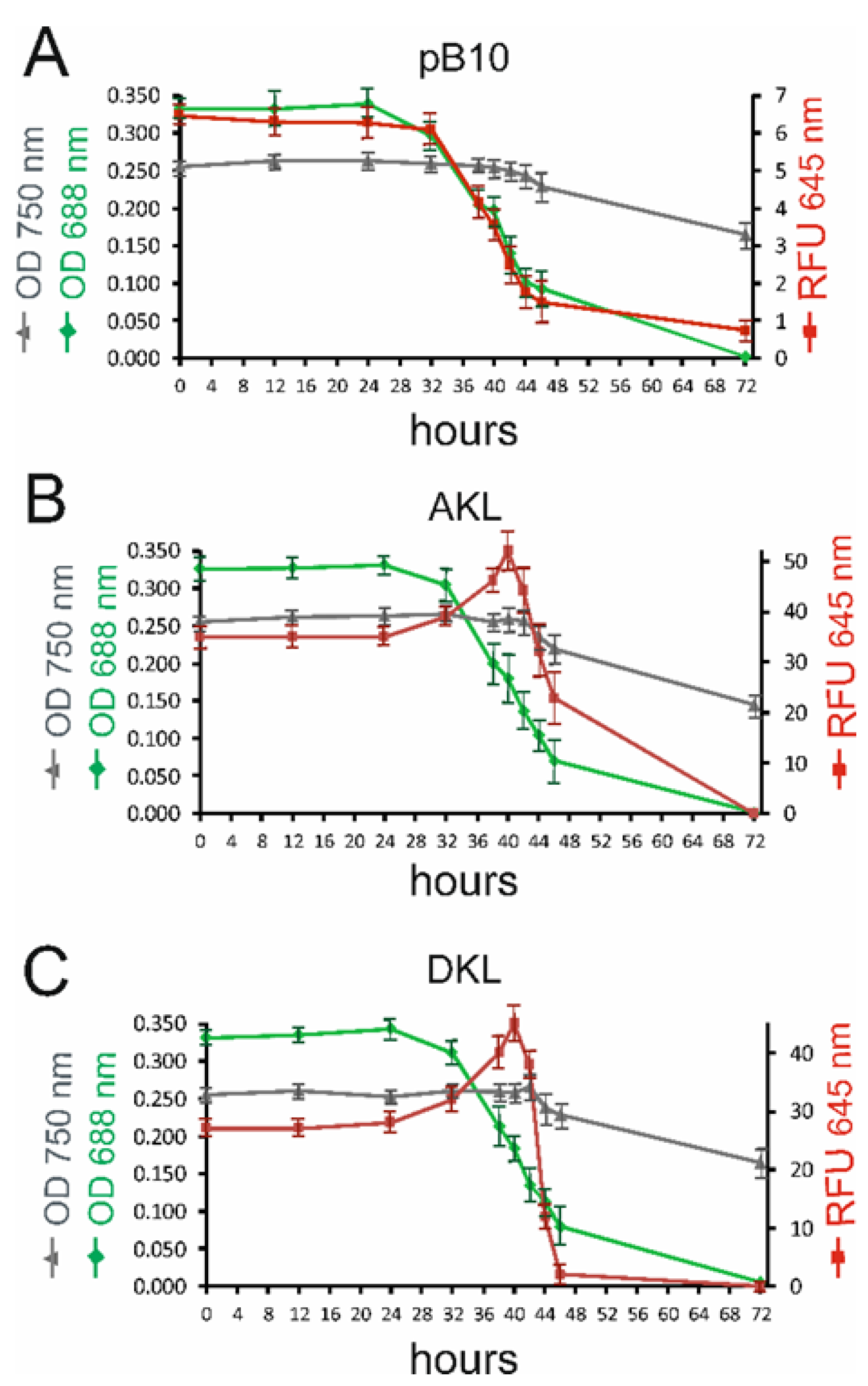
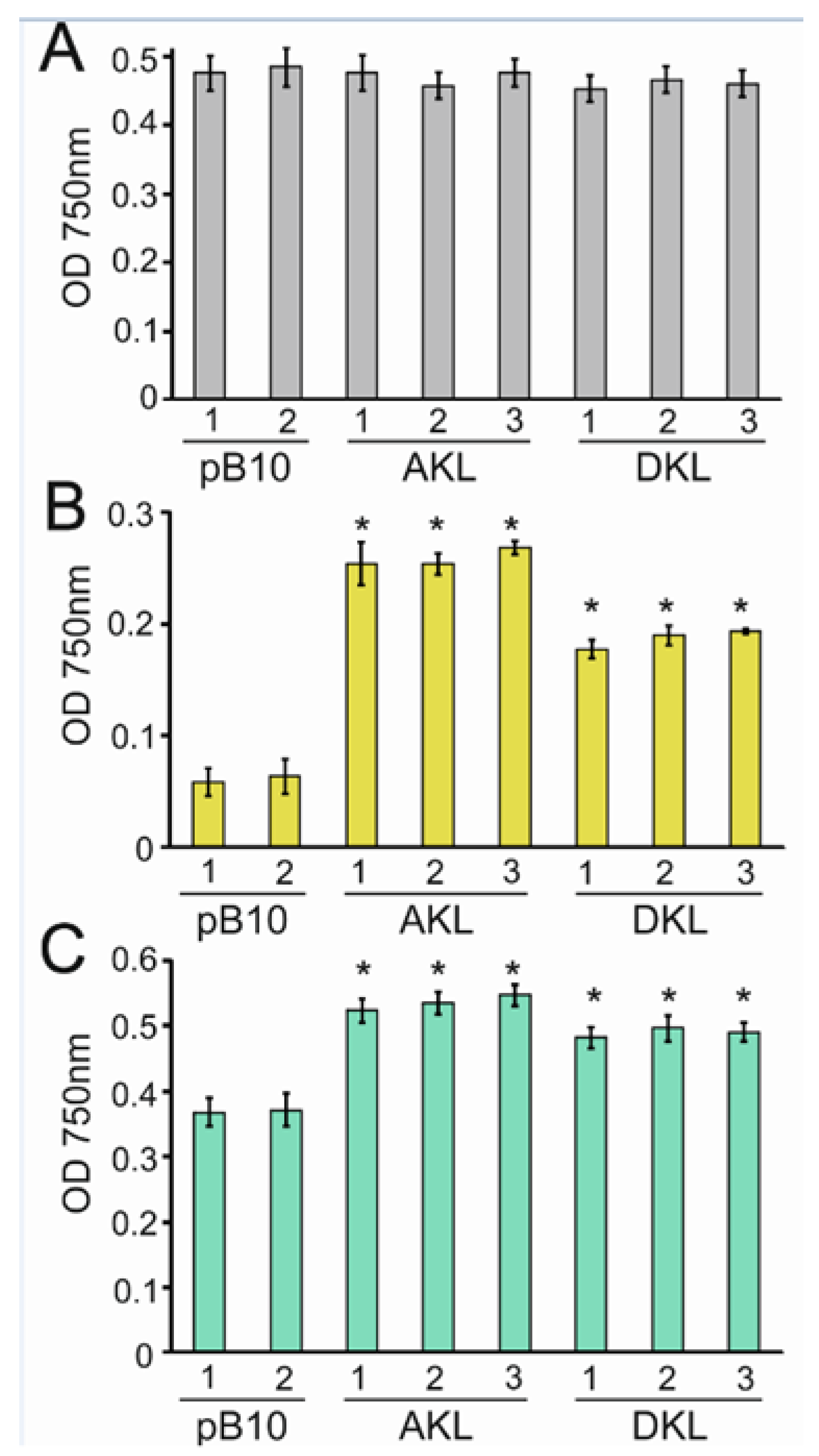
3.5. Yellow Light Promotes the Growth of Transplastomic Lines Expressing Katushka
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yoon, H.; Hackett, J.; Ciniglia, C.; Pinto, G.; Bhattacharya, D. A molecular timeline for the origin of photosynthetic eukaryotes. Mol. Biol. Evol. 2004, 21, 809–818. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef] [PubMed]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for high-value products towards human health and nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef] [PubMed]
- Georgianna, D.; Mayfield, S. Exploiting diversity and synthetic biology for the production of algal biofuels. Nature 2012, 488, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Merchant, S.; Kropat, J.; Liu, B.; Shaw, J.; Warakanont, J. TAG, You’re it! Chlamydomonas as a reference organism for understanding algal triacylglycerol accumulation. Curr. Opin. Biotechnol. 2012, 23, 352–363. [Google Scholar] [CrossRef]
- Mondal, M.; Goswami, S.; Ghosh, A.; Oinam, G.; Tiwari, O.N.; Das, P.; Gayen, K.; Mandal, M.K.; Halder, G.N. Production of biodiesel from microalgae through biological carbon capture: A review. 3 Biotech 2017, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Taunt, H.N.; Stoffels, L.; Purton, S. Green biologics: The algal chloroplast as a platform for making biopharmaceuticals. Bioengineered 2018, 9, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Cutolo, E.A.; Mandalà, G.; Dall’Osto, L.; Bassi, R. Harnessing the algal chloroplast for heterologous protein production. Microorganisms 2022, 10, 743. [Google Scholar] [CrossRef] [PubMed]
- Naduthodi, M.I.S.; Claassens, N.J.; Adamo, S.D.; van der Oost, J.; Barbosa, M.J. Synthetic biology approaches to enhance microalgal productivity. Trends Biotechnol. 2021, 39, 1019–1036. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Silva, S.O.; Baptista, J.M.; Malcata, F.X. Light requirements in microalgal photobioreactors: An overview of biophotonic aspects. Appl. Microbiol. Biotechnol. 2011, 89, 1275–1288. [Google Scholar] [CrossRef]
- Dixon, R.; Cheng, Q.; Shen, G.F.; Day, A.; DowsonDay, M. Nif gene transfer and expression in chloroplasts: Prospects and problems. Plant Soil 1997, 194, 193–203. [Google Scholar] [CrossRef]
