
Impaired VEGF-A-Mediated Neurovascular Crosstalk Induced by SARS-CoV-2 Spike Protein: A Potential Hypothesis Explaining Long COVID-19 Symptoms and COVID-19 Vaccine Side Effects?


Abstract
:1. Introduction
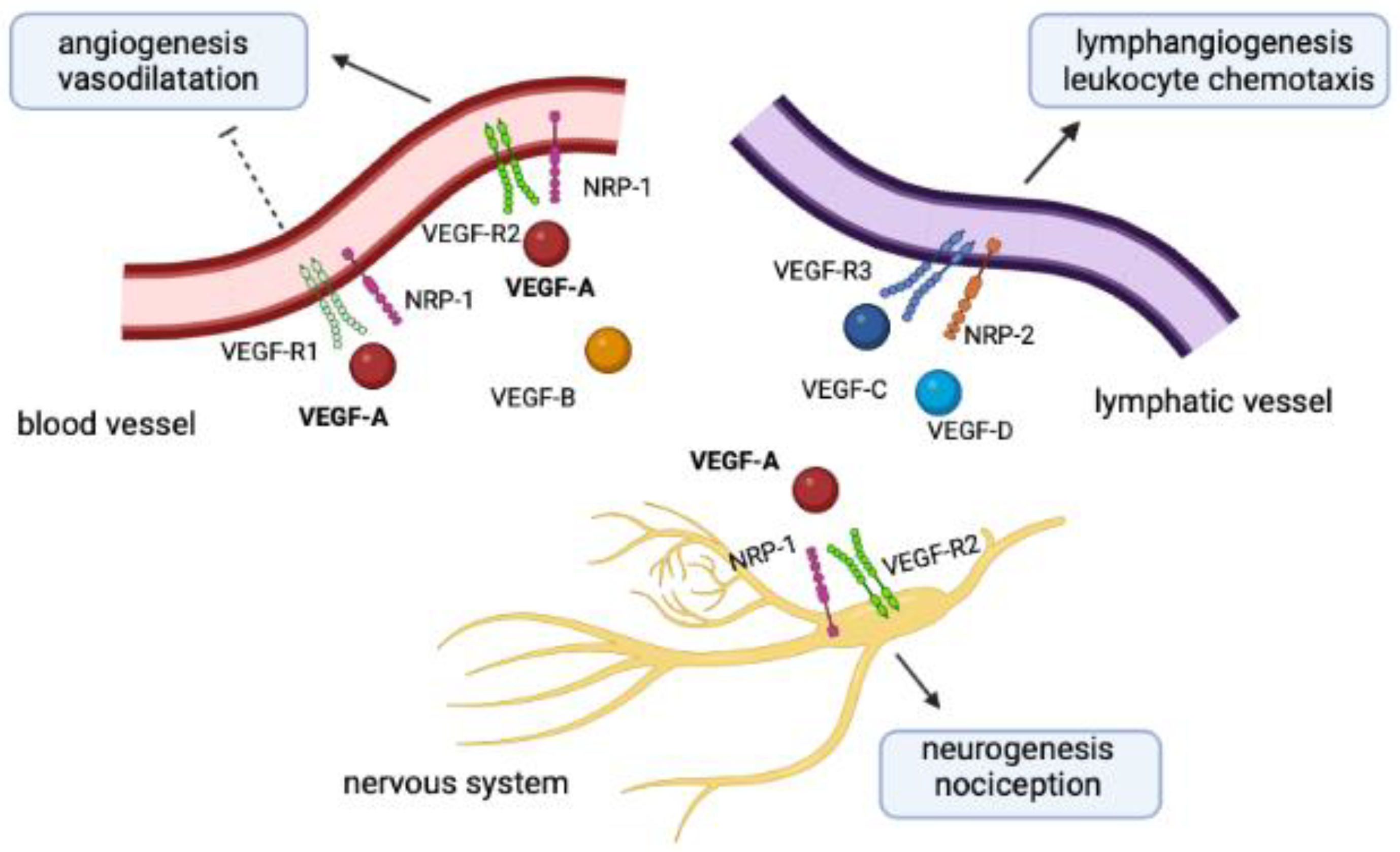
2. The NRP-1/VEGF-A Complex in Health and Disease
2.1. The Family of NRPs
2.2. The Family of VEGFs
2.3. Physiopathological Crosstalk between NRPs and VEGFs
3. The NRP-1/VEGF-A Pathway during Human Viral Infections
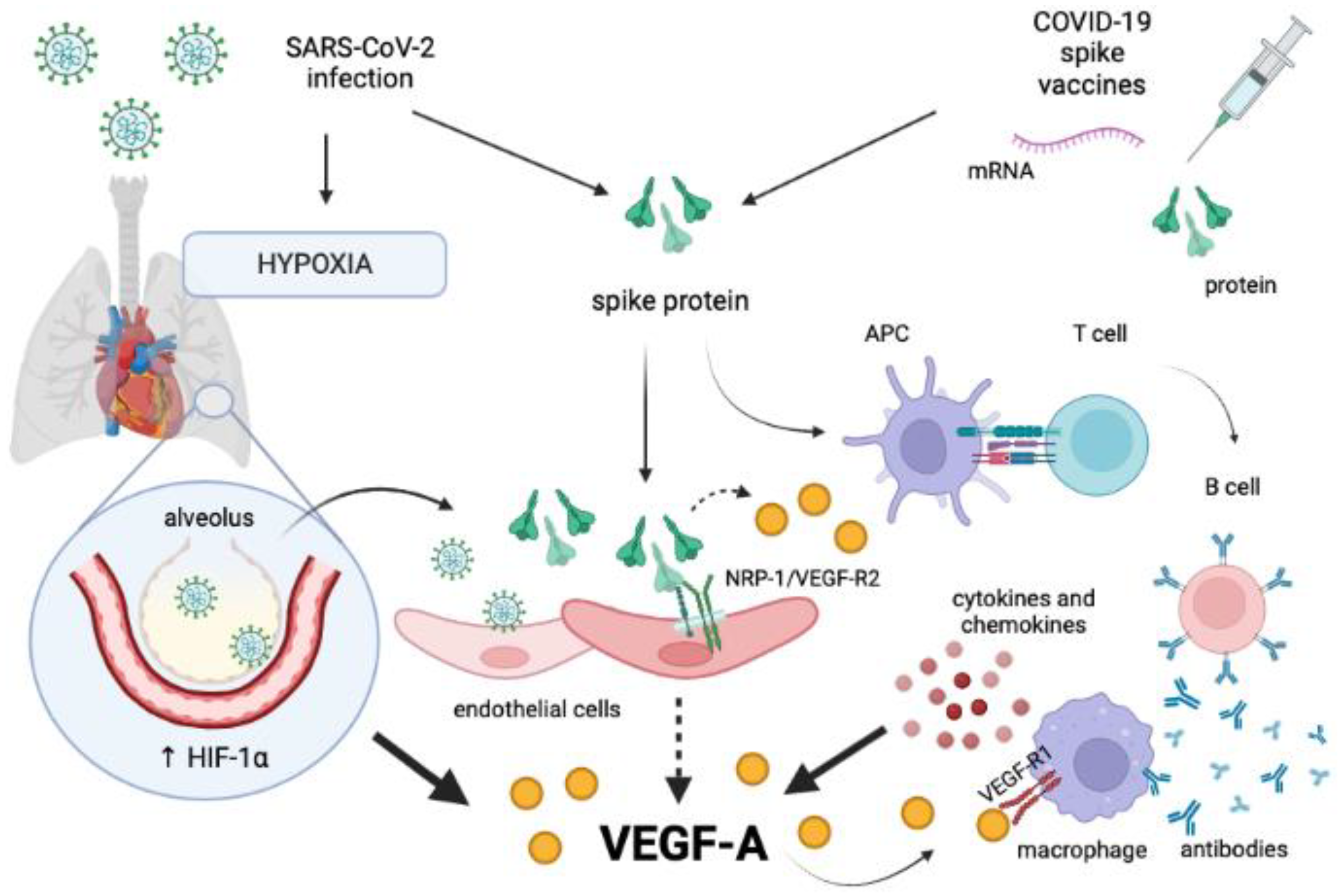
4. Disruption of the NRP-1/VEGF-A Pathway by SARS-CoV-2 in Acute COVID-19
5. Disruption of the NRP-1/VEGF-A Pathway by SARS-CoV-2 in Long COVID-19
6. Disruption of the NRP-1/VEGF-A Pathway by COVID-19 Vaccines
7. SARS-CoV-2 S Protein Impairing the NRP-1/VEGF-A Pathway: A Potential Alternative Hypothesis Unifying Multiple Scenarios?
8. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization Coronavirus Disease (COVID-19) Pandemic. Available online: https://www.who.int/europe/emergencies/situations/COVID-19 (accessed on 7 November 2022).
