
Overview of tRNA Modifications in Chloroplasts



Abstract
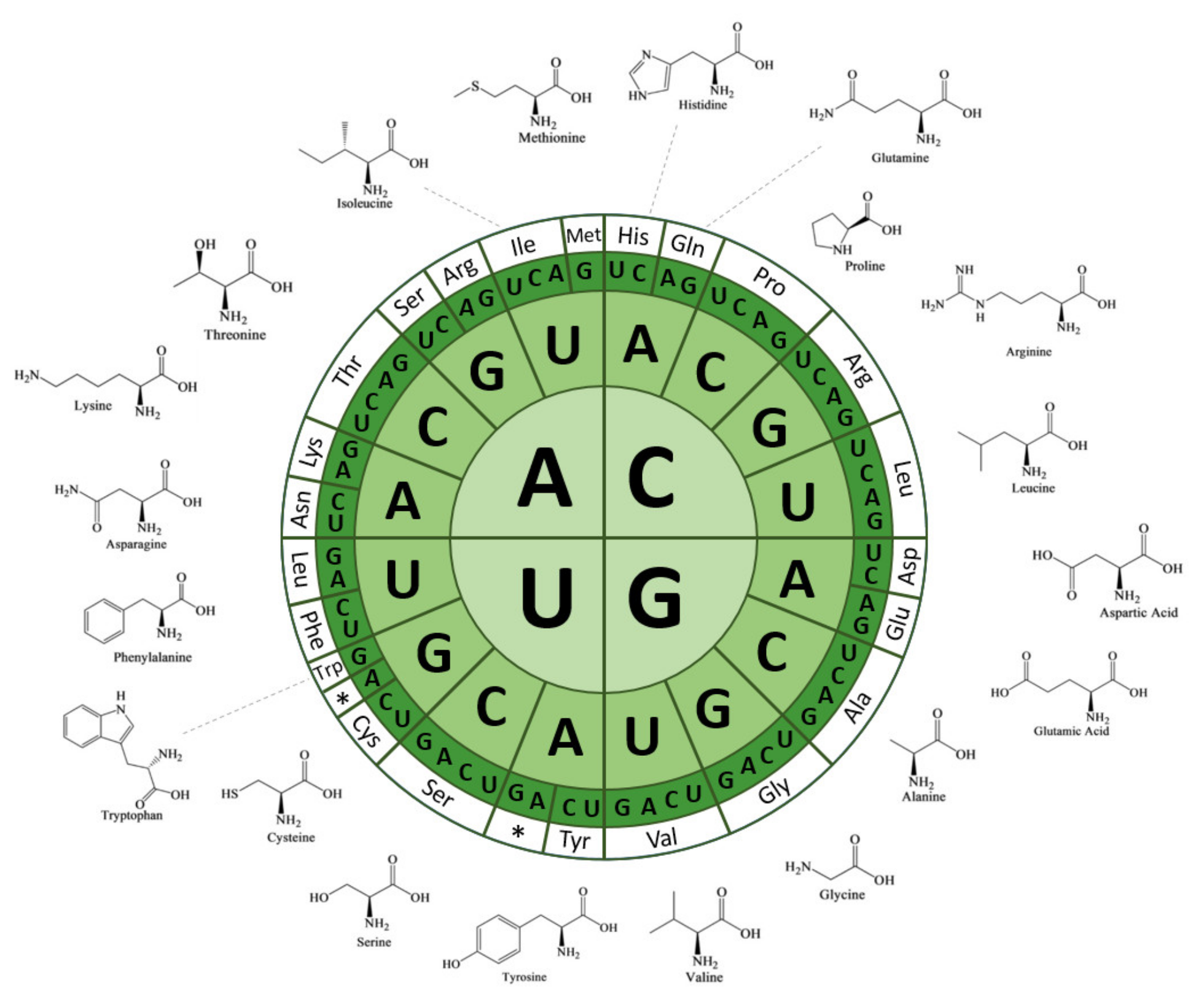
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Common Nucleotide Modifications of tRNA Backbones in Chloroplasts
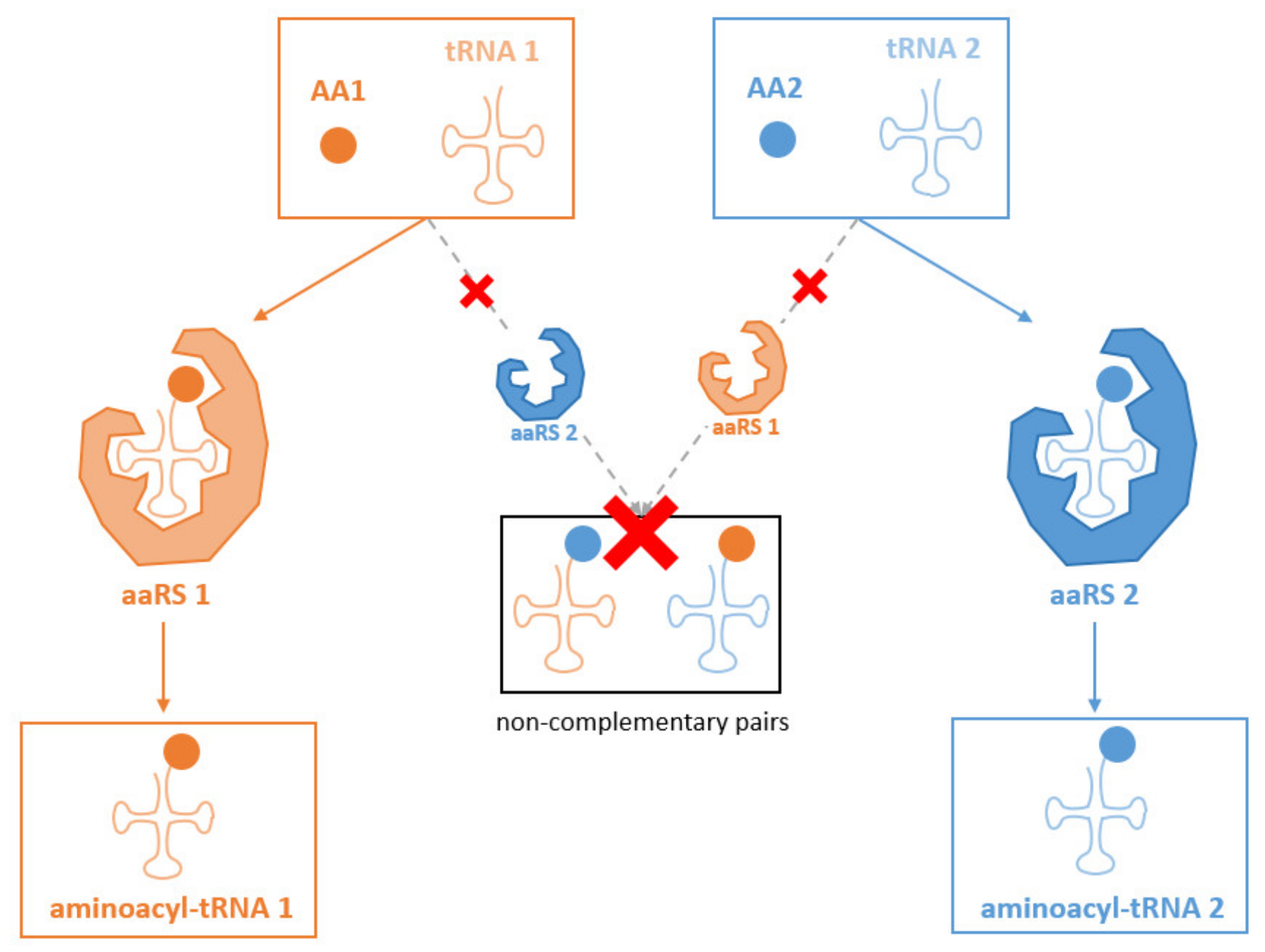
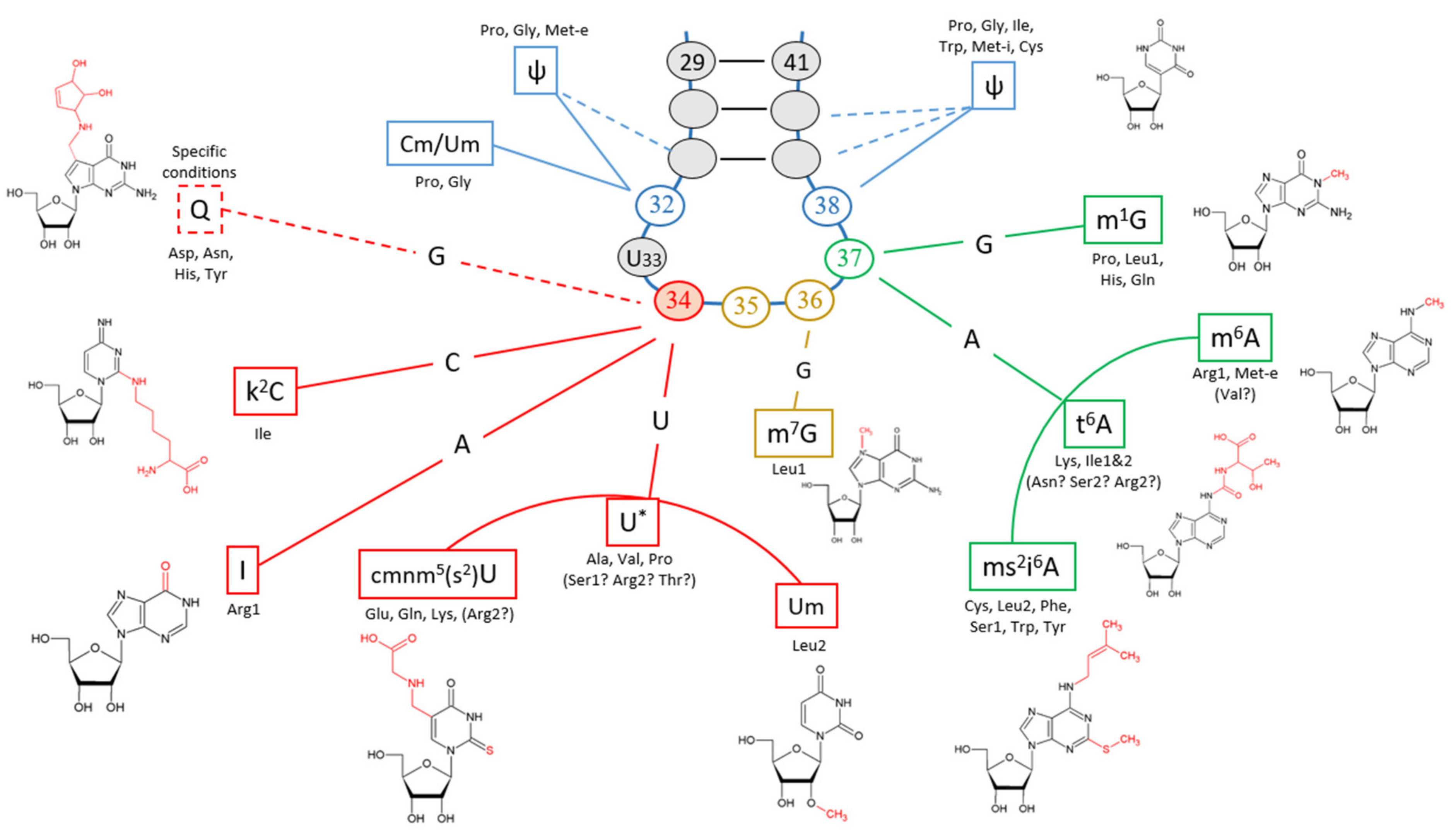
3.2. Modifications of the Anticodon-Adjacent Nucleotide (N37) Maintains the Fidelity of the Genetic Code
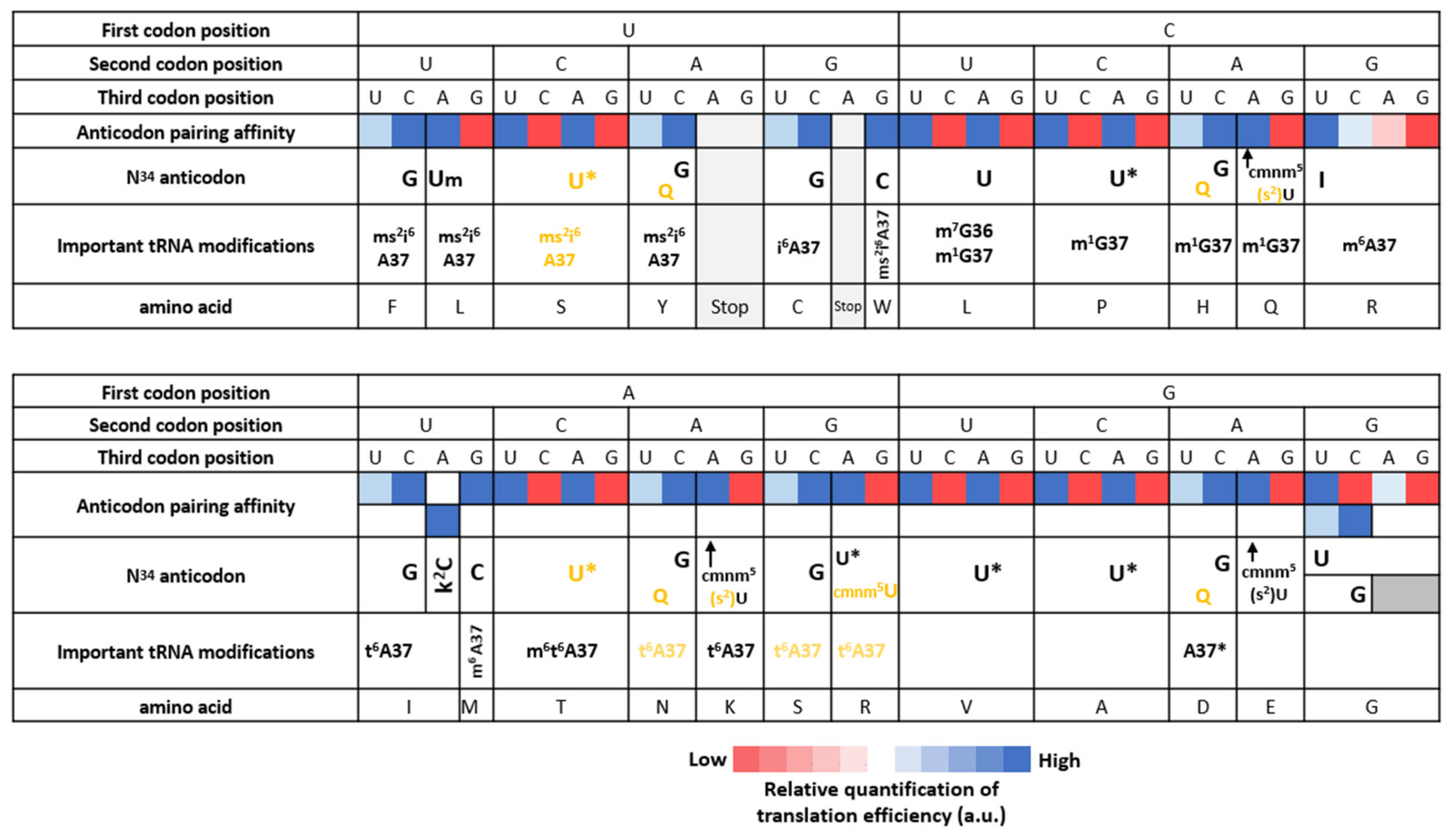
3.3. Nucleotide Modifications Affecting Anticodons Shape Their Deciphering Properties
3.3.1. The Anticodon Base 34 of Single Codon Box
3.3.2. The Anticodon Base 34 of NNU/C Duet Codon Boxes Are Mostly Unmodified
3.3.3. The Anticodon Base 34 of NNA/G Duet Codon Boxes Restricts Deciphering to Purine Codons
3.3.4. The Anticodon Base 34 of Quartet Codon Boxes Expands tRNA Reading Properties by Superwobbling
3.4. Translation Termination
3.5. Comparison of tRNA Modifications within Minimal Genomes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bock, R. Engineering Plastid Genomes: Methods, Tools, and Applications in Basic Research and Biotechnology. Annu. Rev. Plant Biol. 2015, 66, 211–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doron, L.; Segal, N.; Shapira, M. Transgene Expression in Microalgae—From Tools to Applications. Front. Plant Sci. 2016, 7, 505. [Google Scholar] [CrossRef] [PubMed]
- Scaife, M.A.; Smith, A.G. Towards developing algal synthetic biology. Biochem. Soc. Trans. 2016, 44, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Dyo, Y.M.; Purton, S. The algal chloroplast as a synthetic biology platform for production of therapeutic proteins. Microbiology 2018, 164, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Bywater, R.P. Why twenty amino acid residue types suffice(d) to support all living systems. PLoS ONE 2018, 13, e0204883. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.L.; Miller, S.L. Reasons for the occurrence of the twenty coded protein amino acids. J. Mol. Evol. 1981, 17, 273–284. [Google Scholar] [CrossRef]
- Crick, F.H.C. Codon-anticodon pairing: The wobble hypothesis. J. Mol. Biol. 1966, 19, 548–555. [Google Scholar] [CrossRef]
- Agris, P.F. Decoding the genome: A modified view. Nucleic Acids Res. 2004, 32, 223–238. [Google Scholar] [CrossRef] [Green Version]
- Agris, P.F. Wobble position modified nucleosides evolved to select transfer RNA codon recognition: A modified-wobble hypothesis. Biochimie 1991, 73, 1345–1349. [Google Scholar] [CrossRef]
- Agris, P.F. Bringing order to translation: The contributions of transfer RNA anticodon-domain modifications. EMBO Rep. 2008, 9, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Gustilo, E.M.; Vendeix, F.A.; Agris, P.F. tRNA’s modifications bring order to gene expression. Curr. Opin. Microbiol. 2008, 11, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Osawa, S.; Jukes, T.H.; Watanabe, K.; Muto’, A. Recent Evidence for Evolution of the Genetic Code. Microbiol. Rev. 1992, 56, 229–264. [Google Scholar] [CrossRef]
- Osawa, S.; Muto, A.; Ohama, T.; Andachi, Y.; Tanaka, R.; Yamao, F. Prokaryotic genetic code. Exp. Rev. 1990, 46, 1097–1106. [Google Scholar] [CrossRef]
- Yarian, C.; Townsend, H.; Czestkowski, W.; Sochacka, E.; Malkiewicz, A.J.; Guenther, R.; Miskiewicz, A.; Agris, P.F. Accurate Translation of the Genetic Code Depends on tRNA’s Modified Nucleosides. J. Biol. Chem. 2002, 277, 16391–16395. [Google Scholar] [CrossRef] [Green Version]
- Grosjean, H.; Westhof, E. An integrated, structure- and energy-based view of the genetic code. Nucleic Acids Res. 2016, 44, 8020–8040. [Google Scholar] [CrossRef] [Green Version]
