
Cryptosporidiosis: From Prevention to Treatment, a Narrative Review


Abstract
:1. Introduction
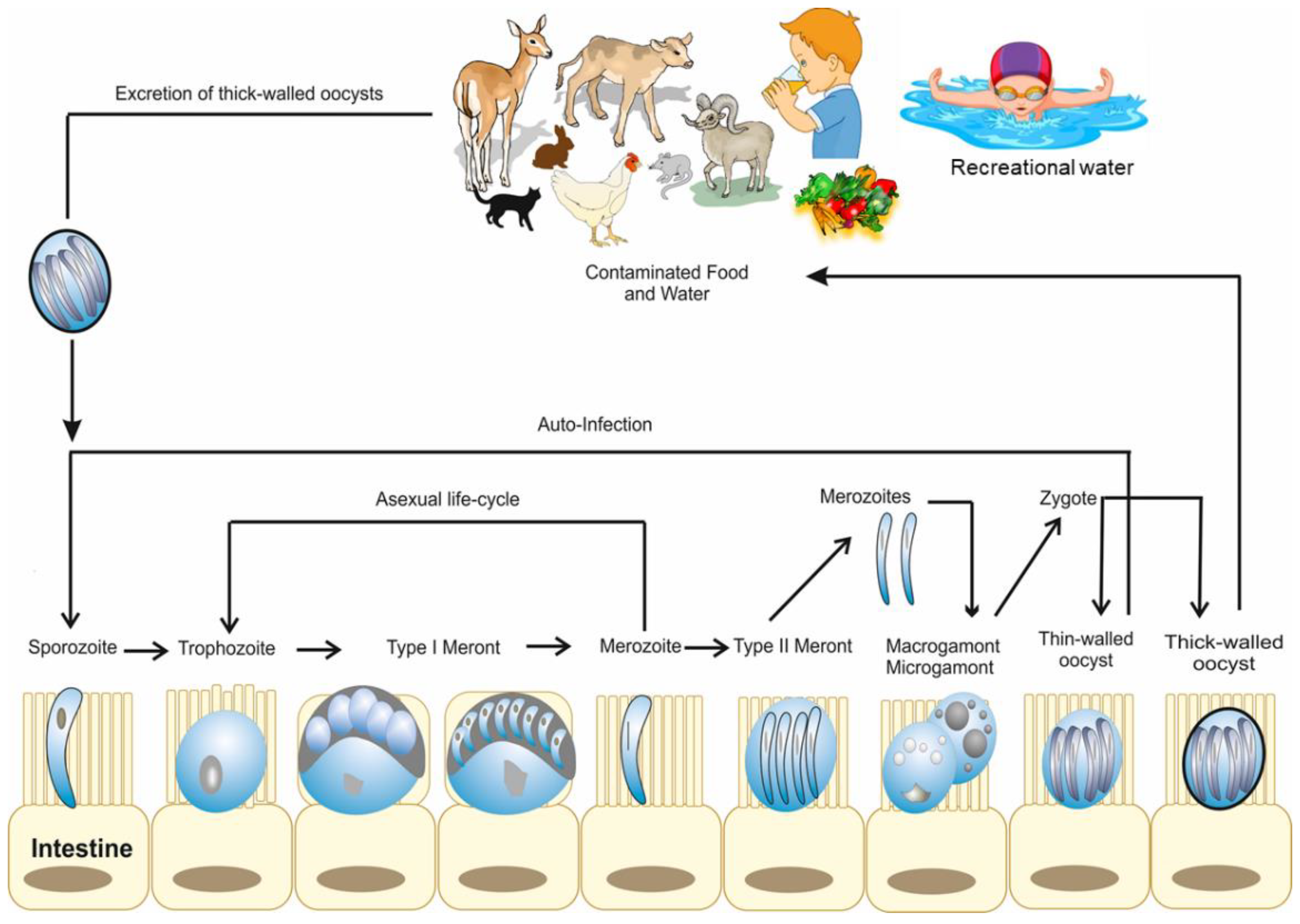
2. Life Cycle and Developmental Stages of Cryptosporidium
3. Pathogenesis of Cryptosporidium
4. Species, Genotypes/Subtypes, and Host Spectrum of Cryptosporidium

{kind=link}
| Cryptosporidium spp. | Hosts | Sporulated Oocyst Size (µm) | Location |
|---|---|---|---|
| C. hominis | Humans | 4.5 × 5.5 | Small intestine |
| C. parvum | Ruminants, humans, deer | 4.5 × 5.5 | Small intestine |
| C. bovis | Ruminants | 4.2–4.8 × 4.8–5.4 | Small intestine |
| C. andersoni | Ruminants, camel | 5.5 × 7.4 | Abomasum |
| C. ryanae | Ruminants | 3.2 × 3.7 | Small intestine |
| C. xiaoi | Sheep | 3.9 × 3.4 | Small intestine |
| C. ubiquitum | Sheep/wildlife | 5.2 × 4.9 | Small intestine |
| C. meleagridis | Chicken, turkey, humans | 4.5–5.0 × 4.6–5.2 | Intestine |
| C. baileyi | Birds | 6.4 × 6.2 | Cloaca, bursa, respiratory tract |
| C. galli | Birds | 8.0–8.5 × 6.2–6.4 | Proventriculus |
| C. avium | Birds | 5.3–6.9 × 4.3–5.5 | Intestine |
| C. ornithophilusis | Ostrich | 6.13 × 5.15 | Intestine |
| C. proventriculi | Psittaciformes birds | 7.4 × 5.8 | Proventriculus |
| Avian genotype II | Birds | 6.0–6.5 × 4.8–6.6 | Intestine |
| Avian genotype IV | Birds | 8.25 × 6.3 | Intestine |
| Eurasian woodcock genotype | Birds | 8.5 × 6.4 | Intestine |
| C. suis | Pigs, humans | 5.1 × 4.4 | Small intestine |
| C. wrairi | Guinea pigs | 4.0–5.0 × 4.8–5.6 | Small intestine |
| C. cuniculus | Rabbits | 5.9 × 5.4 | Small intestine |
| C. canis | Canids, humans, mink, fox, coyote | 5.0 × 4.7 | Small intestine |
| C. felis | Felids, humans | 4.5 × 5.0 | Small intestine |
| C. saurophilum | Lizards, snakes | 4.2–5.2 × 4.4–5.6 | Intestinal and cloacal mucosa |
| C. serpentis | Snakes, lizards | 4.8–5.6 × 5.6–6.6 | Stomach |
| C. fayeri | Red Kangaroo, marsupials | 4.9 × 4.3 | Intestine |
| C. macropodum | Marsupials | 4.9 × 5.4 | Small intestine |
| C. muris | Rodents, humans | 5.6 × 7.4 | Stomach |
| C. ratti | Rodents | 4.5–5.4 × 4.5–5.0 | Small intestine |
| C. tyzzeri | Mice | 4.6 × 4.2 | Small intestine |
| C. molnari | Fish | 4.7 × 4.5 | Stomach |
| C. scophithalmi | Fish | 3.0–4.7 × 3.7–5.0 | Intestine |
| C. nasorum | Fish | 4.3 × 3.2 | Intestine |
5. Epidemiology of Cryptosporidiosis
5.1. Source of Infection and Mode of Transmission
5.1.1. In Humans
5.1.2. In Animals
5.2. Clinical Signs and High-Risk Groups of Cryptosporidiosis
5.2.1. In Humans
5.2.2. In Livestock Animals
5.2.3. In Poultry
5.3. Outbreaks of Cryptosporidiosis in Humans
6. Diagnosis of Cryptosporidium
7. Prevention and Treatment of Cryptosporidium Infection
7.1. One Health Approach for the Control of Cryptosporidiosis
7.2. Preventive Measures for Cryptosporidium Infection
7.3. Treatment of Cryptosporidium Infection
7.4. Vaccines Development
8. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Helmy, Y.A.; El-Adawy, H.; Abdelwhab, E.M. A Comprehensive Review of Common Bacterial, Parasitic and Viral Zoonoses at the Human-Animal Interface in Egypt. Pathogens 2017, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Gharpure, R.; Perez, A.; Miller, A.D.; Wikswo, M.E.; Silver, R.; Hlavsa, M.C. Cryptosporidiosis Outbreaks—United States, 2009–2017. Mmwr. Morb. Mortal Wkly Rep. 2019, 68, 568–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, J.; Hague, H.; Hudgin, G.; Ross, L.; Moore, D. International Perspectives: An Outbreak of Cryptosporidium at a Recreational Water Park in Niagara Region, Canada. J. Environ. Heath 2013, 75, 28–33. [Google Scholar]
- Savioli, L.; Smith, H.; Thompson, A. Giardia and Cryptosporidium join the ‘Neglected Diseases Initiative’. Trends Parasitol. 2006, 22, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Klotz, C.; Wilking, H.; Krucken, J.; Nockler, K.; Von Samson-Himmelstjerna, G.; Zessin, K.H.; Aebischer, T. Epidemiology of Giardia duodenalis infection in ruminant livestock and children in the Ismailia province of Egypt: Insights by genetic characterization. Parasites Vectors 2014, 7, 321. [Google Scholar] [CrossRef] [Green Version]
- Helmy, Y.A.; Spierling, N.G.; Schmidt, S.; Rosenfeld, U.M.; Reil, D.; Imholt, C.; Jacob, J.; Ulrich, R.G.; Aebischer, T.; Klotz, C. Occurrence and distribution of Giardia species in wild rodents in Germany. Parasites Vectors 2018, 11, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadonaite, B.; Ritchie, H.; Roser, M. Diarrheal Diseases. In Our World in Data. Available online: https://ourworldindata.org/diarrheal-diseases (accessed on 26 January 2021).
