

Effects of Variety, Plant Location, and Season on the Phyllosphere Bacterial Community Structure of Alfalfa (Medicago sativa L.)

Abstract
:1. Background
2. Materials and Methods
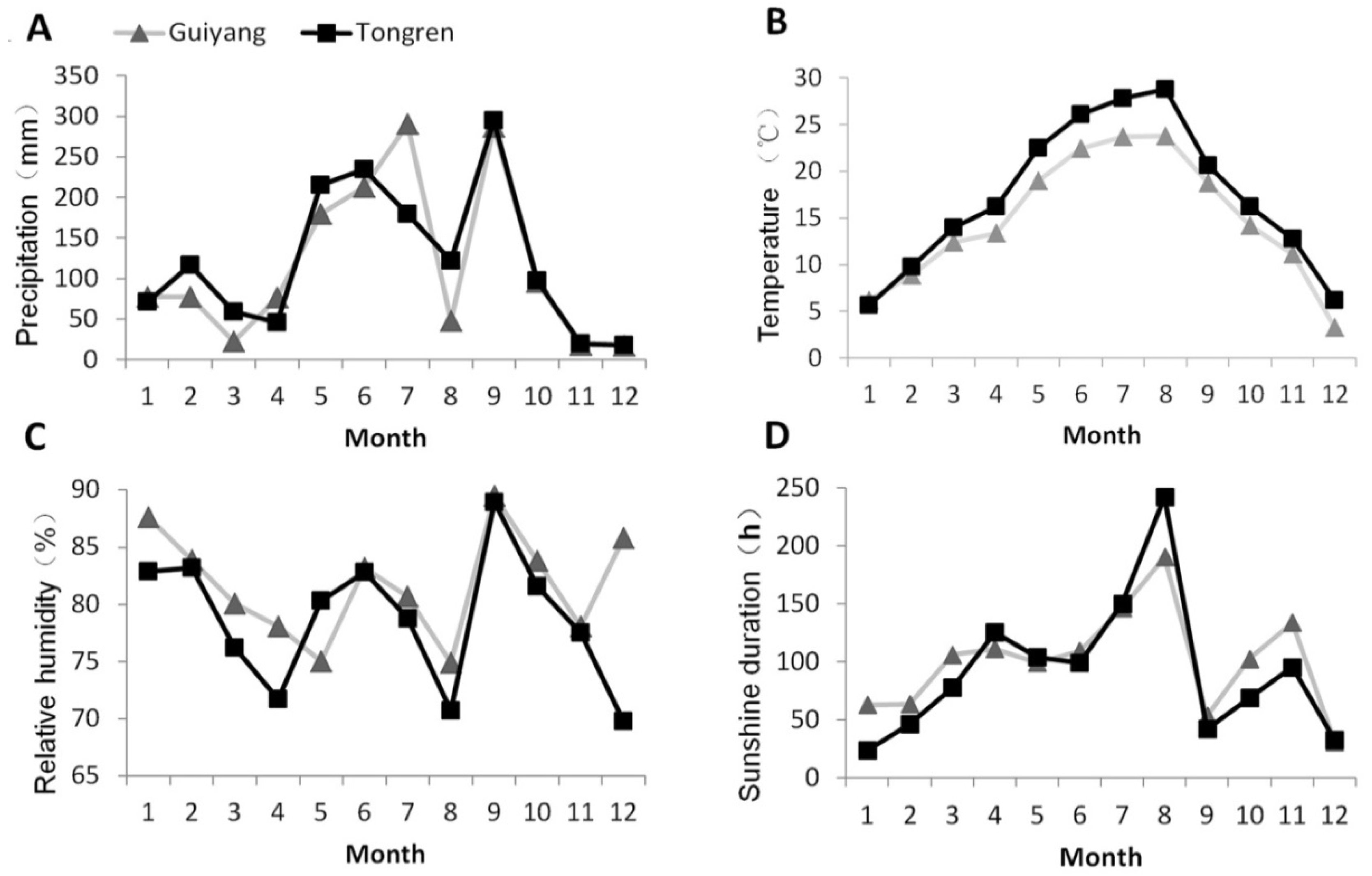
2.1. Study Site
2.2. Bacterial Community Collection
2.3. Microbiome Analysis
2.4. Statistical Analysis
3. Results
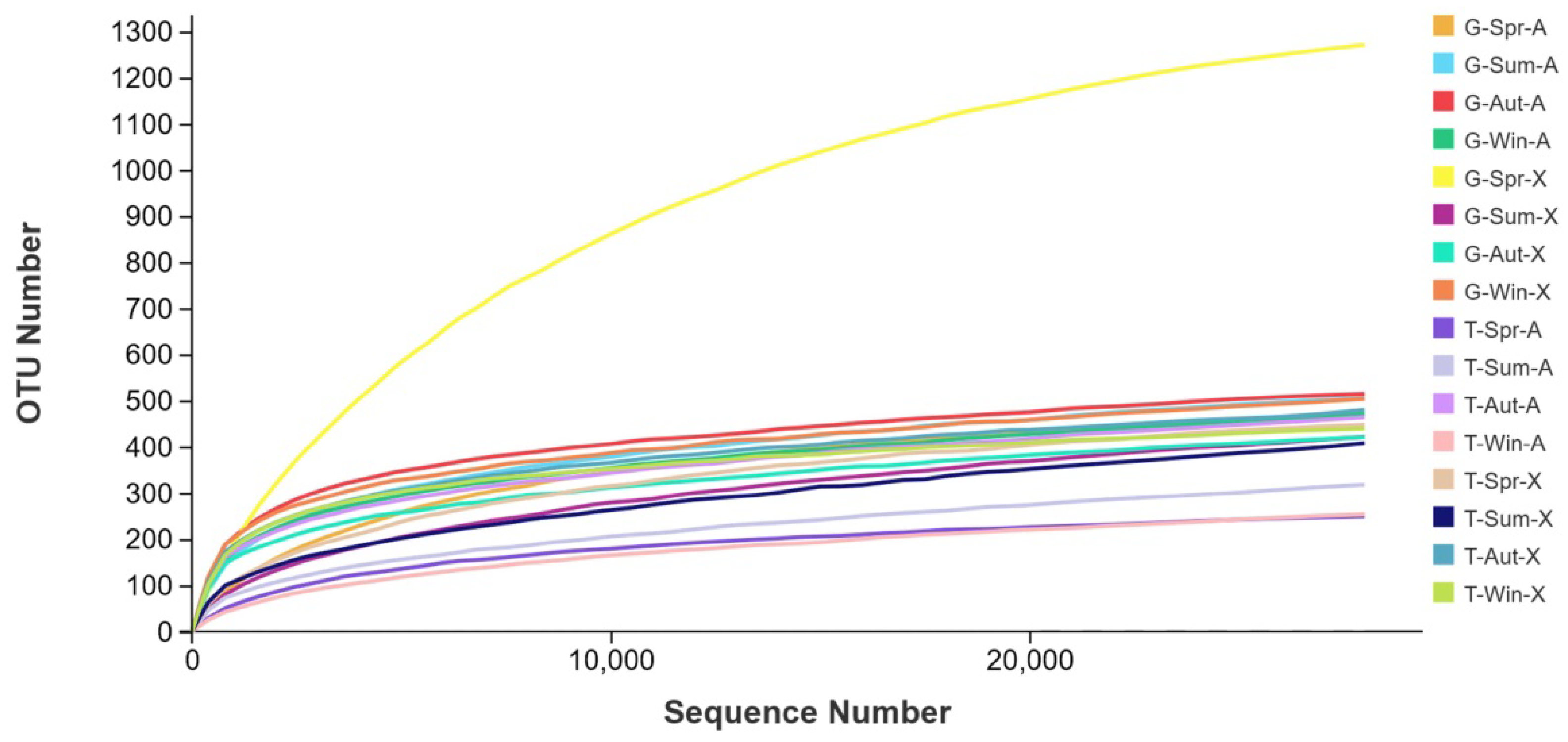
3.1. Phyllosphere Bacterial Diversity and Bacterial Community Dynamics
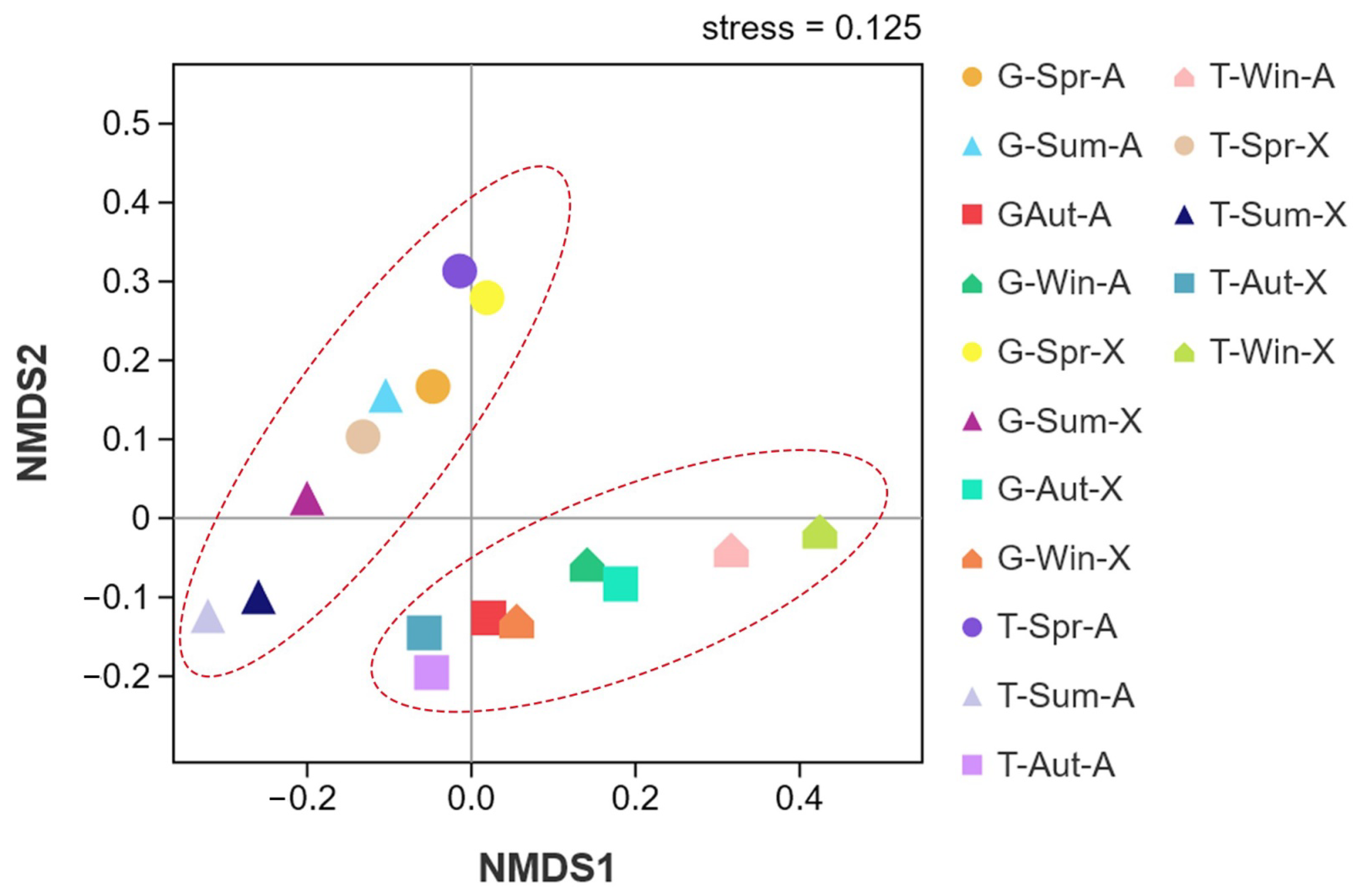
3.2. Drivers of Variation in Phyllosphere Bacterial Community Composition and Diversity
4. Discussion
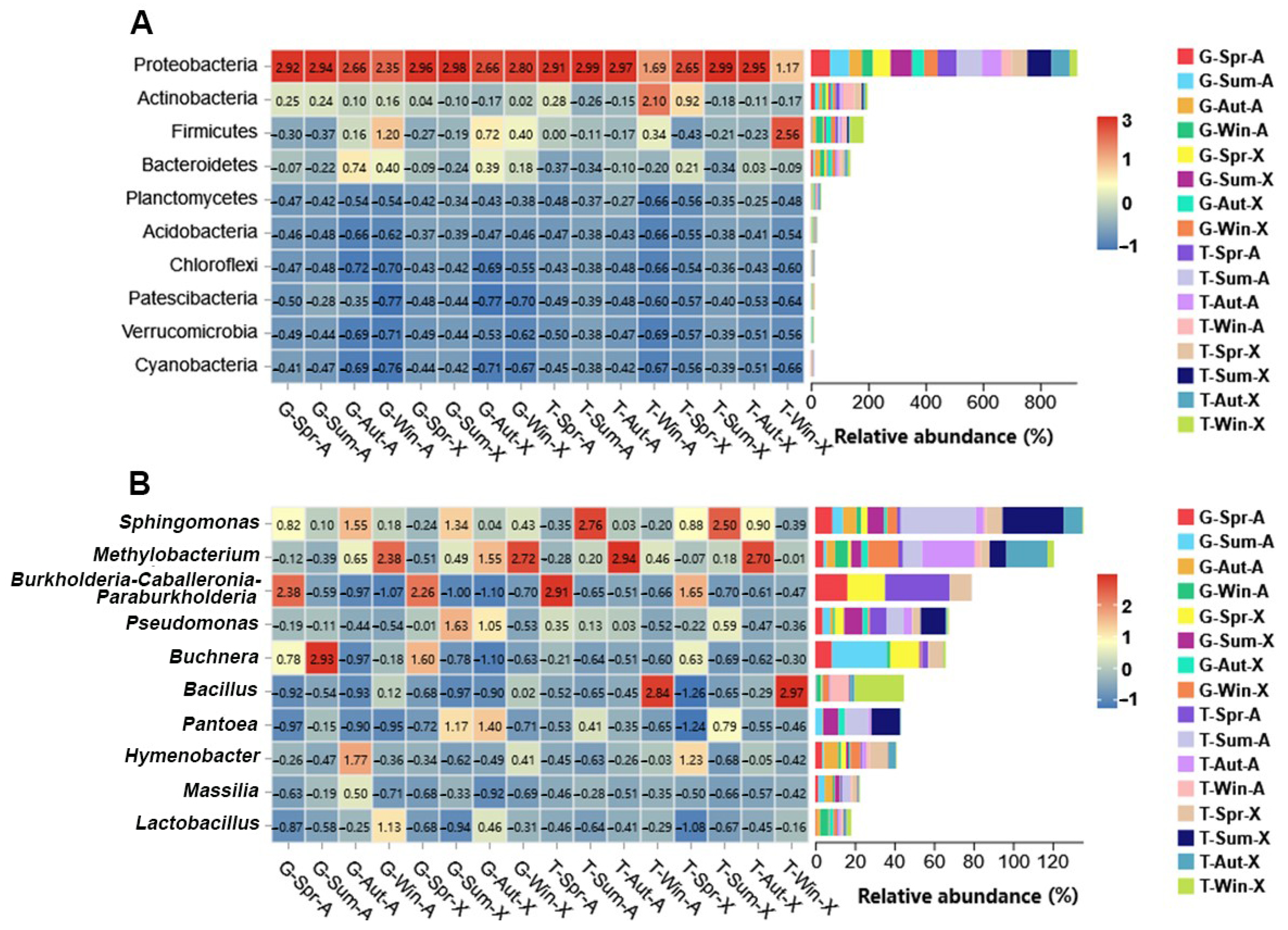
4.1. Analysis of Bacterial Community Composition and Diversity
4.2. Influence of Variety, Site, and Season on the Structure of Alfalfa Phyllosphere Bacterial Communities
4.3. Correlation Analysis between Bacterial Communities and Environmental Factors
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Area | pH | AN mg/kg | TN mg/kg | TK g/kg | TP g/kg | AK mg/kg | TOC g/kg | AP mg/kg | AMn mg/kg | AFe mg/kg | ACu mg/kg | AZn mg/kg |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Guiyang | 5.84 | 33.23 | 655 | 26.13 | 0.76 | 293.33 | 5.68 | 4.74 | 9.68 | 34.77 | 1.04 | 1.34 |
| Tongren | 6.14 | 11.55 | 847.72 | 16.15 | 0.73 | 184.62 | 14.07 | 0.18 | 174.99 | 75.81 | 3.48 | 1.21 |
References
