
Roles of Arbuscular mycorrhizal Fungi as a Biocontrol Agent in the Control of Plant Diseases

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Biocontrol of AMF against Phytopathogenic Fungi
3. Biocontrol of AMF against Phytopathogenic Bacteria and Nematodes
4. AMF and Beneficial Microorganisms Combine to Control Plant Diseases
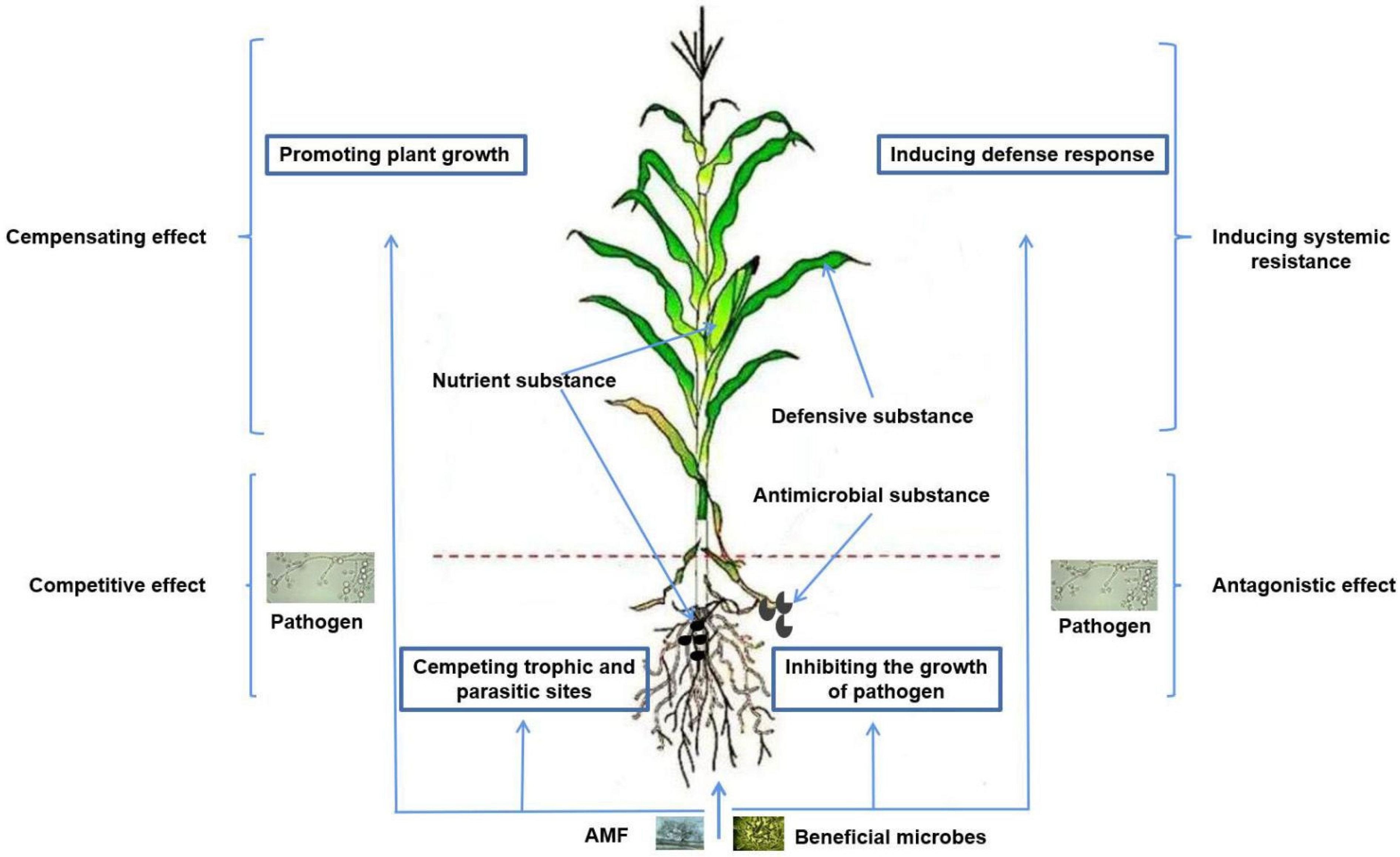
5. The Mechanism of AMF Improving Plant Disease Resistance
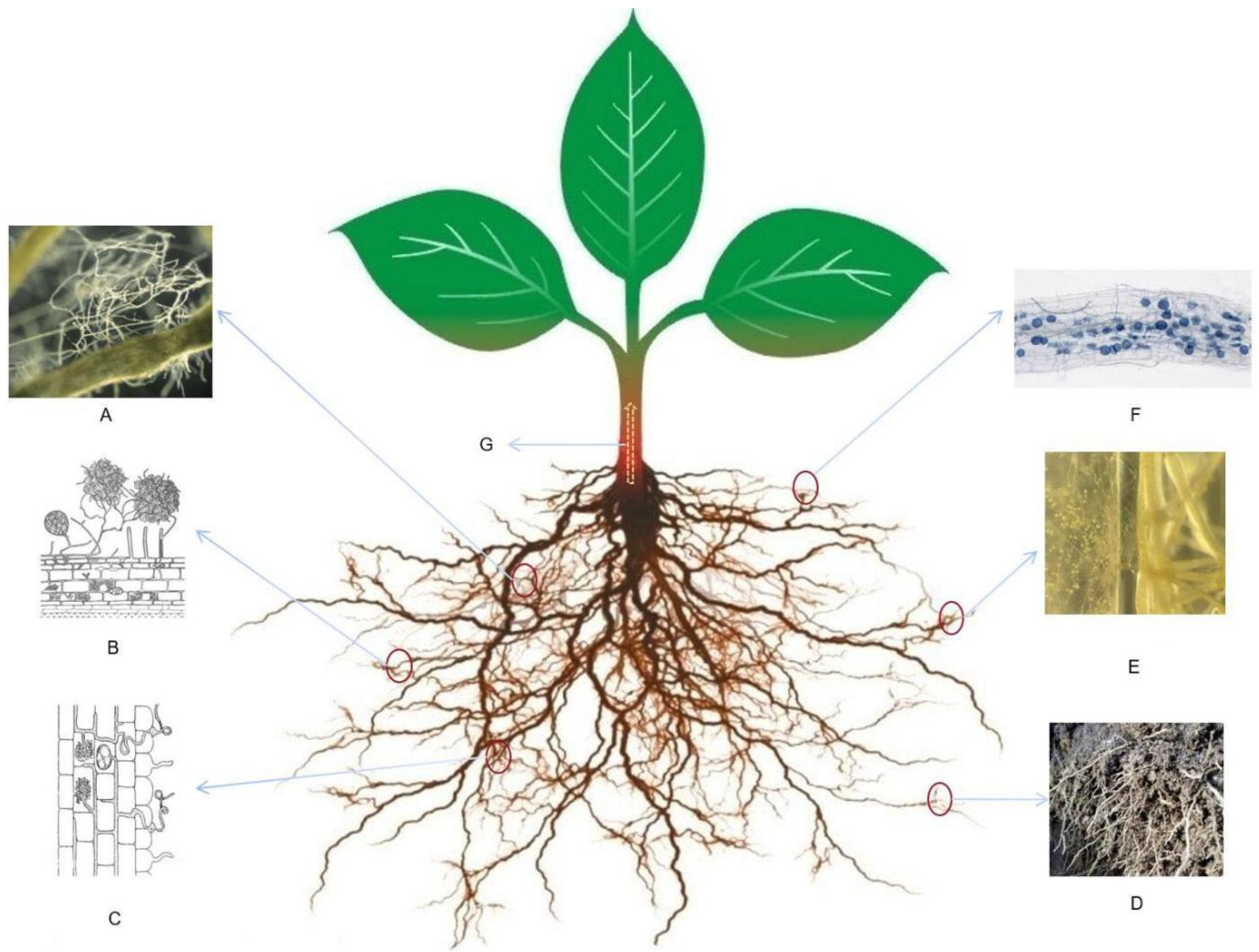
5.1. AMF Regulate Plant Root Morphological Structure
5.2. AMF Improves Plant Nutrition
5.3. AMF Regulate the Synthesis of Plant Secondary Metabolites
5.4. Regulation of Microbial Populations in the Rhizosphere of Plants
5.5. AMF Compete with Pathogenic Microorganisms for Invasion Sites
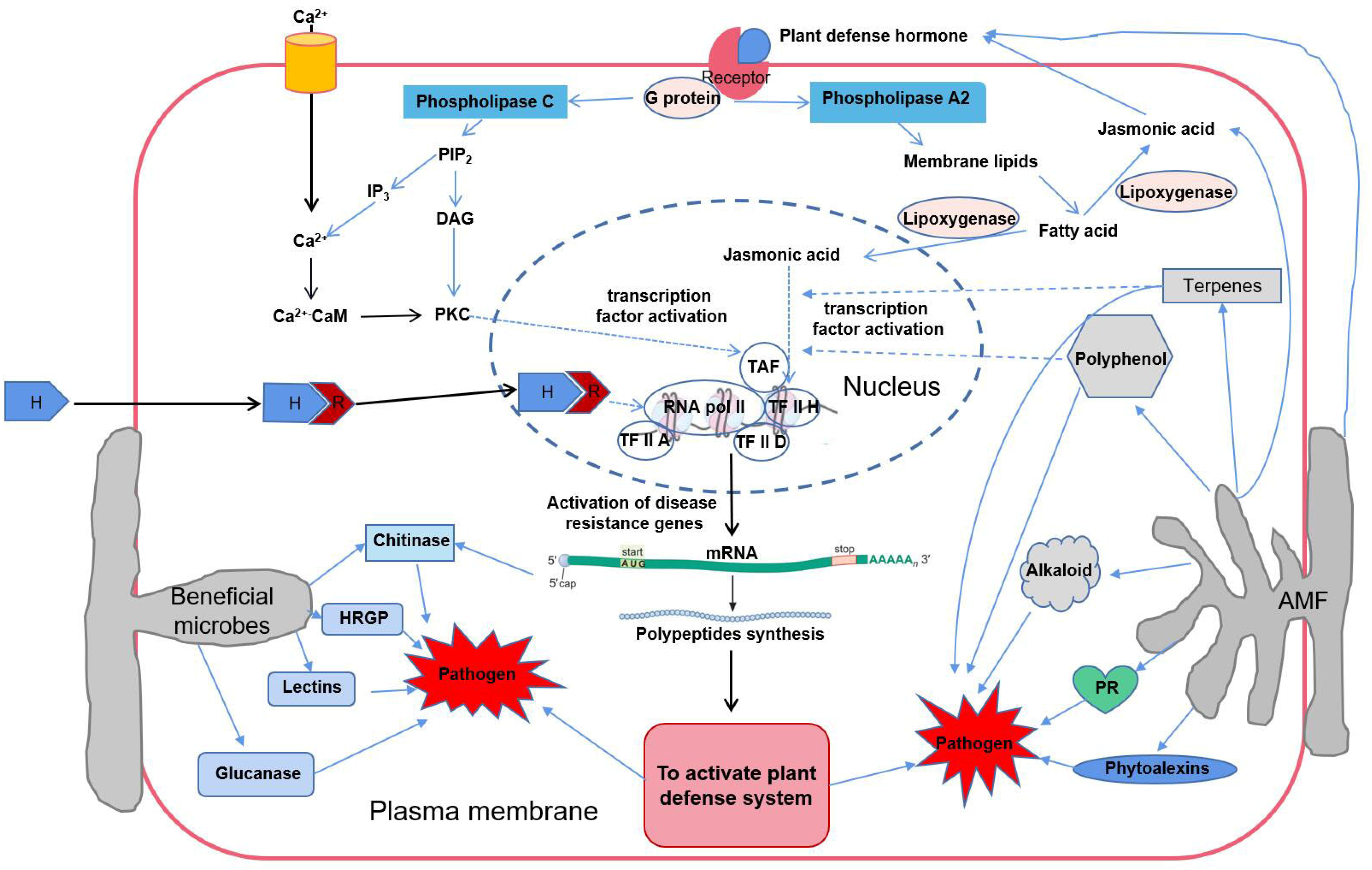
5.6. AMF Activate the Host Defense System
5.6.1. AMF Promote the Synthesis of Plant Defense Hormones
5.6.2. AMF Induce the Synthesis of Plant Signal Substances
5.6.3. AMF Regulate Defense Gene Expression
5.6.4. AMF Improve Defense Enzyme Activity and Induce PR Synthesis
6. Conclusions and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, N.X.; Enkegaard, A.; Osborne, L.S.; Ramakers, P.M.J.; Messelink, G.J.; Pijnakker, J.; Murphy, G. The banker plant method in biological control. Crit. Rev. Plant Sci. 2011, 30, 259–278. [Google Scholar] [CrossRef]
- Hage-Ahmed, K.; Rosner, K.; Steinkellner, S. Arbuscular mycorrhizal fungi and their response to pesticides. Pest. Manag. Sci. 2019, 75, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Razak, N.A.; Gange, A.C. Multitrophic Interactions between Arbuscular Mycorrhizal Fungi, Foliar Endophytic Fungi and Aphids. Microb. Ecol. 2021, 1–11. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Driesche, R.G.; Carruthers, R.I.; Center, T.; Hoddle, M.S.; Hough-Goldstein, J.; Morin, L.; Smith, L.; Wagner, D.L.; Blossey, B.; Brancatini, V.; et al. Classical biological control for the protection of natural ecosystems. Biol. Control 2010, 54 (Suppl. S1), 2–33. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.H.; Eo, J.K.; Ka, K.H.; Eom, A.H. Diversity of arbuscular mycorrhizal fungi and their roles in ecosystems. Mycobiology 2013, 41, 121–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gianinazzi, S.; Gollotte, A.; Binet, M.N.; van Tuinen, D.; Redecker, D.; Wipf, D. Agroecology: The key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 2010, 20, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, U.; Regvar, M.; Bothe, H. Arbuscular mycorrhiza and heavy metal tolerance. Phytochemistry 2007, 68, 139–146. [Google Scholar] [CrossRef]
- Obrycki, J.J.; Harwood, J.D.; Kring, T.J.; O’Neil, R.J. Aphidophagy by coccinellidae: Application of biological control in agroecosystems. Biol. Control 2009, 51, 244–254. [Google Scholar] [CrossRef]
- Allsup, C.M.; Lankau, R.A.; Paige, K.N. Herbivory and Soil Water Availability Induce Changes in Arbuscular Mycorrhizal Fungal Abundance and Composition. Microb. Ecol. 2021, 1–12. [Google Scholar] [CrossRef]
- Gabriele, B. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biot. 2009, 84, 11–18. [Google Scholar]
- Aseel, D.G.; Rashad, Y.M.; Hammad, S.M. Arbuscular Mycorrhizal Fungi Trigger Transcriptional Expression of Flavonoid and Chlorogenic Acid Biosynthetic Pathways Genes in Tomato against Tomato Mosaic Virus. Sci. Rep. 2019, 9, 9692. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Guo, Y.; Christensen, M.J.; Gao, P.; Li, Y.; Duan, T. An arbuscular mycorrhizal fungus and Epichloë festucae var. lolii reduce Bipolaris sorokiniana disease incidence and improve perennial ryegrass growth. Mycorrhiza 2018, 28, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Liu, R.J.; Su, H.R. Mechanism of increasing plant disease resistance by AM fungi. Mycosystema 2001, 20, 435–439. [Google Scholar]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef]