- Grosjean, H.; de Crécy-Lagard, V.; Marck, C. Deciphering synonymous codons in the three domains of life: Co-evolution with specific tRNA modification enzymes. FEBS Lett. 2010, 584, 252–264. [Google Scholar] [CrossRef]
- Jukes, T.H. Possibilities for the evolution of the genetic code from a preceding form. Nature 1973, 246, 22–26. [Google Scholar] [CrossRef]
- Cantara, W.A.; Bilbille, Y.; Kim, J.; Kaiser, R.; Leszczyńska, G.; Malkiewicz, A.; Agris, P.F. Modifications modulate anticodon loop dynamics and codon recognition of E. coli tRNAArg1,2. J. Mol. Biol. 2012, 416, 579–597. [Google Scholar] [CrossRef]
- Van Der Gulik, P.T.S.; Hoff, W.D. Unassigned codons, nonsense suppression, and anticodon modifications in the evolution of the genetic code. J. Mol. Evol. 2011, 73, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.J.; Hemez, C.F.; Barber, K.W.; Rinehart, J.; Isaacs, F.J. Organisms with alternative genetic codes resolve unassigned codons via mistranslation and ribosomal rescue. Elife 2018, 7, e34878. [Google Scholar] [CrossRef]
- Muramatsu, T.; Nishikawa, K.; Nemoto, F.; Kuchino, Y.; Nishimura, S.; Miyazawa, T.; Yokoyama, S. Codon and amino-acid specificities of a transfer RNA are both converted by a single post-transcriptional modification. Nature 1988, 336, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Pütz, J.; Florentz, C.; Benseler, F.; Giegé, R. A single methyl group prevents the mischarging of a tRNA. Nat. Struct. Mol. Biol. 1994, 1, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Seno, T.; Agris, P.F.; Söll, D. Involvement of the anticodon region of Escherichia coli tRNAGln and tRNAGlu in the specific interaction with cognate aminoacyl-tRNA synthetase. Biochim. Biophys. Acta Nucleic Acids Protein Synth. 1974, 349, 328–338. [Google Scholar] [CrossRef]
- Sylvers, L.A.; Rogers, K.C.; Shimizu, M.; Ohtsuka, E.; Soll, D. A 2-thiouridine derivative in tRNAGlu is a positive determinant for aminoacylation by Escherichia coli glutamyl-tRNA synthetase. Biochemistry 1993, 32, 3836–3841. [Google Scholar] [CrossRef] [PubMed]
- Cochella, L. An Active Role for tRNA in Decoding Beyond Codon:Anticodon Pairing. Science 2005, 308, 1178–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gromadski, K.B.; Daviter, T.; Rodnina, M.V. A Uniform Response to Mismatches in Codon-Anticodon Complexes Ensures Ribosomal Fidelity. Mol. Cell 2006, 21, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogle, J.M.; Brodersen, D.E.; Clemons, W.M.; Tarry, M.J.; Carter, A.P.; Ramakrishnan, V. Recognition of Cognate Transfer RNA by the 30 S Ribosomal Subunit. Science 2001, 292, 897–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogle, J.M.; Murphy, F.V.; Tarry, M.J.; Ramakrishnan, V. Selection of tRNA by the Ribosome Requires a Transition from an Open to a Closed Form. Cell 2002, 111, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, F.; Krautwurst, S.; Salentin, S.; Haupt, V.J.; Leberecht, C.; Bittrich, S.; Labudde, D.; Schroeder, M. The structural basis of the genetic code: Amino acid recognition by aminoacyl-tRNA synthetases. Sci. Rep. 2020, 10, 12647. [Google Scholar] [CrossRef]
- Pang, Y.L.J.; Poruri, K.; Martinis, S.A. tRNA synthetase: tRNA aminoacylation and beyond. WIREs RNA 2014, 5, 461–480. [Google Scholar] [CrossRef] [Green Version]
- Cusack, S. Aminoacyl-tRNA synthetases. Curr. Opin. Struct. Biol. 1997, 7, 881–889. [Google Scholar] [CrossRef]