- Mosier, D.A.; Oberst, R.D. Cryptosporidiosis. A global challenge. Ann. N. Y. Acad. Sci. 2000, 916, 102–111. [Google Scholar] [CrossRef]
- Ryan, U.; Hijjawi, N.; Xiao, L. Foodborne cryptosporidiosis. Int. J. Parasitol. 2018, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Putignani, L.; Menichella, D. Global distribution, public health and clinical impact of the protozoan pathogen Cryptosporidium. Interdiscip. Perspect. Infect. Dis. 2010, 2010, 753512. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, W.R.; Hoxie, N.J.; Proctor, M.E.; Gradus, M.S.; Blair, K.A.; Peterson, D.E. A massive outbreak in Milwaukee of Cryptosporidium infection transmitted through the public water supply. N. Engl. J. Med. 1994, 331, 161–167. [Google Scholar] [CrossRef]
- Peng, M.M.; Xiao, L.; Freeman, A.R.; Arrowood, M.J.; Escalante, A.A.; Weltman, A.C.; Ong, C.S.; Mac Kenzie, W.R.; Lal, A.A.; Beard, C.B. Genetic polymorphism among Cryptosporidium parvum isolates: Evidence of two distinct human transmission cycles. Emerg. Infect. Dis. 1997, 3, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Palmer, C.S.; O’Handley, R. The public health and clinical significance of Giardia and Cryptosporidium in domestic animals. Vet. J. 2008, 177, 18–25. [Google Scholar] [CrossRef]
- CDC. Crypto Outbreaks Linked to Swimming Have Doubled since 2014. 2017. Available online: https://www.cdc.gov/media/releases/2017/p0518-cryptosporidium-outbreaks.html (accessed on 26 January 2021).
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States–Major pathogens. Emerg. Inf. Dis. 2011, 17, 7. [Google Scholar] [CrossRef] [PubMed]
- DuPont, H.L. Persistent diarrhea: A clinical review. JAMA Netw. 2016, 315, 2712–2723. [Google Scholar] [CrossRef] [PubMed]
- Khalil, I.A.; Troeger, C.; Rao, P.C.; Blacker, B.F.; Brown, A.; Brewer, T.G.; Colombara, D.V.; De Hostos, E.L.; Engmann, C.; Guerrant, R.L.; et al. Morbidity, mortality, and long-term consequences associated with diarrhoea from Cryptosporidium infection in children younger than 5 years: A meta-analyses study. Lancet Glob. Health 2018, 6, e758–e768. [Google Scholar] [CrossRef] [Green Version]
- Sunnotel, O.; Lowery, C.J.; Moore, J.E.; Dooley, J.S.; Xiao, L.; Millar, B.C.; Rooney, P.J.; Snelling, W.J. Cryptosporidium. Lett. Appl. Microbiol. 2006, 43, 7–16. [Google Scholar] [CrossRef]
- Fayer, R.; Trout, J.M.; Jenkins, M.C. Infectivity of Cryptosporidium parvum oocysts stored in water at environmental temperatures. J. Parasitol. 1998, 84, 1165–1169. [Google Scholar] [CrossRef]
- Smith, H.V.; Caccio, S.M.; Cook, N.; Nichols, R.A.; Tait, A. Cryptosporidium and Giardia as foodborne zoonoses. Vet. Parasitol. 2007, 149, 29–40. [Google Scholar] [CrossRef]
- Gulliksen, S.M.; Jor, E.; Lie, K.I.; Hamnes, I.S.; Loken, T.; Akerstedt, J.; Osteras, O. Enteropathogens and risk factors for diarrhea in Norwegian dairy calves. J. Dairy Sci. 2009, 92, 5057–5066. [Google Scholar] [CrossRef] [Green Version]
- Shahiduzzaman, M.; Daugschies, A. Therapy and prevention of cryptosporidiosis in animals. Vet. Parasitol. 2012, 188, 203–214. [Google Scholar] [CrossRef]
- Stockdale, H.D.; Spencer, J.A.; Blagburn, B.L. Prophylaxis and Chemotherapy. In Cryptosporidium and Cryptosporidiosis; Fayer, R., Xiao, L., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 255–287. [Google Scholar]
- Silverlas, C.; Bjorkman, C.; Egenvall, A. Systematic review and meta-analyses of the effects of halofuginone against calf cryptosporidiosis. Prevent. Vet. Med. 2009, 91, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Brainard, J.; Hammer, C.C.; Hunter, P.R.; Katzer, F.; Hurle, G.; Tyler, K. Efficacy of halofuginone products to prevent or treat cryptosporidiosis in bovine calves: A systematic review and meta-analyses. Parasitology 2021, 148, 408–419. [Google Scholar] [CrossRef]
- Sparks, H.; Nair, G.; Castellanos-Gonzalez, A.; White, A.C., Jr. Treatment of Cryptosporidium: What We Know, Gaps, and the Way Forward. Curr. Trop. Med. Rep. 2015, 2, 181–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijjawi, N.S.; Meloni, B.P.; Ryan, U.M.; Olson, M.E.; Thompson, R.C. Successful in vitro cultivation of Cryptosporidium andersoni: Evidence for the existence of novel extracellular stages in the life cycle and implications for the classification of Cryptosporidium. Int. J. Parasitol. 2002, 32, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.V.; Corcoran, G.D. New drugs and treatment for cryptosporidiosis. Curr. Opin. Infect. Dis. 2004, 17, 557–564. [Google Scholar] [CrossRef]
- Leitch, G.J.; He, Q. Cryptosporidiosis-an overview. J. Biomed. Res. 2012, 25, 1–16. [Google Scholar] [CrossRef]
- Lendner, M.; Daugschies, A. Cryptosporidium infections: Molecular advances. Parasitology 2014, 141, 1511–1532. [Google Scholar] [CrossRef]
- Tandel, J.; English, E.D.; Sateriale, A.; Gullicksrud, J.A.; Beiting, D.P.; Sullivan, M.C.; Pinkston, B.; Striepen, B. Life cycle progression and sexual development of the apicomplexan parasite Cryptosporidium parvum. Nat. Microbiol. 2019, 4, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, S.P.; Chen, X.M. The cell biology of Cryptosporidium infection. Microbes Infect. 2011, 13, 721–730. [Google Scholar] [CrossRef] [Green Version]
- Fayer, R.; Xiao, L. Cryptosporidium and Cryptosporidiosis, 2nd ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: London, UK, 2007; p. 580. [Google Scholar]
- Upton, S.J.; Current, W.L. The species of Cryptosporidium (Apicomplexa: Cryptosporidiidae) infecting mammals. J. Parasitol. 1985, 71, 625–629. [Google Scholar] [CrossRef]
- Current, W.L.; Reese, N.C. A comparison of endogenous development of three isolates of Cryptosporidium in suckling mice. J. Protozool. 1986, 33, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R.; Ungar, B.L. Cryptosporidium spp. and cryptosporidiosis. Microbiol. Rev. 1986, 50, 458–483. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.V.; Nichols, R.A.; Grimason, A.M. Cryptosporidium excystation and invasion: Getting to the guts of the matter. Trends Parasitol. 2005, 21, 133–142. [Google Scholar] [CrossRef]
- O’Hara, S.P.; Yu, J.R.; Lin, J.J. A novel Cryptosporidium parvum antigen, CP2, preferentially associates with membranous structures. Parasitol. Res. 2004, 92, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Zapata, F.; Perkins, M.E.; Riojas, Y.A.; Wu, T.W.; Le Blancq, S.M. The Cryptosporidium parvum ABC protein family. Mol. Biochem. Parasitol. 2002, 120, 157–161. [Google Scholar] [CrossRef]
- Fayer, R.; Gasbarre, L.; Pasquali, P.; Canals, A.; Almeria, S.; Zarlenga, D. Cryptosporidium parvum infection in bovine neonates: Dynamic clinical, parasitic and immunologic patterns. Int. J. Parasitol. 1998, 28, 49–56. [Google Scholar] [CrossRef]
- Sponseller, J.K.; Griffiths, J.K.; Tzipori, S. The evolution of respiratory Cryptosporidiosis: Evidence for transmission by inhalation. Clin. Microbiol. Rev. 2014, 27, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Dillingham, R.A.; Lima, A.A.; Guerrant, R.L. Cryptosporidiosis: Epidemiology and impact. Microbes Infect. 2002, 4, 1059–1066. [Google Scholar] [CrossRef]
- Nascimento, M.F.; Ginoris, Y.P.; Brandão, C.C.S. Cryptosporidium Oocysts Removal by Upflow Direct Filtration: Pilot Scale Assessment. Water 2020, 12, 1328. [Google Scholar] [CrossRef]
