
Screening of Bacteria Inhibiting Clostridium perfringens and Assessment of Their Beneficial Effects In Vitro and In Vivo with Whole Genome Sequencing Analysis
Abstract
:1. Introduction
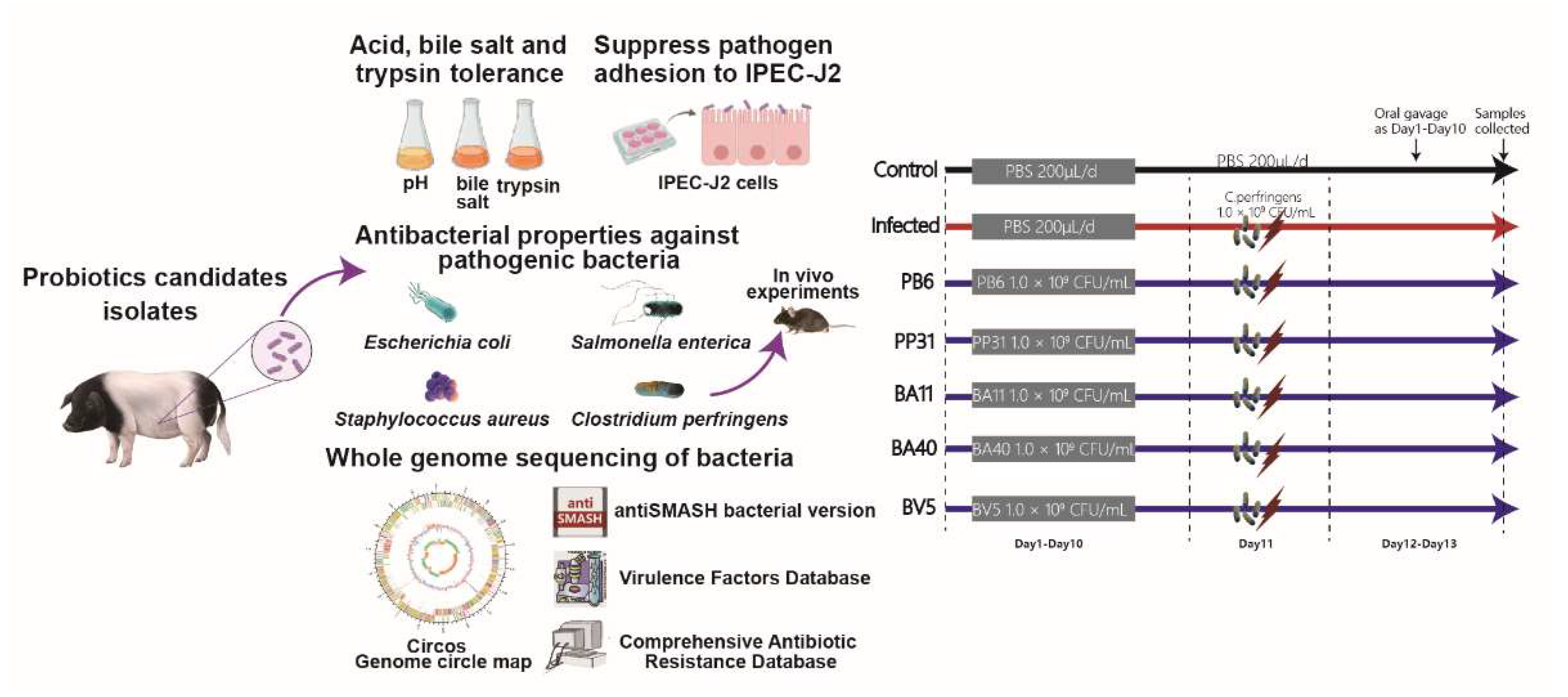
2. Materials and Methods
2.1. Isolation of Microbes from JinHua Pig Intestines
2.2. Bacterial Strains Preparation
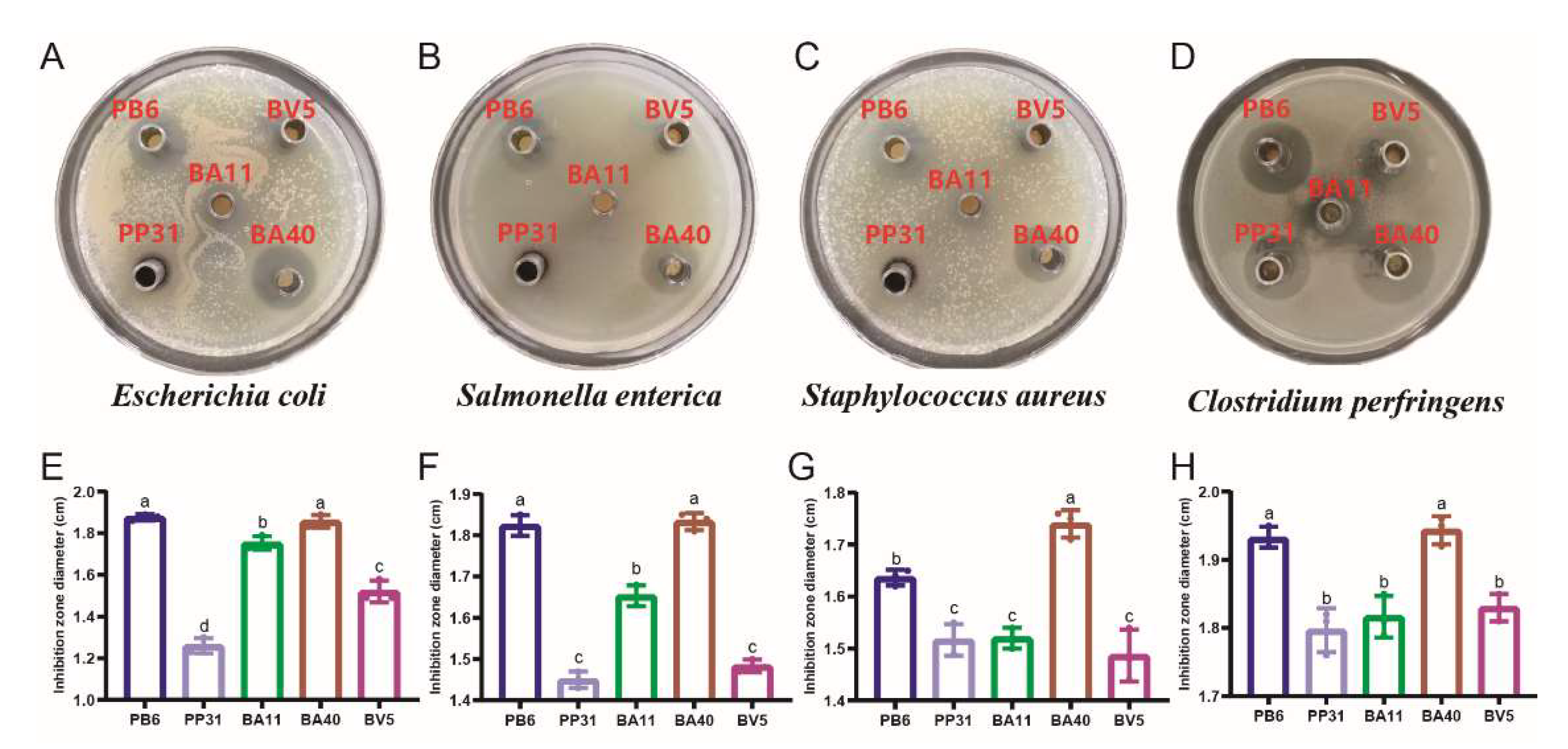
2.3. Preparation of Culture Supernatant and Agar-Well Diffusion Method
2.4. Identification of the Selected Isolates by 16S rRNA Sequencing
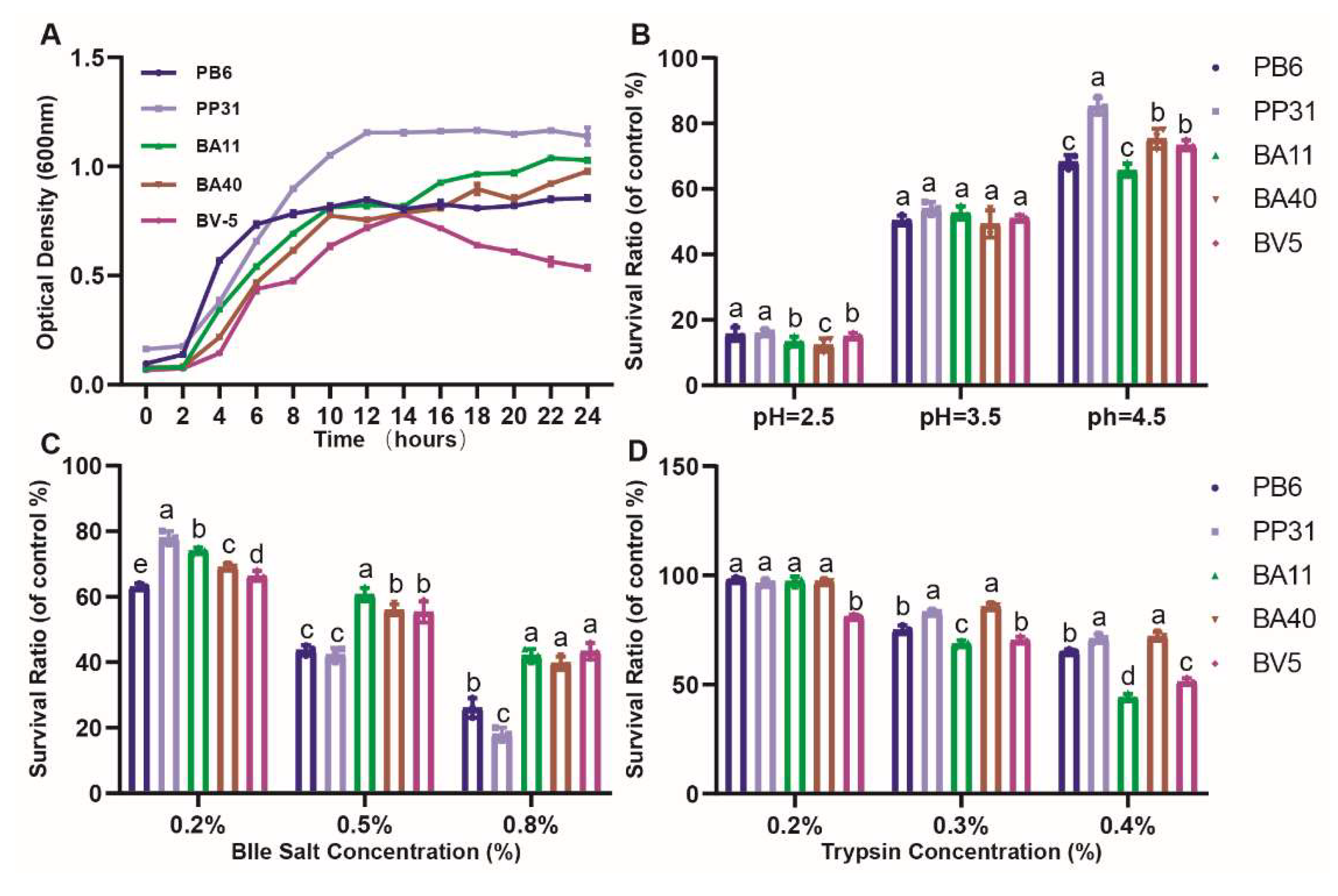
2.5. Growth Characteristics, Acid, Bile Salts and Trypsin Tolerance
2.6. Suppress Pathogen Adhesion to IPEC-J2 Cell
2.7. Whole Genome Sequencing and Analysis
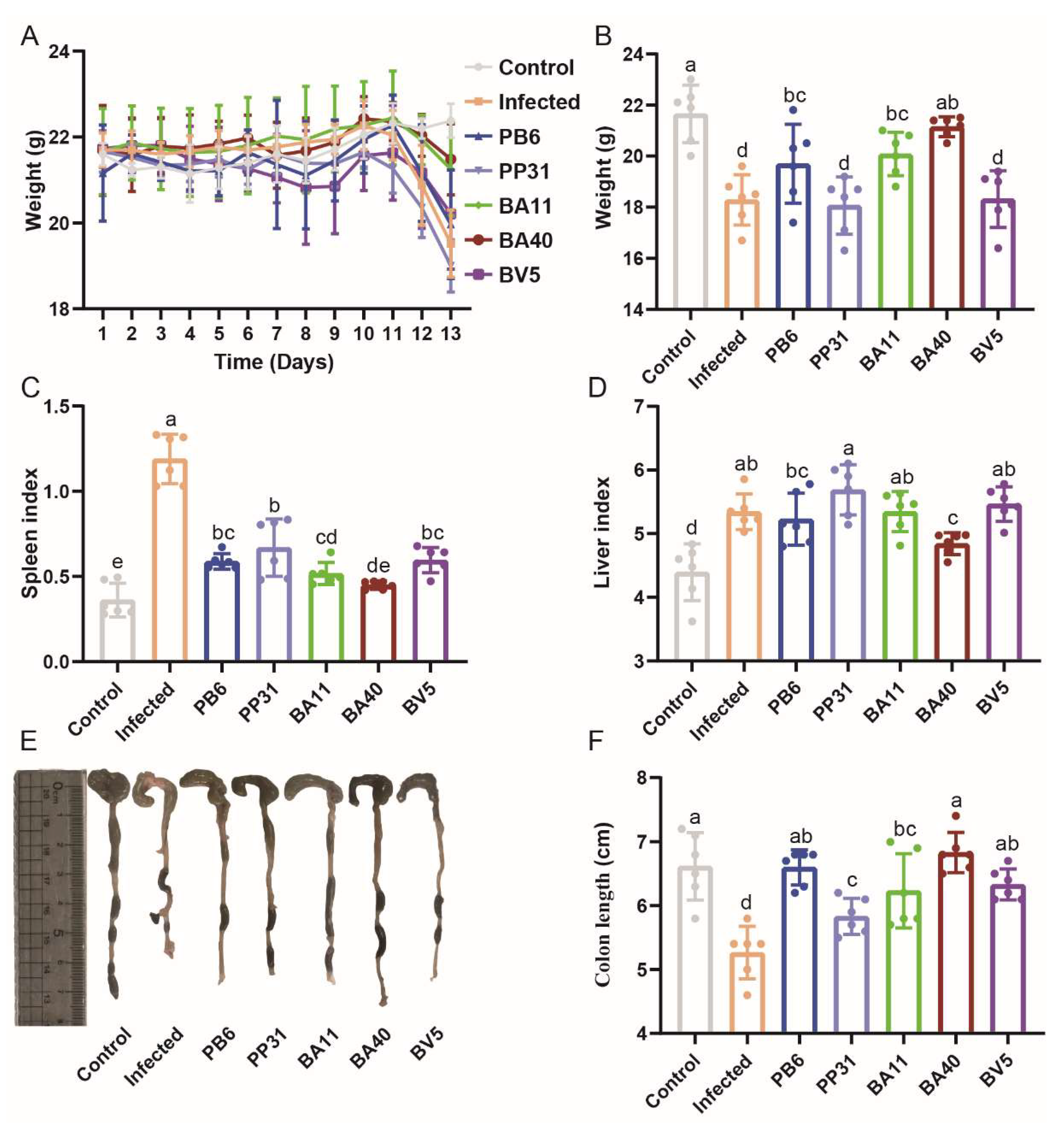
2.8. Animal Experiment
2.9. Sample Collection and Treatment
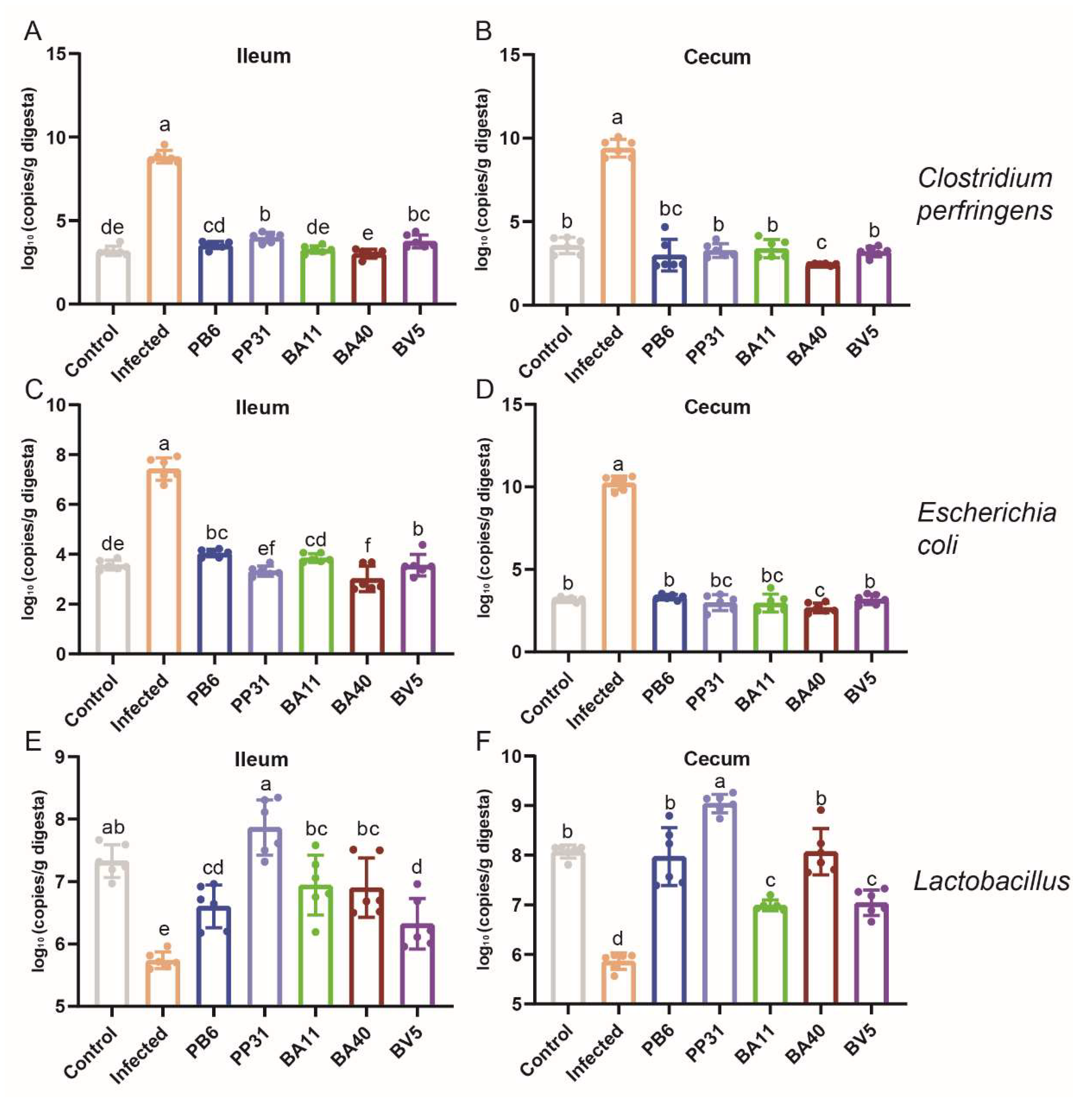
2.10. Bacteria Enumeration of Ileum and Cecum
2.11. Determination of Inflammatory Cytokines, Immunoglobulin, DAO, and DLA Concentrations
2.12. Statistical Analysis
3. Results
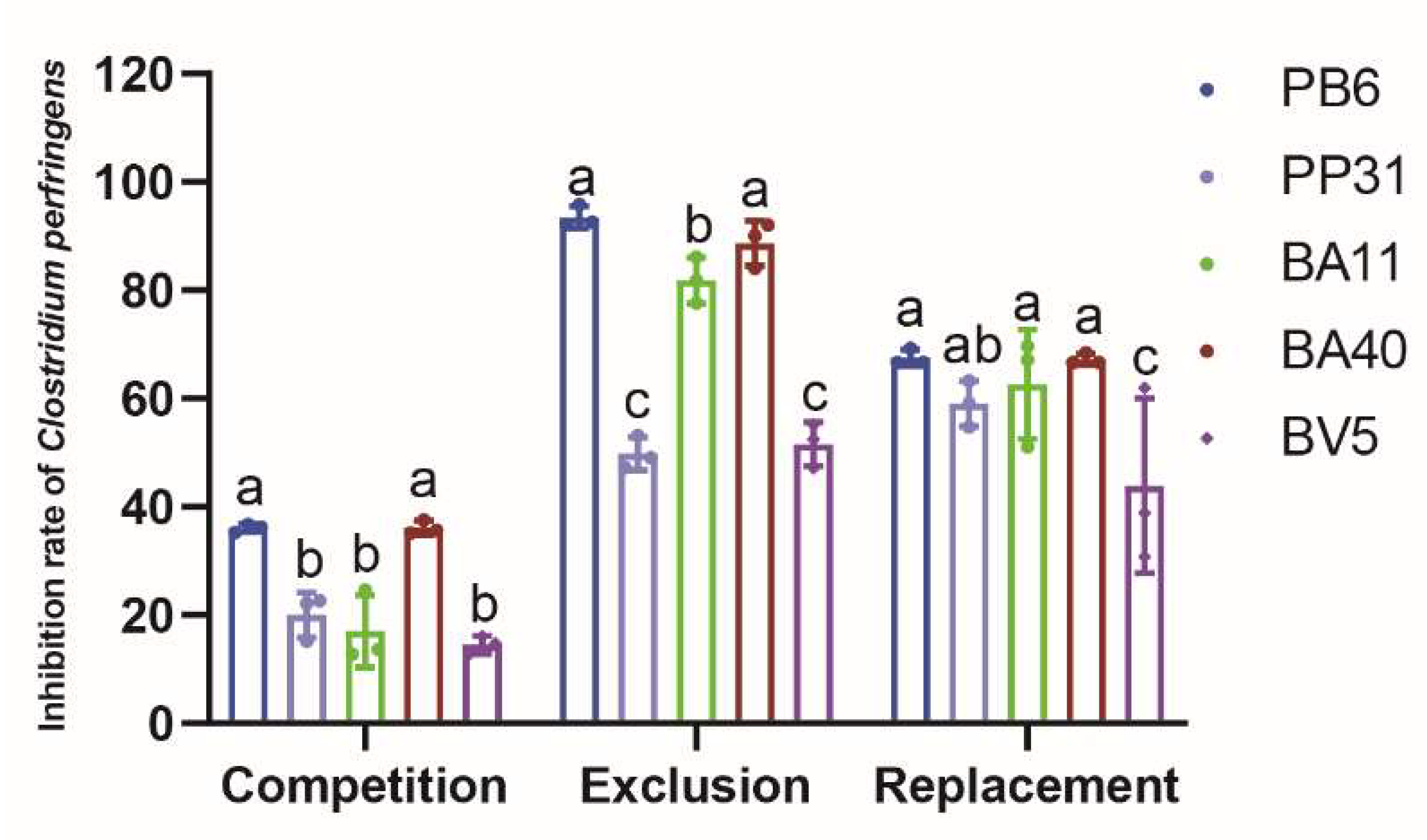
3.1. Pathogen Inhibition Using Well Diffusion Assay
3.2. Phenotypic Characteristics of Isolated Strains
3.3. Adhesion of Isolated Strains to IPEC-J2
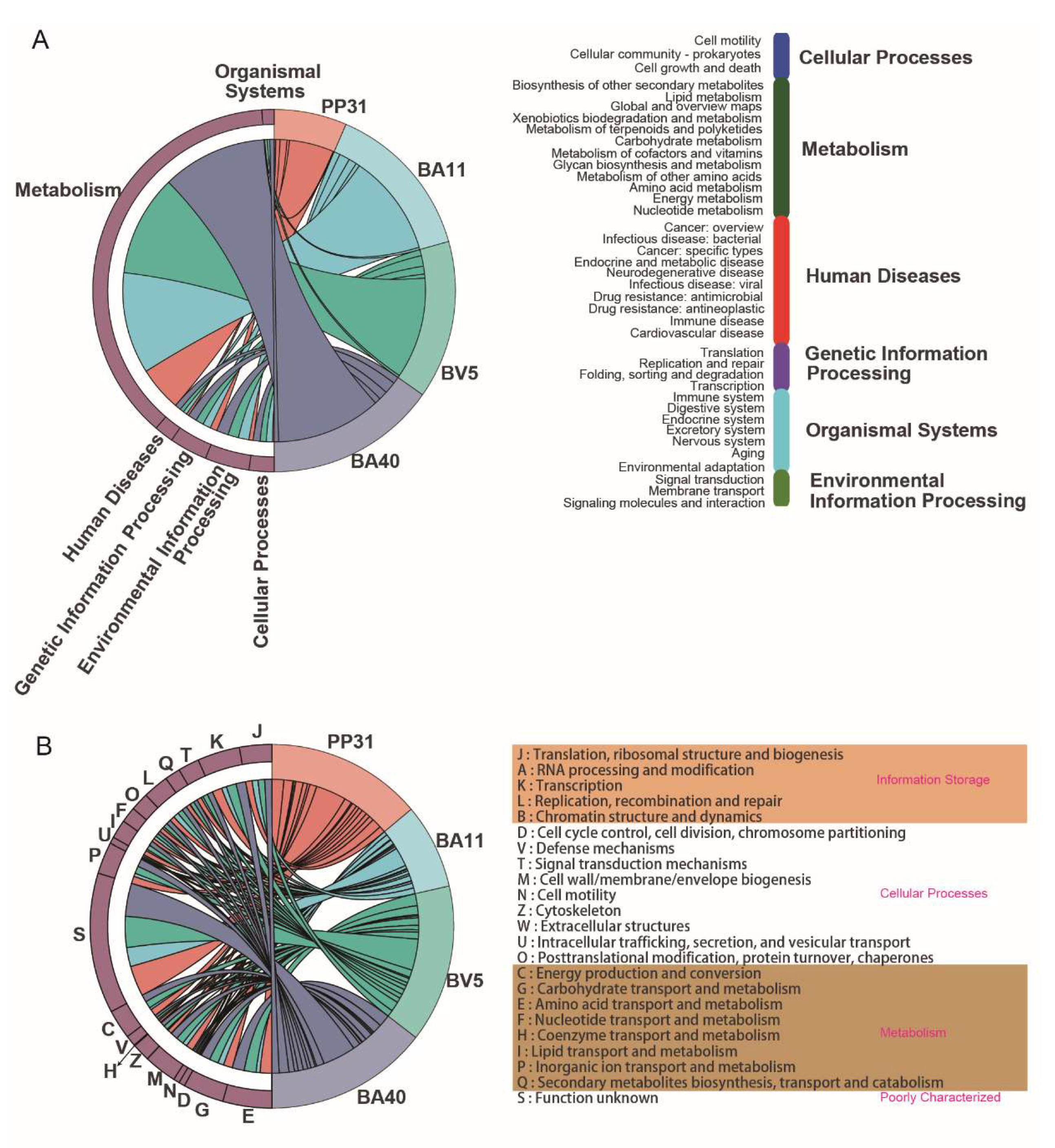
3.4. Whole-Genome Sequence of the Isolated Probiotics
3.5. Effect of Isolated Probiotics Administration on the Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lange, M.E.; Uwiera, R.R.E.; Inglis, G.D. Enteric Escherichia coli O157:H7 in Cattle, and the Use of Mice as a Model to Elucidate Key Aspects of the Host-Pathogen-Microbiota Interaction: A Review. Front. Vet. Sci. 2022, 9, 937866. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Huyghebaert, G.; Haesebrouck, F.; Ducatelle, R. Clostridium perfringens in poultry: An emerging threat for animal and public health. Avian Pathol. 2004, 33, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Song, L.; Wang, X.; Xu, Y.; Liu, Z.; Zhao, D.; Wang, S.; Fan, X.; Wang, Z.; Gao, C.; et al. A bovine lactoferricin-lactoferrampin-encoding Lactobacillus reuteri CO21 regulates the intestinal mucosal immunity and enhances the protection of piglets against enterotoxigenic Escherichia coli K88 challenge. Gut Microbes 2021, 13, 1956281. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.; Bai, Y.; Li, Y.; Huang, Y.; Li, L.; Wang, G.; Qu, L.; Wang, J.; Yu, L.-Y.; Hou, X. Changes in Gut Microbiota by the Lactobacillus casei Anchoring the K88 Fimbrial Protein Prevented Newborn Piglets From Clinical Diarrhea. Front. Cell. Infect. Microbiol. 2022, 12, 842007. [Google Scholar] [CrossRef] [PubMed]