- Cusack, S.; Berthet-Colominas, C.; Härtlein, M.; Nassar, N.; Leberman, R. A second class of synthetase structure revealed by X-ray analysis of Escherichia coli seryl-tRNA synthetase at 2.5 Å. Nature 1990, 347, 249–255. [Google Scholar] [CrossRef]
- Eriani, G.; Delarue, M.; Poch, O.; Gangloff, J.; Moras, D. Partition of tRNA synthetases into two classes based on mutually exclusive sets of sequence motifs. Nature 1990, 347, 203–206. [Google Scholar] [CrossRef]
- Ribas de Pouplana, L.; Schimmel, P. Two Classes of tRNA Synthetases Suggested by Sterically Compatible Dockings on tRNA Acceptor Stem. Cell 2001, 104, 191–193. [Google Scholar] [CrossRef] [Green Version]
- O’Donoghue, P.; Luthey-Schulten, Z. On the Evolution of Structure in Aminoacyl-tRNA Synthetases. Microbiol. Mol. Biol. Rev. 2003, 67, 550–573. [Google Scholar] [CrossRef] [Green Version]
- Arnez, J. Structural and functional considerations of the aminoacylation reaction. Trends Biochem. Sci. 1997, 22, 211–216. [Google Scholar] [CrossRef]
- Prætorius- Ibba, M.; Stange-Thomann, N.; Kitabatake, M.; Ali, K.; Söll, I.; Carter, C.W.; Ibba, M.; Söll, D. Ancient Adaptation of the Active Site of Tryptophanyl-tRNA Synthetase for Tryptophan Binding. Biochemistry 2000, 39, 13136–13143. [Google Scholar] [CrossRef]
- Kaiser, F.; Bittrich, S.; Salentin, S.; Leberecht, C.; Haupt, V.J.; Krautwurst, S.; Schroeder, M.; Labudde, D. Backbone Brackets and Arginine Tweezers delineate Class I and Class II aminoacyl tRNA synthetases. PLoS Comput. Biol. 2018, 14, e1006101. [Google Scholar] [CrossRef] [Green Version]
- Pak, D.; Kim, Y.; Burton, Z.F. Aminoacyl-tRNA synthetase evolution and sectoring of the genetic code. Transcription 2018, 9, 205–224. [Google Scholar] [CrossRef]
- Ibba, M.; Söll, D. Aminoacyl-tRNA Synthesis. Annu. Rev. Biochem. 2000, 69, 617–650. [Google Scholar] [CrossRef]
- Dock-Bregeon, A.-C.; Sankaranarayanan, R.; Romby, P.; Caillet, J.; Springer, M.; Rees, B.; Francklyn, C.S.; Ehresmann, C.; Moras, D. Transfer RNA–Mediated Editing in Threonyl-tRNA Synthetase. Cell 2000, 103, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Hadd, A.; Perona, J.J. Coevolution of Specificity Determinants in Eukaryotic Glutamyl- and Glutaminyl-tRNA Synthetases. J. Mol. Biol. 2014, 426, 3619–3633. [Google Scholar] [CrossRef] [PubMed]
- Douglas, S.E.; Turner, S. Molecular Evidence for the Origin of Plastids from a Cyanobacterium-like Ancestor. J. Mol. Evol. 1991, 33, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.W. The evolutionary origins of organelles. Trends Genet. 1989, 5, 294–299. [Google Scholar] [CrossRef]
- Martin, W.; Kowallik, K. Annotated english translation of mereschkowsky’s 1905 paper ‘Über natur und ursprung der chromatophoren impflanzenreiche’. Eur. J. Phycol. 1999, 34, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Scharff, L.B.; Bock, R. Synthetic biology in plastids. Plant J. 2014, 78, 783–798. [Google Scholar] [CrossRef]
- Gallaher, S.D.; Fitz-Gibbon, S.T.; Strenkert, D.; Purvine, S.O.; Pellegrini, M.; Merchant, S.S. High-throughput sequencing of the chloroplast and mitochondrion of Chlamydomonas reinhardtii to generate improved de novo assemblies, analyze expression patterns and transcript speciation, and evaluate diversity among laboratory strains and wild isolates. Plant J. 2018, 93, 545–565. [Google Scholar] [CrossRef] [Green Version]
- Maul, J.E.; Lilly, J.W.; Cui, L.; Depamphilis, C.W.; Miller, W.; Harris, E.H.; Stern, D.B. The Chlamydomonas reinhardtii Plastid Chromosome: Islands of Genes in a Sea of Repeats. Plant Cell 2002, 14, 2659–2679. [Google Scholar] [CrossRef]