- Okhuysen, P.C.; Chappell, C.L.; Crabb, J.H.; Sterling, C.R.; DuPont, H.L. Virulence of three distinct Cryptosporidium parvum isolates for healthy adults. J. Infect. Dis. 1999, 180, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.A.; Atwill, E.R.; Kirk, J.H.; Brahmbhatt, D.; Alonso, L.H.; Hou, L.; Singer, M.D.; Miller, T.D. Prophylactic use of decoquinate for infections with Cryptosporidium parvum in experimentally challenged neonatal calves. J. Am. Vet. Med. Assoc. 2003, 15, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A. Epidemiological investigations on the public health significance of Cryptosporidium parasites in livestock and people in the Ismailia Canal Zone of Egypt. Ph.D. Dissertation, Freie Universität, Berlin, Germany, 2014; 77p. [Google Scholar]
- Etzold, M.; Lendner, M.; Daugschies, A.; Dyachenko, V. CDPKs of Cryptosporidium parvum-stage-specific expression in vitro. Parasitol. Res. 2014, 113, 2525–2533. [Google Scholar] [CrossRef] [PubMed]
- Tzipori, S.; Ward, H. Cryptosporidiosis: Biology, pathogenesis and disease. Microbes Infect. 2002, 4, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R.; Speer, C.A.; Dubey, J.P. General Biology of Cryptosporidium. In Cryptosporidiosis of Man and Animals; Dubey, J.P., Speer, C.A., Fayer, R., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 1–29. [Google Scholar]
- Tzipori, S.; Smith, M.; Halpin, C.; Angus, K.W.; Sherwood, D.; Campbell, I. Experimental cryptosporidiosis in calves: Clinical manifestations and pathological findings. Vet. Record 1983, 112, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Heine, J.; Pohlenz, J.F.; Moon, H.W.; Woode, G.N. Enteric lesions and diarrhea in gnotobiotic calves monoinfected with Cryptosporidium species. J. Infect. Dis. 1984, 150, 768–775. [Google Scholar] [CrossRef]
- Fayer, R.; Santin, M. Cryptosporidium xiaoi n. sp. (Apicomplexa: Cryptosporidiidae) in sheep (Ovis aries). Vet. Parasitol. 2009, 164, 192–200. [Google Scholar] [CrossRef]
- Elwin, K.; Hadfield, S.J.; Robinson, G.; Crouch, N.D.; Chalmers, R.M. Cryptosporidium viatorum n. sp. (Apicomplexa: Cryptosporidiidae) among travellers returning to Great Britain from the Indian subcontinent, 2007–2011. Int. J. Parasitol. 2012, 42, 675–682. [Google Scholar] [CrossRef]
- Lebbad, M.; Beser, J.; Insulander, M.; Karlsson, L.; Mattsson, J.G.; Svenungsson, B.; Axen, C. Unusual cryptosporidiosis cases in Swedish patients: Extended molecular characterization of Cryptosporidium viatorum and Cryptosporidium chipmunk genotype I. Parasitology 2013, 140, 1735–1740. [Google Scholar] [CrossRef] [Green Version]
- Adamu, H.; Petros, B.; Zhang, G.; Kassa, H.; Amer, S.; Ye, J.; Feng, Y.; Xiao, L. Distribution and clinical manifestations of Cryptosporidium species and subtypes in HIV/AIDS patients in Ethiopia. PLoS Neg. Trop. Dis. 2014, 8, e2831. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.A.; Karanis, P. Cryptosporidium and Cryptosporidiosis: The Perspective from the Gulf Countries. Int. J. Environ. Res. Public Heath 2020, 17, 6824. [Google Scholar] [CrossRef]
- Fayer, R. General Biology. In Cryptosporidium and Cryptosporidiosis; Fayer, R., Xiao, L., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 1–42. [Google Scholar]
- Helmy, Y.A.; Krucken, J.; Abdelwhab, E.M.; von Samson-Himmelstjerna, G.; Hafez, H.M. Molecular diagnosis and characterization of Cryptosporidium spp. in turkeys and chickens in Germany reveals evidence for previously undetected parasite species. PLoS ONE 2017, 12, e0177150. [Google Scholar] [CrossRef] [PubMed]
- Mamedova, S.; Karanis, P. Cryptosporidium spp. Infections in Livestock and Wild Animals in Azerbaijan Territory. Environ. Sci. Proceed. 2020, 2, 44. [Google Scholar]
- Zahedi, A.; Paparini, A.; Jian, F.; Robertson, I.; Ryan, U. Public health significance of zoonotic Cryptosporidium species in wildlife: Critical insights into better drinking water management. Int. J. Parasitol. Parasites Wildl 2016, 5, 88–109. [Google Scholar] [CrossRef] [Green Version]
- Lendner, M.; Etzold, M.; Daugschies, A. Cryptosporidiosis—An update. Berl. Und Munch. Tierarztl. Wochenschr. 2011, 124, 473–484. [Google Scholar]
- Helmy, Y.A.; Krucken, J.; Nockler, K.; von Samson-Himmelstjerna, G.; Zessin, K.H. Molecular epidemiology of Cryptosporidium in livestock animals and humans in the Ismailia province of Egypt. Vet. Parasitol. 2013, 193, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Fayer, R.; Xiao, L. Cryptosporidium species in humans and animals: Current understanding and research needs. Parasitology 2014, 141, 1667–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. Cryptosporidiosis. Available online: https://www.cdc.gov/dpdx/cryptosporidiosis/index.html (accessed on 1 August 2020).
- Nichols, R.A.; Connelly, L.; Sullivan, C.B.; Smith, H.V. Identification of Cryptosporidium species and genotypes in Scottish raw and drinking waters during a one-year monitoring period. Appl. Environ. Microbiol. 2010, 76, 5977–5986. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, O.; Gonzalez-Diaz, M.; Garibay-Escobar, A.; Burgara-Estrella, A.; Cano, M.; Durazo, M.; Bernal, R.M.; Hernandez, J.; Xiao, L. Molecular Characterization of Cryptosporidium spp. in Children from Mexico. PLoS ONE 2014, 9, e96128. [Google Scholar] [CrossRef] [Green Version]
- Jex, A.R.; Gasser, R.B. Genetic richness and diversity in Cryptosporidium hominis and C. parvum reveals major knowledge gaps and a need for the application of “next generation” technologies-research review. Biotechnol. Adv. 2010, 28, 17–26. [Google Scholar] [CrossRef]
- Xiao, L. Molecular epidemiology of cryptosporidiosis: An update. Exp. Parasitol. 2010, 124, 80–89. [Google Scholar] [CrossRef]
- Silverlås, C. Cryptosporidium Infection in Dairy Cattle. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2010. [Google Scholar]
- Zahedi, A.; Gofton, A.W.; Greay, T.; Monis, P.; Oskam, C.; Ball, A.; Bath, A.; Watkinson, A.; Robertson, I.; Ryan, U. Profiling the diversity of Cryptosporidium species and genotypes in wastewater treatment plants in Australia using next generation sequencing. Sci. Total Environ. 2018, 644, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, A.; Gofton, A.W.; Jian, F.; Paparini, A.; Oskam, C.; Ball, A.; Robertson, I.; Ryan, U. Next Generation Sequencing uncovers within-host differences in the genetic diversity of Cryptosporidium gp60 subtypes. Int. J. Parasitol. 2017, 47, 601–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, U.; Zahedi, A.; Feng, Y.; Xiao, L. An update on zoonotic Cryptosporidium species and genotypes in humans. Animal 2021, 11, 3307. [Google Scholar] [CrossRef] [PubMed]
- Caccio, S.M.; Widmer, G. Cryptosporidium: Parasite and Disease; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Holubová, N.; Tůmová, L.; Sak, B.; Hejzlarová, A.; Konečný, R.; McEvoy, J.; Kváč, M. Description of Cryptosporidium ornithophilus n. sp. (Apicomplexa: Cryptosporidiidae) in farmed ostriches. Parasites Vectors 2020, 13, 340. [Google Scholar] [CrossRef]
- Holubová, N.; Sak, B.; Horčičková, M.; Hlásková, L.; Květoňová, D.; Menchaca, S.; McEvoy, J.; Kváč, M. Cryptosporidium avium n. sp.(Apicomplexa: Cryptosporidiidae) in birds. Parasitol. Res. 2016, 115, 2243–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, U. Cryptosporidium in birds, fish and amphibians. Exp. Parasitol. 2010, 124, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Karanis, P. Cryptosporidium: Waterborne and foodborne transmission and worldwide outbreaks. In Proceedings of the Euro-Mediterranean Conference for Environmental Integration, Sousse, Tunisia, 22–25 November 2017; pp. 41–44. [Google Scholar]
- Graczyk, T.K.; Fayer, R.; Cranfield, M.R.; Mhangami-Ruwende, B.; Knight, R.; Trout, J.M.; Bixler, H. Filth flies are transport hosts of Cryptosporidium parvum. Emerg. Infect. Dis. 1999, 5, 726–727. [Google Scholar] [CrossRef]