- Knief, C.; Frances, L.; Vorholt, J.A. Competitiveness of Diverse Methylobacterium Strains in the Phyllosphere of Arabidopsis thaliana and Identification of Representative Models, Including M. extorquens PA1. Microb. Ecol. 2010, 60, 440–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martirosyan, V.; Unc, A.; Miller, G.; Doniger, T.; Wachtel, C.; Steinberger, Y. Desert Perennial Shrubs Shape the Microbial-Community Miscellany in Laimosphere and Phyllosphere Space. Microb. Ecol. 2016, 72, 659–668. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Guttman, D.S.; McHardy, A.C.; Schulze-Lefert, P. Microbial genome-enabled insights into plant-microorganism interactions. Nat. Rev. Genet. 2014, 15, 797–813. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.J.; Cai, W.Y.; Zhang, X.F.; Liu, J.H.; Chen, H.; Wei, Y.S.; Jia, X.X.; Bai, Z.H. Distinct Microbial Community of Phyllosphere Associated with Five Tropical Plants on Yongxing Island, South China Sea (vol 7, 525, 2019). Microorganisms 2020, 8, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, F.M.; Marina, M.; Pieckenstain, F.L. Novel components of leaf bacterial communities of field-grown tomato plants and their potential for plant growth promotion and biocontrol of tomato diseases. Res. Microbiol. 2016, 167, 222–233. [Google Scholar] [CrossRef]
- Devarajan, A.K.; Sabarinathan, K.G.; Gomathy, M.; Kannan, R.; Balachandar, D. Mitigation of drought stress in rice crop with plant growth promoting abiotic stress-tolerant rice phyllosphere bacteria. J. Basic Microbiol. 2020, 60, 768–786. [Google Scholar] [CrossRef]