- Stern, D.B.; Goldschmidt-Clermont, M.; Hanson, M.R. Chloroplast RNA Metabolism. Annu. Rev. Plant Biol. 2010, 61, 125–155. [Google Scholar] [CrossRef]
- Alkatib, S.; Scharff, L.B.; Rogalski, M.; Fleischmann, T.T.; Matthes, A.; Seeger, S.; Schöttler, M.A.; Ruf, S.; Bock, R. The Contributions of Wobbling and Superwobbling to the Reading of the Genetic Code. PLoS Genet. 2012, 8, e1003076. [Google Scholar] [CrossRef] [Green Version]
- Rogalski, M.; Karcher, D.; Bock, R. Superwobbling facilitates translation with reduced tRNA sets. Nat. Struct. Mol. Biol. 2008, 15, 192–198. [Google Scholar] [CrossRef]
- Fages-Lartaud, M.; Hundvin, K.; Hohmann-Marriott, M.F. Mechanisms governing codon usage bias and the implications for protein expression in the chloroplast. Cell Press Mol. Plant 2022. [Google Scholar]
- Ikemura, T. Correlation between the abundance of Escherichia coli transfer RNAs and the occurrence of the respective codons in its protein genes: A proposal for a synonymous codon choice that is optimal for the E. coli translational system. J. Mol. Biol. 1981, 151, 389–409. [Google Scholar] [CrossRef]
- Duchene, A.-M.; Giritch, A.; Hoffmann, B.; Cognat, V.; Lancelin, D.; Peeters, N.M.; Zaepfel, M.; Marechal-Drouard, L.; Small, I.D. Dual targeting is the rule for organellar aminoacyl-tRNA synthetases in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2005, 102, 16484–16489. [Google Scholar] [CrossRef] [Green Version]
- Francis, M.A.; Suh, E.R.; Dudock, B.S. The Nucleotide Sequence and Characterization of Four Chloroplast tRNAs from the Alga Codium fragile. J. Biol. Chem. 1989, 264, 17243–17249. [Google Scholar] [CrossRef]
- Francis, M.A.; Dudock, B.S. Nucleotide Sequence of a Spinach Chloroplast Isoleucine tRNA. J. Biol. Chem. 1982, 257, 11195–11198. [Google Scholar] [CrossRef]
- Francis, M.; Kashdan, M.; Sprouse, H.; Dudock, B. Nucleotide sequence of a spinach chloroplast proline tRNA. Nucleic Acids Res. 1982, 10, 2755–2758. [Google Scholar] [CrossRef] [Green Version]
- Canaday, J.; Guillemaut, P.; Gloeckler, R.; Weil, J.-H. The nucleotide sequence of spinach chloroplast tryptophan transfer RNA. Nucleic Acids Res. 1981, 9, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Canaday, J.; Guillemaut, P.; Gloeckler, R.; Weil, J.-H. Comparison of the nucleotide sequences of chloroplast tRNAs Phe and tRNAs 3 Leu from spinach and bean. Plant Sci. Lett. 1980, 20, 57–62. [Google Scholar] [CrossRef]
- Guillemaut, P.; Weil, J.H. The nucleotide sequence of the maize and spinach chloroplast isoleucine transfer RNA encoded in the 16S to 23S rDNA spacer. Nucleic Acids Res. 1982, 10, 1653–1659. [Google Scholar] [CrossRef] [Green Version]
- Pillay, D.T.N.; Guillemaut, P.; Weil, J.H. Nucletotide sequences of three soybean chloroplast tRNAs LEU and re-examination of bean tRNA LEU2 chloroplast sequence. Nucleic Acids Res. 1984, 12, 2997–3001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marck, C.; Grosjean, H. tRNomics: Analysis of tRNA genes from 50 genomes of Eukarya, Archaea, and Bacteria reveals anticodon-sparing strategies and domain-specific features. RNA 2002, 8, 1189–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konevega, A.L.; Soboleva, N.G.; Makhno, V.I.; Semenkov, Y.P.; Wintermeyer, W.; Rodnina, M.V.; Katunin, V.I. Purine bases at position 37 of tRNA stabilize codon-anticodon interaction in the ribosomal A site by stacking and Mg2+-dependent interactions. RNA 2004, 10, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schon, A.; Kannangara, C.G.; Gough, S.; Sollt, D. Protein biosynthesis in organelles requires misaminoacylation of tRNA. Nature 1988, 331, 187–190. [Google Scholar] [CrossRef]