- Li, X.; Wang, L.; Yan, S.; Yang, F.; Xiang, L.; Zhu, J.; Shen, B.; Gong, Z. Clinical Characteristics of 25 Death Cases with COVID-19: A Retrospective Review of Medical Records in a Single Medical Center, Wuhan, China. Int. J. Infect. Dis. 2020, 94, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical Course and Outcomes of Critically Ill Patients with SARS-CoV-2 Pneumonia in Wuhan, China: A Single-Centered, Retrospective, Observational Study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S.W. Structural and Functional Properties of SARS-CoV-2 Spike Protein: Potential Antivirus Drug Development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Trougakos, I.P.; Stamatelopoulos, K.; Terpos, E.; Tsitsilonis, O.E.; Aivalioti, E.; Paraskevis, D.; Kastritis, E.; Pavlakis, G.N.; Dimopoulos, M.A. Insights to SARS-CoV-2 Life Cycle, Pathophysiology, and Rationalized Treatments That Target COVID-19 Clinical Complications. J. Biomed. Sci. 2021, 28, 9. [Google Scholar] [CrossRef] [PubMed]
- Fenizia, C.; Galbiati, S.; Vanetti, C.; Vago, R.; Clerici, M.; Tacchetti, C.; Daniele, T. SARS-CoV-2 Entry: At the Crossroads of CD147 and ACE2. Cells 2021, 10, 1434. [Google Scholar] [CrossRef] [PubMed]
- Jobe, A.; Vijayan, R. Neuropilins: C-End Rule Peptides and Their Association with Nociception and COVID-19. Comput. Struct. Biotechnol. J. 2021, 19, 1889–1895. [Google Scholar] [CrossRef]
- Saleki, K.; Banazadeh, M.; Miri, N.S.; Azadmehr, A. Triangle of Cytokine Storm, Central Nervous System Involvement, and Viral Infection in COVID-19: The Role of SFasL and Neuropilin-1. Rev. Neurosci. 2022, 33, 147–160. [Google Scholar] [CrossRef]
- Pavan, M.; Bassani, D.; Sturlese, M.; Moro, S. From the Wuhan-Hu-1 Strain to the XD and XE Variants: Is Targeting the SARS-CoV-2 Spike Protein Still a Pharmaceutically Relevant Option against COVID-19? J. Enzym. Inhib. Med. Chem. 2022, 37, 1704–1714. [Google Scholar] [CrossRef]
- Kumari, M.; Lu, R.-M.; Li, M.-C.; Huang, J.-L.; Hsu, F.-F.; Ko, S.-H.; Ke, F.-Y.; Su, S.-C.; Liang, K.-H.; Yuan, J.P.-Y.; et al. A Critical Overview of Current Progress for COVID-19: Development of Vaccines, Antiviral Drugs, and Therapeutic Antibodies. J. Biomed. Sci. 2022, 29, 68. [Google Scholar] [CrossRef]
- Zappa, M.; Verdecchia, P.; Angeli, F. Knowing the New Omicron BA.2.75 Variant (‘Centaurus’): A Simulation Study. Eur. J. Intern. Med. 2022, 105, 107–108. [Google Scholar] [CrossRef]
- Consolazio, D.; Murtas, R.; Tunesi, S.; Lamberti, A.; Senatore, S.; Faccini, M.; Russo, A.G. A Comparison Between Omicron and Earlier COVID-19 Variants’ Disease Severity in the Milan Area, Italy. Front. Epidemiol. 2022, 2, 891162. [Google Scholar] [CrossRef]
- Christie, B. COVID-19: Early Studies Give Hope Omicron Is Milder than Other Variants. BMJ 2021, 375, n3144. [Google Scholar] [CrossRef] [PubMed]
- Wolter, N.; Jassat, W.; Walaza, S.; Welch, R.; Moultrie, H.; Groome, M.; Amoako, D.G.; Everatt, J.; Bhiman, J.N.; Scheepers, C.; et al. Early Assessment of the Clinical Severity of the SARS-CoV-2 Omicron Variant in South Africa: A Data Linkage Study. Lancet 2022, 399, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Iyengar, K.P.; Jain, V.K.; Vaishya, R.; Ish, P. Long COVID-19: An Emerging Pandemic in Itself. Adv. Respir. Med. 2021, 89, 234–236. [Google Scholar] [CrossRef]
- Antonelli, M.; Pujol, J.C.; Spector, T.D.; Ourselin, S.; Steves, C.J. Risk of Long COVID Associated with Delta versus Omicron Variants of SARS-CoV-2. Lancet 2022, 399, 2263–2264. [Google Scholar] [CrossRef]
- Carfì, A.; Bernabei, R.; Landi, F. Persistent Symptoms in Patients after Acute COVID-19. JAMA—J. Am. Med. Assoc. 2020, 324, 603–605. [Google Scholar] [CrossRef]
- Yong, S.J. Long COVID or Post-COVID-19 Syndrome: Putative Pathophysiology, Risk Factors, and Treatments. Infect. Dis. 2021, 53, 737–754. [Google Scholar] [CrossRef]
- EudraVigilance—European Database of Suspected Adverse Drug Reaction Reports. Available online: https://www.adrreports.eu/en/search_subst.html# (accessed on 13 October 2022).
- SeyedAlinaghi, S.; Karimi, A.; Pashaei, Z.; Afzalian, A.; Mirzapour, P.; Ghorbanzadeh, K.; Ghasemzadeh, A.; Dashti, M.; Nazarian, N.; Vahedi, F.; et al. Safety and Adverse Events Related to COVID-19 MRNA Vaccines; a Systematic Review. Arch. Acad. Emerg. Med. 2022, 10, e41. [Google Scholar] [CrossRef]
- Rabail, R.; Ahmed, W.; Ilyas, M.; Rajoka, M.S.R.; Hassoun, A.; Khalid, A.R.; Khan, M.R.; Aadil, R.M. The Side Effects and Adverse Clinical Cases Reported after COVID-19 Immunization. Vaccines 2022, 10, 488. [Google Scholar] [CrossRef]
- Khokhar, F.; Khan, A.; Hussain, Z.; Yu, J. Small Fiber Neuropathy Associated with the Moderna SARS-CoV-2 Vaccine. Cureus 2022, 14, e25969. [Google Scholar] [CrossRef] [PubMed]
- Watad, A.; De Marco, G.; Mahajna, H.; Druyan, A.; Eltity, M.; Hijazi, N.; Haddad, A.; Elias, M.; Zisman, D.; Naffaa, M.E.; et al. Immune-Mediated Disease Flares or New-Onset Disease in 27 Subjects Following Mrna/Dna Sars-Cov-2 Vaccination. Vaccines 2021, 9, 435. [Google Scholar] [CrossRef] [PubMed]
- Schelke, M.W.; Barcavage, S.; Lampshire, E.; Brannagan, T.H. Post–COVID-19 vaccine small-fiber neuropathy and tinnitus treated with plasma exchange. Muscle Nerve 2022, 66, E21–E23. [Google Scholar] [CrossRef]
- Abbott, M.G.; Allawi, Z.; Hofer, M.; Ansorge, O.; Brady, S.; Fadic, R.; Torres, G.; Knight, R.; Calvo, M.; Bennett, D.L.H.; et al. Acute small fiber neuropathy after Oxford-AstraZeneca ChAdOx1-S vaccination: A report of three cases and review of the literature. J. Peripher. Nerv. Syst. 2022, 27, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Moutal, A.; Martin, L.F.; Boinon, L.; Gomez, K.; Ran, D.; Zhou, Y.; Stratton, H.J.; Cai, S.; Luo, S.; Gonzalez, K.B.; et al. SARS-CoV-2 Spike Protein Co-Opts VEGF-A/Neuropilin-1 Receptor Signaling to Induce Analgesia. Pain 2021, 162, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Perez-Miller, S.; Patek, M.; Moutal, A.; Duran, P.; Cabel, C.R.; Thorne, C.A.; Campos, S.K.; Khanna, R. Novel Compounds Targeting Neuropilin Receptor 1 with Potential to Interfere with SARS-CoV-2 Virus Entry. ACS Chem. Neurosci. 2021, 12, 1299–1312. [Google Scholar] [CrossRef] [PubMed]
- Gudowska-Sawczuk, M.; Mroczko, B. The Role of Neuropilin-1 (Nrp-1) in Sars-Cov-2 Infection: Review. J. Clin. Med. 2021, 10, 2772. [Google Scholar] [CrossRef] [PubMed]
- Angeli, F.; Reboldi, G.; Trapasso, M.; Zappa, M.; Spanevello, A.; Verdecchia, P. COVID-19, Vaccines and Deficiency of ACE2 and Other Angiotensinases. Closing the Loop on the “Spike Effect”. Eur. J. Intern. Med. 2022, 103, 23–28. [Google Scholar] [CrossRef]
- Angeli, F.; Spanevello, A.; Reboldi, G.; Visca, D.; Verdecchia, P. SARS-CoV-2 Vaccines: Lights and Shadows. Eur. J. Intern. Med. 2021, 88, 1–8. [Google Scholar] [CrossRef]
- Ramos, S.G.; da Cruz Rattis, B.A.; Ottaviani, G.; Celes, M.R.N.; Dias, E.P. ACE2 Down-Regulation May Act as a Transient Molecular Disease Causing RAAS Dysregulation and Tissue Damage in the Microcirculatory Environment Among COVID-19 Patients. Am. J. Pathol. 2021, 191, 1154–1164. [Google Scholar] [CrossRef]