- Carrión, O.; Gibson, L.; Elias, D.; Mcnamara, N.P.; Murrell, J.C. Diversity of isoprene-degrading bacteria in phyllosphere and soil communities from a high isoprene-emitting environment: A Malaysian oil palm plantation. Microbiome 2020, 8, 81. [Google Scholar] [CrossRef]
- Qza, C.; Wla, C.; Yan, L.B.; Mk, B.; Qian, Q.B.; Wy, B.; Xpa, B.; Hqa, B. Enantioselective effects of imazethapyr residues on Arabidopsis thaliana metabolic profile and phyllosphere microbial communities—ScienceDirect. J. Environ. Sci. 2020, 93, 57–65. [Google Scholar] [CrossRef]
- Aydogan, E.L.; Gerald, M.; Christoph, M.; Peter, K.; Glaeser, S.P. Long-Term Warming Shifts the Composition of Bacterial Communities in the Phyllosphere of Galium album in a Permanent Grassland Field-Experiment. Front. Microbiol. 2018, 9, 144. [Google Scholar] [CrossRef]
- Truchado, P.; Gil, M.I.; Moreno-Candel, M.; Allende, A. Impact of weather conditions, leaf age and irrigation water disinfection on the major epiphytic bacterial genera of baby spinach grown in an open field. Food Microbiol. 2019, 78, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Prasanna, R. Prospecting the characteristics and significance of the phyllosphere microbiome. Ann. Microbiol. 2018, 68, 229–245. [Google Scholar] [CrossRef]
- Tian, L.; Lin, X.L.; Tian, J.; Ji, L.; Chen, Y.L.; Tran, L.S.P.; Tian, C.J. Research Advances of Beneficial Microbiota Associated with Crop Plants. Int. J. Mol. Sci. 2020, 21, 1792. [Google Scholar] [CrossRef] [Green Version]