- Miransari, M. Interactions between arbuscular mycorrhizal fungi and soil bacteria. Appl. Microbiol. Biot. 2011, 89, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Cruz, A.F.; Ishii, T. Arbuscular mycorrhizal fungal spores host bacteria that affect nutrient biodynamics and biocontrol of soil-borne plant pathogens. Biol. Open 2012, 1, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Duan, T.; Nan, Z.; Li, Y. Arbuscular mycorrhizal fungus alleviates alfalfa leaf spots caused by Phoma medicaginis revealed by RNA-seq analysis. J. Appl. Microbiol. 2021, 130, 547–560. [Google Scholar] [CrossRef]
- Lin, P.; Zhang, M.; Wang, M.; Li, Y.; Liu, J.; Chen, Y. Inoculation with arbuscular mycorrhizal fungus modulates defense-related genes expression in banana seedlings susceptible to wilt disease. Plant Signal Behav. 2021, 16, 1884782. [Google Scholar] [CrossRef]
- Schönbeck, F.; Dehne, H.W. Damage to mycorrhizal and nonmycorrhizal cotton seedlings by Thielaviopsis basicola. Plant Dis. Rep. 1977, 61, 266–276. [Google Scholar]
- Baltruschat, H.; Schönbeck, F. The influence of endotrophic mycorrhiza on the infestation of tobacco by Thielaviopsis basicola. Phytopathol. Z 1975, 84, 172–188. [Google Scholar] [CrossRef]
- Chou, L.G.; Schmitthenner, A.F. Effect of Rhiz obium japonicum and Endogone mosseae on soybean root rot caused by Pythium ultimum and Phytophthora megasperma var. sojae. Plant Dis. Rep. 1974, 58, 221–227. [Google Scholar]
- Aguk, J.A.; Karanja, N.; Schulte-Geldermann, E.; Bruns, C.; Kinyua, Z.; Parker, M. Control of bacterial wilt (Ralstonia solanacearum) in potato (Solanum tuberosum) using rhizobacteria and arbuscular mycorrhiza fungi. Afr. J. Food Agr. Nutr. Dev. 2018, 18, 13371–13387. [Google Scholar]
- Becker, W.N. Quantification of Onion Vesicular-Arbuscular Mycorrhizae and Their Resistance to Pyrenochaeta terrestris; University of Illinois at Urbana-Champaign: Champaign, IL, USA, 1976. [Google Scholar]
- Steinkellner, S.; Hage-Ahmed, K.; García-Garrido, J.M.; Illana, A.; Ocampo, J.A.; Vierheilig, H. A comparison of wild-type, old and modern tomato cultivars in the interaction with the arbuscular mycorrhizal fungus Glomus mosseae and the tomato pathogen Fusarium oxysporum f. sp. lycopersici. Mycorrhiza 2012, 22, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Eke, P.; Adamou, S.; Fokom, R.; Dinango Nya, V.; Tsouh Fokou, P.V.; Nana Wakam, L.; Nwaga, D.; Fekam Boyom, F. Arbuscular mycorrhizal fungi alter antifungal potential of lemongrass essential oil against Fusarium solani, causing root rot in common bean (Phaseolus vulgaris L.). Heliyon 2020, 6, e05737. [Google Scholar] [CrossRef] [PubMed]
- Guzman, A.; Montes, M.; Hutchins, L.; DeLaCerda, G.; Yang, P.; Kakouridis, A.; Dahlquist-Willard, R.M.; Firestone, M.K.; Bowles, T.; Kremen, C. Crop diversity enriches arbuscular mycorrhizal fungal communities in an intensive agricultural landscape. New Phytol. 2021, 231, 447–459. [Google Scholar] [CrossRef]
- D Aljawasim, B.; M Khaeim, H.; A Manshood, M. Assessment of arbuscular mycorrhizal fungi (Glomus spp.) as potential biocontrol agents against damping-off disease Rhizoctonia solani on cucumber. J. Crop Prot. 2020, 9, 141–147. [Google Scholar]