- Cheng, Q.; Day, A.; Dowson-Day, M.; Shen, G.; Dixon, R. The Klebsiella pneumoniae nitrogenase Fe protein gene (nifH) functionally substitutes for the chlL gene in Chlamydomonas reinhardtii. Biochem. Biophys. Res. Commun. 2005, 329, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Jackson, H.O.; Taunt, H.N.; Mordaka, P.M.; Smith, A.G.; Purton, S. The algal chloroplast as a testbed for synthetic biology designs aimed at radically rewiring plant metabolism. Front. Plant Sci. 2021, 12, 708370. [Google Scholar] [CrossRef] [PubMed]
- Perrine, Z.; Negi, S.; Sayre, R.T. Optimization of photosynthetic light energy utilization by microalgae. Algal Res. Biomass Biofuels Bioprod. 2012, 1, 134–142. [Google Scholar] [CrossRef]
- Blankenship, R.E.; Chen, M. Spectral expansion and antenna reduction can enhance photosynthesis for energy production. Curr. Opin. Chem. Biol. 2013, 17, 457–461. [Google Scholar] [CrossRef]
- Croce, R.; van Amerongen, H. Natural strategies for photosynthetic light harvesting. Nat. Chem. Biol. 2014, 10, 492–501. [Google Scholar] [CrossRef]
- Negi, S.; Perrine, Z.; Friedland, N.; Kumar, A.; Tokutsu, R.; Minagawa, J.; Berg, H.; Barry, A.N.; Govindjee, G.; Sayre, R. Light regulation of light-harvesting antenna size substantially enhances photosynthetic efficiency and biomass yield in green algae. Plant J. 2020, 103, 584–603. [Google Scholar] [CrossRef]
- Hitchcock, A.; Hunter, C.N.; Sobotka, R.; Komenda, J.; Dann, M.; Leister, D. Redesigning the photosynthetic light reactions to enhance photosynthesis-the PhotoRedesign consortium. Plant J. 2022, 109, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Blankenship, R.E. Expanding the solar spectrum used by photosynthesis. Trends Plant Sci. 2011, 16, 427–431. [Google Scholar] [CrossRef]
- Blankenship, R.E.; Tiede, D.M.; Barber, J.; Brudvig, G.W.; Fleming, G.; Ghirardi, M.; Gunner, M.R.; Junge, W.; Kramer, D.M.; Melis, A.; et al. Comparing photosynthetic and photovoltaic efficiencies and recognizing the potential for improvement. Science 2011, 332, 805–809. [Google Scholar] [CrossRef] [Green Version]
- Ho, M.Y.; Shen, G.Z.; Canniffe, D.P.; Zhao, C.; Bryant, D.A. Light-dependent chlorophyll f synthase is a highly divergent paralog of PsbA of photosystem II. Science 2016, 353, aaf9178. [Google Scholar] [CrossRef] [PubMed]
