
New Cell Lines Derived from European Tick Species

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ticks
2.2. Preparation and Monitoring of Primary Tick Cell Cultures
2.3. Generation and Cryopreservation of Tick Cell Lines
2.4. Confirmation of Species Origin and Screening for Contaminating Microorganisms
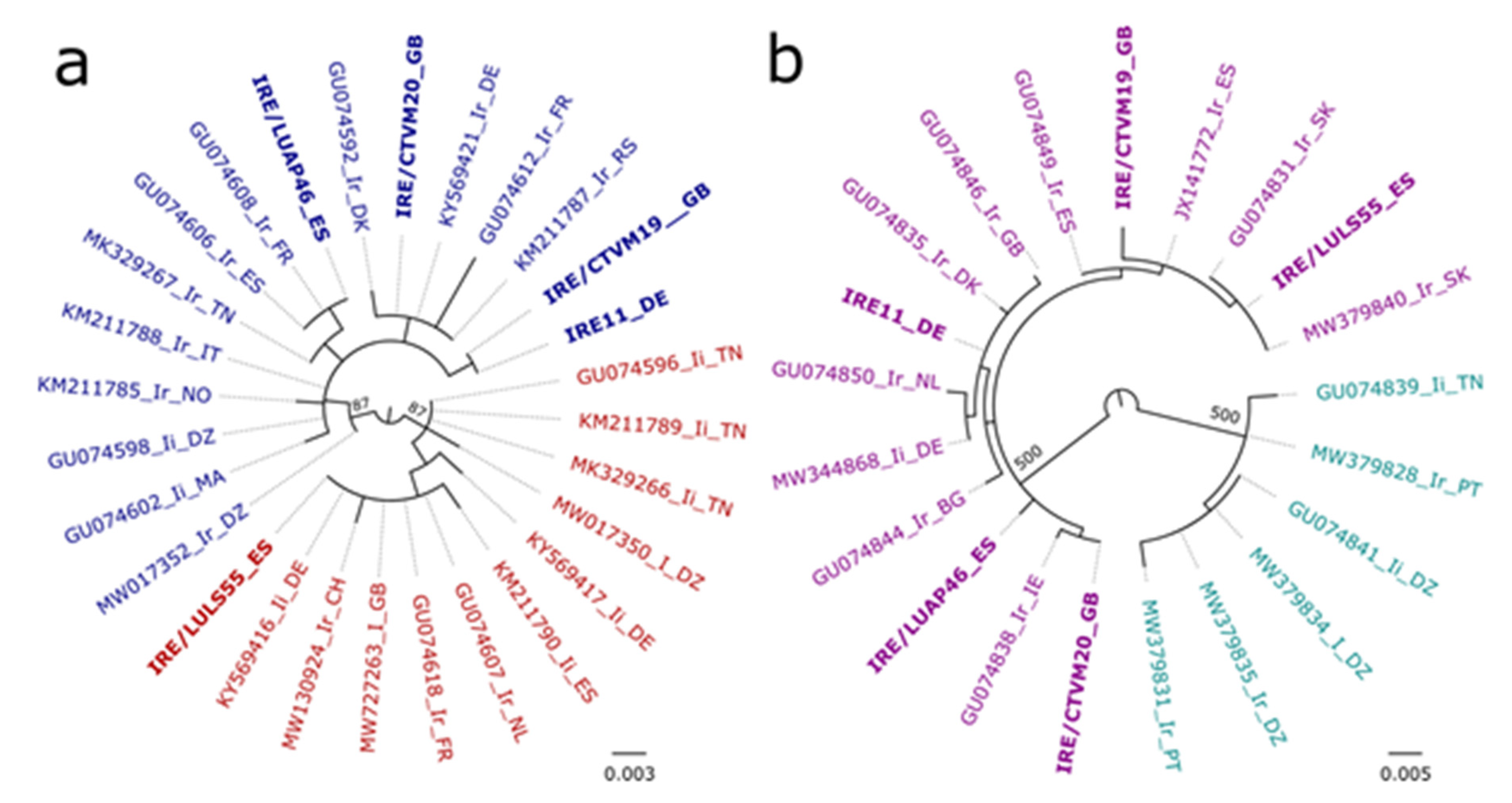
2.5. Sequence and Phylogenetic Analyses