- Pfitzinger, H.; Weil, J.H.; Pillay, D.T.N.; Guillemaut, P. Codon recognition mechanisms in plant chloroplasts. Plant Mol. Biol. 1990, 14, 805–814. [Google Scholar] [CrossRef]
- Delannoy, E.; Le Ret, M.; Faivre-Nitschke, E.; Estavillo, G.M.; Bergdoll, M.; Taylor, N.L.; Pogson, B.J.; Small, I.; Imbault, P.; Gualberto, J.M. Arabidopsis tRNA adenosine deaminase arginine edits the wobble nucleotide of chloroplast tRNAArg(ACG) and is essential for efficient chloroplast translation. Plant Cell 2009, 21, 2058–2071. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.M.; Masuda, I.; Gamper, H. Codon-specific translation by m1G37 methylation of tRNA. Front. Genet. 2019, 9, 713. [Google Scholar] [CrossRef]
- Green, G.A.; Jones, D.S. The nucleotide sequences of a cytoplasmic and a chloroplast tRNA-Tyr from Scenedesmus obliquus. Nucleic Acids Res. 1985, 13, 1659–1663. [Google Scholar] [CrossRef] [Green Version]
- Urban, C.; Beier, H. Cysteine tRNAs of plant origin as novel UGA suppressors. Nucleic Acids Res. 1995, 23, 4591–4597. [Google Scholar] [CrossRef] [Green Version]
- Kashdan, M.A.; Pirtle, R.M.; Pirtle, I.L.; Calagan, J.L.; Vreman, H.J.; Dudockfj, B.S. Nucleotide Sequence of a Spinach Chloroplast Threonine tRNA. J. Biol. Chem. 1980, 255, 8831–8835. [Google Scholar] [CrossRef]
- Esberg, B.; Leung, H.-C.E.; Tsui, H.-C.T.; Björk, G.R.; Winkler, M.E. Identification of the miaB Gene, Involved in Methylthiolation of Isopentenylated A37 Derivatives in the tRNA of Salmonella typhimurium and Escherichia coli. J. Bacteriol. 1999, 181, 7256–7265. [Google Scholar] [CrossRef] [Green Version]
- Soderberg, T.; Poulter, C.D. Escherichia coli Dimethylallyl Diphosphate:tRNA Dimethylallyltransferase: Essential Elements for Recognition of tRNA Substrates Within the Anticodon Stem−Loop. Biochemistry 2000, 39, 6546–6553. [Google Scholar] [CrossRef]
- Leung, H.-C.E.; Chen, Y.; Winkler, M.E. Regulation of Substrate Recognition by the MiaA tRNA Prenyltransferase Modification Enzyme of Escherichia coliK-12. J. Biol. Chem. 1997, 272, 13073–13083. [Google Scholar] [CrossRef] [Green Version]
- Mccoy, J.M.; Jones, D.S. The nucleotide sequence of scenedesmus obliquus chloroplast tRNAfMet. Nucleic Acids Res. 1980, 8, 5089–5093. [Google Scholar] [CrossRef] [Green Version]
- Schön, A.; Krupp, G.; Gough, S.; Berry-Lowe, S.; Kannangara, C.G.; Söll, D. The RNA required in the first step of chlorophyll biosynthesis is a chloroplast glutamate tRNA. Nature 1986, 322, 281–284. [Google Scholar] [CrossRef]
- Sprouse, H.M.; Kashdan, M.; Otis, L.; Dudock, B. Nucleotide sequence of a spinach chloroplast valine tRNA. Nucleic Acids Res. 1981, 9, 2543–2547. [Google Scholar] [CrossRef] [Green Version]
- Schön, A.; Gough, S.; Söll, D. Chloroplast tRNA Asp: Nucleotide sequence and variation of in vivo levels during plastid maturation. Plant Mol. Biol. 1992, 20, 601–607. [Google Scholar] [CrossRef]
- Esposito, D. In vivo evidence for the prokaryotic model of extended codon-anticodon interaction in translation initiation. EMBO J. 2003, 22, 651–656. [Google Scholar] [CrossRef] [Green Version]
- Stern, L.; Schulman, L.H. The role of the minor base N4-acetylcytidine in the function of the Escherichia coli noninitiator methionine transfer RNA. J. Biol. Chem. 1978, 253, 6132–6139. [Google Scholar] [CrossRef]