- Trinugroho, J.P.; Beckova, M.; Shao, S.X.; Yu, J.F.; Zhao, Z.Y.; Murray, J.W.; Sobotka, R.; Komenda, J.; Nixon, P.J. Chlorophyll f synthesis by a super-rogue photosystem II complex. Nat. Plants 2020, 6, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y.; Aikawa, S.; Kondo, A.; Akimoto, S. Adaptation of light-harvesting functions of unicellular green algae to different light qualities. Photosyn Res. 2019, 139, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Kageshima, Y.; Shinagawa, T.; Kuwata, T.; Nakata, J.; Minegishi, T.; Takanabe, K.; Domen, K. A miniature solar device for overall water splitting consisting of series-connected spherical silicon solar cells. Sci. Rep. 2016, 6, 24633. [Google Scholar] [CrossRef]
- Schlichter, D.; Fricke, H. Coral host improves photosynthesis of endosymbiotic algae. Naturwissenschaften 1990, 77, 447–450. [Google Scholar] [CrossRef]
- Smith, E.G.; D’Angelo, C.; Sharon, Y.; Tchernov, D.; Wiedenmann, J. Acclimatization of symbiotic corals to mesophotic light environments through wavelength transformation by fluorescent protein pigments. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170320. [Google Scholar] [CrossRef]
- Aihara, Y.; Maruyama, S.; Baird, A.H.; Iguchi, A.; Takahashi, S.; Minagawa, J. Green fluorescence from cnidarian hosts attracts symbiotic algae. Proc. Natl. Acad. Sci. USA 2019, 116, 2118–2123. [Google Scholar] [CrossRef]
- Ben-Zvi, O.; Lindemann, Y.; Eyal, G.; Loya, Y. Coral fluorescence: A prey-lure in deep habitats. Commun. Biol. 2022, 5, 537. [Google Scholar] [CrossRef]
- Lambert, T.J. FPbase: A community-editable fluorescent protein database. Nat. Methods 2019, 16, 277–278. [Google Scholar] [CrossRef]
- Shcherbo, D.; Merzlyak, E.; Chepurnykh, T.; Fradkov, A.; Ermakova, G.; Solovieva, E.; Lukyanov, K.; Bogdanova, E.; Zaraisky, A.; Lukyanov, S.; et al. Bright far-red fluorescent protein for whole-body imaging. Nat. Methods 2007, 4, 741–746. [Google Scholar] [CrossRef]
- Wang, L.; Jackson, W.; Steinbach, P.; Tsien, R. Evolution of new nonantibody proteins via iterative somatic hypermutation. Proc. Natl. Acad. Sci. USA 2004, 101, 16745–16749. [Google Scholar] [CrossRef] [PubMed]