- Duhain, G.L.; Minnaar, A.; Buys, E.M. Effect of chlorine, blanching, freezing, and microwave heating on Cryptosporidium parvum viability inoculated on green peppers. J. Food Prot. 2012, 75, 936–941. [Google Scholar] [CrossRef]
- Macarisin, D.; Bauchan, G.; Fayer, R. Spinacia oleracea L. leaf stomata harboring Cryptosporidium parvum oocysts: A potential threat to food safety. Appl. Environ. Microbiol. 2010, 76, 555–559. [Google Scholar] [CrossRef] [Green Version]
- Macarisin, D.; Santin, M.; Bauchan, G.; Fayer, R. Infectivity of Cryptosporidium parvum oocysts after storage of experimentally contaminated apples. J. Food Prot. 2010, 73, 1824–1829. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo Lorenzo, M.J.; Ares-Mazas, E.; de Maturana, I.V.M. Detection of oocysts and IgG antibodies to Cryptosporidium parvum in asymptomatic adult cattle. Vet. Parasitol. 1993, 47, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Villacorta, I.; Ares-Mazas, E.; Lorenzo, M.J. Cryptosporidium parvum in cattle, sheep and pigs in Galicia (N.W. Spain). Vet. Parasitol. 1991, 38, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P.L.; Schwarzenberger, R. Free-living amoebae as vectors of cryptosporidia. Parasitol. Res. 2011, 109, 499–504. [Google Scholar] [CrossRef]
- Pumipuntu, N.; Piratae, S. Cryptosporidiosis: A zoonotic disease concern. Vet. World 2018, 11, 681–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santin, M.; Trout, J.M.; Fayer, R. A longitudinal study of cryptosporidiosis in dairy cattle from birth to 2 years of age. Vet. Parasitol. 2008, 155, 15–23. [Google Scholar] [CrossRef]
- Fayer, R.; Santin, M.; Trout, J.M. Prevalence of Cryptosporidium species and genotypes in mature dairy cattle on farms in eastern United States compared with younger cattle from the same locations. Vet. Parasitol. 2007, 145, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Santin, M.; Trout, J.M. Livestock. In Cryptosporidium and Cryptosporidiosis; Fayer, R., Xiao, L., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 451–483. [Google Scholar]
- Korpe, P.S.; Valencia, C.; Haque, R.; Mahfuz, M.; McGrath, M.; Houpt, E.; Kosek, M.; McCormick, B.J.J.; Penataro Yori, P.; Babji, S.; et al. Epidemiology and Risk Factors for Cryptosporidiosis in Children From 8 Low-income Sites: Results From the MAL-ED Study. Clin. Infect. Dis. 2018, 67, 1660–1669. [Google Scholar] [CrossRef] [Green Version]
- Lal, A.; Fearnley, E.; Wilford, E. Local weather, flooding history and childhood diarrhoea caused by the parasite Cryptosporidium spp.: A systematic review and meta-analysis. Sci. Total Environ. 2019, 674, 300–306. [Google Scholar] [CrossRef]
- CDC. Parasites-Cryptosporidium (also known as “Crypto”). Available online: https://www.cdc.gov/parasites/crypto/infection-sources.html (accessed on 1 August 2020).
- Chen, X.M.; Keithly, J.S.; Paya, C.V.; LaRusso, N.F. Cryptosporidiosis. N. Engl. J. Med. 2002, 346, 1723–1731. [Google Scholar] [CrossRef]
- Tzipori, S. Cryptosporidiosis in perspective. Adv. Parasitol. 1988, 27, 63–129. [Google Scholar]
- Current, W.L.; Garcia, L.S. Cryptosporidiosis. Clin. Microbiol. Rev. 1991, 4, 325–358. [Google Scholar] [CrossRef]
- Farthing, M.J. Clinical aspects of Human Cryptosporidiosis. In Cryptosporidiosis and Microsporidiosis; Tetry, F., Ed.; Karger: Basel, Switzerland, 2000; pp. 50–74. [Google Scholar]
- Hunter, P.R.; Nichols, G. Epidemiology and clinical features of Cryptosporidium infection in immunocompromised patients. Clin. Microbiol. Rev. 2002, 15, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Cama, V.A.; Ross, J.M.; Crawford, S.; Kawai, V.; Chavez-Valdez, R.; Vargas, D.; Vivar, A.; Ticona, E.; Navincopa, M.; Williamson, J.; et al. Differences in clinical manifestations among Cryptosporidium species and subtypes in HIV-infected persons. J. Infect. Dis. 2007, 196, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Hunter, P.R.; Hughes, S.; Woodhouse, S.; Raj, N.; Syed, Q.; Chalmers, R.M.; Verlander, N.Q.; Goodacre, J. Health sequelae of human cryptosporidiosis in immunocompetent patients. Clin. Infect. Dis. 2004, 39, 504–510. [Google Scholar] [CrossRef]
- Hatam-Nahavandi, K.; Ahmadpour, E.; Carmena, D.; Spotin, A.; Bangoura, B.; Xiao, L. Cryptosporidium infections in terrestrial ungulates with focus on livestock: A systematic review and meta-analysis. Parasites Vectors 2019, 12, 453. [Google Scholar] [CrossRef]
- Kaske, M.; Smolka, K.; Andresen, U. Neonatal diarrhea in the calf—Etiology and path physiology. Der. Prakt. Tierarzt 2008, 89, 852–859. [Google Scholar]
- De Graaf, D.C.; Vanopdenbosch, E.; Ortega-Mora, L.M.; Abbassi, H.; Peeters, J.E. A review of the importance of cryptosporidiosis in farm animals. Int. J. Parasitol. 1999, 29, 1269–1287. [Google Scholar] [CrossRef]
- McDonald, V. Host cell-mediated responses to infection with Cryptosporidium. Parasit. Immunol. 2000, 22, 597–604. [Google Scholar] [CrossRef]
- Bertoni, E.; Barragán, A.A.; Bok, M.; Vega, C.; Martínez, M.; Gil, J.F.; Cimino, R.O.; Parreño, V. Assessment of Influential Factors for Scours Associated with Cryptosporidium sp., Rotavirus and Coronavirus in Calves from Argentinean Dairy Farms. Animals 2021, 11, 2652. [Google Scholar] [CrossRef]
- Lombardelli, J.A.; Tomazic, M.L.; Schnittger, L.; Tiranti, K.I. Prevalence of Cryptosporidium parvum in dairy calves and GP60 subtyping of diarrheic calves in central Argentina. Parasitol. Res. 2019, 118, 2079–2086. [Google Scholar] [CrossRef]
- Xiao, L.; Fayer, R.; Ryan, U.; Upton, S.J. Cryptosporidium taxonomy: Recent advances and implications for public health. Clin. Microbiol. Rev. 2004, 17, 72–97. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, D.S.; Blagburn, B.L. Cryptosporidiosis in birds. In Cryptosporidiosis Man and Animals; CRC Press: Boca Raton, FL, USA, 1990; pp. 133–148. [Google Scholar]
- Makino, I.; Abe, N.; Reavill, D.R. Cryptosporidium avian genotype III as a possible causative agent of chronic vomiting in peach-faced lovebirds (Agapornis roseicollis). Avian Dis. 2010, 54, 1102–1107. [Google Scholar] [CrossRef]
- Jellison, K.L.; Distel, D.L.; Hemond, H.F.; Schauer, D.B. Phylogenetic analysis of the hypervariable region of the 18S rRNA gene of Cryptosporidium oocysts in feces of Canada geese (Branta canadensis): Evidence for five novel genotypes. App. Environ. Microbiol. 2004, 70, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Jellison, K.L.; Lynch, A.E.; Ziemann, J.M. Source tracking identifies deer and geese as vectors of human-infectious Cryptosporidium genotypes in an urban/suburban watershed. Environ. Sci. Technol. 2009, 43, 4267–4272. [Google Scholar] [CrossRef]
- Zhou, L.; Kassa, H.; Tischler, M.L.; Xiao, L. Host-adapted Cryptosporidium spp. in Canada geese (Branta canadensis). Appl. Environ. Microbiol. 2004, 70, 4211–4215. [Google Scholar] [CrossRef]
- da Cunha, M.J.R.; Cury, M.C.; Santín, M. Molecular characterization of Cryptosporidium spp. in poultry from Brazil. Res. Vet. Sci. 2018, 118, 331–335. [Google Scholar] [CrossRef]
- Santana, B.N.; Kurahara, B.; Nakamura, A.A.; da Silva Camargo, V.; Ferrari, E.D.; da Silva, G.S.; Nagata, W.B.; Meireles, M.V. Detection and characterization of Cryptosporidium species and genotypes in three chicken production systems in Brazil using different molecular diagnosis protocols. Prev. Vet. Med. 2018, 151, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Xu, M.; Jiang, Y.; Liu, H.; Yuan, Z.; Sun, L.; Cao, J.; Shen, Y. Prevalence and Genetic Characterization of Cryptosporidium, Giardia and Enterocytozoon in Chickens From Ezhou, Hubei, China. Fron. Vet. Sci. 2020, 7, 30. [Google Scholar] [CrossRef]