3. Results
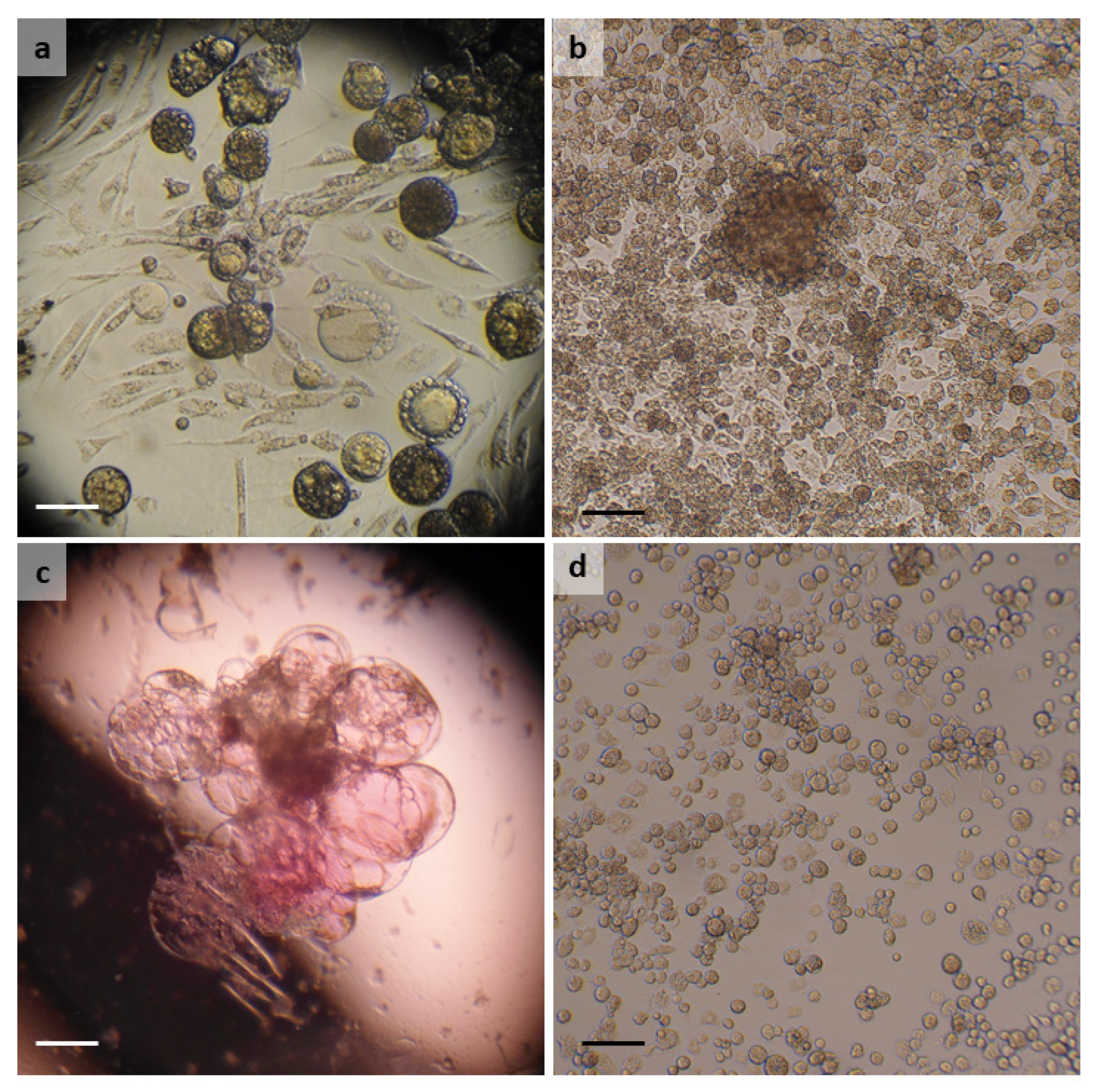
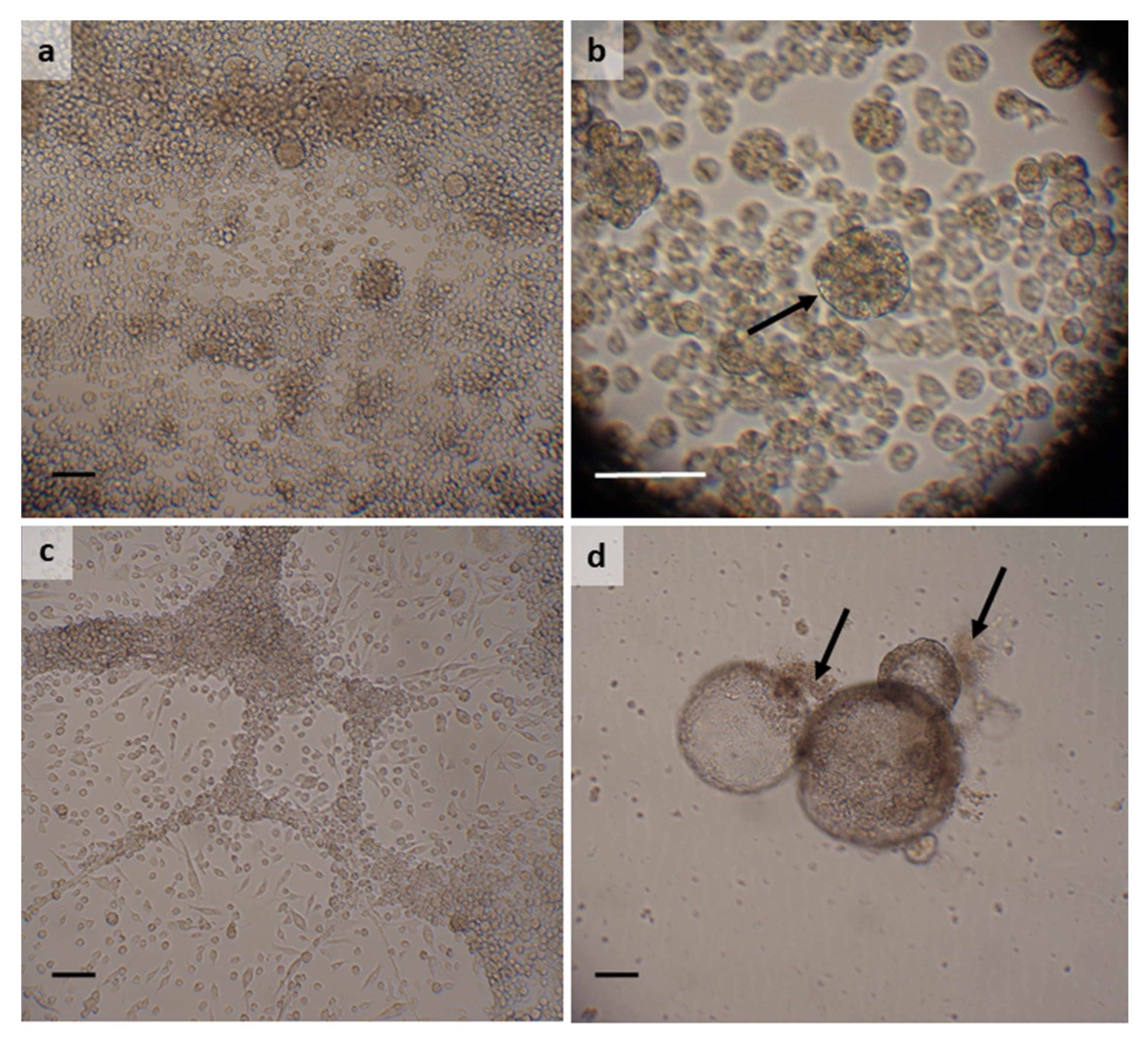
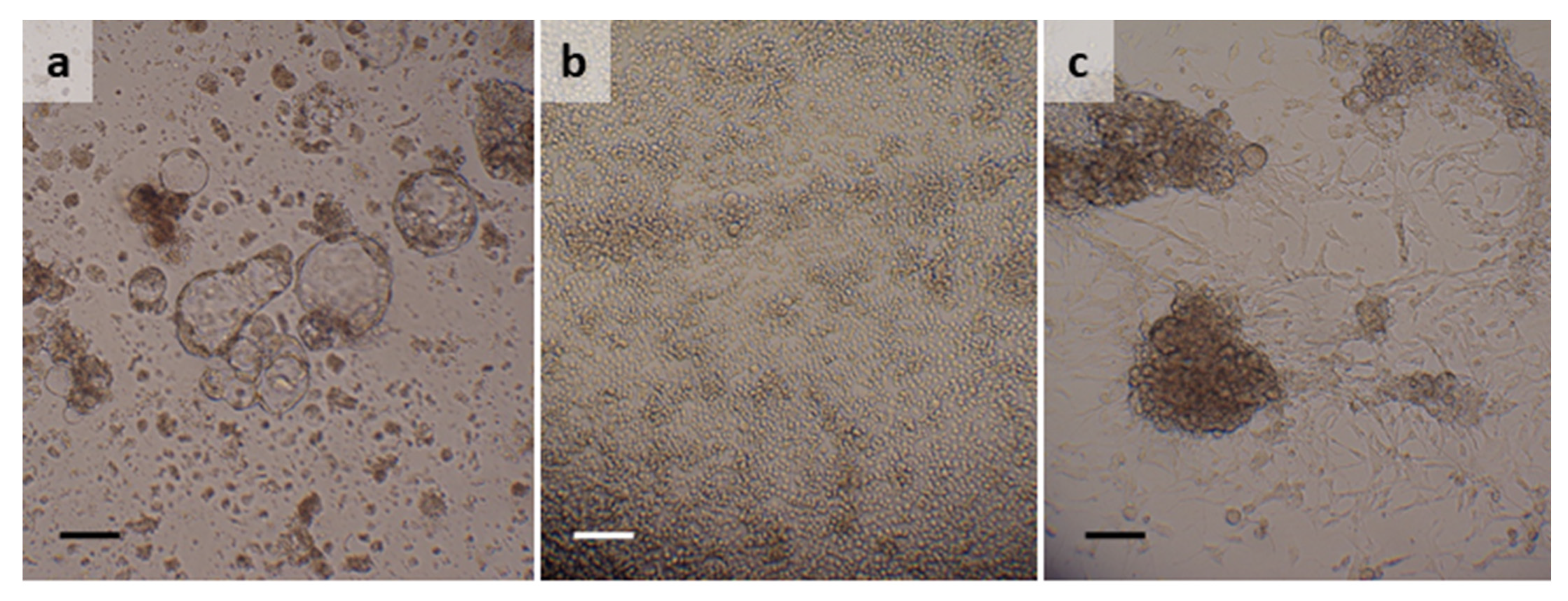
3.1. Argas reflexus
3.2. Dermacentor reticulatus
3.3. Hyalomma lusitanicum
3.4. Hyalomma scupense
3.5. Ixodes ricinus
3.6. Rhipicephalus bursa
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, 643–645. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; Kahl, O.; Zintl, A. What do we still need to know about Ixodes ricinus? Ticks Tick Borne Dis. 2021, 12, 101682. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control and European Food Safety Authority. Tick maps [internet]. Stockholm: ECDC. 2022. Available online: https://www.ecdc.europa.eu/en/publications-data/ixodes-ricinus-current-known-distribution-march-2022 (accessed on 25 April 2022).