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Gómez-Lama Cabanás, C.; Mercado-Blanco, J. Biological Control Agents against Fusarium Wilt of Banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef] [Green Version]
- Sudhasha, S. Chapter-1 Constructiveness of the Biocontrol Agents on Fusarial Wilt of Tomato Incited by the Destructive Pathogen Fusarium oxysporum f. sp. lycopersici. In Current Research and Innovations in Plant Pathology; AkiNik Publications: Delhi, India, 2020. [Google Scholar]
- Slezack, S.; Dumas-Gaudot, E.; Paynot, M.; Gianinazzi, S. Is a fully established arbuscular mycorrhizal symbiosis required for bioprotection of Pisum sativum roots against Aphanomyces euteiches? Mol. Plant Microbe Interact. 2000, 13, 238–241. [Google Scholar] [CrossRef] [Green Version]
- Krzyzaniak, Y.; Magnin-Robert, M.; Randoux, B.; Fotaine, J.; Sahraoui, A.L. Combined use of beneficial bacteria and arbuscular mycorrhizal fungi for the biocontrol of plant cryptogamic diseases: Evidence, methodology, and limits. In Symbiotic Soil Microorganisms; Springer: Cham, Switzerland, 2021; pp. 429–468. [Google Scholar]
- Weaver, D.J.; Wehunt, E.J. Effect of soil pH on susceptibility of peach to Pseudomonas syringae. Phytopathology 1975, 65, 984–989. [Google Scholar] [CrossRef]
- Kamble, V.R.; Agre, D.G. New report on AMF colonization in root parasite Striga gesnerioides and its host Lepidagathis hamiltoniana from high altitude region of Maharashtra. Int. Multidiscip. Res. J. 2014, 3, 27–31. [Google Scholar]
- Liu, R.J.; Dai, M.; Wu, X.; Li, M.; Liu, X.Z. Suppression of the root-knot nematode Meloidogyne incognita (Kofoid & White) Chitwood] on tomato by dual inoculation with arbuscular mycorrhizal fungi and plant growth-promoting rhizobacteria. Mycorrhiza 2012, 22, 289–296. [Google Scholar] [PubMed]
- De Sá, C.S.B.; Campos, M.A.S. Arbuscular mycorrhizal fungi decrease Meloidogyne enterolobii infection of Guava seedlings. J. Helminthol. 2020, 94, e183. [Google Scholar] [CrossRef]
- Rodrigues, E.; Silva, M.T.; Calandrelli, A.; Miamoto, A.; Rinaldi, L.K.; Moreno, B.P.; da Silva, C.; Dias-Arieira, C.R. Pre-inoculation with arbuscular mycorrhizal fungi affects essential oil quality and the reproduction of root lesion nematode in Cymbopogon citratus. Mycorrhiza 2021, 31, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Shrinkhala, M. Study on the Bioprotective Effect of Endomycorrhizae against M. graminicola in Rice; Laboratory of Tropical Crop Improvement, Department of Biosys, Faculty of Bioscience Engineering, Catholic University of Leuven: Leuven, Belgium, 2011. [Google Scholar]
- Vos, C.; Schouteden, N.; van Tuinen, D.; Chatagnier, O.; Elsen, A.; De Waele, D.; Panis, B.; Gianinazzi-Pearson, V. Mycorrhiza-induced resistance against the root-knot nematode Meloidogyne incognita involves priming of defense gene responses in tomato. Soil Biol. Biochem. 2013, 60, 45–54. [Google Scholar] [CrossRef]
- Vos, C.; Geerinckx, K.; Mkandawire, R.; Panis, B.; De Waele, D.; Elsen, A. Arbuscular mycorrhizal fungi affect both penetration and further life stage development of root-knot nematodes in tomato. Mycorrhiza 2012, 22, 157–163. [Google Scholar] [CrossRef]
- Tanwar, A.; Aggarwal, A.; Panwar, V. Arbuscular mycorrhizal fungi and Trichoderma viride mediated Fusarium wilt control in tomato. Biocontrol. Sci. Technol. 2013, 23, 485–498. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Roldán, A.; Pascual, J.A. Interaction between arbuscular mycorrhizal fungi and Trichoderma harzianum under conventional and low input fertilization field condition in melon crops: Growth response and Fusarium wilt biocontrol. Appl. Soil Ecol. 2011, 47, 98–105. [Google Scholar] [CrossRef]