- AL-Ghezey, M.R.; Al-Zubaidi, M.T. Molecular detection of Cryptosporidium parasite in chickens (Broiler and layer) in Thi-Qar province. Iraqi J. Vet. Sci. 2020, 34, 441–445. [Google Scholar]
- Altamimi, M.K.; Al-Zubaidi, M.T. Cryptosporidium Spp.: Conventional and molecular and molecular detection in wild pigeons (COLUMBA LIVIA) in Babylon province, Iraq. Plan. Arch. 2020, 20, 1544–1548. [Google Scholar]
- Shahbazi, P.; Aligolzadeh, A.; Khordadmehr, M.; Farhang, H.H.; Katiraee, F. Molecular study and genotyping of Cryptosporidium baileyi and Cryptosporidium parvum from free-range and commercial broiler chickens in Guilan province, Iran. Comp. Immunol. Microbiol. Infect. Dis. 2020, 69, 101411. [Google Scholar] [CrossRef]
- Jian, Y.; Zhang, X.; Li, X.; Schou, C.; Charalambidou, I.; Ma, L.; Karanis, P. Occurrence of Cryptosporidium and Giardia in wild birds from Qinghai Lake on the Qinghai-Tibetan Plateau, China. Parasitol. Res. 2021, 120, 615–628. [Google Scholar] [CrossRef]
- Kabir, M.H.B.; Han, Y.; Lee, S.-H.; Nugraha, A.B.; Recuenco, F.; Murakoshi, F.; Xuan, X.; Kato, K. Prevalence and molecular characterization of Cryptosporidium species in poultry in Bangladesh. One Health 2020, 9, 100122. [Google Scholar] [CrossRef]
- Liao, C.; Wang, T.; Koehler, A.V.; Fan, Y.; Hu, M.; Gasser, R.B. Molecular investigation of Cryptosporidium in farmed chickens in Hubei Province, China, identifies ‘zoonotic’subtypes of C. meleagridis. Parasit Vectors 2018, 11, 484. [Google Scholar] [CrossRef]
- Reboredo-Fernandez, A.; Ares-Mazas, E.; Caccio, S.M.; Gomez-Couso, H. Occurrence of Giardia and Cryptosporidium in wild birds in Galicia (Northwest Spain). Parasitology 2015, 142, 917–925. [Google Scholar] [CrossRef]
- Li, J.; Lin, X.; Zhang, L.; Qi, N.; Liao, S.; Lv, M.; Wu, C.; Sun, M. Molecular characterization of Cryptosporidium spp. in domestic pigeons (Columba livia domestica) in Guangdong Province, Southern China. Parasitol. Res. 2015, 114, 2237–2241. [Google Scholar] [CrossRef]
- Máca, O.; Pavlásek, I. First finding of spontaneous infections with Cryptosporidium baileyi and C. meleagridis in the red-legged partridge Alectoris rufa from an aviary in the Czech Republic. Vet. Parasitol. 2015, 209, 164–168. [Google Scholar] [CrossRef]
- Wang, L.; Xue, X.; Li, J.; Zhou, Q.; Yu, Y.; Du, A. Cryptosporidiosis in broiler chickens in Zhejiang Province, China: Molecular characterization of oocysts detected in fecal samples. Parasite 2014, 21, 36. [Google Scholar] [CrossRef]
- Qi, M.; Huang, L.; Wang, R.; Xiao, L.; Xu, L.; Li, J.; Zhang, L. Natural infection of Cryptosporidium muris in ostriches (Struthio camelus). Vet. Parasitol. 2014, 205, 518–522. [Google Scholar] [CrossRef]
- Laatamna, A.E.; Holubova, N.; Sak, B.; Kvac, M. Cryptosporidium meleagridis and C. baileyi (Apicomplexa) in domestic and wild birds in Algeria. Folia Parasitol. 2017, 64, 2017.018. [Google Scholar] [CrossRef]
- Baroudi, D.; Khelef, D.; Goucem, R.; Adjou, K.T.; Adamu, H.; Zhang, H.; Xiao, L. Common occurrence of zoonotic pathogen Cryptosporidium meleagridis in broiler chickens and turkeys in Algeria. Vet. Parasitol. 2013, 196, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, F.; Zhao, J.; Qi, M.; Ning, C.; Zhang, L.; Xiao, L. Cryptosporidium spp. in quails (Coturnix coturnix japonica) in Henan, China: Molecular characterization and public health significance. Vet. Parasitol. 2012, 187, 534–537. [Google Scholar] [CrossRef]
- Qi, M.; Wang, R.; Ning, C.; Li, X.; Zhang, L.; Jian, F.; Sun, Y.; Xiao, L. Cryptosporidium spp. in pet birds: Genetic diversity and potential public health significance. Exp. Parasitol. 2011, 128, 336–340. [Google Scholar] [CrossRef] [PubMed]
- da Paixão Sevá, A.; Funada, M.R.; Richtzenhain, L.; Guimarães, M.B.; de Oliveira Souza, S.; Allegretti, L.; Sinhorini, J.A.; Duarte, V.V.; Soares, R.M. Genotyping of Cryptosporidium spp. from free-living wild birds from Brazil. Vet. Parasitol. 2011, 175, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Qi, M.; Jingjing, Z.; Sun, D.; Ning, C.; Zhao, J.; Zhang, L.; Xiao, L. Prevalence of Cryptosporidium baileyi in ostriches (Struthio camelus) in Zhengzhou, China. Vet. Parasitol. 2011, 175, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Jian, F.; Sun, Y.; Hu, Q.; Zhu, J.; Wang, F.; Ning, C.; Zhang, L.; Xiao, L. Large-scale survey of Cryptosporidium spp. in chickens and Pekin ducks (Anas platyrhynchos) in Henan, China: Prevalence and molecular characterization. Avian Pathol. 2010, 39, 447–451. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, J.; Giddings, C. Cryptosporidium in commercially produced turkeys on-farm and postslaughter. Lett. App. Microbiol. 2009, 48, 302–306. [Google Scholar] [CrossRef]
- Nakamura, A.A.; Simões, D.C.; Antunes, R.G.; da Silva, D.C.; Meireles, M.V. Molecular characterization of Cryptosporidium spp. from fecal samples of birds kept in captivity in Brazil. Vet. Parasitol. 2009, 166, 47–51. [Google Scholar] [CrossRef]
- Ng, J.; Pavlasek, I.; Ryan, U. Identification of novel Cryptosporidium genotypes from avian hosts. Appl. Environ. Microbiol. 2006, 72, 7548–7553. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, A.; Ryan, U. Cryptosporidium—An update with an emphasis on foodborne and waterborne transmission. Res. Vet. Sci. 2020, 132, 500–512. [Google Scholar] [CrossRef]
- Feng, Y.; Li, N.; Roellig, D.M.; Kelley, A.; Liu, G.; Amer, S.; Tang, K.; Zhang, L.; Xiao, L. Comparative genomic analysis of the IId subtype family of Cryptosporidium parvum. Int. J. Parasitol. 2017, 47, 281–290. [Google Scholar] [CrossRef]
- CDC. National Outbreak Reporting System Dashboard; U.S. Department of Health and Human Services, CDC: Atlanta, Georgia.
- Doman, B.; Selvage, W.D.; Ferrreira, J.; Roelling, D.M.; Kahler, A. Cryptosporidium Outbreak Associated with the Consumption of Unpasteurized Cow Milk, New Mexico. Available online: https://cste.confex.com/cste/2017/webprogram/Paper8610.html (accessed on 1 August 2020).
- Fill, M.-M.A.; Lloyd, J.; Chakraverty, T.; Sweat, D.; Manners, J.; Garman, K.; Hlavsa, M.C.; Roellig, D.M.; Dunn, J.R.; Schaffner, W. Cryptosporidiosis outbreak associated with a single hotel. J. Environ. Health 2017, 79, 16–23. [Google Scholar]
- Rosenthal, M.; Pedersen, R.; Leibsle, S.; Hill, V.; Carter, K.; Roellig, D.M. Cryptosporidiosis Associated with Consumption of Unpasteurized Goat Milk—Idaho, 2014. Morb. Mortal. Wkly. Rep. 2015, 64, 194. [Google Scholar]
- Chalmers, R.M.; Robinson, G.; Elwin, K.; Elson, R. Analysis of the Cryptosporidium spp. and gp60 subtypes linked to human outbreaks of cryptosporidiosis in England and Wales, 2009 to 2017. Parasites Vectors 2019, 12, 1–13. [Google Scholar] [CrossRef]
- McKerr, C.; Adak, G.K.; Nichols, G.; Gorton, R.; Chalmers, R.M.; Kafatos, G.; Cosford, P.; Charlett, A.; Reacher, M.; Pollock, K.G. An outbreak of Cryptosporidium parvum across England & Scotland associated with consumption of fresh pre-cut salad leaves, May 2012. PLoS ONE 2015, 10, e0125955. [Google Scholar]
- Chalmers, R.M. Waterborne outbreaks of cryptosporidiosis. Ann. Dell’istituto Super. Di Sanita 2012, 48, 429–446. [Google Scholar] [CrossRef]
- McCann, R.; Jones, R.; Snow, J.; Cleary, P.; Burgess, S.; Bothra, V.; Chalmers, R. An outbreak of cryptosporidiosis at a swimming club–can rapid field epidemiology limit the spread of illness? Epidemiol. Infect. 2014, 142, 51–55. [Google Scholar] [CrossRef]
- Whitworth, J. Food Safety News. Available online: https://www.foodsafetynews.com/2020/03/five-foodborne-outbreaks-added-to-cryptosporidium-rise-in-sweden/ (accessed on 1 August 2020).