- Mora, Z.V.; Macias-Rodriguez, M.E.; Arratia-Quijada, J.; Gonzalez-Torres, Y.S.; Nuno, K.; Villarruel-Lopez, A. Clostridium perfringens as Foodborne Pathogen in Broiler Production: Pathophysiology and Potential Strategies for Controlling Necrotic Enteritis. Animals 2020, 10, 1718. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Navarro, M.A.; Li, J.; Freedman, J.C.; Shrestha, A.; McClane, B.A. Comparative pathogenesis of enteric clostridial infections in humans and animals. Anaerobe 2018, 53, 11–20. [Google Scholar] [CrossRef]
- Walsh, T.R.; Wu, Y. China bans colistin as a feed additive for animals. Lancet Infect. Dis. 2016, 16, 1102–1103. [Google Scholar] [CrossRef]
- Cao, X.; Tang, L.; Zeng, Z.; Wang, B.; Zhou, Y.; Wang, Q.; Zou, P.; Li, W. Effects of Probiotics BaSC06 on Intestinal Digestion and Absorption, Antioxidant Capacity, Microbiota Composition, and Macrophage Polarization in Pigs for Fattening. Front. Vet. Sci. 2020, 7, 570593. [Google Scholar] [CrossRef]
- Ayalew, H.; Zhang, H.; Wang, J.; Wu, S.; Qiu, K.; Qi, G.; Tekeste, A.; Wassie, T.; Chanie, D. Potential Feed Additives as Antibiotic Alternatives in Broiler Production. Front. Vet. Sci. 2022, 9, 916473. [Google Scholar] [CrossRef]
- Kulkarni, R.R.; Gaghan, C.; Gorrell, K.; Sharif, S.; Taha-Abdelaziz, K. Probiotics as Alternatives to Antibiotics for the Prevention and Control of Necrotic Enteritis in Chickens. Pathogens 2022, 11, 692. [Google Scholar] [CrossRef]
- Ogbuewu, I.P.; Mabelebele, M.; Sebola, N.A.; Mbajiorgu, C. Bacillus Probiotics as Alternatives to In-feed Antibiotics and Its Influence on Growth, Serum Chemistry, Antioxidant Status, Intestinal Histomorphology, and Lesion Scores in Disease-Challenged Broiler Chickens. Front. Vet. Sci. 2022, 9, 876725. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Marco, M.L.; Sanders, M.E.; Ganzle, M.; Arrieta, M.C.; Cotter, P.D.; De Vuyst, L.; Hill, C.; Holzapfel, W.; Lebeer, S.; Merenstein, D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on fermented foods. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 196–208. [Google Scholar] [CrossRef]
- van Zyl, W.F.; Deane, S.M.; Dicks, L.M.T. Molecular insights into probiotic mechanisms of action employed against intestinal pathogenic bacteria. Gut Microbes 2020, 12, 1831339. [Google Scholar] [CrossRef]
- Bai, Y.; Ma, K.; Li, J.; Ren, Z.; Zhang, J.; Shan, A. Lactobacillus rhamnosus GG ameliorates DON-induced intestinal damage depending on the enrichment of beneficial bacteria in weaned piglets. J. Anim. Sci. Biotechnol. 2022, 13, 90. [Google Scholar] [CrossRef]
- Kruse, S.; Schenk, M.; Pierre, F.; Morlock, G.E. Bacillus subtilis spores in probiotic feed quantified via bacterial metabolite using planar chromatography. Anal. Chim. Acta 2022, 1221, 340124. [Google Scholar] [CrossRef]
- Lu, S.; Na, K.; Li, Y.; Zhang, L.; Fang, Y.; Guo, X. Bacillus-derived probiotics: Metabolites and mechanisms involved in bacteria-host interactions. Crit. Rev. Food Sci. Nutr. 2022, 1–14. [Google Scholar] [CrossRef]