- Chimnaronk, S.; Suzuki, T.; Manita, T.; Ikeuchi, Y.; Yao, M.; Suzuki, T.; Tanaka, I. RNA helicase module in an acetyltransferase that modifies a specific tRNA anticodon. EMBO J. 2009, 28, 1362–1373. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Ren, D.; Jiang, L.; Li, X.; Yao, Y.; Mi, L.; Chen, W.; Mo, A.; Jiang, N.; Yang, J.; et al. A natural variation in pleiotropic developmental defects uncovers a crucial role for chloroplast tRNA modification in translation and plant development. Plant Cell 2020, 32, 2345–2366. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, Y.; Bessho, Y.; Osawa, S. Lack of peptide-release activity responding to codon UGA in Mycoplasma capricolum. Nucleic Acids Res. 1993, 21, 1335–1338. [Google Scholar] [CrossRef] [Green Version]
- Young, R.E.B.; Purton, S. Codon reassignment to facilitate genetic engineering and biocontainment in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2016, 14, 1251–1260. [Google Scholar] [CrossRef] [Green Version]
- Alkatib, S.; Fleischmann, T.T.; Scharff, L.B.; Bock, R. Evolutionary constraints on the plastid tRNA set decoding methionine and isoleucine. Nucleic Acids Res. 2012, 40, 6713–6724. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.; Ch’ng, J.H.; Wahlgren, M.; Thutkawkorapin, J. Frequent GU wobble pairings reduce translation efficiency in Plasmodium falciparum. Sci. Rep. 2017, 7, 723. [Google Scholar] [CrossRef]
- Grosjean, H.J.; De Henau, S.; Crotherst, D.M. On the physical basis for ambiguity in genetic coding interactions (wobble rules/tRNA/codon-anticodon interactions/base pairing). Biochemistry 1978, 75, 610–614. [Google Scholar] [CrossRef] [Green Version]
- Sharp, P.M.; Emery, L.R.; Zeng, K. Forces that influence the evolution of codon bias. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 1203–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadler, M.; Fire, A. Wobble base-pairing slows in vivo translation elongation in metazoans. RNA 2011, 17, 2063–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beier, H.; Zech, U.; Zubrod, E.; Kersten, H. Queuine in plants and plant tRNAs: Differences between embryonic tissue and mature leaves. Plant Mol. Biol. 1987, 8, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Kersten, H.; Kersten, W. Biosynthesis and function of queuine and queuosine tRNAs. In Chromatography and Modification of Nucleosides: Biological Roles and Function of Modification; 1990; Volume 45, pp. B69–B108. Elsevier: Amsterdam, The Netherlands.
- Kirtland, G.M.; Morris, T.D.; Moore, P.H.; O’brian, J.J.; Edmonds, C.G.; Mccloskey, J.A.; Katze, J.R. Novel Salvage of Queuine from Queuosine and Absence of Queuine Synthesis in Chlorella pyrenoidosa and Chlamydomonas reinhardtii. J. Bacteriol. 1988, 170, 5633–5641. [Google Scholar] [CrossRef] [Green Version]
- Vinayak, M.; Pathak, C. Queuosine modification of tRNA: Its divergent role in cellular machinery. Biosci. Rep. 2010, 30, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Begley, T.J.; Dedon, P.C. TRNA modifications regulate translation during cellular stress. FEBS Lett. 2014, 588, 4287–4296. [Google Scholar] [CrossRef] [Green Version]
- Endres, L.; Dedon, P.C.; Begley, T.J. Codon-biased translation can be regulated by wobble-base tRNA modification systems during cellular stress responses. RNA Biol. 2015, 12, 603–614. [Google Scholar] [CrossRef] [Green Version]
- Dewez, M.; Bauer, F.; Dieu, M.; Raes, M.; Vandenhaute, J.; Hermand, D. The conserved Wobble uridine tRNA thiolase Ctu1-Ctu2 is required to maintain genome integrity. Proc. Natl. Acad. Sci. USA 2008, 105, 5459–5464. [Google Scholar] [CrossRef] [Green Version]