- Estrada-Pena, A.; Pfäffle, M.; Baneth, G.; Kleinerman, G.; Petney, T.N. Ixodoidea of the Western Palaearctic: A review of available literature for identification of species. Ticks Tick Borne Dis. 2017, 8, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Darby, A.C.; Baylis, M.; Makepeace, B.L. The Tick Cell Biobank: A global resource for in vitro research on ticks, other arthropods and the pathogens they transmit. Ticks Tick Borne Dis. 2018, 9, 1364–1371. [Google Scholar] [CrossRef]
- Al-Rofaai, A.; Bell-Sakyi, L. Tick cell lines in research on tick control. Front. Physiol. 2020, 11, 152. [Google Scholar] [CrossRef] [Green Version]
- Loginov, D.S.; Böttinger, K.; Loginova, Y.F.; Dycka, F.; Vechtova, P.; Sterba, J. Biotyping of IRE/CTVM19 tick cell line infected by tick-borne encephalitis virus. Ticks Tick Borne Dis. 2020, 11, 101420. [Google Scholar] [CrossRef]
- Nelson, C.M.; Herron, M.J.; Wang, X.-R.; Baldridge, G.D.; Oliver, J.D.; Munderloh, U.G. Global transcription profiles of Anaplasma phagocytophilum at key stages of infection in tick and human cell lines and granulocytes. Front. Vet. Sci. 2020, 7, 111. [Google Scholar] [CrossRef]
- Mateos-Hernandez, L.; Pipova, N.; Allain, E.; Henry, C.; Rouxel, C.; Lagree, A.-C.; Haddad, N.; Boulois, H.-J.; Valdes, J.J.; Alberdi, P.; et al. Enlisting the Ixodes scapularis embryonic ISE6 cell line to investigate the neuronal basis of tick—pathogen interactions. Pathogens 2021, 10, 70. [Google Scholar] [CrossRef]
- Salata, C.; Moutailler, S.; Attoui, H.; Zweygarth, E.; Decker, L.; Bell-Sakyi, L. How relevant are in vitro culture models for study of tick-pathogen interactions? Pathog. Glob. Health 2021, 115, 437–455. [Google Scholar] [CrossRef]
- Simser, J.; Palmer, A.; Fingerle, V.; Wilske, B.; Kurtti, T.; Munderloh, U. Rickettsia monacensis sp. nov., a spotted fever group Rickettsia, from ticks (Ixodes ricinus) collected in a European city park. Appl. Environ. Microbiol. 2002, 68, 4559–4566. [Google Scholar] [CrossRef] [Green Version]
- Bell-Sakyi, L.; Zweygarth, E.; Blouin, E.F.; Gould, E.A.; Jongejan, F. Tick cell lines: Tools for tick and tick-borne disease re-search. Trends Parasitol. 2007, 23, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Koh-Tan, H.H.C.; Strachan, E.; Cooper, K.; Bell-Sakyi, L.; Jonsson, N.N. Identification of a novel β-adrenergic octopamine receptor-like gene (βAOR-like) and increased ATP-binding cassette B10 (ABCB10) expression in a Rhipicephalus microplus cell line derived from acaricide-resistant ticks. Parasites Vectors 2016, 9, 425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kröber, T.; Guerin, P.M. In vitro feeding assays for hard ticks. Trends Parasitol. 2007, 23, 445–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munderloh, U.G.; Kurtti, T.J. Formulation of medium for tick cell culture. Exp. Appl. Acarol. 1989, 7, 219–229. [Google Scholar] [CrossRef]
- Black, W.C.; Piesman, J. Phylogeny of hard and soft tick taxa (Acari:Ixodida) based on mitochondrial 16S rDNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Noureddine, R.; Chauvin, A.; Plantard, O. Lack of genetic structure among Eurasian populations of the tick Ixodes ricinus contrasts with marked divergence from north-African populations. Int. J. Parasitol. 2011, 41, 183–192. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Meth. 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomar, A.M.; Veiga, J.; Portillo, A.; Santibanez, S.; Vaclav, R.; Santibanez, P.; Oteo, J.A.; Valera, F. Novel genotypes of nidicolous Argas ticks and their associated microorganisms from Spain. Front. Vet. Sci. 2021, 8, 637837. [Google Scholar] [CrossRef] [PubMed]
- Dwużnik, D.; Mierzejewska, E.J.; Kowalec, M.; Alsarraf, M.; Stańczak, Ł.; Opalińska, P.; Krokowska-Paluszak, M.; Górecki, G.; Bajer, A. Ectoparasites of red foxes (Vulpes vulpes) with a particular focus on ticks in subcutaneous tissues. Parasitology 2020, 147, 1359–1368. [Google Scholar] [CrossRef]