- Sennoi, R.; Singkham, N.; Jogloy, S.; Boonlue, S.; Saksirirat, W.; Kesmala, T.; Patanothai, A. Biological control of southern stem rot caused by Sclerotium rolfsii using Trichoderma harzianum and arbuscular mycorrhizal fungi on Jerusalem artichoke (Helianthus tuberosus L.). Crop Prot. 2013, 54, 148–153. [Google Scholar] [CrossRef]
- Singh, K. Organic amendments to soil inoculated arbuscular mycorrhizal fungi and Pseudomonas fluorescens treatments reduce the development of root-rot disease and enhance the yield of Phaseolus vulgaris L. Eur. J. Soil Biol. 2011, 47, 288–295. [Google Scholar]
- Srivastava, R.; Khalid, A.; Singh, U.S.; Sharma, A.K. Evaluation of arbuscular mycorrhizal fungus, fluorescent Pseudomonas and Trichoderma harzianum formulation against Fusarium oxysporum f. sp. lycopersici for the management of tomato wilt. Biol. Control 2010, 53, 24–31. [Google Scholar] [CrossRef]
- Akköprü, A.; Demir, S. Biological control of Fusarium wilt in tomato caused by Fusarium oxysporum f. sp. lycopersici by AMF Glomus intraradices and some rhizobacteria. J. Phytopathol. 2005, 153, 544–550. [Google Scholar] [CrossRef]
- Hernández-Montiel, L.G.; Rueda-Puente, E.O.; Cordoba-Matson, M.V.; Holguín-Peña, J.R.; Zulueta-Rodríguez, R. Mutualistic interaction of rhizobacteria with arbuscular mycorrhizal fungi and its antagonistic effect on Fusarium oxysporum in Carica papaya seedlings. Crop Prot. 2013, 47, 61–66. [Google Scholar] [CrossRef]
- Behn, O. Influence of Pseudomonas fluorescens and arbuscular mycorrhiza on the growth, yield, quality and resistance of wheat infected with Gaeumannomyces graminis. J. Plant Dis. Protect. 2008, 115, 4–8. [Google Scholar] [CrossRef]
- Parvin, W.; Govender, N.; Othman, R.; Jaafar, H.; Rahman, M.; Wong, M.Y. Phenazine from Pseudomonas aeruginosa UPMP3 induced the host resistance in oil palm (Elaeis guineensis Jacq.)-Ganoderma boninense pathosystem. Sci. Rep. 2020, 10, 15621. [Google Scholar] [CrossRef]
- Hoda, H.; Ghalia, A. Effect of arbuscular mycorrhizal Fungus Glomus mosseae and Glomus subtilis on the infection of tomato roots by Fusarium oxysporum. Egyp. J. Microbiol. 2007, 42, 29–49. [Google Scholar]
- Cai, X.; Zhao, H.; Liang, C.; Li, M.; Liu, R. Effects and Mechanisms of Symbiotic Microbial Combination Agents to Control Tomato Fusarium Crown and Root Rot Disease. Front. Microbiol. 2021, 12, 629793. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Raza, W.; Wang, X.; Ran, W.; Shen, Q. Systemic modification of cotton root exudates induced by arbuscular mycorrhizal fungi and Bacillus vallismortis HJ-5 and their effects on Verticillium wilt disease. Appl. Soil Ecol. 2012, 61, 85–91. [Google Scholar] [CrossRef]
- Rashad, Y.M.; Abbas, M.A.; Soliman, H.M.; Abdel-Fattah, G.G.; Abdel-Fattah, G.M. Synergy between endophytic Bacillus amyloliquefaciens GGA and arbuscular mycorrhizal fungi induces plant defense responses against white rot of garlic and improves host plant growth. Phytopathol. Mediterr. 2020, 59, 169–186. [Google Scholar] [CrossRef]
- Tahmatsidou, V.; O’Sullivan, J.; Cassells, A.C.; Voyiatzis, D.; Paroussi, G. Comparison of AMF and PGPR inoculants for the suppression of Verticillium wilt of strawberry (Fragaria × ananassa cv. Selva). Appl. Soil Ecol. 2006, 32, 316–324. [Google Scholar] [CrossRef]