- Wagner, M.R.; Roberts, J.H.; Balint-Kurti, P.; Holland, J.B. Heterosis of leaf and rhizosphere microbiomes in field-grown maize. New Phytol. 2020, 228, 1055–1069. [Google Scholar] [CrossRef]
- Jackson, E.F.; Echlin, H.L.; Jackson, C.R. Changes in the phyllosphere community of the resurrection fern, Polypodium polypodioides, associated with rainfall and wetting. FEMS Microbiol. Ecol. 2006, 58, 236–246. [Google Scholar] [CrossRef] [Green Version]
- Kadivar, H.; Stapleton, A.E. Ultraviolet radiation alters maize phyllosphere bacterial diversity. Microb. Ecol. 2003, 45, 353–361. [Google Scholar] [CrossRef]
- Yang, C.H.; Crowley, D.E.; Borneman, J.; Keen, N.T. Microbial phyllosphere populations are more complex than previously realized. Proc. Natl. Acad. Sci. USA 2001, 98, 3889–3894. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, G.; Sbodio, A.; Tech, J.J.; Suslow, T.V.; Coaker, G.L.; Leveau, J.H.J. Leaf microbiota in an agroecosystem: Spatiotemporal variation in bacterial community composition on field-grown lettuce. ISEM J. 2012, 6, 1812–1822. [Google Scholar] [CrossRef]
- Stone, B.W.G.; Jackson, C.R. Biogeographic Patterns Between Bacterial Phyllosphere Communities of the Southern Magnolia (Magnolia grandiflora) in a Small Forest. Microb. Ecol. 2016, 71, 954–961. [Google Scholar] [CrossRef]