- Remesar, S.; Fernandez, P.D.; Venzal, J.M.; Perez-Creo, A.; Prieto, A.; Estrada-Pena, A.; Lopez, C.M.; Panadero, R.; Fernandez, G.; Diez-Banos, P.; et al. Tick species diversity and population dynamics of Ixodes ricinus in Galicia (north-western Spain). Ticks Tick Borne Dis. 2019, 10, 132–137. [Google Scholar] [CrossRef]
- Karger, A.; Kampen, H.; Bettin, B.; Dautel, H.; Ziller, M.; Hoffmann, B.; Süss, J.; Klaus, C. Species determination and characterization of developmental stages of ticks by whole-animal matrix-assisted laser desorption/ionization mass spectrometry. Ticks Tick Borne Dis. 2012, 3, 78–89. [Google Scholar] [CrossRef]
- Sands, A.F.; Apanaskevich, D.A.; Matthee, S.; Horak, I.G.; Harrison, A.; Karim, S.; Mohammad, M.K.; Mumcuoglu, K.Y.; Rajakaruna, R.S.; Santos-Silva, M.M.; et al. Effects of tectonics and large scale climatic changes on the evolutionary history of Hyalomma ticks. Mol. Phylogenet. Evol. 2017, 114, 153–165. [Google Scholar] [CrossRef]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef]
- Hornok, S.; Grima, A.; Takacs, N.; Szekeres, S.; Kontschan, J. First records and molecular-phylogenetic analyses of three tick species (Ixodes kaiseri, Hyalomma lusitanicum and Ornithodoros coniceps) from Malta. Ticks Tick Borne Dis. 2020, 11, 101379. [Google Scholar] [CrossRef]
- Lv, J.; Wu, S.; Zhang, Y.; Chen, Y.; Feng, C.; Yuan, X.; Jia, G.; Deng, J.; Wang, C.; Wang, Q.; et al. Assessment of four DNA fragments (COI, 16S rDNA, ITS2, 12S rDNA) for species identification of the Ixodida (Acari: Ixodida). Parasites Vectors 2014, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Apanaskevich, D.A.; Filippova, N.A.; Horak, I.G. The genus Hyalomma Koch, 1844. X. Redescription of all parasitic stages of H. (Euhyalomma) scupense Schulze, 1919 (=H. detritum Schulze) (Acari: Ixodidae) and notes on its biology. Folia Parasitol. 2010, 57, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghafar, A.; Gasser, R.B.; Rashid, I.; Ghafoor, A.; Jabbar, A. Exploring the prevalence and diversity of bovine ticks in five agro-ecological zones of Pakistan using phenetic and genetic tools. Ticks Tick Borne Dis. 2020, 11, 101472. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.S.; de Bruin, A.; Veloso, A.R.; Marques, C.; da Fonseca, I.P.; de Sousa, R.; Sprong, H.; Santos-Silva, M.M. Detection of Anaplasma phagocytophilum, Candidatus Neoehrlichia sp., Coxiella burnetii and Rickettsia spp. in questing ticks from a recreational park, Portugal. Ticks Tick Borne Dis. 2018, 9, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Luu, L.; Palomar, A.M.; Farrington, G.; Schilling, A.-K.; Premchand-Branker, S.; McGarry, J.; Makepeace, B.L.; Meredith, A.; Bell-Sakyi, L. Bacterial pathogens and symbionts harboured by Ixodes ricinus ticks parasitising red squirrels in the United Kingdom. Pathogens 2021, 10, 458. [Google Scholar] [CrossRef]
- Estrada-Pena, A.; Nava, S.; Petney, T. Description of all the stages of Ixodes inopinatus n. sp. (Acari: Ixodidae). Ticks Tick Borne Dis. 2014, 5, 734–743. [Google Scholar] [CrossRef]
- Jaenson, T.G.T.; Varv, K.; Frojdman, I.; Jaaskelainen, A.; Rundgren, K.; Versteirt, V.; Estrada-Pena, A.; Medlock, J.M.; Golovljova, I. First evidence of established populations of the taiga tick Ixodes persulcatus (Acari: Ixodidae) in Sweden. Parasites Vectors 2016, 9, 377. [Google Scholar] [CrossRef]
- Carpi, G.; Kitchen, A.; Kim, H.L.; Ratan, A.; Drautz-Moses, D.I.; McGraw, J.J.; Kazimirova, M.; Rizzoli, A.; Schuster, S.C. Mitogenomes reveal diversity of the European Lyme borreliosis vector Ixodes ricinus in Italy. Mol. Phylog. Evol. 2016, 101, 194–202. [Google Scholar] [CrossRef]
- Kotsarenko, K.; Vechtova, P.; Lieskovska, J.; Füssy, Z.; Cabral-de-Mello, D.; Rego, R.; Alberdi, P.; Collins, M.; Bell-Sakyi, L.; Sterba, J.; et al. Karyotype changes in long-term cultured tick cell lines. Sci. Rep. 2020, 10, 13443. [Google Scholar] [CrossRef]
- Hornok, S.; Daccord, J.; Takács, N.; Kontschán, J.; Tuska-Szalay, B.; Sándor, A.D.; Szekeres, S.; Meli, M.L.; Hofmann-Lehmann, R. Investigation on haplotypes of ixodid ticks and retrospective finding of Borrelia miyamotoi in bank vole (Myodes glareolus) in Switzerland. Ticks Tick Borne Dis. 2022, 13, 101865. [Google Scholar] [CrossRef]