- Lowe, A.; Rafferty-McArdle, S.M.; Cassells, A.C. Effects of AMF-and PGPR-root inoculation and a foliar chitosan spray in single and combined treatments on powdery mildew disease in strawberry. Agr. Food Sci. 2012, 21, 28–38. [Google Scholar] [CrossRef]
- Chen, Q.; Wu, W.W.; Qi, S.S.; Cheng, H.; Li, Q.; Ran, Q.; Dai, Z.C.; Du, D.L.; Egan, S.; Thomas, T. Arbuscular mycorrhizal fungi improve the growth and disease resistance of the invasive plant Wedelia trilobata. J. Appl. Microbiol. 2021, 130, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Jaiti, F.; Meddich, A.; El-Hadrami, I. Effectiveness of arbuscular mycorrhizal fungi in the protection of date palm (Phoenix dactylifera L.) against bayoud disease. Physiol. Mol. Plant Pathol. 2007, 71, 166–173. [Google Scholar] [CrossRef]
- Song, Y.Y.; Cao, M.; Xie, L.J.; Liang, X.T.; Zeng, R.S.; Su, Y.J.; Huang, J.H.; Wang, R.L.; Luo, S.M. Induction of DIMBOA accumulation and systemic defense responses as mechanism of enhanced resistance of mycorrhizal corn (Zea mays L.) to sheath blight. Mycorrhiza 2011, 21, 721–731. [Google Scholar] [CrossRef]
- Lee, C.S.; Lee, Y.J.; Jeun, Y.C. Observations of infection structures on the leaves of cucumber plants pre-treated with arbuscular mycorrhiza Glomus intraradices after challenge inoculation with Colletotrichum orbiculare. Plant Pathol. 2005, 21, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Shaw, L.J.; Morris, P.; Hooker, J.E. Perception and modification of plant flavonoid signals by rhizosphere microorganisms. Environ. Microbiol. 2006, 8, 1867–1880. [Google Scholar] [CrossRef]
- Tatsumi, C.; Hyodo, F.; Taniguchi, T.; Shi, W.; Koba, K.; Fukushima, K.; Du, S.; Yamanaka, N.; Templer, P.; Tateno, R. Arbuscular Mycorrhizal Community in Roots and Nitrogen Uptake Patterns of Understory Trees Beneath Ectomycorrhizal and Non-ectomycorrhizal Overstory Trees. Front. Plant Sci. 2021, 11, 583585. [Google Scholar] [CrossRef]
- Basyal, B.; Emery, S.M. An arbuscular mycorrhizal fungus alters switchgrass growth, root architecture, and cell wall chemistry across a soil moisture gradient. Mycorrhiza 2021, 31, 251–258. [Google Scholar] [CrossRef]
- Boutaj, H.; Meddich, A.; Wahbi, S.; Moukhli, A.; El Alaoui-Talibi, Z.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Effect of Arbuscular Mycorrhizal Fungi on Verticillium wilt development of olive trees caused by Verticillium dahliae. Res. J. Biotechnol. 2019, 14, 8. [Google Scholar]
- Pozo, M.J.; Cordier, C.; Dumas-Gaudot, E.; Gianinazzi, S.; Barea, J.M.; Azcón-Aguilar, C. Localized versus systemic effect of arbuscular mycorrhizal fungi on defence responses to Phytophthora infection in tomato plants. J. Exp. Bot. 2002, 53, 525–534. [Google Scholar] [CrossRef]
- Balestrini, R.; Bonfante, P. Cell wall remodeling in mycorrhizal symbiosis: A way towards biotrophism. Front. Plant Sci. 2014, 5, 237. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Li, X.; Ludewig, U. Arbuscular mycorrhizal colonization outcompetes root hairs in maize under low phosphorus availability. Ann. Bot. 2021, 127, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Artursson, V.; Finlay, R.D.; Jansson, J.K. Interactions between arbuscular mycorrhizal fungi and bacteria and their potential for stimulating plant growth. Environ. Microbiol. 2006, 8, 1–10. [Google Scholar] [CrossRef]
- Silvestri, A.; Pérez-Tienda, J.; López-Ráez, J.A. Arbuscular Mycorrhizal Fungal Gene Expression Analysis by Real-Time PCR. Methods Mol. Biol. 2020, 2146, 157–170. [Google Scholar]