- Shamshirgaran, M.A.; Golchin, M.; Mohammadi, E. Lactobacillus casei displaying Clostridium perfringens NetB antigen protects chickens against necrotic enteritis. Appl. Microbiol. Biotechnol. 2022, 106, 6441–6453. [Google Scholar] [CrossRef]
- Aljumaah, M.R.; Alkhulaifi, M.M.; Abudabos, A.M.; Aljumaah, R.S.; Alsaleh, A.N.; Stanley, D. Bacillus subtilis PB6 based probiotic supplementation plays a role in the recovery after the necrotic enteritis challenge. PLoS ONE 2020, 15, e0232781. [Google Scholar] [CrossRef]
- Foligne, B.; Peys, E.; Vandenkerckhove, J.; Van Hemel, J.; Dewulf, J.; Breton, J.; Pot, B. Spores from two distinct colony types of the strain Bacillus subtilis PB6 substantiate anti-inflammatory probiotic effects in mice. Clin. Nutr. 2012, 31, 987–994. [Google Scholar] [CrossRef]
- Guo, S.; Xv, J.; Li, Y.; Bi, Y.; Hou, Y.; Ding, B. Interactive effects of dietary vitamin K3 and Bacillus subtilis PB6 on the growth performance and tibia quality of broiler chickens with sex separate rearing. Animal 2020, 14, 1610–1618. [Google Scholar] [CrossRef]
- Jayaraman, S.; Thangavel, G.; Kurian, H.; Mani, R.; Mukkalil, R.; Chirakkal, H. Bacillus subtilis PB6 improves intestinal health of broiler chickens challenged with Clostridium perfringens-induced necrotic enteritis. Poult. Sci. 2013, 92, 370–374. [Google Scholar] [CrossRef]
- Wang, C.; Wei, S.; Chen, N.; Xiang, Y.; Wang, Y.; Jin, M. Characteristics of gut microbiota in pigs with different breeds, growth periods and genders. Microb. Biotechnol. 2022, 15, 793–804. [Google Scholar] [CrossRef]
- Ripamonti, B.; Agazzi, A.; Bersani, C.; De Dea, P.; Pecorini, C.; Pirani, S.; Rebucci, R.; Savoini, G.; Stella, S.; Stenico, A.; et al. Screening of species-specific lactic acid bacteria for veal calves multi-strain probiotic adjuncts. Anaerobe 2011, 17, 97–105. [Google Scholar] [CrossRef]
- Lone, A.; Mottawea, W.; Ait Chait, Y.; Hammami, R. Dual Inhibition of Salmonella enterica and Clostridium perfringens by New Probiotic Candidates Isolated from Chicken Intestinal Mucosa. Microorganisms 2021, 9, 166. [Google Scholar] [CrossRef]
- Fu, J.; Wang, T.; Xiao, X.; Cheng, Y.; Wang, F.; Jin, M.; Wang, Y.; Zong, X. Clostridium Butyricum ZJU-F1 Benefits the Intestinal Barrier Function and Immune Response Associated with Its Modulation of Gut Microbiota in Weaned Piglets. Cells 2021, 10, 527. [Google Scholar] [CrossRef]