- Lim, V.I. Analysis of Action of Wobble Nucleoside Modifications on Codon-Anticodon Pairing within the Ribosome. J. Mol. Biol. 1994, 240, 8–19. [Google Scholar] [CrossRef]
- Kurata, S.; Weixlbaumer, A.; Ohtsuki, T.; Shimazaki, T.; Wada, T.; Kirino, Y.; Takai, K.; Watanabe, K.; Ramakrishnan, V.; Suzuki, T. Modified uridines with C5-methylene substituents at the first position of the tRNA anticodon stabilize U·G wobble pairing during decoding. J. Biol. Chem. 2008, 283, 18801–18811. [Google Scholar] [CrossRef] [Green Version]
- Takai, K.; Yokoyama, S. Roles of 5-substituents of tRNA wobble uridines in the recognition of purine-ending codons. Nucleic Acids Res. 2003, 31, 6383–6391. [Google Scholar] [CrossRef] [Green Version]
- Björk, G.R.; Hagervall, T.G. Transfer RNA Modification. EcoSal Plus 2005, 1, 2. [Google Scholar] [CrossRef]
- Lagerkvist, U. “Two out of three”: An alternative method for codon reading (codon-anticodon recognition/translational fidelity/wobbling/organization of the genetic code). Proc. Natl. Acad. Sci. USA 1978, 75, 1759–1762. [Google Scholar] [CrossRef] [Green Version]
- Claesson, C.; Lustig, F.; Boré, T.; Simonsson, C.; Barciszewska, M.; Lagerkvist, U. Glycine Codon Discrimination and the Nucleotide in Position 32 of the Anticodon Loop. J. Mol. Biol. 1995, 247, 191–196. [Google Scholar] [CrossRef]
- Lustig, F.; Bortn, T.; Claesson, C.; Simonsson, C.; Barciszewskat, M.; Lagerkvist, U. The nucleotide in position 32 of the tRNA anticodon loop determines ability of anticodon UCC to discriminate among glycine codons. Proc. Natl. Acad. Sci. USA 1993, 90, 3343–3347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.K.; Lustig, F.; Akesson, B.; Lagerkvist, U. Codon-Anticodon Recognition in the Valine Codon Family. J. Biol. Chem. 1977, 252, 471–478. [Google Scholar] [CrossRef]
- Mitra, S.K.; Lustig, F.; Akesson, B.; Axberg, T.; Lagerkvists, U. Relative Efficiency of Anticodons in Reading the Valine Codons during Protein Synthesis in vitro. J. Biol. Chem. 1979, 254, 6397–6401. [Google Scholar] [CrossRef]
- Weixlbaumer, A.; Murphy IV, F.V.; Dziergowska, A.; Malkiewicz, A.; Vendeix, F.A.P.; Agris, P.F.; Ramakrishnan, V. Mechanism for expanding the decoding capacity of transfer RNAs by modification of uridines. Nat. Struct. Mol. Biol. 2007, 14, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Murphy, F.V.; Ramakrishnan, V. Structure of a purine-purine wobble base pair in the decoding center of the ribosome. Nat. Struct. Mol. Biol. 2004, 11, 1251–1252. [Google Scholar] [CrossRef]
- Ikemura, T. Correlation Between the Abundance of Yeast Transfer RNAs and the Occurrence of the Respective Codons in Protein Genes Differences in Synonymous Codon Choice Patterns of Yeast and Escherichia coli with Reference to the Abundance of Isoaccepting Transfer RNAs. J. Mol. Biol. 1982, 158–573. [Google Scholar] [CrossRef]
- Karcher, D.; Bock, R. Identification of the chloroplast adenosine-to-inosine tRNA editing enzyme. RNA 2009, 15, 1251–1257. [Google Scholar] [CrossRef] [Green Version]
- Ohyamat, K.; Fukuzawat, H.; Kohchi, T.; Sano, T.; Sano, S.; Shirai, H.; Umesonoy, K.; Shiki, Y.; Takeuchi, M.; Chang, Z.; et al. Structure and Organization of Marchantia polymorpha Chloroplast Genome I. Cloning and Gene Identification. J. Mol. Biol. 1988, 203, 281–298. [Google Scholar] [CrossRef]