- Scervino, J.M.; Ponce, M.A.; Erra-Bassells, R.; Bompadre, J.; Vierheilig, H.; Ocampo, J.A.; Godeas, A. The effect of flavones and flavonols on colonization of tomato plants by arbuscular mycorrhizal fungi of the genera Gigaspora and Glomus. Can. J. Microbiol. 2007, 53, 702–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Rosa-Mera, C.J.; Ferrera-Cerrato, R.; Alarcón, A.; Sánchez-Colín, M.D.; Muoz-Muiz, O.D. Arbuscular mycorrhizal fungi and potassium bicarbonate enhance the foliar content of the vinblastine alkaloid in Catharanthus roseus. Plant Soil 2011, 349, 367–376. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Graham, J.H.; Syvertsen, J.P.; Drouillard, D.L. Carbon economy of sour orange in relation to mycorrhizal colonization and phosphprus status. Ann. Bot. 1993, 71, 1–10. [Google Scholar] [CrossRef]
- Kuila, D.; Ghosh, S. Aspects, problems and utilization of Arbuscular Mycorrhizal (AM) application as bio-fertilizer in sustainable agriculture. Curr. Res. Microb. Sci. 2022, 3, 100107. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, H.; Wang, D.; Guo, X.; Yang, T.; Xiang, X.; Walder, F.; Chu, H. Differential Responses of Arbuscular Mycorrhizal Fungal Communities to Long-Term Fertilization in the Wheat Rhizosphere and Root Endosphere. Appl. Environ. Microbiol. 2021, 87, e0034921. [Google Scholar] [CrossRef]
- Wang, Y.; Bao, X.; Li, S. Effects of Arbuscular Mycorrhizal Fungi on Rice Growth Under Different Flooding and Shading Regimes. Front. Microbiol. 2021, 12, 756752. [Google Scholar] [CrossRef]
- Liang, M.; Shi, L.; Burslem, D.F.R.P.; Johnson, D.; Fang, M.; Zhang, X.; Yu, S. Soil fungal networks moderate density-dependent survival and growth of seedlings. New Phytol. 2021, 230, 2061–2071. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Hermosa, R.; Monte, E.; Nicolás, C. Trichoderma harzianum favours the access of arbuscular mycorrhizal fungi to non-host Brassicaceae roots and increases plant productivity. Sci. Rep. 2019, 9, 11650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, A.J.; Daniell, T.J.; Cameron, D.D.; Field, K.J. A commercial arbuscular mycorrhizal inoculum increases root colonization across wheat cultivars but does not increase assimilation of mycorrhiza-acquired nutrients. Plants People Planet 2021, 3, 588–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.H.; Yanagi, A.; Miyawaki, Y.; Okada, T.; Matsubara, Y. Disease tolerance and changes in antioxidative abilities in mycorrhizal strawberry plants. J. Jpn. Soc. Hortic. Sci. 2010, 79, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Tahat, M.M.; Sijam, K.; Othman, R. Ultrastructural changes of tomatoes (Lycopersicon esculentum) root colonized by Glomus mosseae and Ralstonia solanacearum. Afr. J. Biotechnol. 2012, 25, 668–6686. [Google Scholar]
- Jaiti, F.; Kassami, M.; Meddich, A.; Hadrami, I.E. Effect of arbuscular mycorrhization on the accumulation of hydroxycinnamic acid derivatives in data palm seedlings challenged with Fusarium oxysporum f. sp. Albedinis. J. Phytopathol. 2008, 156, 641–646. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Medina, A.; Pascual, J.A.; Pérez-Alfocea, F.; Albacete, A.; Roldán, A. Trichoderma harzianum and Glomus intraradices modify the hormone disruption induced by Fusarium oxysporum infection in melon plants. Phytopathology 2010, 100, 682–688. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.L.; Smith, A.F.; Smith, S.E. The rmc locus does not affect plant interactions or defense- related gene expression when tomato (Solanum lycopersicum) is infected with the root fungal parasite, Rhizoctonia. Funct. Plant Biol. 2006, 33, 289–296. [Google Scholar] [CrossRef]