- Tan, L.L.; Tan, C.H.; Ng, N.K.J.; Tan, Y.H.; Conway, P.L.; Loo, S.C.J. Potential Probiotic Strains From Milk and Water Kefir Grains in Singapore-Use for Defense Against Enteric Bacterial Pathogens. Front. Microbiol. 2022, 13, 857720. [Google Scholar] [CrossRef] [PubMed]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Li, W.; Su, W.; Wen, C.; Gong, T.; Zhang, Y.; Wang, Y.; Jin, M.; Lu, Z. Protective Effects of Bacillus amyloliquefaciens 40 Against Clostridium perfringens Infection in Mice. Front. Nutr. 2021, 8, 733591. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhang, J.; Li, Y.; Li, K.; Gong, S.; Li, F.; Wang, P.; Iqbal, M.; Kulyar, M.F.; Li, J. Probiotic Potential of Bacillus licheniformis and Bacillus pumilus Isolated from Tibetan Yaks, China. Probiotics Antimicrob. Proteins 2022, 14, 579–594. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhang, J.; Zhao, Y.; Lin, Z.; Ji, L.; Ma, X. Tibetan Pig-Derived Probiotic Lactobacillus amylovorus SLZX20-1 Improved Intestinal Function via Producing Enzymes and Regulating Intestinal Microflora. Front. Nutr. 2022, 9, 846991. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Zhang, Q.; Tan, B.; Li, M.; Zhang, W.; Feng, J. A metagenomic view of how different carbon sources enhance the aniline and simultaneous nitrogen removal capacities in the aniline degradation system. Bioresour. Technol. 2021, 335, 125277. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Cao, X.; Wang, H.; Xiao, X.; Wang, Y.; Lu, Z. Cathelicidin-WA Facilitated Intestinal Fatty Acid Absorption Through Enhancing PPAR-gamma Dependent Barrier Function. Front. Immunol. 2019, 10, 1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinttila, T.; Kassinen, A.; Malinen, E.; Krogius, L.; Palva, A. Development of an extensive set of 16S rDNA-targeted primers for quantification of pathogenic and indigenous bacteria in faecal samples by real-time PCR. J. Appl. Microbiol. 2004, 97, 1166–1177. [Google Scholar] [CrossRef]
- Jiang, Z.; Su, W.; Wen, C.; Li, W.; Zhang, Y.; Gong, T.; Du, S.; Wang, X.; Lu, Z.; Jin, M.; et al. Effect of Porcine Clostridium perfringens on Intestinal Barrier, Immunity, and Quantitative Analysis of Intestinal Bacterial Communities in Mice. Front. Vet. Sci. 2022, 9, 881878. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Cheng, Y.; Fu, J.; Lu, Z.; Wang, F.; Jin, M.; Zong, X.; Wang, Y. Gut Immunity and Microbiota Dysbiosis Are Associated with Altered Bile Acid Metabolism in LPS-Challenged Piglets. Oxid. Med. Cell Longev. 2021, 2021, 6634821. [Google Scholar] [CrossRef] [PubMed]
- Acharjee, M.; Hasan, F.; Islam, T.; Nur, I.T.; Begum, N.; Mazumder, C.; Lubna, M.A.; Zerin, N.; Shahriar, A.; Mahmud, M.R. In-vitro antibacterial activity of commercially available probiotics on food-borne pathogens along with their synergistic effects with synthetic drugs. Metab. Open 2022, 14, 100187. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Micciche, A.C.; Foley, S.L.; Pavlidis, H.O.; McIntyre, D.R.; Ricke, S.C. A Review of Prebiotics Against Salmonella in Poultry: Current and Future Potential for Microbiome Research Applications. Front. Vet. Sci. 2018, 5, 191. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.O.; Lobato, F.C. Clostridium perfringens: A review of enteric diseases in dogs, cats and wild animals. Anaerobe 2015, 33, 14–17. [Google Scholar] [CrossRef]