- Scolnick, E.; Tompkins, R.; Caskey, T.; Nirenberg, M. Release factors differing in specificity for terminator codons. Proc. Natl. Acad. Sci. USA 1968, 61, 768–774. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Ito, K.; Isaksson, L.A. Emerging Understanding of Translation Termination. Cell 1996, 87, 147–150. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Uno, M.; Nakamura, Y. A tripeptide ‘anticodon’ deciphers stop codons in messenger RNA. Nature 2000, 403, 680–684. [Google Scholar] [CrossRef]
- Fu, Z.; Indrisiunaite, G.; Kaledhonkar, S.; Shah, B.; Sun, M.; Chen, B.; Grassucci, R.A.; Ehrenberg, M.; Frank, J. The structural basis for release-factor activation during translation termination revealed by time-resolved cryogenic electron microscopy. Nat. Commun. 2019, 10, 2579. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, R.; Yamazaki, T.; Myouga, F.; Ito, T.; Ito, K.; Satou, M.; Kobayashi, M.; Nagata, N.; Yoshida, S.; Nagashima, A.; et al. Chloroplast ribosome release factor 1 (AtcpRF1) is essential for chloroplast development. Plant Mol. Biol. 2007, 64, 481–497. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, J.; Xia, X. Coevolution between Stop Codon Usage and Release Factors in Bacterial Species. Mol. Biol. Evol. 2016, 33, 2357–2367. [Google Scholar] [CrossRef] [Green Version]
- Young, R.; Purton, S. CITRIC: Cold-inducible translational readthrough in the chloroplast of Chlamydomonas reinhardtii using a novel temperature-sensitive transfer RNA. Microb. Cell Fact. 2018, 17, 186. [Google Scholar] [CrossRef] [Green Version]
- Letzring, D.P.; Dean, K.M.; Grayhack, E.J. Control of translation efficiency in yeast by codon-anticodon interactions. RNA 2010, 16, 2516–2528. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modified Base | Codons | E. Coli | M. Capricolum | Mammalian Mitochondria | Chloroplast | ||
|---|---|---|---|---|---|---|---|
| N37 | A37 | (U1NN) | ms2i6 A | ms2i6 A | ms2i6 A | ||
| m6 A | i6 A | i6 A | |||||
| unmodified A | |||||||
| (C1NN) | m2 A | m6 A | unmodified A | m6 A | |||
| (G1NN) | m2 A | m6 A | A* | ||||
| unmodified A | unmodified A | unmodified A | unmodified A | ||||
| (A1NN) | m6t6 A | m6t6 A | |||||
| ct6A | ct6A | ct6A | t6A | ||||
| m6 A | m6 A | ||||||
| unmodified A | unmodified A | (unmodified A) | |||||
| G37 | (C1NN) | m1G | m1G | m1G | m1G | ||
| N34 | NNU/C (duet boxes) | NNU/C | unmodified G | unmodified G | unmodified G | unmodified G | |
| NAU/C | Q | unmodified G | Q | unmodified G (maybe Q) | |||
| GluQ | |||||||
| NNA/G (duet boxes) | mnm5s2U | ||||||
| mnm5Um | cmnm5U | τm5U | cmnm5U | ||||
| cmnm5s2U | cmnm5s2U | τm5s2U | cmnm5s2U | ||||
| cmnm5Um | cmnm5Um | Um | |||||
| Cm | |||||||
| NNU/A/C/G (quartet boxes) | cmo5U | unmodified U | unmodified U | unmodified U | |||
| unmodified G | U* | ||||||
| unmodified C | |||||||
| quartet Arginine | I | I | unmodified U | I | |||
| NNG | Trp | Cm | Cm | τm5U | unmodified C | ||
| cmnm5Um | |||||||
| Met | ac4C | unmodified C | f5C | unmodified C | |||
| Isoleucine AUC | k2C | k2C | (NA) | k2C | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fages-Lartaud, M.; Hohmann-Marriott, M.F. Overview of tRNA Modifications in Chloroplasts. Microorganisms 2022, 10, 226. https://doi.org/10.3390/microorganisms10020226
Fages-Lartaud M, Hohmann-Marriott MF. Overview of tRNA Modifications in Chloroplasts. Microorganisms. 2022; 10(2):226. https://doi.org/10.3390/microorganisms10020226
Chicago/Turabian StyleFages-Lartaud, Maxime, and Martin Frank Hohmann-Marriott. 2022. "Overview of tRNA Modifications in Chloroplasts" Microorganisms 10, no. 2: 226. https://doi.org/10.3390/microorganisms10020226