- Lioussanne, L.; Jolicoeur, M.; St-Arnaud, M. Mycorrhizal colonization with Glomus intraradices and development stage of transformed tomato roots significantly modify the chemotactic response of zoospores of the pathogen Phytophthora nicotianae. Soil Biol. Biochem. 2008, 40, 2217–2224. [Google Scholar] [CrossRef]
- Chandanie, W.A.; Kubota, M.; Hyakumachi, M. Interactions between the arbuscular mycorrhizal fungus Glomus mosseae and plant growth-promoting fungi and their significance for enhancing plant growth and suppressing damping-off of cucumber (Cucumis sativus L.). Appl. Soil Ecol. 2009, 41, 336–341. [Google Scholar] [CrossRef]
- Zhu, H.H.; Yao, Q. Localized and systemic increase of phenols in tomato roots induced by Glomus versiforme inhibits Ralstonia solanacearum. J. Phytopathol. 2004, 152, 537–542. [Google Scholar] [CrossRef]
- Miransari, M.; Abrishamchi, A.; Khoshbakht, K.; Niknam, V. Plant hormones as signals in arbuscular mycorrhizal symbiosis. Crit. Rev. Biotechnol. 2014, 34, 123–133. [Google Scholar] [CrossRef]
- Cordier, C.; Gianinazzi, S.; Gianinazzi-Pearson, V. Colonisation patterns of root tissues by Phytophthora nicotianae var. parasitica related to reduced disease in mycorrhizal tomato. Plant Soil 1996, 185, 223–232. [Google Scholar] [CrossRef]
- Whipps, J.M. Prospects and limitations for mycorrhizas in biocontrol of root pathogens. Can. J. Bot. 2004, 82, 1198–1227. [Google Scholar] [CrossRef]
- Tabin, T.; Arunachalam, A.; Shrivastava, K.; Arunachalam, K. Effect of arbuscular mycorrhizal fungi on damping-off disease in Aquilaria agallocha Roxb. Seedlings. Trop. Ecol. 2009, 50, 243–248. [Google Scholar]
- Zhou, J.; Chai, X.; Zhang, L.; George, T.S.; Wang, F.; Feng, G. Different Arbuscular Mycorrhizal Fungi Cocolonizing on a Single Plant Root System Recruit Distinct Microbiomes. mSystems 2020, 5, e00929-20. [Google Scholar] [CrossRef]
- Castillo, P.; Nico, A.I.; Azcón-Aguilar, C.; Del Río Rincón, C.; Calvet, C.; Jiménez-Díaz, R.M. Protection of olive planting stocks against parasitism of root-knot nematodes by arbuscular mycorrhizal fungi. Plant Pathol. 2006, 55, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, A.M.; Harrison, M.J. Signaling events during initiation of arbuscular mycorrhizal symbiosis. J. Integr. Plant Biol. 2014, 56, 250–261. [Google Scholar] [CrossRef]
- Metwally, R.A.; Al-Amri, S.M. Individual and interactive role of Trichoderma viride and arbuscular mycorrhizal fungi on growth and pigment content of onion plants. Lett. Appl. Microbiol. 2020, 70, 79–86. [Google Scholar] [CrossRef]
- Ortu, G.; Balestrini, R.; Pereira, P.A.; Becker, J.D.; Küster, H.; Bonfante, P. Plant genes related to gibberellin biosynthesis and signaling are differentially regulated during the early stages of AM fungal interactions. Mol. Plant 2012, 5, 951–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hause, B.; Mrosk, C.; Isayenkov, S.; Strack, D. Jasmonates in arbuscular mycorrhizal interactions. Phytochemistry 2007, 68, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Calcagno, C.; Novero, M.; Genre, A.; Bonfante, P.; Lanfranco, L. The exudate from an arbuscular mycorrhizal fungus induces nitric oxide accumulation in Medicago truncatula roots. Mycorrhiza 2012, 22, 259–269. [Google Scholar] [CrossRef] [PubMed]
- He, Z.Q.; Li, H.X.; Tang, H.R. Effect of arbuscular mycorrhizal fungi on endogenous in Cucumber after Rhizoctonia solani inoculation. Chin. Agric. Sci. Bull. 2010, 26, 187–190. [Google Scholar]