- Rehn, M.; Wallensten, A.; Widerström, M.; Lilja, M.; Grunewald, M.; Stenmark, S.; Kark, M.; Lindh, J. Post-infection symptoms following two large waterborne outbreaks of Cryptosporidium hominis in Northern Sweden, 2010–2011. BMC Public Health 2015, 15, 529. [Google Scholar] [CrossRef] [Green Version]
- Widerström, M.; Schönning, C.; Lilja, M.; Lebbad, M.; Ljung, T.; Allestam, G.; Ferm, M.; Björkholm, B.; Hansen, A.; Hiltula, J. Large outbreak of Cryptosporidium hominis infection transmitted through the public water supply, Sweden. Emerg. Infec. Dis. 2014, 20, 581. [Google Scholar] [CrossRef]
- Gherasim, A.; Lebbad, M.; Insulander, M.; Decraene, V.; Kling, A.; Hjertqvist, M.; Wallensten, A. Two geographically separated food-borne outbreaks in Sweden linked by an unusual Cryptosporidium parvum subtype, October 2010. Eurosurveillance 2012, 17, 20318. [Google Scholar] [CrossRef]
- Mosnier, E.; Martin, N.; Razakandrainibe, R.; Dalle, F.; Roux, G.; Buteux, A.; Favennec, L.; Brousse, P.; Guarmit, B.; Blanchet, D. Cryptosporidiosis outbreak in immunocompetent children from a remote area of French Guiana. Am. J Trop. Med. Hyg. 2018, 98, 1727–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gertler, M.; Dürr, M.; Renner, P.; Poppert, S.; Askar, M.; Breidenbach, J.; Frank, C.; Preußel, K.; Schielke, A.; Werber, D. Outbreak of Cryptosporidium hominis following river flooding in the city of Halle (Saale), Germany, August 2013. BMC Infect. Dis. 2015, 15, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahon, M.; Doyle, S. Waterborne outbreak of cryptosporidiosis in the South East of Ireland: Weighing up the evidence. Irish J. Med. Sci. 2017, 186, 989–994. [Google Scholar] [CrossRef]
- Robertson, L.; Temesgen, T.T.; Tysnes, K.; Eikås, J. An apple a day: An outbreak of cryptosporidiosis in Norway associated with self-pressed apple juice. Epidemiol. Infect. 2019, 147, e139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, H.; Johansen, Ø.; Vold, L.; Robertson, L.; Anthonisen, I.; Nygard, K. Second outbreak of infection with a rare Cryptosporidium parvum genotype in schoolchildren associated with contact with lambs/goat kids at a holiday farm in Norway. Epidemiol. Infect. 2014, 142, 2105–2113. [Google Scholar] [CrossRef]
- Åberg, R.; Sjöman, M.; Hemminki, K.; Pirnes, A.; Räsänen, S.; Kalanti, A.; Pohjanvirta, T.; Caccio, S.; Pihlajasaari, A.; Toikkanen, S. Cryptosporidium parvum caused a large outbreak linked to frisée salad in Finland, 2012. Zoonoses Public Health 2015, 62, 618–624. [Google Scholar] [CrossRef]
- Moon, S.; Kwak, W.; Lee, S.; Kim, W.; Oh, J.; Youn, S.-K. Epidemiological characteristics of the first water-borne outbreak of cryptosporidiosis in Seoul, Korea. J. Korean Med. Sci. 2013, 28, 983. [Google Scholar] [CrossRef] [Green Version]
- Ng-Hublin, J.; Hargrave, D.; Combs, B.; Ryan, U. Investigation of a swimming pool-associated cryptosporidiosis outbreak in the Kimberley region of Western Australia. Epidemiol. Infect. 2015, 143, 1037–1041. [Google Scholar] [CrossRef] [Green Version]
- Chartier, C.; Rieux, A.; Delafosse, A.; Lehebel, A.; Paraud, C. Detection of Cryptosporidium oocysts in fresh calf faeces: Characteristics of two simple tests and evaluation of a semi-quantitative approach. Vet. J. 2013, 198, 148–152. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Krucken, J.; Nockler, K.; von Samson-Himmelstjerna, G.; Zessin, K.H. Comparison between two commercially available serological tests and polymerase chain reaction in the diagnosis of Cryptosporidium in animals and diarrhoeic children. Parasitol. Res. 2014, 113, 211–216. [Google Scholar] [CrossRef]
- El-Missiry, A.; Abd El-Hameed, L.; Saad, G.; El-Badry, A.; Helmy, Y.; Shehata, M. Molecular genetic characterization of human Cryptosporidium isolates and their respective demographic, environmental and clinical manifestations in Egyptian diarrheic patients. Parasitol. United J. 2019, 12, 187–196. [Google Scholar] [CrossRef]
- Papini, R.; Cardini, G. Evaluation of a rapid Cryptosporidium/Giardia immunochromatographic test for diagnosis of giardiasis in dogs. Revue Méd. Vét. 2006, 157, 490–493. [Google Scholar]
- Brescia, C.C.; Griffin, S.M.; Ware, M.W.; Varughese, E.A.; Egorov, A.I.; Villegas, E.N. Cryptosporidium propidium monoazide-PCR, a molecular biology-based technique for genotyping of viable Cryptosporidium oocysts. Appl. Environ. Microbiol. 2009, 75, 6856–6863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caccio, S.M.; Thompson, R.C.; McLauchlin, J.; Smith, H.V. Unravelling Cryptosporidium and Giardia epidemiology. Trends Parasitol. 2005, 21, 430–437. [Google Scholar] [CrossRef]
- Egyed, Z.; Sreter, T.; Szell, Z.; Beszteri, B.; Dobos-Kovacs, M.; Marialigeti, K.; Cornelissen, A.W.; Varga, I. Polyphasic typing of Cryptosporidium baileyi: A suggested model for characterization of cryptosporidia. J. Parasitol. 2002, 88, 237–243. [Google Scholar] [CrossRef]
- Morgan, U.M.; Constantine, C.C.; O’Donoghue, P.; Meloni, B.P.; O’Brien, P.A.; Thompson, R.C. Molecular characterization of Cryptosporidium isolates from humans and other animals using random amplified polymorphic DNA analysis. Am. J. Trop. Med. Hyg. 1995, 52, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Helmy, Y.A.; Von Samson-Himmelstjerna, G.; Nockler, K.; Zessin, K.H. Frequencies and spatial distributions of Cryptosporidium in livestock animals and children in the Ismailia province of Egypt. Epidemiol. Infect. 2015, 143, 1208–1218. [Google Scholar] [CrossRef] [Green Version]
- Ghaffari, S.; Kalantari, N.; Hart, C.A. A Multi-Locus Study for Detection of Cryptosporidium Species Isolated from Calves Population, Liverpool; UK. Int. J. Mol. Cell Med. 2014, 3, 35–42. [Google Scholar]
- Morrison, L.J.; Mallon, M.E.; Smith, H.V.; MacLeod, A.; Xiao, L.; Tait, A. The population structure of the Cryptosporidium parvum population in Scotland: A complex picture. Infect. Gen. Evol. 2008, 8, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Fayer, R. Molecular characterisation of species and genotypes of Cryptosporidium and Giardia and assessment of zoonotic transmission. Int. J. Parasitol. 2008, 38, 1239–1255. [Google Scholar] [CrossRef]
- Cevallos, A.M.; Zhang, X.; Waldor, M.K.; Jaison, S.; Zhou, X.; Tzipori, S.; Neutra, M.R.; Ward, H.D. Molecular cloning and expression of a gene encoding Cryptosporidium parvum glycoproteins gp40 and gp15. Infect. Immun. 2000, 68, 4108–4116. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Xiao, L.; Cama, V.A.; Ortega, Y.; Gilman, R.H.; Guo, M.; Feng, Y. Genetic recombination and Cryptosporidium hominis virulent subtype IbA10G2. Emerg. Infect. Dis. 2013, 19, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Fawzy, M.; Helmy, Y.A. The one health approach is necessary for the control of Rift Valley fever infections in Egypt: A comprehensive review. Viruses 2019, 11, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmy, Y.A.; Fawzy, M.; Elaswad, A.; Sobieh, A.; Kenney, S.P.; Shehata, A.A. The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control. J. Clin. Med. 2020, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- Innes, E.A.; Chalmers, R.M.; Wells, B.; Pawlowic, M.C. A one health approach to tackle cryptosporidiosis. Trends Parasitol. 2020, 36, 290–303. [Google Scholar] [CrossRef] [Green Version]