- Copeland, J.K.; Yuan, L.; Layeghifard, M.; Wang, P.W.; Guttman, D.S. Seasonal community succession of the phyllosphere microbiome. Mol. Plant-Microbe Interact. 2015, 28, 274–285. [Google Scholar] [CrossRef]
- Samapundo, S.; Devliehgere, F.; De Meulenaer, B.; Debevere, J. Effect of water activity and temperature on growth and the relationship between fumonisin production and the radial growth of Fusarium verticillioides and Fusarium proliferatum on corn. J. Food Prot. 2005, 68, 1054–1059. [Google Scholar] [CrossRef] [PubMed]
- Laforest-Lapointe, I.; Messier, C.; Kembel, S.W. Host species identity, site and time drive temperate tree phyllosphere bacterial community structure. Microbiome 2016, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, L.; Yang, J.; Jia, Y.; Hu, X.; Liu, Y.; Xu, H.; Yin, H.; Li, J.; Yi, Z. Effects of ecological environment and host genotype on the phyllosphere bacterial communities of cigar tobacco (Nicotiana tabacum L.). Ecol. Evol. 2021, 11, 10892–10903. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.J.; Gu, L.K.; Sun, B.; Cai, W.Y.; Zhang, S.W.; Zhuang, G.Q.; Bai, Z.H.; Zhuang, X.L. Seasonal variation of epiphytic bacteria in the phyllosphere of Gingko biloba, Pinus bungeana and Sabina chinensis. FEMS Microbiol. Ecol. 2020, 96, fiaa017. [Google Scholar] [CrossRef] [PubMed]
- Joung, Y.S.; Ge, Z.F.; Buie, C.R. Bioaerosol generation by raindrops on soil. Nat. Commun. 2017, 8, 14668. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.D.; Castillo, J.A. Influence of Light on Plant-Phyllosphere Interaction. Front. Plant Sci. 2018, 9, 1482. [Google Scholar] [CrossRef] [Green Version]