- Sfera, A.; Osorio, C.; Jafri, N.; Diaz, E.L.; Campo Maldonado, J.E. Intoxication With Endogenous Angiotensin II: A COVID-19 Hypothesis. Front. Immunol. 2020, 11, 1472. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Hu, R.; Zhang, C.; Ren, W.; Yu, A.; Zhou, X. Elevation of Plasma Angiotensin II Level Is a Potential Pathogenesis for the Critically Ill COVID-19 Patients. Crit. Care 2020, 24, 290. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, Y.; Zhang, J.; Xing, B.; Xuan, W.; Wang, H.; Huang, H.; Yang, J.; Tang, J. NRP-2 in Tumor Lymphangiogenesis and Lymphatic Metastasis. Cancer Lett. 2018, 418, 176–184. [Google Scholar] [CrossRef]
- Roy, S.; Bag, A.K.; Singh, R.K.; Talmadge, J.E.; Batra, S.K.; Datta, K. Multifaceted Role of Neuropilins in the Immune System: Potential Targets for Immunotherapy. Front. Immunol. 2017, 8, 1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.D.; Zhong, L.P.; He, J.; Zhao, Y.X. Targeting Neuropilin-1 Interactions Is a Promising Anti-Tumor Strategy. Chin. Med. J. 2021, 134, 508–517. [Google Scholar] [CrossRef]
- Graziani, G.; Lacal, P.M. Neuropilin-1 as Therapeutic Target for Malignant Melanoma. Front. Oncol. 2015, 5, 125. [Google Scholar] [CrossRef] [Green Version]
- Van Acker, N.; Ragé, M.; Vermeirsch, H.; Schrijvers, D.; Nuydens, R.; Byttebier, G.; Timmers, M.; De Schepper, S.; Streffer, J.; Andries, L.; et al. NRP-1 Receptor Expression Mismatch in Skin of Subjects with Experimental and Diabetic Small Fiber Neuropathy. PLoS ONE 2016, 11, e0161441. [Google Scholar] [CrossRef] [Green Version]
- Harvey, N.L. Lymphatic Vascular Development. In Heart Development and Regeneration; Rosenthal, N., Harvey, R.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 543–565. ISBN 9780123813329. [Google Scholar]
- Schellenburg, S.; Schulz, A.; Poitz, D.M.; Muders, M.H. Role of Neuropilin-2 in the Immune System. Mol. Immunol. 2017, 90, 239–244. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhu, X.; Cui, H.; Shi, J.; Yuan, G.; Shi, S.; Hu, Y. The Role of the VEGF Family in Coronary Heart Disease. Front. Cardiovasc. Med. 2021, 8, 738325. [Google Scholar] [CrossRef]
- Woolard, J.; Bevan, H.S.; Harper, S.J.; Bates, D.O. Molecular Diversity of VEGF-A as a Regulator of Its Biological Activity. Microcirculation 2009, 16, 572–592. [Google Scholar] [CrossRef]
- Wallensten, J.; Mobarrez, F.; Åsberg, M.; Borg, K.; Beser, A.; Wilczek, A.; Nager, A. Isoforms of Soluble Vascular Endothelial Growth Factor in Stress-Related Mental Disorders: A Cross-Sectional Study. Sci. Rep. 2021, 11, 16693. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chen, H.; Lu, L.; Wang, L.; Zhang, X.; Guo, X. New Insights into the Role of Co-Receptor Neuropilins in Tumour Angiogenesis and Lymphangiogenesis and Targeted Therapy Strategies. J. Drug Target. 2021, 29, 155–167. [Google Scholar] [CrossRef] [PubMed]
- King, C.; Wirth, D.; Workman, S.; Hristova, K. Interactions between NRP1 and VEGFR2 Molecules in the Plasma Membrane. Biochim. Biophys. Acta-Biomembr. 2018, 1860, 2118–2125. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, M. Vascular Endothelial Growth Factor Receptor-1 (VEGFR-1/Flt-1): A Dual Regulator for Angiogenesis. Angiogenesis 2006, 9, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Colotti, G.; Failla, C.M.; Lacal, P.M.; Ungarelli, M.; Ruffini, F.; Di Micco, P.; Orecchia, A.; Morea, V. Neuropilin-1 Is Required for Endothelial Cell Adhesion to Soluble Vascular Endothelial Growth Factor Receptor 1. FEBS J. 2022, 289, 183–198. [Google Scholar] [CrossRef]
- Lampropoulou, D.I.; Bala, V.M.; Zerva, E.; Pliakou, E.; Filippou, D.; Gazouli, M.; Aravantinos, G. The Potential Role of the Combined PARP-1 and VEGF Inhibition in Severe SARS-CoV-2 (COVID-19) Infection. J. Infect. Dev. Ctries 2022, 16, 101–111. [Google Scholar] [CrossRef]
- Nowacka, M.M.; Obuchowicz, E. Vascular Endothelial Growth Factor (VEGF) and Its Role in the Central Nervous System: A New Element in the Neurotrophic Hypothesis of Antidepressant Drug Action. Neuropeptides 2012, 46, 1–10. [Google Scholar] [CrossRef]
- Hulse, R.P. Role of VEGF-A in Chronic Pain. Oncotarget 2017, 8, 10775–10776. [Google Scholar] [CrossRef]
- Mukouyama, Y.S.; Gerber, H.P.; Ferrara, N.; Gu, C.; Anderson, D.J. Peripheral Nerve-Derived VEGF Promotes Arterial Differentiation via Neuropilin 1-Mediated Positive Feedback. Development 2005, 132, 941–952. [Google Scholar] [CrossRef] [Green Version]
- Schuch, G.; Machluf, M.; Bartsch, G.; Nomi, M.; Richard, H.; Atala, A.; Soker, S. In Vivo Administration of Vascular Endothelial Growth Factor (VEGF) and Its Antagonist, Soluble Neuropilin-1, Predicts a Role of VEGF in the Progression of Acute Myeloid Leukemia in Vivo. Blood 2002, 100, 4622–4628. [Google Scholar] [CrossRef]
- Lu, L.; Zhang, L.; Xiao, Z.; Lu, S.; Yang, R.; Han, Z.C. Neuropilin-1 in Acute Myeloid Leukemia: Expression and Role in Proliferation and Migration of Leukemia Cells. Leuk. Lymphoma 2008, 49, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Ruffini, F.; D’Atri, S.; Lacal, P.M. Neuropilin-1 Expression Promotes Invasiveness of Melanoma Cells through Vascular Endothelial Growth Factor Receptor-2-Dependent and -Independent Mechanisms. Int. J. Oncol. 2013, 43, 297–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.W.; Lee, J.E.; Yoo, C.Y.; Ko, M.S.; Park, C.S.; Yang, S.H. NRP-1 Expression Is Strongly Associated with the Progression of Pituitary Adenomas. Oncol. Rep. 2014, 32, 1537–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.; Hou, L.; Li, J.; Shao, S.; Huang, S.; Meng, D.; Liu, L.; Feng, L.; Xia, P.; Qin, T.; et al. VEGF/NRP-1axis Promotes Progression of Breast Cancer via Enhancement of Epithelial-Mesenchymal Transition and Activation of NF-ΚB and β-Catenin. Cancer Lett. 2016, 373, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Jiang, X. Role of NRP-1 in VEGF-VEGFR2-Independent Tumorigenesis. Target. Oncol. 2016, 11, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.; Hurwitz, H.I.; Sandler, A.B.; Miles, D.; Coleman, R.L.; Deurloo, R.; Chinot, O.L. Bevacizumab (Avastin®) in Cancer Treatment: A Review of 15 Years of Clinical Experience and Future Outlook. Cancer Treat. Rev. 2020, 86, 102017. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, Y.; Yamada, S.; Thirunavukkarasu, M.; Nin, V.; Joshi, M.; Rishi, M.T.; Bhattacharya, S.; Camacho-Pereira, J.; Sharma, A.K.; et al. Cardiomyopathy and Worsened Ischemic Heart Failure in SM22-α Cre-Mediated Neuropilin-1 Null Mice: Dysregulation of PGC1α and Mitochondrial Homeostasis. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1401–1412. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, E.; Zhang, Y.; Madamsetty, V.S.; Ji, B.; Radisky, D.C.; Grande, J.P.; Misra, S.; Mukhopadhyay, D. Neuropilin-1 Maintains Dimethylarginine Dimethylaminohydrolase 1 Expression in Endothelial Cells, and Contributes to Protection from Angiotensin II-Induced Hypertension. FASEB J. 2019, 33, 494–500. [Google Scholar] [CrossRef]