- Myers, A.; Grant, D.; Rabert, D.; Harris, E.; Boynton, J.; Gillham, N. Mutants of Chlamydomonas reinhardtii with physical alterations in their chloroplast DNA. Plasmid 1982, 7, 133–151. [Google Scholar] [CrossRef]
- DeShaw, A.E.; Figueroa-Martinez, F.; Reyes-Prieto, A. Complete chloroplast genomes of the Chlamydomonas reinhardtii nonphotosynthetic mutants CC-1375, CC-373, CC-4199, CC-2359 and CC-1051. Mitochondrial DNA B Resour. 2017, 2, 405–407. [Google Scholar] [CrossRef]
- Harris, E.H. The Chlamydomonas Sourcebook; Academic Press: London, UK, 1989. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.; Maniatis, T. Molecular Cloning: A Laboratory Manual (3 Volume Set), 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Day, A.; Schirmerrahire, M.; Kuchka, M.R.; Mayfield, S.P.; Rochaix, J.D. A transposon with an unusual arrangement of long terminal repeats in the green alga Chlamydomonas reinhardtii. EMBO J. 1988, 7, 1917–1927. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Maul, J.; Lilly, J.; Cui, L.; dePamphilis, C.; Miller, W.; Harris, E.; Stern, D. The Chlamydomonas reinhardtti plastid chromosome: Islands of genes in a sea of repeats. Plant Cell 2002, 14, 2659–2679. [Google Scholar] [CrossRef]
- Rochaix, J.D. Restriction endonuclease map of the chloroplast DNA of Chlamydomonas reinhardtii. J. Mol. Biol. 1978, 126, 597–617. [Google Scholar] [CrossRef]
- Altingmees, M.; Short, J. pBluescript II: Gene mapping vectors. Nucleic Acids Res. 1989, 17, 9494. [Google Scholar] [CrossRef]
- Yanisch-Perron, C.; Vieira, J.; Messing, J. Improved M13 phage cloning vectors and host strains: Nucleotide sequences of the M13mp18 and pUC19 vectors. Gene 1985, 33, 103–119. [Google Scholar] [CrossRef]
- Goldschmidt-Clermont, M. Transgenic expression of aminoglycoside adenine transferase in the chloroplast: A selectable marker for site-directed transformation of Chlamydomonas. Nucleic Acids Res. 1991, 19, 4083–4089. [Google Scholar] [CrossRef]