- Dugassa, G.D.; Vonalten HSchonbeck, F. Effects of arbuscular mycorrhizal (AM) on health of linumusita tissimuml infected by fungal pathogens. Plant Soil 1996, 185, 173–182. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopath. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Kapoor, R. Induced resistance in mycorrhizal tomato is correlated to concentration of jasmonic acid. Online J. Biol. Sci. 2008, 8, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Jalali, B.L.; Bhargava, S.; Kamble, A. Signal transduction and transcriptional regulation of plant defense responses. J. Phytopathol. 2006, 154, 65–74. [Google Scholar] [CrossRef]
- Riedel, T.; Groten, K.; Baldwin, I.T. Symbiosis between Nicotiana attenuata and Glomus intraradices: Ethylene plays a role, jasmonic acid does not. Plant Cell Environ. 2008, 31, 1203–1213. [Google Scholar] [CrossRef]
- Li, H.Y.; Liu, R.J.; Shu, H.; Li, Y. Chib1 and PAL5 directly involved in the defense responses induced by the arbuscular mycorrhizal fungus Glomus fasciculatus against nematode. Mycosystema 2005, 24, 385–393. [Google Scholar]
- Liu, J.Y.; Maldonado-Mendoza, I.; Lopez-Meyer, M.; Cheung, F.C.; Town CDHarrison, M.J. Arbuscular mycorrhizal symbiosis is accompanied by local and systemic alterations in gene expression and an increase in disease resistance in the shoots. Plant J. 2007, 50, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Tahiri-Alaoui, A.; Frigotto, L.; Manville, N.; Ibrahim, J.; Romby, P.; James, W. High affinity nucleic acid aptamers for streptavidin incorporated into bi-specific capture ligands. Nucleic Acids Res. 2002, 30, 30–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallou, A.; Mosquera, H.P.L.; Cranenbrouck, S.; Suárezb, J.P.; Declerck, S. Mycorrhiza induced resistance in potato plantlets challenged by Phytophthora infestans. Physiol. Mol. Plant Pathol. 2011, 76, 20–26. [Google Scholar] [CrossRef]
- Isayenkov, S.; Mrosk, C.; Stenzel, I.; Strack, D.; Hause, B. Suppression of allene oxide cyclase in hairy roots of Medicago truncatula reduces jasmonate levels and the degree of mycorrhization with Glomus intraradices. Plant Physiol. 2005, 139, 1401–1410. [Google Scholar] [CrossRef] [Green Version]
- Fester, T.; Hause, G. Accumulation of reactive oxygen species in arbuscular mycorrhizal roots. Mycorrhiza 2005, 15, 373–379. [Google Scholar] [CrossRef]
- De Román, M.; Fernández, I.; Wyatt, T.; Sahrawy, M.; Heil, M.; Poze, M.J. Elicitation of foliar resistance mechanisms transiently impairs root association with arbuscular mycorrhizal fungi. J. Ecol. 2011, 99, 36–45. [Google Scholar] [CrossRef]
- Kloppholz, S.; Kuhn, H.; Requena, N. A secreted fungal effector of Glomus intraradices promotes symbiotic biotrophy. Curr. Biol. 2011, 21, 1204–1209. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, W.; Yan, J.; Zhou, M.; Yao, X.; Gao, A.; Ma, C.; Cheng, J.; Ruan, J. Roles of Arbuscular mycorrhizal Fungi as a Biocontrol Agent in the Control of Plant Diseases. Microorganisms 2022, 10, 1266. https://doi.org/10.3390/microorganisms10071266
Weng W, Yan J, Zhou M, Yao X, Gao A, Ma C, Cheng J, Ruan J. Roles of Arbuscular mycorrhizal Fungi as a Biocontrol Agent in the Control of Plant Diseases. Microorganisms. 2022; 10(7):1266. https://doi.org/10.3390/microorganisms10071266
Chicago/Turabian StyleWeng, Wenfeng, Jun Yan, Meiliang Zhou, Xin Yao, Aning Gao, Chao Ma, Jianping Cheng, and Jingjun Ruan. 2022. "Roles of Arbuscular mycorrhizal Fungi as a Biocontrol Agent in the Control of Plant Diseases" Microorganisms 10, no. 7: 1266. https://doi.org/10.3390/microorganisms10071266