- Costa, C.; Martínez-Sáez, E.; Gutiérrez-Franco, A.; Eixarch, H.; Castro, Z.; Ortega-Aznar, A.; Ramón, Y.C.S.; Montalban, X.; Espejo, C. Expression of Semaphorin 3A, Semaphorin 7A and Their Receptors in Multiple Sclerosis Lesions. Mult. Scler. 2015, 21, 1632–1643. [Google Scholar] [CrossRef]
- Solomona, B.D.; Muellera, C.; Chaeb, W.J.; Alabanzaa, L.M.; Bynoea, M.S. Neuropilin-1 Attenuates Autoreactivity in Experimental Autoimmune Encephalomyelitis. Proc. Natl. Acad. Sci. USA 2011, 108, 2040–2045. [Google Scholar] [CrossRef]
- Nissen, J.C.; Tsirka, S.E. Tuftsin-Driven Experimental Autoimmune Encephalomyelitis Recovery Requires Neuropilin-1. Glia 2016, 64, 923–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollock, C.E.; Sutherland, H.G.; Maher, B.H.; Lea, R.A.; Haupt, L.M.; Frith, A.; Anne MacGregor, E.; Griffiths, L.R. The NRP1 Migraine Risk Variant Shows Evidence of Association with Menstrual Migraine. J. Headache Pain 2018, 19, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, T.; Qazilbash, M.H. POEMS Syndrome: A Multisystem Clonal Disorder. Eur. J. Haematol. 2021, 106, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Dispenzieri, A. POEMS Syndrome: 2021 Update on Diagnosis, Risk-Stratification, and Management. Am. J. Hematol. 2021, 96, 872–888. [Google Scholar] [CrossRef]
- Arima, K.; Origuchi, T.; Tamai, M.; Iwanaga, N.; Izumi, Y.; Huang, M.; Tanaka, F.; Kamachi, M.; Aratake, K.; Nakamura, H.; et al. RS3PE Syndrome Presenting as Vascular Endothelial Growth Factor Associated Disorder. Ann. Rheum. Dis. 2005, 64, 1653–1655. [Google Scholar] [CrossRef] [Green Version]
- Smets, P.; Devauchelle-Pensec, V.; Rouzaire, P.O.; Pereira, B.; Andre, M.; Soubrier, M. Vascular Endothelial Growth Factor Levels and Rheumatic Diseases of the Elderly. Arthritis Res. Ther. 2016, 18, 283. [Google Scholar] [CrossRef] [Green Version]
- Harada, M.; Mitsuyama, K.; Yoshida, H.; Sakisaka, S.; Taniguchi, E.; Kawaguchi, T.; Ariyoshi, M.; Saiki, T.; Sakamoto, M.; Nagata, K.; et al. Vascular Endothelial Growth Factor in Patients with Rheumatoid Arthritis. Scand. J. Rheumatol. 1998, 27, 377–380. [Google Scholar] [CrossRef]
- Meliconi, R.; Pulsatelli, L.; Dolzani, P.; Boiardi, L.; Macchioni, P.; Salvarani, C.; Silvestri, T.; Frizziero, L.; Facchini, A. Vascular Endothelial Growth Factor Production in Polymyalgia Rheumatica. Arthritis Rheum. 2000, 43, 2472–2480. [Google Scholar] [CrossRef]
- Lambert, S.; Bouttier, M.; Vassy, R.; Seigneuret, M.; Petrow-Sadowski, C.; Janvier, S.; Heveker, N.; Ruscetti, F.W.; Perret, G.; Jones, K.S.; et al. HTLV-1 Uses HSPG and Neuropilin-1 for Entry by Molecular Mimicry of VEGF165. Blood 2009, 113, 5176–5185. [Google Scholar] [CrossRef] [Green Version]
- Afonso, P.V.; Ozden, S.; Cumont, M.C.; Seilhean, D.; Cartier, L.; Rezaie, P.; Mason, S.; Lambert, S.; Huerre, M.; Gessain, A.; et al. Alteration of Blood-Brain Barrier Integrity by Retroviral Infection. PLoS Pathog. 2008, 4, e1000205. [Google Scholar] [CrossRef]
- Haeberle, H.A.; Dürrstein, C.; Rosenberger, P.; Hosakote, Y.M.; Kuhlicke, J.; Kempf, V.A.J.; Garofalo, R.P.; Eitzschig, H.K. Oxygen-Independent Stabilization of Hypoxia Inducible Factor (HIF)-1 during RSV Infection. PLoS ONE 2008, 3, e3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morichi, S.; Morishita, N.; Takeshita, M.; Ishida, Y.; Oana, S.; Yamanaka, G.; Kashiwagi, Y.; Kawashima, H. Vascular Endothelial Growth Factor (VEGF) and Platelet-Derived Growth Factor (PDGF) Levels in the Cerebrospinal Fluid of Children with Influenza-Associated Encephalopathy. J. Infect. Chemother. 2017, 23, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Alkharsah, K.R. VEGF Upregulation in Viral Infections and Its Possible Therapeutic Implications. Int. J. Mol. Sci. 2018, 19, 1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.S.; Oshansky, C.M.; Guo, X.Z.J.; Ralston, J.; Wood, T.; Seeds, R.; Newbern, C.; Waite, B.; Reynolds, G.; Widdowson, M.A.; et al. Severe Influenza Is Characterized by Prolonged Immune Activation: Results from the SHIVERS Cohort Study. J. Infect. Dis. 2018, 217, 245–256. [Google Scholar] [CrossRef]
- Bautista, E.; Arcos, M.; Jimenez-Alvarez, L.; García-Sancho, M.C.; Vázquez, M.E.; Peña, E.; Higuera, A.; Ramírez, G.; Fernández-Plata, R.; Cruz-Lagunas, A.; et al. Angiogenic and Inflammatory Markers in Acute Respiratory Distress Syndrome and Renal Injury Associated to A/H1N1 Virus Infection. Exp. Mol. Pathol. 2013, 94, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Pino, M.; Kelvin, D.J.; Bermejo-Martin, J.F.; Alonso, A.; Matías, V.; Tenorio, A.; Rico, L.; Eiros, J.M.; Castrodeza, J.; Blanco-Quiros, A.; et al. Nasopharyngeal Aspirate Cytokine Levels 1 Yr after Severe Respiratory Syncytial Virus Infection. Pediatr. Allergy Immunol. 2009, 20, 791–795. [Google Scholar] [CrossRef]
- Bermejo-Martin, J.F.; Garcia-Arevalo, M.C.; De Lejarazu, R.O.; Ardura, J.; Eiros, J.M.; Alonso, A.; Matías, V.; Pino, M.; Bernardo, D.; Arranz, E.; et al. Predominance of Th2 Cytokines, CXC Chemokines and Innate Immunity Mediators at the Mucosal Level during Severe Respiratory Syncytial Virus Infection in Children. Eur. Cytokine Netw. 2007, 18, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Rahmatpanah, F.; Agrawal, S.; Jaiswal, N.; Ngyuen, H.M.; McClelland, M.; Agrawal, A. Airway Epithelial Cells Prime Plasmacytoid Dendritic Cells to Respond to Pathogens via Secretion of Growth Factors. Mucosal Immunol. 2019, 12, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Oldford, S.A.; Salsman, S.P.; Portales-Cervantes, L.; Alyazidi, R.; Anderson, R.; Haidl, I.D.; Marshall, J.S. Interferon A2 and Interferon γ Induce the Degranulation Independent Production of VEGF-A and IL-1 Receptor Antagonist and Other Mediators from Human Mast Cells. Immun. Inflamm. Dis. 2018, 6, 176–189. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.G.; Yoon, H.J.; Zhu, Z.; Link, H.; Wang, Z.; Gwaltney, J.; Landry, M.; Elias, J.A. Respiratory Syncytial Virus Stimulation of Vascular Endothelial Cell Growth Factor/Vascular Permeability Factor. Am. J. Respir. Cell Mol. Biol. 2000, 23, 662–669. [Google Scholar] [CrossRef]
- Killerby, M.E.; Biggs, H.M.; Haynes, A.; Dahl, R.M.; Mustaquim, D.; Gerber, S.I.; Watson, J.T. Human Coronavirus Circulation in the United States 2014–2017. J. Clin. Virol. 2018, 101, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Rabaan, A.A.; Al-Ahmed, S.H.; Haque, S.; Sah, R.; Tiwari, R.; Malik, Y.S.; Dhama, K.; Yatoo, M.I.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. SARS-CoV-2, SARS-CoV, and MERS-CoV: A Comparative Overview. Infez. Med. 2020, 28, 174–184. [Google Scholar] [PubMed]