- Norte, A.C.; Boyer, P.H.; Castillo-Ramirez, S.; Chvostáč, M.; Brahami, M.O.; Rollins, R.E.; Woudenberg, T.; Didyk, Y.M.; Derdakova, M.; Núncio, M.S.; et al. The population structure of Borrelia lusitaniae is reflected by a population division of its Ixodes vector. Microorganisms 2021, 9, 933. [Google Scholar] [CrossRef]
- Orkun, O.; Vatansever, Z. Rediscovery and first genetic description of some poorly known tick species: Haemaphysalis kopetdaghica Kerbabaev, 1962 and Dermacentor raskemensis Pomerantzev, 1946. Ticks Tick Borne Dis. 2021, 12, 101726. [Google Scholar] [CrossRef] [PubMed]
- Socolovschi, C.; Mediannikov, O.; Raoult, D.; Parola, P. Update on tick-borne bacterial diseases in Europe. Parasite 2009, 16, 259–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyman, P.; Cochez, C.; Hofhuis, A.; van der Giessen, J.; Sprong, H.; Porter, S.R.; Losson, B.; Saegerman, C.; Donoso-Mantke, O.; Niedrig, M.; et al. A clear and present danger: Tick-borne diseases in Europe. Expert Rev. Anti Infect. Ther. 2010, 8, 33–50. [Google Scholar] [CrossRef]
- Moutailler, S.; Moro, C.V.; Vaumourin, E.; Michelet, L.; Tran, F.H.; Devillers, E.; Cosson, J.-F.; Gasqui, P.; Van, V.T.; Mavangui, P.; et al. Co-infection of ticks: The rule rather than the exception. PloS Negl. Trop. Dis. 2016, 10, e0004539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Khafaji, A.M.; Clegg, S.R.; Pinder, A.C.; Luu, L.; Hansford, K.M.; Seeliog, F.; Dinnis, R.E.; Margos, G.; Medlock, J.M.; Feil, E.J.; et al. Multi-locus sequence typing of Ixodes ricinus and its symbiont Candidatus Midichloria mitochondrii across Europe reveals evidence of local co-cladogenesis in Scotland. Ticks Tick Borne Dis. 2019, 10, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Sprong, H.; Fonville, M.; van Leeuwen, A.D.; Devillers, E.; Ibanez-Justicia, A.; Stroo, A.; Hansford, K.; Cull, B.; Medlock, J.; Heyman, P.; et al. Detection of pathogens in Dermacentor reticulatus in northwestern Europe: Evaluation of a high-throughput array. Heliyon 2019, 5, e01270. [Google Scholar] [CrossRef] [Green Version]
- Grochowska, A.; Milewski, R.; Pancewicz, S.; Dunaj, J.; Czupryna, P.; Milewska, A.J.; Rog-Makal, M.; Grygorczuk, S.; Moniuszko-Malinowska, A. Comparison of tick-borne pathogen prevalence in Ixodes ricinus ticks collected in urban areas of Europe. Sci. Rep. 2020, 10, 6975. [Google Scholar] [CrossRef]
- Alberdi, M.P.; Nijhof, A.M.; Jongejan, F.; Bell-Sakyi, L. Tick cell culture isolation and growth of Rickettsia raoultii from Dutch Dermacentor reticulatus ticks. Ticks Tick Borne Dis. 2012, 3, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Palomar, A.M.; Premchand-Branker, S.; Alberdi, P.; Belova, O.; Moniuszko-Malinowska, A.; Kahl, O.; Bell-Sakyi, L. Isolation of known and potentially pathogenic tick-borne microorganisms from European ixodid ticks using tick cell lines. Ticks Tick Borne Dis. 2019, 10, 628–638. [Google Scholar] [CrossRef]
- Alberdi, M.P.; Dalby, M.J.; Rodriguez-Andres, J.; Fazakerley, J.K.; Kohl, A.; Bell-Sakyi, L. Detection and identification of putative bacterial endosymbionts and endogenous viruses in tick cell lines. Ticks Tick Borne Dis. 2012, 3, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Darby, A.C.; Al-Khafaji, A.M.; Whitehead, M.; Hartley, C.S.; Robinson, G.; Armstrong, S.D.; Beliavskaia, A.Y.; Bah, G.S.; Githaka, N.; Bell-Sakyi, L.; et al. Amblyomma variegatum, vector of African tick-bite fever, contains an integrated Rickettsia africae chromosome in its nuclear genome. Am. J. Trop. Med. Hyg. 2019, 101, 211. [Google Scholar]
- Hubalek, Z.; Rudolf, I. Tick-borne viruses in Europe. Parasitol. Res. 2012, 111, 9–36. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, C.L.; Mansfield, K.L.; Phipps, L.P.; Wakeley, P.R.; Mearns, R.; Schock, A.; Bell, S.; Breed, A.C.; Fooks, A.R.; Johnson, N. Louping ill virus: An endemic tick-borne disease of Great Britain. J. Gen. Virol. 2014, 95, 1005–1014. [Google Scholar] [CrossRef] [Green Version]