- King, B.J.; Keegan, A.R.; Monis, P.T.; Saint, C.P. Environmental temperature controls Cryptosporidium oocyst metabolic rate and associated retention of infectivity. App. Environ. Microbiol. 2005, 71, 3848–3857. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, M.; Trout, J.M.; Higgins, J.; Dorsch, M.; Veal, D.; Fayer, R. Comparison of tests for viable and infectious Cryptosporidium parvum oocysts. Parasitol. Res. 2003, 89, 1–5. [Google Scholar] [CrossRef]
- Fayer, R. Effect of high temperature on infectivity of Cryptosporidium parvum oocysts in water. Appl. Environ. Microbiol. 1994, 60, 2732–2735. [Google Scholar] [CrossRef] [Green Version]
- Sivaganesan, M.; Sivaganesan, S. Effect of lot variability on ultraviolet radiation inactivation kinetics of Cryptosporidium parvum oocysts. Environ. Sci. Technol. 2005, 39, 4166–4171. [Google Scholar] [CrossRef]
- Stott, R.; May, E.; Ramirez, E.; Warren, A. Predation of Cryptosporidium oocysts by protozoa and rotifers: Implications for water quality and public health. Water Sci. Technol. 2003, 47, 77–83. [Google Scholar] [CrossRef]
- Fayer, R.; Trout, J.M.; Walsh, E.; Cole, R. Rotifers Ingest Oocysts of Cryptosporidium parvum. J. Eukar. Microbiol. 2000, 47, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Mead, J.R. Cryptosporidiosis and the challenges of chemotherapy. Drug Resis. Updates 2002, 5, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Wendt, S.; Lübbert, C.; Trawinski, H. Current pharmacotherapy of cryptosporidiosis: An update of the state-of-the-art. Expert Opin. Pharmacother. 2021, 22, 2337–2342. [Google Scholar] [CrossRef] [PubMed]
- Van Voorhis, W.C.; Hulverson, M.A.; Choi, R.; Huang, W.; Arnold, S.L.M.; Schaefer, D.A.; Betzer, D.P.; Vidadala, R.S.R.; Lee, S.; Whitman, G.R.; et al. One health therapeutics: Target-Based drug development for cryptosporidiosis and other apicomplexa diseases. Vet. Parasitol. 2021, 289, 109336. [Google Scholar] [CrossRef]
- Cabada, M.M.; White, A.C., Jr. Treatment of cryptosporidiosis: Do we know what we think we know? Curr. Opin. Infect. Dis. 2010, 23, 494–499. [Google Scholar] [CrossRef]
- Johnson, A.M.; Linden, K.; Ciociola, K.M.; De Leon, R.; Widmer, G.; Rochelle, P.A. UV inactivation of Cryptosporidium hominis as measured in cell culture. Appl. Environ. Microbiol. 2005, 71, 2800–2802. [Google Scholar] [CrossRef] [Green Version]
- Rochelle, P.A.; Fallar, D.; Marshall, M.M.; Montelone, B.A.; Upton, S.J.; Woods, K. Irreversible UV inactivation of Cryptosporidium spp. despite the presence of UV repair genes. J. Euk. Microbiol. 2004, 51, 553–562. [Google Scholar] [CrossRef]
- Innes, E.A.; Bartley, P.M.; Rocchi, M.; Benavidas-Silvan, J.; Burrells, A.; Hotchkiss, E.; Chianini, F.; Canton, G.; Katzer, F. Developing vaccines to control protozoan parasites in ruminants: Dead or alive? Vet. Parasitol. 2011, 180, 155–163. [Google Scholar] [CrossRef]
- Mead, J.R. Prospects for immunotherapy and vaccines against Cryptosporidium. Human Vac. Immun. 2014, 10, 1505–1513. [Google Scholar] [CrossRef] [Green Version]
- Ryan, U.; Zahedi, A.; Paparini, A. Cryptosporidium in humans and animals–A one health approach to prophylaxis. Parasit. Immunol. 2016, 38, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Ludington, J.G.; Ward, H.D. Systemic and mucosal immune responses to cryptosporidium–Vaccine development. Curr. Trop. Med. Rep. 2015, 2, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borad, A.; Ward, H. Human immune responses in cryptosporidiosis. Future Microbiol. 2010, 5, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Ming, Z.; Zhou, R.; Chen, X.-M. Regulation of host epithelial responses to Cryptosporidium infection by microRNAs. Parasit. Immunol. 2017, 39, e12408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, B.; Huston, C.; Wagner, D.; Noel, F.; Rouzier, P.; Pape, J.; Bois, G.; Larsson, C.; Alston, W.; Tenney, K. Serum mannose-binding lectin deficiency is associated with cryptosporidiosis in young Haitian children. Clin. Infect. Dis. 2006, 43, 289–294. [Google Scholar] [CrossRef] [Green Version]
- Kelly, P.; Jack, D.L.; Naeem, A.; Mandanda, B.; Pollok, R.C.; Klein, N.J.; Turner, M.W.; Farthing, M.J. Mannose-binding lectin is a component of innate mucosal defense against Cryptosporidium parvum in AIDS. Gastroenterology 2000, 119, 1236–1242. [Google Scholar] [CrossRef]
- Carmolli, M.; Duggal, P.; Haque, R.; Lindow, J.; Mondal, D.; Petri, W.A., Jr.; Mourningstar, P.; Larsson, C.J.; Sreenivasan, M.; Khan, S. Deficient serum mannose-binding lectin levels and MBL2 polymorphisms increase the risk of single and recurrent Cryptosporidium infections in young children. J. Infect. Dis. 2009, 200, 1540–1547. [Google Scholar] [CrossRef] [Green Version]
- Lucio-Forster, A.; Griffiths, J.K.; Cama, V.A.; Xiao, L.; Bowman, D.D. Minimal zoonotic risk of cryptosporidiosis from pet dogs and cats. Trends Parasitol. 2010, 26, 174–179. [Google Scholar] [CrossRef]
- Sagodira, S.; Buzoni-Gatel, D.; Iochmann, S.; Naciri, M.; Bout, D. Protection of kids against Cryptosporidium parvum infection after immunization of dams with CP15-DNA. Vaccine 1999, 17, 2346–2355. [Google Scholar] [CrossRef]
- Roche, J.K.; Rojo, A.L.; Costa, L.B.; Smeltz, R.; Manque, P.; Woehlbier, U.; Bartelt, L.; Galen, J.; Buck, G.; Guerrant, R.L. Intranasal vaccination in mice with an attenuated Salmonella enterica Serovar 908htr A expressing Cp15 of Cryptosporidium: Impact of malnutrition with preservation of cytokine secretion. Vaccine 2013, 31, 912–918. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Zai, D.; Zhang, D.; Wei, Q.; Han, G.; Gao, H.; Huang, B. Divalent Cp15–23 vaccine enhances immune responses and protection against Cryptosporidium parvum infection. Parasit. Immunol. 2010, 32, 335–344. [Google Scholar] [CrossRef]
- Riggs, M.W. Recent advances in cryptosporidiosis: The immune response. Microb. Inf. 2002, 4, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Frost, F.J.; Tollestrup, K.; Craun, G.F.; Fairley, C.K.; Sinclair, M.I.; Kunde, T.R. Protective immunity associated with a strong serological response to a Cryptosporidium-specific antigen group, in HIV-infected individuals. J. Inf. Dis. 2005, 192, 618–621. [Google Scholar] [CrossRef] [PubMed]

| Continents | Animal Species | Diagnostic Test * | Prevalence Range |
|---|---|---|---|
| South America | Buffalo | CM, PCR | 9.4–48.2% |
| Cattle | CM, ICT, PCR | 3.0–56.1% | |
| Goat | CM | 4.8–100% | |
| Sheep | CM, PCR | 0.0–25.0% | |
| Pig | CM, PCR | 0.0–2.2% | |
| Horse | CM | 0.0–100% | |
| Calves | CM, ELISA, PCR | 84.2% | |
| North America | Cattle | CM, IFA, PCR | 1.1–78.0% |
| Goat | CM | 20.0–72.5% | |
| Sheep | CM, IFA, PCR | 20.0–77.4% | |
| Pig | CM, IFA | 2.8–19.6% | |
| Horse | CM, IFA, PCR | 0.0–17.0% | |
| Africa | Buffalo | CM, PCR | 1.3–52.0% |
| Cattle | CM, ELISA, PCR | 0.5–86.7% | |
| Goat | CM, ELISA | 0.0–76.5% | |
| Sheep | CM, ELISA, PCR | 1.3–41.8% | |
| Pig | CM, ELISA, IFA, PCR | 13.9–44.9% | |
| Horse | CM, PCR | 0.0–2.9% | |
| Asia | Buffalo | CM, ICT, PCR | 3.6–50.0% |