- Mohammed, M.E.A. The Percentages of SARS-CoV-2 Protein Similarity and Identity with SARS-CoV and BatCoV RaTG13 Proteins Can Be Used as Indicators of Virus Origin. J. Proteins Proteom. 2021, 12, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; He, Y.; Zhou, Y.; Liu, S.; Zheng, B.J.; Jiang, S. The Spike Protein of SARS-CoV—A Target for Vaccine and Therapeutic Development. Nat. Rev. Microbiol. 2009, 7, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Hatmal, M.M.; Alshaer, W.; Al-Hatamleh, M.A.I.; Hatmal, M.; Smadi, O.; Taha, M.O.; Oweida, A.J.; Boer, J.C.; Mohamud, R.; Plebanski, M. Comprehensive Structural and Molecular Comparison of Spike Proteins of SARS-CoV-2, SARS-CoV and MERS-CoV, and Their Interactions with ACE2. Cells 2020, 9, 2638. [Google Scholar] [CrossRef] [PubMed]
- Rabaan, A.A.; Bazzi, A.M.; Al-Ahmed, S.H.; Al-Tawfiq, J.A. Molecular Aspects of MERS-CoV. Front. Med. 2017, 11, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Verma, J.; Subbarao, N. A Comparative Study of Human Betacoronavirus Spike Proteins: Structure, Function and Therapeutics. Arch. Virol. 2021, 166, 697–714. [Google Scholar] [CrossRef]
- Millet, J.K.; Jaimes, J.A.; Whittaker, G.R. Molecular Diversity of Coronavirus Host Cell Entry Receptors. FEMS Microbiol. Rev. 2021, 45, fuaa057. [Google Scholar] [CrossRef]
- Ghosh, P.; Jayaram, S.; Patwardhan, D.; Marimuthu, S.; Lenehan, P.; Venkatakrishnan, A.; Hughes, T.; Zemmour, D.; Anand, P.; Soundararajan, V. Diversity of Coronavirus Receptors. Preprints 2021, 2021080071. [Google Scholar] [CrossRef]
- Talotta, R.; Robertson, E. Autoimmunity as the Comet Tail of COVID-19 Pandemic. World J. Clin. Cases 2020, 8, 3621–3644. [Google Scholar] [CrossRef]
- Lechien, J.R.; Chiesa-Estomba, C.M.; Place, S.; Van Laethem, Y.; Cabaraux, P.; Mat, Q.; Huet, K.; Plzak, J.; Horoi, M.; Hans, S.; et al. Clinical and Epidemiological Characteristics of 1420 European Patients with Mild-to-Moderate Coronavirus Disease 2019. J. Intern. Med. 2020, 288, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Stokes, E.K.; Zambrano, L.D.; Anderson, K.N.; Marder, E.P.; Raz, K.M.; El Burai Felix, S.; Tie, Y.; Fullerton, K.E. Coronavirus Disease 2019 Case Surveillance—United States, January 22–May 30 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Long, B.; Carius, B.M.; Chavez, S.; Liang, S.Y.; Brady, W.J.; Koyfman, A.; Gottlieb, M. Clinical Update on COVID-19 for the Emergency Clinician: Presentation and Evaluation. Am. J. Emerg. Med. 2022, 54, 46–57. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, K.Y.; Huang, Y.; Lui, K.O. Endothelial Contribution to COVID-19: An Update on Mechanisms and Therapeutic Implications. J. Mol. Cell. Cardiol. 2022, 164, 69–82. [Google Scholar] [CrossRef]
- Sulli, A.; Gotelli, E.; Bica, P.F.; Schiavetti, I.; Pizzorni, C.; Aloè, T.; Grosso, M.; Barisione, E.; Paolino, S.; Smith, V.; et al. Detailed Videocapillaroscopic Microvascular Changes Detectable in Adult COVID-19 Survivors. Microvasc. Res. 2022, 142, 104361. [Google Scholar] [CrossRef]
- Çakmak, F.; Demirbuga, A.; Demirkol, D.; Gümüş, S.; Torun, S.H.; Kayaalp, G.K.; Ömeroglu, R.E.; Somer, A.; Uysalol, M.; Yıldız, R.; et al. Nailfold Capillaroscopy: A Sensitive Method for Evaluating Microvascular Involvement in Children with SARS-CoV-2 Infection. Microvasc. Res. 2021, 138, 104196. [Google Scholar] [CrossRef]
- Natalello, G.; De Luca, G.; Gigante, L.; Campochiaro, C.; De Lorenzis, E.; Verardi, L.; Paglionico, A.; Petricca, L.; Martone, A.M.; Calvisi, S.; et al. Nailfold Capillaroscopy Findings in Patients with Coronavirus Disease 2019: Broadening the Spectrum of COVID-19 Microvascular Involvement. Microvasc. Res. 2021, 133, 104071. [Google Scholar] [CrossRef]
- Rovas, A.; Osiaevi, I.; Buscher, K.; Sackarnd, J.; Tepasse, P.R.; Fobker, M.; Kühn, J.; Braune, S.; Göbel, U.; Thölking, G.; et al. Microvascular Dysfunction in COVID-19: The MYSTIC Study. Angiogenesis 2021, 24, 145–157. [Google Scholar] [CrossRef]
- Lim, J.; Puan, K.J.; Wang, L.W.; Teng, K.W.W.; Loh, C.Y.; Tan, K.P.; Carissimo, G.; Chan, Y.H.; Poh, C.M.; Lee, C.Y.P.; et al. Data-Driven Analysis of COVID-19 Reveals Persistent Immune Abnormalities in Convalescent Severe Individuals. Front. Immunol. 2021, 12, 710217. [Google Scholar] [CrossRef]
- Medeiros, T.; Guimarães, G.M.C.; Carvalho, F.R.; Alves, L.S.; Faustino, R.; Campi-Azevedo, A.C.; Peruhype-Magalhães, V.; Teixeira-Carvalho, A.; de Souza Gomes, M.; Rodrigues do Amaral, L.; et al. Acute Kidney Injury Associated to COVID-19 Leads to a Strong Unbalance of Circulant Immune Mediators. Cytokine 2022, 157, 155974. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Meini, S.; Giani, T.; Tascini, C. Intussusceptive Angiogenesis in COVID-19: Hypothesis on the Significance and Focus on the Possible Role of FGF2. Mol. Biol. Rep. 2020, 47, 8301–8304. [Google Scholar] [CrossRef] [PubMed]
- Smadja, D.M.; Mentzer, S.J.; Fontenay, M.; Laffan, M.A.; Ackermann, M.; Helms, J.; Jonigk, D.; Chocron, R.; Pier, G.B.; Gendron, N.; et al. COVID-19 Is a Systemic Vascular Hemopathy: Insight for Mechanistic and Clinical Aspects. Angiogenesis 2021, 24, 755–788. [Google Scholar] [CrossRef]
- Mayi, B.S.; Leibowitz, J.A.; Woods, A.T.; Ammon, K.A.; Liu, A.E.; Raja, A. The Role of Neuropilin-1 in COVID-19. PLoS Pathog. 2021, 17, e1009153. [Google Scholar] [CrossRef]
- Alipoor, S.D.; Mirsaeidi, M. SARS-CoV-2 Cell Entry beyond the ACE2 Receptor. Mol. Biol. Rep. 2022, 49, 10715–10727. [Google Scholar] [CrossRef]
- Simonetti, B.; Daly, J.L.; Simón-Gracia, L.; Klein, K.; Weeratunga, S.; Antón-Plágaro, C.; Tobi, A.; Hodgson, L.; Lewis, P.A.; Heesom, K.J.; et al. ESCPE-1 Mediates Retrograde Endosomal Sorting of the SARS-CoV-2 Host Factor Neuropilin-1. Proc. Natl. Acad. Sci. USA 2022, 119, e2201980119. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Mei, H.; Sun, J.; Li, H.; Huang, Y.; Tang, Y.; Duan, L.; Liu, D.; Pang, Y.; Wang, Q.; et al. Neuropilin-1-Mediated SARS-CoV-2 Infection in Bone Marrow-Derived Macrophages Inhibits Osteoclast Differentiation. Adv. Biol. 2022, 6, 2200007. [Google Scholar] [CrossRef]
- Benedetti, F.; Silvestri, G.; Mavian, C.; Weichseldorfer, M.; Munawwar, A.; Cash, M.N.; Dulcey, M.; Vittor, A.Y.; Ciccozzi, M.; Salemi, M.; et al. Comparison of Sars-Cov-2 Receptors Expression in Primary Endothelial Cells and Retinoic Acid-Differentiated Human Neuronal Cells. Viruses 2021, 13, 2193. [Google Scholar] [CrossRef]
- Zhang, F.; Li, W.; Feng, J.; da Silva, S.R.; Ju, E.; Zhang, H.; Chang, Y.; Moore, P.S.; Guo, H.; Gao, S.J. SARS-CoV-2 Pseudovirus Infectivity and Expression of Viral Entry-Related Factors ACE2, TMPRSS2, Kim-1, and NRP-1 in Human Cells from the Respiratory, Urinary, Digestive, Reproductive, and Immune Systems. J. Med. Virol. 2021, 93, 6671–6685. [Google Scholar] [CrossRef]
- Eldrid, C.; Zloh, M.; Fotinou, C.; Yelland, T.; Yu, L.; Mota, F.; Selwood, D.L.; Djordjevic, S. VEGFA, B, C: Implications of the C-Terminal Sequence Variations for the Interaction with Neuropilins. Biomolecules 2022, 12, 372. [Google Scholar] [CrossRef]
- Li, Z.-L.; Buck, M. Neuropilin-1 Assists SARS-CoV-2 Infection by Stimulating the Separation of Spike Protein S1 and S2. Biophys. J. 2021, 120, 2828–2837. [Google Scholar] [CrossRef] [PubMed]