- Ruzek, D.; Bell-Sakyi, L.; Kopecky, J.; Grubhoffer, L. Growth of tick-borne encephalitis virus (European subtype) in cell lines from vector and non-vector ticks. Virus Res. 2008, 137, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Weisheit, S.; Villar, M.; Tykalová, H.; Popara, M.; Loecherbach, J.; Watson, M.; Růžek, D.; Grubhoffer, L.; de la Fuente, J.; Fazakerley, J.K.; et al. Ixodes scapularis and Ixodes ricinus tick cell lines respond to infection with tick-borne encephalitis virus: Transcriptomic and proteomic analysis. Parasites Vectors 2015, 8, 599. [Google Scholar] [CrossRef] [Green Version]
- Belova, O.A.; Litov, A.G.; Kholodilov, I.S.; Kozlovskaya, L.I.; Bell-Sakyi, L.; Romanova, L.I.; Karganova, G.G. Properties of tick-borne encephalitis virus populations during persistent infection of ixodid ticks and tick cell lines. Ticks Tick Borne Dis. 2017, 8, 895–906. [Google Scholar] [CrossRef]
- Helmova, R.; Honig, V.; Tykalova, H.; Palus, M.; Bell-Sakyi, L.; Grubhoffer, L. Tick-borne encephalitis virus adaptation in different host environments and existence of quasispecies. Viruses 2020, 12, 902. [Google Scholar] [CrossRef]
- Lattová, E.; Straková, P.; Pokorná-Formanová, P.; Grubhoffer, L.; Bell-Sakyi, L.; Zdráhal, Z.; Palus, M.; Ružek, D. Comprehensive N-glycosylation mapping of envelope glycoprotein from tick-borne encephalitis virus grown in human and tick cells. Sci. Rep. 2020, 10, 13204. [Google Scholar] [CrossRef]
- Lemasson, M.; Caignard, G.; Unterfinger, Y.; Attoui, H.; Bell-Sakyi, L.; Hirchaud, E.; Moutailler, S.; Johnson, N.; Vitour, D.; Richardson, J.; et al. Exploration of binary protein-protein interactions between tick-borne flaviviruses and Ixodes ricinus. Parasites Vectors 2021, 14, 144. [Google Scholar] [CrossRef]
- Lickova, M.; Havlikova, S.F.; Slavikova, M.; Slovak, M.; Drexler, J.F.; Klempa, B. Dermacentor reticulatus is a vector of tick-borne encephalitis virus. Ticks Tick Borne Dis. 2020, 11, 101414. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Kohl, A.; Bente, D.A.; Fazakerley, J.F. Tick cell lines for study of Crimean-Congo hemorrhagic fever virus and other arboviruses. Vector-Borne Zoonot. Dis. 2012, 12, 769–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salata, C.; Monteil, V.; Karlberg, H.; Celestino, M.; Devignot, S.; Leijon, M.; Bell-Sakyi, L.; Bergeron, E.; Weber, F.; Mirazimi, A. The DEVD motif of Crimean-Congo hemorrhagic fever virus nucleoprotein is essential for viral replication in tick cells. Emerg. Microbes Infect. 2018, 7, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvati, M.V.; Salaris, C.; Monteil, V.; Del Vecchio, C.; Palu, G.; Parolin, C.; Calistri, A.; Bell-Sakyi, L.; Mirazimi, A.; Salata, C. Virus-derived DNA forms mediate the persistent infection of tick cells by Hazara virus and Crimean-Congo hemorrhagic fever virus. J. Virol. 2021, 95, e0163821. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Attoui, H. Virus discovery using tick cell lines. Evol. Bioinform. 2016, 12, 31–34. [Google Scholar]
- Kholodilov, I.S.; Litov, A.G.; Klimentov, A.S.; Belova, O.A.; Polienko, A.E.; Nikitin, N.A.; Shchetinin, A.M.; Ivannikova, A.Y.; Bell-Sakyi, L.; Yakovlev, A.S.; et al. Isolation and characterization of Alongshan virus in Russia. Viruses 2020, 12, 362. [Google Scholar] [CrossRef] [Green Version]
- Nakao, R.; Matsuno, K.; Qiu, Y.; Maruyama, J.; Eguchi, N.; Nao, N.; Kajihara, M.; Yoshii, K.; Sawa, H.; Takada, A.; et al. Putative RNA viral sequences detected in an Ixodes scapularis-derived cell line. Ticks Tick Borne Dis. 2017, 8, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Daveu, R.; Hervet, C.; Sigrist, L.; Sassera, D.; Jex, A.; Labadie, K.; Aury, J.-M.; Plantard, O.; Rispe, C. Sequence diversity and evolution of a group of iflaviruses associated with ticks. Arch. Virol. 2021, 166, 1843–1852. [Google Scholar] [CrossRef]
- Drolesky, R.E.; Holman, P.J.; Wagner, G.G. Babesia bovis infection of a tick cell line studied by electron microscopy. Vitro 1981, 17, 209. [Google Scholar]