- Silva, R.O.S.; Oliveira Junior, C.A.; Guedes, R.M.C.; Lobato, F.C.F. Clostridium perfringens: A review of the disease in pigs, horses and broiler chickens. Ciência Rural 2015, 45, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Lo Verso, L.; Lessard, M.; Talbot, G.; Fernandez, B.; Fliss, I. Isolation and Selection of Potential Probiotic Bacteria from the Pig Gastrointestinal Tract. Probiotics Antimicrob. Proteins 2018, 10, 299–312. [Google Scholar] [CrossRef]
- Adhikari, B.; Kwon, Y.M. Characterization of the Culturable Subpopulations of Lactobacillus in the Chicken Intestinal Tract as a Resource for Probiotic Development. Front. Microbiol. 2017, 8, 1389. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Li, Y.; Zhang, J.; Yang, Q. Co-administration of Bacillus subtilis RJGP16 and Lactobacillus salivarius B1 strongly enhances the intestinal mucosal immunity of piglets. Res. Vet. Sci. 2013, 94, 62–68. [Google Scholar] [CrossRef]
- Foligne, B.; Daniel, C.; Pot, B. Probiotics from research to market: The possibilities, risks and challenges. Curr. Opin. Microbiol. 2013, 16, 284–292. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Forsythe, S.J.; El-Nezami, H. Probiotics interaction with foodborne pathogens: A potential alternative to antibiotics and future challenges. Crit. Rev. Food Sci. Nutr. 2019, 59, 3320–3333. [Google Scholar] [CrossRef]
- Dowarah, R.; Verma, A.K.; Agarwal, N.; Singh, P.; Singh, B.R. Selection and characterization of probiotic lactic acid bacteria and its impact on growth, nutrient digestibility, health and antioxidant status in weaned piglets. PLoS ONE 2018, 13, e0192978. [Google Scholar] [CrossRef] [Green Version]
- Musikasang, H.; Tani, A.; H-kittikun, A.; Maneerat, S. Probiotic potential of lactic acid bacteria isolated from chicken gastrointestinal digestive tract. World J. Microbiol. Biotechnol. 2009, 25, 1337–1345. [Google Scholar] [CrossRef]
- Chen, G.; Fang, Q.; Liao, Z.; Xu, C.; Liang, Z.; Liu, T.; Zhong, Q.; Wang, L.; Fang, X.; Wang, J. Detoxification of Aflatoxin B1 by a Potential Probiotic Bacillus amyloliquefaciens WF2020. Front. Microbiol. 2022, 13, 891091. [Google Scholar] [CrossRef]
- Mulaw, G.; Sisay Tessema, T.; Muleta, D.; Tesfaye, A. In Vitro Evaluation of Probiotic Properties of Lactic Acid Bacteria Isolated from Some Traditionally Fermented Ethiopian Food Products. Int. J. Microbiol. 2019, 2019, 7179514. [Google Scholar] [CrossRef]