- Baindara, P.; Roy, D.; Mandal, S.M.; Schrum, A.G. Conservation and Enhanced Binding of SARS-CoV-2 Omicron Spike Protein to Coreceptor Neuropilin-1 Predicted by Docking Analysis. Infect. Dis. Rep. 2022, 14, 243–249. [Google Scholar] [CrossRef]
- Kong, W.; Montano, M.; Corley, M.J.; Helmy, E.; Kobayashi, H.; Kinisu, M.; Suryawanshi, R.; Luo, X.; Royer, L.A.; Roan, N.R.; et al. Neuropilin-1 Mediates SARS-CoV-2 Infection of Astrocytes in Brain Organoids, Inducing Inflammation Leading to Dysfunction and Death of Neurons. mBio 2022, e0230822. [Google Scholar] [CrossRef]
- Chaudhary, J.K.; Yadav, R.; Chaudhary, P.K.; Maurya, A.; Roshan, R.; Azam, F.; Mehta, J.; Handu, S.; Prasad, R.; Jain, N.; et al. Host Cell and SARS-CoV-2-Associated Molecular Structures and Factors as Potential Therapeutic Targets. Cells 2021, 10, 2427. [Google Scholar] [CrossRef]
- Needham, E.J.; Chou, S.H.Y.; Coles, A.J.; Menon, D.K. Neurological Implications of COVID-19 Infections. Neurocrit. Care 2020, 32, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Tangos, M.; Budde, H.; Kolijn, D.; Sieme, M.; Zhazykbayeva, S.; Lódi, M.; Herwig, M.; Gömöri, K.; Hassoun, R.; Robinson, E.L.; et al. SARS-CoV-2 Infects Human Cardiomyocytes Promoted by Inflammation and Oxidative Stress. Int. J. Cardiol. 2022, 362, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Mezache, L.; Struckman, H.L.; Greer-Short, A.; Baine, S.; Györke, S.; Radwański, P.B.; Hund, T.J.; Veeraraghavan, R. Vascular Endothelial Growth Factor Promotes Atrial Arrhythmias by Inducing Acute Intercalated Disk Remodeling. Sci. Rep. 2020, 10, 20463. [Google Scholar] [CrossRef]
- Choreño-Parra, J.A.; Jiménez-Álvarez, L.A.; Cruz-Lagunas, A.; Rodríguez-Reyna, T.S.; Ramírez-Martínez, G.; Sandoval-Vega, M.; Hernández-García, D.L.; Choreño-Parra, E.M.; Balderas-Martínez, Y.I.; Martinez-Sánchez, M.E.; et al. Clinical and Immunological Factors That Distinguish COVID-19 From Pandemic Influenza A(H1N1). Front. Immunol. 2021, 12, 593595. [Google Scholar] [CrossRef]
- Zhang, Z.Z.; Chen, D.P.; Liu, Q.B.; Gan, C.; Jiang, L.; Zhu, K.; Zhang, X.Y.; Xu, H.M.; Huang, A.L.; Long, Q.X.; et al. Clinical Features of Chinese Children with COVID-19 and Other Viral Respiratory Infections. Pediatr. Pulmonol. 2022, 57, 49–56. [Google Scholar] [CrossRef]
- Pang, J.; Xu, F.; Aondio, G.; Li, Y.; Fumagalli, A.; Lu, M.; Valmadre, G.; Wei, J.; Bian, Y.; Canesi, M.; et al. Efficacy and Tolerability of Bevacizumab in Patients with Severe COVID-19. Nat. Commun. 2021, 12, 814. [Google Scholar] [CrossRef]
- Charoute, H.; Elkarhat, Z.; Elkhattabi, L.; El Fahime, E.; Oukkache, N.; Rouba, H.; Barakat, A. Computational Screening of Potential Drugs against COVID-19 Disease: The Neuropilin-1 Receptor as Molecular Target. VirusDisease 2022, 33, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Perez-Miller, S.; Patek, M.; Moutal, A.; Cabel, C.R.; Thorne, C.A.; Campos, S.K.; Khanna, R. In Silico Identification and Validation of Inhibitors of the Interaction between Neuropilin Receptor 1 and SARS-CoV-2 Spike Protein. bioRxiv 2020. [Google Scholar] [CrossRef]
- Joshee, S.; Vatti, N.; Chang, C. Long-Term Effects of COVID-19. Mayo Clin. Proc. 2022, 97, 579–599. [Google Scholar] [CrossRef] [PubMed]
- Blomberg, B.; Mohn, K.G.I.; Brokstad, K.A.; Zhou, F.; Linchausen, D.W.; Hansen, B.A.; Lartey, S.; Onyango, T.B.; Kuwelker, K.; Sævik, M.; et al. Long COVID in a Prospective Cohort of Home-Isolated Patients. Nat. Med. 2021, 27, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- Sahanic, S.; Tymoszuk, P.; Ausserhofer, D.; Rass, V.; Pizzini, A.; Nordmeyer, G.; Hüfner, K.; Kurz, K.; Weber, P.M.; Sonnweber, T.; et al. Phenotyping of Acute and Persistent Coronavirus Disease 2019 Features in the Outpatient Setting: Exploratory Analysis of an International Cross-Sectional Online Survey. Clin. Infect. Dis. 2022, 75, e418–e431. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.-S.; Al-kuraishy, H.M.; Al-Gareeb, A.I.; Welson, N.N. Pathophysiology of Post-COVID Syndromes: A New Perspective. Virol. J. 2022, 19, 158. [Google Scholar] [CrossRef] [PubMed]
- Torres-Ruiz, J.; Lomelín-Gascón, J.; Lira-Luna, J.; Pérez-Fragoso, A.; Tapia-Conyer, R.; Nuñez-Aguirre, M.; Alcalá-Carmona, B.; Absalón-Aguilar, A.; Maravillas-Montero, J.L.; Mejía-Domínguez, N.R.; et al. FANSY POSTCOV: A Composite Clinical Immunological Predictive Index for Post-COVID-19 Syndrome Unveils Distinctive Features in a Cohort Study of Mild to Critical Patients. Clin. Transl. Med. 2021, 11, e623. [Google Scholar] [CrossRef]
- Du, J.; Wei, L.; Li, G.; Hua, M.; Sun, Y.; Wang, D.; Han, K.; Yan, Y.; Song, C.; Song, R.; et al. Persistent High Percentage of HLA-DR+CD38high CD8+ T Cells Associated With Immune Disorder and Disease Severity of COVID-19. Front. Immunol. 2021, 12, 735125. [Google Scholar] [CrossRef]
- Bobcakova, A.; Petriskova, J.; Vysehradsky, R.; Kocan, I.; Kapustova, L.; Barnova, M.; Diamant, Z.; Jesenak, M. Immune Profile in Patients With COVID-19: Lymphocytes Exhaustion Markers in Relationship to Clinical Outcome. Front. Cell. Infect. Microbiol. 2021, 11, 646688. [Google Scholar] [CrossRef]
- Bobcakova, A.; Barnova, M.; Vysehradsky, R.; Petriskova, J.; Kocan, I.; Diamant, Z.; Jesenak, M. Activated CD8+CD38+ Cells Are Associated With Worse Clinical Outcome in Hospitalized COVID-19 Patients. Front. Immunol. 2022, 13, 861666. [Google Scholar] [CrossRef]
- Garner, L.C.; Klenerman, P.; Provine, N.M. Insights into Mucosal-Associated Invariant T Cell Biology from Studies of Invariant Natural Killer T Cells. Front. Immunol. 2018, 9, 1478. [Google Scholar] [CrossRef] [Green Version]
- Maloney, J.P.; Gao, L. Proinflammatory Cytokines Increase Vascular Endothelial Growth Factor Expression in Alveolar Epithelial Cells. Mediat. Inflamm. 2015, 2015, 387842. [Google Scholar] [CrossRef] [Green Version]
- Arslan, N.G.; Aksakal, Ş.; Yılmam, İ.; Görgün, S. VEGF, IL-17 and IgG4 Levels of Patients with Lung Sequelae in Post-COVID-19 Period. Tuberk Toraks 2022, 70, 179–186. [Google Scholar] [CrossRef]
- Bonny, T.S.; Patel, E.U.; Zhu, X.; Bloch, E.M.; Grabowski, M.K.; Abraham, A.G.; Littlefield, K.; Shrestha, R.; Benner, S.E.; Laeyendecker, O.; et al. Cytokine and Chemokine Levels in Coronavirus Disease 2019 Convalescent Plasma. Open Forum Infect. Dis. 2021, 8, ofaa574. [Google Scholar] [CrossRef]
- Chi, Y.; Ge, Y.; Wu, B.; Zhang, W.; Wu, T.; Wen, T.; Liu, J.; Guo, X.; Huang, C.; Jiao, Y.; et al. Serum Cytokine and Chemokine Profile in Relation to the Severity of Coronavirus Disease 2019 in China. J. Infect. Dis. 2020, 222, 746–754. [Google Scholar] [CrossRef]
- Velikova, T.; Georgiev, T. SARS-CoV-2 Vaccines and Autoimmune Diseases amidst the COVID-19 Crisis. Rheumatol. Int. 2021, 41, 509–518. [Google Scholar] [CrossRef]
- European Medicines Agency Safety of COVID-19 Vaccines. Available online: https://www.ema.europa.eu/en/human-regulatory/overview/public-health-threats/coronavirus-disease-COVID-19/treatments-vaccines/vaccines-COVID-19/safety-COVID-19-vaccines (accessed on 13 October 2022).