- Ribeiro, M.F.B.; Bastos, C.V.; Vasconcelos, M.M.C.; Passos, L.M.F. Babesia bigemina: In vitro multiplication of sporokinetes in Ixodes scapularis (IDE8) cells. Exp. Parasitol. 2009, 122, 192–195. [Google Scholar] [CrossRef]
- Poli, P.; Lenoir, J.; Plantard, O.; Ehrmann, S.; Røed, K.H.; Leinaas, H.P.; Panning, M.; Guiller, A. Strong genetic structure among populations of the tick Ixodes ricinus across its range. Ticks Tick Borne Dis. 2020, 11, 101509. [Google Scholar] [CrossRef]
- Chitimia-Dobler, L.; Rieß, R.; Kahl, O.; Wölfel, S.; Dobler, G.; Nava, S.; Estrada-Pena, A. Ixodes inopinatus—Occurring also outside the Mediterranean region. Ticks Tick Borne Dis. 2018, 9, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Mattila, J.T.; Burkhardt, N.Y.; Hutcheson, H.J.; Munderloh, U.G.; Kurtti, T.J. Isolation of cell lines and a rickettsial endosymbiont from the soft tick Carios capensis (Acari: Argasidae: Ornithodorinae). J. Med. Entomol. 2007, 44, 1091–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell-Sakyi, L.; Ruzek, D.; Gould, E.A. Continuous cell lines from the soft tick Ornithodoros moubata. Exp. Appl. Acarol. 2009, 49, 209–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forth, J.H.; Forth, L.F.; Lycett, S.; Bell-Sakyi, L.; Keil, G.M.; Blome, S.; Calvignac-Spencer, S.; Wissgott, A.; Krause, J.; Höper, D.; et al. Identification of African swine fever virus-like elements in the soft tick genome provides insights into the virus’ evolution. BMC Biol. 2020, 18, 136. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tick Species | Location (Month and Year of Collection) | Host Species |
|---|---|---|
| Argas reflexus | Laboratory colony at FLI (September 2015) | Artificial feeding system 1 |
| Dermacentor reticulatus | Berlin, Germany (November 2018) | Wild boar |
| Hyalomma lusitanicum | Castilla-La Mancha, Spain (February and May 2014) | Deer |
| Hyalomma scupense | Tobía, La Rioja, Spain (February 2016) | Cattle |
| Ixodes ricinus | Tobía, La Rioja, Spain (September 2015) Jubera, La Rioja, Spain (February 2018) | Cattle |
| Rhipicephalus bursa | Jubera, La Rioja, Spain (February 2018) | Cattle |
| Tick Species | Cell Line | Primary Culture (Date) | Culture Medium | 1st Successful Passage | Passage Level Reached | 1st Successful Cryopreservation (Passage Level) |
|---|---|---|---|---|---|---|
| Argas reflexus | ARE/LULS41 | December 2015 | L-15 | 11 months | 9 | 23 months (3) |
| Dermacentor reticulatus | DRE/LULS60 | January 2019 | L-15B | 15 months | 10 | 29 months (3) |
| Hyalomma lusitanicum | HLE/LULS42 | March 2014 | L-15/L-15B 1 | 14 months | 10 2 | 45 months (3) |
| HLE/LULS43 | June 2014 | L-15/H-lac/L-15B 1 | 14 months | 13 2 | 42 months (4) | |
| HLE/LULS48 | March 2014 | L-15/H-lac/L-15B 1 | 26 months | 15 2 | 75 months (12) | |
| Hyalomma scupense | HSE/LULS51 | April 2016 | L-15 | 26 months | 11 | 54 months (4) |
| HSE/LULS59 | April 2016 | L-15/H-lac 1 | 56 months | 9 | 63 months (5) | |
| Ixodes ricinus | IRE/LUAP46 | December 2015 | L-15 | 33 months | 6 2 | 45 months (2) |
| IRE/LULS55 | July 2018 | L-15/H-lac/L-15B 1 | 20 months | 23 | 29 months (3) | |
| Rhipicephalus bursa | RBE/LULS58 | March 2018 | L-15/MEM | 31 months | 8 | 35 months (3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bell-Sakyi, L.; Hartley, C.S.; Khoo, J.J.; Forth, J.H.; Palomar, A.M.; Makepeace, B.L. New Cell Lines Derived from European Tick Species. Microorganisms 2022, 10, 1086. https://doi.org/10.3390/microorganisms10061086
Bell-Sakyi L, Hartley CS, Khoo JJ, Forth JH, Palomar AM, Makepeace BL. New Cell Lines Derived from European Tick Species. Microorganisms. 2022; 10(6):1086. https://doi.org/10.3390/microorganisms10061086
Chicago/Turabian StyleBell-Sakyi, Lesley, Catherine S. Hartley, Jing Jing Khoo, Jan Hendrik Forth, Ana M. Palomar, and Benjamin L. Makepeace. 2022. "New Cell Lines Derived from European Tick Species" Microorganisms 10, no. 6: 1086. https://doi.org/10.3390/microorganisms10061086