- Ngamsomchat, A.; Kaewkod, T.; Konkit, M.; Tragoolpua, Y.; Bovonsombut, S.; Chitov, T. Characterisation of Lactobacillus plantarum of Dairy-Product Origin for Probiotic Chevre Cheese Production. Foods 2022, 11, 934. [Google Scholar] [CrossRef]
- WoldemariamYohannes, K.; Wan, Z.; Yu, Q.; Li, H.; Wei, X.; Liu, Y.; Wang, J.; Sun, B. Prebiotic, Probiotic, Antimicrobial, and Functional Food Applications of Bacillus amyloliquefaciens. J. Agric. Food Chem. 2020, 68, 14709–14727. [Google Scholar] [CrossRef]
- Kelly, C.G.; Younson, J.S. Anti-adhesive strategies in the prevention of infectious disease at mucosal surfaces. Expert Opin. Investig. Drugs 2000, 9, 1711–1721. [Google Scholar] [CrossRef]
- Anonye, B.O.; Hassall, J.; Patient, J.; Detamornrat, U.; Aladdad, A.M.; Schuller, S.; Rose, F.; Unnikrishnan, M. Probing Clostridium difficile Infection in Complex Human Gut Cellular Models. Front. Microbiol. 2019, 10, 879. [Google Scholar] [CrossRef]
- Misawa, Y.; Kelley, K.A.; Wang, X.; Wang, L.; Park, W.B.; Birtel, J.; Saslowsky, D.; Lee, J.C. Staphylococcus aureus Colonization of the Mouse Gastrointestinal Tract Is Modulated by Wall Teichoic Acid, Capsule, and Surface Proteins. PLoS Pathog. 2015, 11, e1005061. [Google Scholar] [CrossRef]
- Redondo, L.M.; Carrasco, J.M.; Redondo, E.A.; Delgado, F.; Miyakawa, M.E. Clostridium perfringens type E virulence traits involved in gut colonization. PLoS ONE 2015, 10, e0121305. [Google Scholar] [CrossRef] [Green Version]
- Tsilia, V.; Uyttendaele, M.; Kerckhof, F.M.; Rajkovic, A.; Heyndrickx, M.; Van de Wiele, T. Bacillus cereus Adhesion to Simulated Intestinal Mucus Is Determined by Its Growth on Mucin, Rather Than Intestinal Environmental Parameters. Foodborne Pathog. Dis. 2015, 12, 904–913. [Google Scholar] [CrossRef]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen elimination by probiotic Bacillus via signalling interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Zhang, X. Alien species invasion of deep-sea bacteria into mouse gut microbiota. J. Adv. Res. 2022, In press. [Google Scholar] [CrossRef]
- Liu, M.; Liu, L.; Jia, S.; Li, S.; Zou, Y.; Zhong, C. Complete genome analysis of Gluconacetobacter xylinus CGMCC 2955 for elucidating bacterial cellulose biosynthesis and metabolic regulation. Sci. Rep. 2018, 8, 6266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ji, H.; Zhang, D.; Liu, H.; Wang, S.; Wang, J.; Wang, Y. Complete Genome Sequencing of Lactobacillus plantarum ZLP001, a Potential Probiotic That Enhances Intestinal Epithelial Barrier Function and Defense Against Pathogens in Pigs. Front. Physiol. 2018, 9, 1689. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yi, D.; Xu, H.; Tan, Z.; Meng, Y.; Wu, T.; Wang, L.; Zhao, D.; Hou, Y. Dietary supplementation with sodium gluconate improves the growth performance and intestinal function in weaned pigs challenged with a recombinant Escherichia coli strain. BMC Vet. Res. 2022, 18, 303. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Fu, J.; Xu, B.; Wang, Y.; Jin, M. Interplay between gut microbiota and antimicrobial peptides. Anim. Nutr. 2020, 6, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Torchinsky, M.B.; Garaude, J.; Martin, A.P.; Blander, J.M. Innate immune recognition of infected apoptotic cells directs T(H)17 cell differentiation. Nature 2009, 458, 78–82. [Google Scholar] [CrossRef]
- Gong, L.; Wang, B.; Zhou, Y.; Tang, L.; Zeng, Z.; Zhang, H.; Li, W. Protective Effects of Lactobacillus plantarum 16 and Paenibacillus polymyxa 10 Against Clostridium perfringens Infection in Broilers. Front. Immunol. 2020, 11, 628374. [Google Scholar] [CrossRef]
- Djaldetti, M.; Bessler, H. Probiotic strains modulate cytokine production and the immune interplay between human peripheral blood mononucear cells and colon cancer cells. FEMS Microbiol. Lett. 2017, 364, fnx014. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Wang, X.; Zhu, L.; Tu, Y.; Chen, W.; Gong, L.; Pan, T.; Lin, H.; Lin, J.; Sun, H.; et al. Lactobacillus rhamnosus GG combined with inosine ameliorates alcohol-induced liver injury through regulation of intestinal barrier and Treg/Th1 cells. Toxicol. Appl. Pharmacol. 2022, 439, 115923. [Google Scholar] [CrossRef]
- Garcia-Ortiz, A.; Serrador, J.M. Nitric Oxide Signaling in T Cell-Mediated Immunity. Trends Mol. Med. 2018, 24, 412–427. [Google Scholar] [CrossRef]
- Chen, Y.; Miao, L.; Yao, Y.; Wu, W.; Wu, X.; Gong, C.; Qiu, L.; Chen, J. Dexmedetomidine Ameliorate CLP-Induced Rat Intestinal Injury via Inhibition of Inflammation. Mediat. Inflamm. 2015, 2015, 918361. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Zhang, L.; Gan, Z.; Xiong, H.; Yu, C.; Du, H.; Wang, Y. High therapeutic efficacy of Cathelicidin-WA against postweaning diarrhea via inhibiting inflammation and enhancing epithelial barrier in the intestine. Sci. Rep. 2016, 6, 25679. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer Sequence (5′ to 3′) | Amplicon Size, bp | Reference |
|---|---|---|---|
| Clostridium perfringens | F: ATGCAAGTCGAGCGAKG R: TATGCGGTATTAATCTYCCTTT | 105 | [35] |
| Lactobacillus subgroup | F: AGCAGTAGGGAATCTTCCA R: CACCGCTACACATGGAG | 341 | [35] |
| Escherichia subgroup | F: GTTAATACCTTTGCTCATTGA R: ACCAGGGTATCTAATCCTGT | 340 | [35] |
| Strains | 16S rRNA Identity | Scaffold Number | Genome Size (bp) | GC % | N50 (bp) | Sequencing Depth (x) | Completeness (%) | Genebank Accession Number |
|---|---|---|---|---|---|---|---|---|
| PP31 | Pediococcuspentosaceus | 36 | 1818617 | 37.3 | 260845 | 659.14 | 97.3 | ON228197 |
| BA11 | Bacillus amyloliquefaciens | 36 | 3927418 | 46.42 | 565069 | 317.09 | 99.3 | ON227093 |