- Boynton, J.; Gillham, N.; Harris, E.; Hosler, J.; Johnson, A.; Jones, A.; Randolphanderson, B.; Robertson, D.; Klein, T.; Shark, K.; et al. Chloroplast transformation in Chlamydomonas with high-velocity microprojectiles. Science 1988, 240, 1534–1538. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Fu, Y.; Guo, Y.; Pan, J. Chlamydomonas (Chlorophyceae) colony PCR. Protoplasma 2009, 235, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Suarez, J.V.; Banks, S.; Thomas, P.G.; Day, A. A new F131V mutation in Chlamydomonas phytoene desaturase locates a cluster of norflurazon resistance mutations near the FAD-binding site in 3D protein models. PLoS ONE 2014, 9, e99894. [Google Scholar] [CrossRef]
- Carpenter, J.H. The accuracy of the winkler method for dissolved oxygen analysis 1. Limnol. Oceanogr. 1965, 10, 135–140. [Google Scholar] [CrossRef]
- Labasque, T.; Chaumery, C.; Aminot, A.; Kergoat, G. Spectrophotometric Winkler determination of dissolved oxygen: Re-examination of critical factors and reliability. Mar. Chem. 2004, 88, 53–60. [Google Scholar] [CrossRef]
- Ritchie, R.J. Consistent sets of spectrophotometric chlorophyll equations for acetone, methanol and ethanol solvents. Photosynth. Res. 2006, 89, 27–41. [Google Scholar] [CrossRef]
- Day, A.; Goldschmidt-Clermont, M. The chloroplast transformation toolbox: Selectable markers and marker removal. Plant Biotechnol. J. 2011, 9, 540–553. [Google Scholar] [CrossRef]
- Pletneva, N.V.; Pletnev, V.Z.; Shemiakina, I.I.; Chudakov, D.M.; Artemyev, I.; Wlodawer, A.; Dauter, Z.; Pletnev, S. Crystallographic study of red fluorescent protein eqFP578 and its far-red variant Katushka reveals opposite pH-induced isomerization of chromophore. Protein Sci. 2011, 20, 1265–1274. [Google Scholar] [CrossRef]
- Oldenhof, H.; Zachleder, V.; Van den Ende, H. Blue- and red-light regulation of the cell cycle in Chlamydomonas reinhardtii (Chlorophyta). Eur. J. Phycol. 2006, 41, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.Y.; Sun, H.; Deng, J.Q.; Zhang, Y.; Li, Y.L.; Huang, J.C.; Chen, F. Coordinating carbon metabolism and cell cycle of Chlamydomonas reinhardtii with light strategies under nitrogen recovery. Microorganisms 2021, 9, 2480. [Google Scholar] [CrossRef]
- Heining, M.; Buchholz, R. Photobioreactors with internal illumination-A survey and comparison. Biotechnol. J. 2015, 10, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Swift, T.A.; Oliver, T.A.A.; Galan, M.C.; Whitney, H.M. Functional nanomaterials to augment photosynthesis: Evidence and considerations for their responsible use in agricultural applications. Interface Focus 2019, 9, 20180048. [Google Scholar] [CrossRef] [PubMed]