- Garg, R.K.; Paliwal, V.K. Spectrum of Neurological Complications Following COVID-19 Vaccination. Neurol. Sci. 2021, 43, 3–40. [Google Scholar] [CrossRef]
- Patone, M.; Mei, X.W.; Handunnetthi, L.; Dixon, S.; Zaccardi, F.; Shankar-Hari, M.; Watkinson, P.; Khunti, K.; Harnden, A.; Coupland, C.A.C.; et al. Risks of Myocarditis, Pericarditis, and Cardiac Arrhythmias Associated with COVID-19 Vaccination or SARS-CoV-2 Infection. Nat. Med. 2021, 28, 410–422. [Google Scholar] [CrossRef]
- Allahyari, F.; Molaee, H.; Hosseini Nejad, J. COVID-19 Vaccines and Neurological Complications: A Systematic Review. Z. Naturforsch. C 2022. [Google Scholar] [CrossRef]
- Roberts, K.; Sidhu, N.; Russel, M.; Abbas, M.J. Psychiatric Pathology Potentially Induced by COVID-19 Vaccine. Prog. Neurol. Psychiatry 2021, 25, 8–10. [Google Scholar] [CrossRef]
- Escudero, C.; Prieto-Montaño, P.; Audicana, M.T. Adverse Reactions to Anti-Infective Vaccines: An Emerging Problem in the COVID-19 Era. Curr. Treat. Options Allergy 2022, 9, 250–272. [Google Scholar] [CrossRef]
- Won, T.; Gilotra, N.A.; Wood, M.K.; Hughes, D.M.; Talor, M.V.; Lovell, J.; Milstone, A.M.; Steenbergen, C.; Čiháková, D. Increased Interleukin 18-Dependent Immune Responses Are Associated With Myopericarditis After COVID-19 MRNA Vaccination. Front. Immunol. 2022, 13, 851620. [Google Scholar] [CrossRef]
- Kuodi, P.; Gorelik, Y.; Zayyad, H.; Wertheim, O.; Wiegler, K.B.; Abu Jabal, K.; Dror, A.A.; Nazzal, S.; Glikman, D.; Edelstein, M. Association between BNT162b2 Vaccination and Reported Incidence of Post-COVID-19 Symptoms: Cross-Sectional Study 2020-21, Israel. NPJ Vaccines 2022, 7, 101. [Google Scholar] [CrossRef]
- Parperis, K.; Constantinou, M. Remitting Seronegative Symmetrical Synovitis with Pitting Oedema Following BNT162b2 MRNA COVID-19 Vaccination. BMJ Case Rep. 2021, 14, e244479. [Google Scholar] [CrossRef]
- Arino, H.; Muramae, N.; Okano, M.; Mori, K.; Otsui, K.; Sakaguchi, K. Acute Onset of Remitting Seronegative Symmetrical Synovitis with Pitting Edema (RS3PE) Two Weeks after COVID-19 Vaccination with MRNA-1273 with Possible Activation of Parvovirus B19: A Case Report With Literature Review. Cureus 2022, 14, e24952. [Google Scholar] [CrossRef]
- Hovaguimian, A.; Gibbons, C.H. Diagnosis and Treatment of Pain in Small-Fiber Neuropathy. Curr. Pain Headache Rep. 2011, 15, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Abrams, R.M.C.; Simpson, D.M.; Navis, A.; Jette, N.; Zhou, L.; Shin, S.C. Small Fiber Neuropathy Associated with SARS-CoV-2 Infection. Muscle Nerve 2022, 65, 440–443. [Google Scholar] [CrossRef]
- Agnihotri, S.P.; Luis, C.V.S.; Kazamel, M. Autonomic Neuropathy as Post-Acute Sequela of SARS-CoV-2 Infection: A Case Report. J. Neurovirol. 2022, 28, 158–161. [Google Scholar] [CrossRef]
- Novak, P. Post COVID-19 Syndrome Associated with Orthostatic Cerebral Hypoperfusion Syndrome, Small Fiber Neuropathy and Benefit of Immunotherapy: A Case Report. Eneurologicalsci 2020, 21, 100276. [Google Scholar] [CrossRef]
- Oaklander, A.L.; Mills, A.J.; Kelley, M.; Toran, L.S.; Smith, B.; Dalakas, M.C.; Nath, A. Peripheral Neuropathy Evaluations of Patients With Prolonged Long COVID. Neurol. Neuroimmunol. Neuroinflamm. 2022, 9, e1146. [Google Scholar] [CrossRef]
- Shouman, K.; Vanichkachorn, G.; Cheshire, W.P.; Suarez, M.D.; Shelly, S.; Lamotte, G.J.; Sandroni, P.; Benarroch, E.E.; Berini, S.E.; Cutsforth-Gregory, J.K.; et al. Autonomic Dysfunction Following COVID-19 Infection: An Early Experience. Clin. Auton. Res. 2021, 31, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Schratzberger, P.; Walter, D.H.; Rittig, K.; Bahlmann, F.H.; Pola, R.; Curry, C.; Silver, M.; Krainin, J.G.; Weinberg, D.H.; Ropper, A.H.; et al. Reversal of Experimental Diabetic Neuropathy by VEGF Gene Transfer. J. Clin. Investig. 2001, 107, 1083–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibuya, M. VEGF-VEGFR System as a Target for Suppressing Inflammation and Other Diseases. Endocr. Metab. Immune Disord. Targets 2015, 15, 135–144. [Google Scholar] [CrossRef]
- Zhang, Q.; Lu, S.; Li, T.; Yu, L.; Zhang, Y.; Zeng, H.; Qian, X.; Bi, J.; Lin, Y. ACE2 Inhibits Breast Cancer Angiogenesis via Suppressing the VEGFa/VEGFR2/ERK Pathway. J. Exp. Clin. Cancer Res. 2019, 38, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Nan, B.; Yan, S.; Li, M.; Yao, Q.; Chen, C. C-Reactive Protein Decreases Expression of VEGF Receptors and Neuropilins and Inhibits VEGF165-Induced Cell Proliferation in Human Endothelial Cells. Biochem. Biophys. Res. Commun. 2005, 333, 1003–1010. [Google Scholar] [CrossRef]
- Prieto-Fernández, E.; Egia-Mendikute, L.; Vila-Vecilla, L.; Bosch, A.; Barreira-Manrique, A.; Lee, S.Y.; García-del Río, A.; Antoñana-Vildosola, A.; Jiménez-Lasheras, B.; Moreno-Cugnon, L.; et al. Hypoxia Reduces Cell Attachment of SARS-CoV-2 Spike Protein by Modulating the Expression of ACE2, Neuropilin-1, Syndecan-1 and Cellular Heparan Sulfate. Emerg. Microbes Infect. 2021, 10, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wei, Q.; Lin, Q.; Fang, J.; Wang, H.; Kwok, H.; Tang, H.; Nishiura, K.; Peng, J.; Tan, Z.; et al. Anti-Spike IgG Causes Severe Acute Lung Injury by Skewing Macrophage Responses during Acute SARS-CoV Infection. JCI Insight 2019, 4, e123158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, D.O.; Catalano, P.J.; Symonds, K.E.; Varey, A.H.R.; Ramani, P.; O’Dwyer, P.J.; Giantonio, B.J.; Meropol, N.J.; Benson, A.B.; Harper, S.J. Association between VEGF Splice Isoforms and Progression-Free Survival in Metastatic Colorectal Cancer Patients Treated with Bevacizumab. Clin. Cancer Res. 2012, 18, 6384–6391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hormbrey, E.; Gillespie, P.; Turner, K.; Han, C.; Roberts, A.; McGrouther, D.; Harris, A.L. A Critical Review of Vascular Endothelial Growth Factor (VEGF) Analysis in Peripheral Blood: Is the Current Literature Meaningful? Clin. Exp. Metastasis 2002, 19, 651–663. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Author, Year | Virus | Type of Study | Population Studied | Main Results |
|---|---|---|---|---|
| Morichi et al., 2017 [75] | Influenza | Case control study | -11 pts with IAE -6 pts with BM -24 pts with non-central nervous system infection | Increased levels of VEGF and PDGF in cerebrospinal fluid of IAE and BM pts compared with controls; positive correlation between VEGF and PDGF cerebrospinal fluid levels and worse prognosis according to the PCPC score |