| BA40 | Bacillus amyloliquefaciens | 143 | 3969383 | 46.4 | 382464 | 322.08 | 99.3 | ON227058 |
| BV5 | Bacillus velezensis | 36 | 3867471 | 46.49 | 608593 | 328.7 | 99.3 | ON227128 |
| Strains | Cluster Type | MIBiG Accession | Similarity | Location (Start–End) | Gene Number |
|---|---|---|---|---|---|
| PP31 | Coagulin | BGC0000617 | 40% | 14107–19888 | 6 |
| BA11 | Bacillaene | BGC0001089 | 100% | 142968–252589 | 52 |
| Macrolactin H | BGC0000181 | 100% | 471746–559949 | 44 | |
| Bacilysin | BGC0001184 | 100% | 514804–556223 | 42 | |
| Bacillibactin | BGC0000309 | 100% | 73512–122977 | 44 | |
| Difficidin | BGC0000176 | 53% | 131390–177054 | 31 | |
| Locillomycin | BGC0001005 | 35% | 23265–45904 | 20 | |
| Fengycin | BGC0001095 | 20% | 1–13205 | 3 | |
| BA40 | Bacillibactin | BGC0000309 | 100% | 73491–125283 | 45 |
| Bacilysin | BGC0001184 | 100% | 661592–703011 | 42 | |
| Macrolactin H | BGC0000181 | 100% | 53617–141453 | 44 | |
| Bacillaene | BGC0001089 | 100% | 360981–470835 | 52 | |
| Fengycin | BGC0001095 | 80% | 535580–623198 | 46 | |
| Difficidin | BGC0000176 | 46% | 273837–319624 | 30 | |
| Plipastatin | BGC0000407 | 30% | 1–13991 | 12 | |
| BV5 | Macrolactin H | BGC0000181 | 100% | 507723–595523 | 44 |
| Bacilysin | BGC0001184 | 100% | 265403–306822 | 43 | |
| Mersacidin | BGC0000527 | 100% | 443880–467069 | 19 | |
| Amylocyclicin | BGC0000616 | 100% | 118402–125992 | 9 | |
| Fengycin | BGC0001095 | 80% | 1–86692 | 46 | |
| Bacillaene | BGC0001089 | 71% | 817963–854795 | 25 | |
| Difficidin | BGC0000176 | 53% | 1–45718 | 30 | |
| Surfactin | BGC0000433 | 39% | 681718–707084 | 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Z.; Su, W.; Yang, M.; Li, W.; Gong, T.; Zhang, Y.; Wen, C.; Wang, X.; Wang, Y.; Jin, M.; et al. Screening of Bacteria Inhibiting Clostridium perfringens and Assessment of Their Beneficial Effects In Vitro and In Vivo with Whole Genome Sequencing Analysis. Microorganisms 2022, 10, 2056. https://doi.org/10.3390/microorganisms10102056
Jiang Z, Su W, Yang M, Li W, Gong T, Zhang Y, Wen C, Wang X, Wang Y, Jin M, et al. Screening of Bacteria Inhibiting Clostridium perfringens and Assessment of Their Beneficial Effects In Vitro and In Vivo with Whole Genome Sequencing Analysis. Microorganisms. 2022; 10(10):2056. https://doi.org/10.3390/microorganisms10102056
Chicago/Turabian StyleJiang, Zipeng, Weifa Su, Mingzhi Yang, Wentao Li, Tao Gong, Yu Zhang, Chaoyue Wen, Xinxia Wang, Yizhen Wang, Mingliang Jin, and et al. 2022. "Screening of Bacteria Inhibiting Clostridium perfringens and Assessment of Their Beneficial Effects In Vitro and In Vivo with Whole Genome Sequencing Analysis" Microorganisms 10, no. 10: 2056. https://doi.org/10.3390/microorganisms10102056