| Cattle | CM, ICT, IFA, PCR | 1.5–93.0% | |
| Goat | CM, ICT, IFA | 0.0–42.9% | |
| Sheep | CM, ELISA, ICT, PCR | 1.8–66.6% | |
| Pig | CM, IFA, PCR | 0.4–55.8% | |
| Horse | CM, PCR | 2.7–37.0% | |
| Europe | Buffalo | ELISA | 14.7% |
| Cattle | CM, ELISA, ICT, IFA, PCR, QLAT | 0.0–71.7% | |
| Goat | CM, ELISA, IFA | 0.0–93.0% | |
| Sheep | CM, IFA, ELISA | 1.4–100% | |
| Pig | CM, IFA, PCR | 0.1–40.9% | |
| Horse | CM, ELISA, IFA, PCR | 3.4–25.0% | |
| Australia | Buffalo | PCR | 13.1–30.0% |
| Cattle | CM, IFA, PCR | 3.6–73.5% | |
| Goat | PCR | 4.4% | |
| Sheep | PCR | 2.2–81.3% | |
| Pig | CM, PCR | 0.3–22.1% |
| Country | Species/Genotype | Host | Prevalence | Reference |
|---|---|---|---|---|
| Brazil | C. meleagridis, C. baileyi | Chicken, turkey, quail | 14.8% | [111] |
| Brazil | C. baileyi, C. parvum, C. meleagridis | Chickens | 12.6% | [112] |
| China | C. baileyi | Chickens | 2.4% | [113] |
| Iraq | C. baileyi, C. parvum, C.galli, C. meliagredis | Broilers, layers | 50% | [114] |
| Iraq | C. parvum and C. baileyi | Wild pigeons | 6.0% | [115] |
| Iran | C. parvum and C. baileyi | Broilers | 8.0% | [116] |
| China | C. parvum and C. baileyi | Wild birds | 8.9% | [117] |
| Bangladesh | C. baileyi, C. meleagridis, C. parvum | Layers, broilers, pigeons | 15.7% | [118] |
| China | C. avium, C. baileyi, C. galli, C. meleagridis | Chickens | 13.7% | [119] |
| Germany | C. parvum, C. baileyi | Turkey, broilers, layers | 7.0% | [58] |
| Spain | C. meleagridis, C. parvum | Wild birds | 8.3% | [120] |
| China | C. baileyi, C. meleagridis | Pigeons | 0.8% | [121] |
| Czech Republic | C. baileyi, C. meleagridis | Red-legged partridge | 22% | [122] |
| China | Avian genotype II, C. baileyi, C. meleagridis | Chickens | 9.9% | [123] |
| China | C. baileyi, C. muris | Ostrich | 10.2% | [123] |
| Vietnam | Avian genotype II | Ostrich | 23.7% | [124] |
| Algeria | C. baileyi, C. meleagridis | Broilers | 9–69.0% | [125] |
| Algeria | C. meleagridis | Turkey | 43.9% | [126] |
| China | C. baileyi, C. meleagridis | Japanese quail | 13.1% | [127] |
| China | C. galli, C. meleagridis, C. baileyi, C. parvum, Avian genotypes I, II, III, V | Pet birds | 8.1% | [128] |
| Brazil | C. baileyi, Avian genotype II, C. galli | Wild birds | 6.6% | [129] |
| China | C. baileyi | Ostrich | 11.7% | [130] |
| China | C. baileyi | Pekin ducks | 16.6% | [131] |
| China | C. baileyi, C. meleagridis | Chickens | 8.9% | [131] |
| USA | C. parvum | Turkey | 6.3% | [132] |
| Brazil | C. baileyi, Avian genotypes I, II, III, C. galli, C. meleagridis, C. parvum | Captive birds | 4.9% | [133] |
| Australia | Avian genotypes I, II, III, C. andersoni, C. baileyi, C. galli, C. muris | Several avian species | 6.3% | [134] |
| Country | Year | No. of Cases | Species/Subtype | Source | No. of Outbreaks | References |
|---|---|---|---|---|---|---|
| USA | 2017 | 41 | C. hominis IfA12G1 and IaA15R3 | Swimming pool | 3 | [137] |
| 2016 | 1373 | C. hominis IbA10G2 and IfA12G1, and C. parvum IIaA17G1R1 and IIaA15G2R1 | Swimming pool and water park | 16 | [137] | |
| 2016 | 10 | C. parvum IIaA15G2R1 and IIaA18G3R1 | Raw cow milk | 2 | [137,138] | |
| 2015 | 55 | C. hominis IfA12G1 | Swimming pool | 1 | [139] | |
| 2015 | 103 | C. parvum IIaA17G2R2 | Raw milk | 1 | [137] | |
| 2014 | 68 | C. hominis IdA17and IfA12G1 | Swimming pool, water slide, and fountain | 4 | [137] | |
| 2014 | 11 | C. parvum IIaA16G3R1 | Unpasteurized goat milk | 1 | [140] | |
| 2013 | 67 | C. hominis IaA28R4 and IfA12G1, and C. parvum (unknown subtype) | Swimming pool, lake, water park, and fountain | 6 | [137] | |
| 2013 | 172 | C. parvum IIaA15G2R1 and C. parvum (unknown subtype) | Drinking water | 3 | [137] | |
| 2013 | 21 | C. parvum IIaA17G2R1 and C. hominis (unknown subtype) | Unknown | 3 | [137] | |
| 2012 | 182 | C. hominis IbA10G2, and C. parvum IIaA16G3R1, IIaA15G2R1, and IIaA16G2R2 | Lake, fountain, water park, and swimming pool | 9 | [137] | |
| 2011 | 44 | C. hominis IaA15R3 and IaA28R4 | Water park and swimming pool | 2 | [137] | |
| 2010 | 162 | C. hominis IaA24R4, IaA28R4, IbA10G2 and IdA15G1 | Splashpad, lake, water park, and swimming pool | 4 | [137] | |
| UK | 2017 | 43 | C. hominis IbA10G2 and IbA12G3 | Swimming pool | 2 | [141] |
| 2016 | 111 | C. hominis IbA10G2 and IdA16, and C. parvum (unknown subtype) | Swimming pool | 10 | [141] | |
| 2015 | 83 | C. hominis IbA10G2 and IaA14R3, and C. parvum IIaA15G2R1 and IIaA26G1R1 | Swimming pool and hydrotherapy pool | 11 | [141] | |
| 2015 | 424 | C. parvum IIdA24G1 | Salad | 1 | [135] | |
| 2014 | 109 | C. hominis IaA14R3, IaA20R3, IbA10G2 and IdA25, and C. parvum IIaA15G2R1 and IIdA17G1 | Swimming pool and hydrotherapy pool | 11 | [141] | |
| 2014 | 12 | C. parvum IIaA15G2R1 | Drinking water | 1 | [141] | |
| 2013 | 94 | C. hominis IbA10G2 and IA14R3 | Swimming pool and paddling pool | 5 | [141] | |
| 2013 | 23 | C. hominis IbA10G2 and IdA18 | Public drinking water supply | 1 | [141] | |
| 2013 | 11 | C. parvum IIaA15G1R1 | Unpasteurization dairy milk | [141] | ||
| 2012 | 176 | C. hominis IbA10G2 and C. hominis (unknown subtype) | Swimming pool and hydrotherapy pool | 10 | [141] | |
| 2012 | 648 | C. parvum IIaA15G2R1 | Pre-cut mixed salad leaves | 1 | [142] | |
| 2011 | 21 | C. hominis IbA10G2 and C. hominis (unknown subtype) | Swimming pool | 1 | [141] | |
| 2010 | 78 | C. hominis (unknown subtype) | Swimming pool | 2 | [143,144] | |
| Sweden | 2019 | 122 | C. parvum IIdA22G1c | Spinach in vegetable juice | 1 | [145] |
| 2011 | 872 + 730 | C. hominis | Public drinking water source | 2 | [146] | |
| 2010 | 27,000 | C. hominis IbA10G2 | Public drinking water source | 1 | [147] | |
| 2010 | 16 + 89 | C. parvum IIdA20G1e and C. parvum IIdA24G1 | Salad garnish on chanterelle sauce | 2 | [148] | |
| French Guiana | 2014 | 12 | C. hominis IbA9G2, IbA10G2, IbA15G1 | Playing and bathing in a river | 1 | [149] |
| Germany | 2013 | 167 | C. hominis IbA9G2 | Playing and bathing in a river | 1 | [150] |
| Ireland | 2012 | 12 | C. parvum IIaA20G3R1 | Public drinking water supply | 1 | [151] |
| Norway | 2018 | 6 | C. parvum IIaA14G1R1 | Apple juice | 1 | [152] |
| 2012 | 145 | C. parvum IIaA19G1R1 | Goat kids and lambs | 1 | [153] | |
| Finland | 2012 | >250 | C. parvum IIdA17G1 | Salad | 5 | [154] |
| South Korea | 2012 | 126 | C. parvum (unknown subtype) | Tap water from the underground water tank | 1 | [155] |
| Australia | 2012 | 18 | C. hominis IbA10G2 | Swimming pool | 1 | [156] |
| Canada | 2010 | 12 | C. hominis (unknown subtype) | Recreational water park | 1 | [3] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helmy, Y.A.; Hafez, H.M. Cryptosporidiosis: From Prevention to Treatment, a Narrative Review. Microorganisms 2022, 10, 2456. https://doi.org/10.3390/microorganisms10122456
Helmy YA, Hafez HM. Cryptosporidiosis: From Prevention to Treatment, a Narrative Review. Microorganisms. 2022; 10(12):2456. https://doi.org/10.3390/microorganisms10122456
Chicago/Turabian StyleHelmy, Yosra A., and Hafez M. Hafez. 2022. "Cryptosporidiosis: From Prevention to Treatment, a Narrative Review" Microorganisms 10, no. 12: 2456. https://doi.org/10.3390/microorganisms10122456