| Wong et al., 2018 [77] | Influenza | Case control study | -27 pts with SARI -27 non-hospitalized pts with ILI | Higher VEGF-A serum levels during both acute disease and convalescence (2 weeks later) in pts with SARI compared to those with ILI |
| Bautista et al., 2013 [78] | Influenza | Case control study | -32 pts with A/H1N1 virus infection (17 with ARDS-related AKI and 15 ARDS pts without AKI) -18 HCs | Higher serum levels of VEGF in ARDS/AKI pts; significant association between VEGF serum levels and the risk of AKI development in ARDS pts; acute tubular damage and diffuse expression of VEGF in the cytoplasm of renal tubules with hydropic degeneration observed at immunohistochemistry assays in ARDS/AKI pts with fatal outcome |
| Pino et al., 2009 [79] | RSV | Observational, longitudinal study | -20 hospitalized children due to severe infection -12 HCs | Hypersecretion of VEGF in NPAs from hospitalized children at discharge and 1 yr later; potential role of VEGF in contributing to the pathogenesis of post-infectious wheezing |
| Bermejo-Martin et al., 2007 [80] | RSV | Case control study | -22 children aged <2 yrs with severe infection involving the lower respiratory tract -22 children with innocent heart murmurs | Positive correlation between plasma levels of VEGF and disease severity |
| Lee et al., 2000 [83] | RSV | Prospective study | -25 pts with documented influenza infection -47 pts with documented RSV infection -21 controls without documentable virus infection | Higher levels of VEGF in NPA samples from RSV-infected pts compared to the other groups; detection of VEGF 165– and 121–amino acid isoforms as the major VEGF representatives in nasal secretions of RSV-infected individuals |
| NRP Ligand | Binding Sequence (C-Terminus) |
|---|---|
| VEGF-A165 | DKPRR |
| VEGF-B167 | RKLRR |
| VEGF-B186 | RPQPR |
| VEGF-C | SIIRR |
| spike S1 subdomain | TNSPRRAR |
| Author, Year | Type of Study | Population Studied | Main Results |
|---|---|---|---|
| Rovas et al., 2021 [101] | Prospective, observational, cross-sectional study | -23 pts with moderate–severe COVID-19 -15 HCs | Increase in plasma VEGF-A levels and significant correlation with 60-day in-hospital mortality |
| Lim et al., 2021 [102] | Cross-sectional study | -37 pts with acute COVID-19 -40 convalescent pts -10 HCs | Increase in VEGF-A plasma levels during both acute disease and convalescence; positive correlation between VEGF-A plasma levels and convalescence severity; inverse correlation between VEGF-A plasma levels and CD8+CD56- MAIT cells during convalescence; positive association between VEGF-A plasma levels and HLA-DR+CD38+ CD8+ T cells in convalescent pts |
| Medeiros et al., 2022 [103] | Longitudinal study | -82 hospitalized moderate-to-severe COVID-19 pts, 41.5% of whom developing AKI | Increased serum levels of VEGF in pts with COVID-19-related AKI compared to pts without renal involvement |
| Choreño-Parra et al., 2021 [121] | Prospective cohort study | -10 pts with acute moderate COVID-19 -24 pts with acute severe COVID-19 -23 pts with pandemic influenza A(H1N1) | Increased serum levels of VEGF along with systemic, Th1 and Th2 cell cytokines in pts with COVID-19 but not in those with influenza |
| Zhang et al., 2022 [122] | Case control study | -20 hospitalized COVID-19 children -58 children with ARTI caused by RSV, influenza virus, and ADV -20 HCs | Significantly increased VEGF serum levels in SARS-CoV-2-infected pts compared to the other groups |
| Pang et al., 2021 [123] | Single-arm trial | -26 pts with severe COVID-19 treated with a single dose of bevacizumab | Improvement in PaO2/FiO2 parameters at days 1 and 7 from baseline; improvement in oxygen-support status in 92% of pts at day 28 from baseline; decrease in lung lesions on chest CT or X-ray within 7 days; normalization of body temperature within 72 h in 93% treated pts |
| Torres-Ruiz et al., 2021 [130] | Observational cohort study | -103 pts with previous COVID-19, 46.6% of whom diagnosed with long COVID-19 | Higher VEGF serum levels in pts with long COVID-19 than in pts without; VEGF appearing as the sole biomarker strongly associated with long COVID-19 in univariate analysis |
| Arslan et al., 2022 [136] | Observational, cross-sectional study | -32 pts with a previous COVID-19 diagnosis and no lung fibrosis on CT scan -32 pts with a previous COVID-19 diagnosis and lung fibrosis on CT scan -26 HCs | Higher VEGF serum concentration in pts with a previous COVID-19 diagnosis and no lung fibrosis compared to the other groups |
| Bonny et al., 2021 [137] | Observational, cross-sectional study | -20 healthy blood donors without previous SARS-CoV-2 infection -140 COVID-19 convalescent plasma donors | Higher plasma levels of IFN-γ, IL-10, IL-15, IL-21, and MIP-1 and lower levels of IL-1RA, IL-8, IL-16, and VEGF-A in convalescent plasma donors compared to controls |
| Chi et al., 2020 [138] | Observational, cross-sectional study | -70 SARS-CoV-2-infected pts (4 asymptomatic; 66 symptomatic) -4 convalescent pts -4 HCs | Higher VEGF serum levels in symptomatic cases compared with asymptomatic cases; lower serum levels of VEGF in convalescent pts than in symptomatic pts; positive correlation between VEGF serum concentration and male gender; weakly positive correlation between VEGF serum concentration and SARS-CoV-2 viral load |
| Clinical Domain | Long COVID-19 | COVID-19 Vaccine Side Effects |
|---|---|---|
| Constitutional |
|
|
| Musculoskeletal |
|
|
| Neuropsychiatric |
|
|
| Cardiovascular |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Talotta, R. Impaired VEGF-A-Mediated Neurovascular Crosstalk Induced by SARS-CoV-2 Spike Protein: A Potential Hypothesis Explaining Long COVID-19 Symptoms and COVID-19 Vaccine Side Effects? Microorganisms 2022, 10, 2452. https://doi.org/10.3390/microorganisms10122452
Talotta R. Impaired VEGF-A-Mediated Neurovascular Crosstalk Induced by SARS-CoV-2 Spike Protein: A Potential Hypothesis Explaining Long COVID-19 Symptoms and COVID-19 Vaccine Side Effects? Microorganisms. 2022; 10(12):2452. https://doi.org/10.3390/microorganisms10122452
Chicago/Turabian StyleTalotta, Rossella. 2022. "Impaired VEGF-A-Mediated Neurovascular Crosstalk Induced by SARS-CoV-2 Spike Protein: A Potential Hypothesis Explaining Long COVID-19 Symptoms and COVID-19 Vaccine Side Effects?" Microorganisms 10, no. 12: 2452. https://doi.org/10.3390/microorganisms10122452