- Rasala, B.; Barrera, D.; Ng, J.; Plucinak, T.; Rosenberg, J.; Weeks, D.; Oyler, G.; Peterson, T.; Haerizadeh, F.; Mayfield, S. Expanding the spectral palette of fluorescent proteins for the green microalga Chlamydomonas reinhardtii. Plant J. 2013, 74, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, M.; Oertel, W.; Hegemann, P. A synthetic gene coding for the green fluorescent protein (GFP) is a versatile reporter in Chlamydomonas reinhardtii. Plant J. 1999, 19, 353–361. [Google Scholar] [CrossRef]
- Franklin, S.; Ngo, B.; Efuet, E.; Mayfield, S. Development of a GFP reporter gene for Chlamydomonas reinhardtii chloroplast. Plant J. 2002, 30, 733–744. [Google Scholar] [CrossRef]
- Michelet, L.; Lefebvre-Legendre, L.; Burr, S.E.; Rochaix, J.D.; Goldschmidt-Clermont, M. Enhanced chloroplast transgene expression in a nuclear mutant of Chlamydomonas. Plant Biotechnol. J. 2011, 9, 565–574. [Google Scholar] [CrossRef]
- Barnes, D.; Franklin, S.; Schultz, J.; Henry, R.; Brown, E.; Coragliotti, A.; Mayfield, S. Contribution of 5′- and 3′-untranslated regions of plastid mRNAs to the expression of Chlamydomonas reinhardtii chloroplast genes. Mol. Genet. Genom. 2005, 274, 625–636. [Google Scholar] [CrossRef]
- Jares-Erijman, E.; Jovin, T. FRET imaging. Nat. Biotechnol. 2003, 21, 1387–1395. [Google Scholar] [CrossRef]
- Schneider, A.; Geissler, P. Coexistence of fluid and crystalline phases of proteins in photosynthetic membranes. Biophys. J. 2013, 105, 1161–1170. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suarez, J.V.; Mudd, E.A.; Day, A. A Chloroplast-Localised Fluorescent Protein Enhances the Photosynthetic Action Spectrum in Green Algae. Microorganisms 2022, 10, 1770. https://doi.org/10.3390/microorganisms10091770
Suarez JV, Mudd EA, Day A. A Chloroplast-Localised Fluorescent Protein Enhances the Photosynthetic Action Spectrum in Green Algae. Microorganisms. 2022; 10(9):1770. https://doi.org/10.3390/microorganisms10091770
Chicago/Turabian StyleSuarez, Julio V., Elisabeth A. Mudd, and Anil Day. 2022. "A Chloroplast-Localised Fluorescent Protein Enhances the Photosynthetic Action Spectrum in Green Algae" Microorganisms 10, no. 9: 1770. https://doi.org/10.3390/microorganisms10091770