- Noble, A.S.; Noe, S.; Clearwater, M.J.; Lee, C.K. A core phyllosphere microbiome exists across distant populations of a tree species indigenous to New Zealand. PLoS ONE 2020, 15, e0237079. [Google Scholar] [CrossRef]
- Truchado, P.; Gil, M.I.; Reboleiro, P.; Rodelas, B.; Allende, A. Impact of solar radiation exposure on phyllosphere bacterial community of red-pigmented baby leaf lettuce. Food Microbiol. 2017, 66, 77–85. [Google Scholar] [CrossRef]
- Wallace, J.; Laforest-Lapointe, I.; Kembel, S.W. Variation in the leaf and root microbiome of sugar maple (Acer saccharum) at an elevational range limit. PeerJ 2018, 6, e5293. [Google Scholar] [CrossRef] [Green Version]
- Rao, Y.; Qian, Y.; She, X.; Yang, J.T.; He, P.H.; Jiang, Y.L.; Wang, M.; Xiang, W.L. Pellicle formation, microbial succession and lactic acid utilisation during the aerobic deteriorating process of Sichuan pickle. Int. J. Food Sci. Technol. 2018, 53, 767–775. [Google Scholar] [CrossRef]
- Avila, C.L.S.; Carvalho, B.F. Silage fermentation-updates focusing on the performance of micro-organisms. J. Appl. Microbiol. 2020, 128, 966–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, R.; Tian, Z. College of Animal Science, Guizhou University, Guiyang, Guizhou Province, China. 2022; manuscript in preparation. [Google Scholar]
- Hewitt, K.M.; Gerba, C.P.; Maxwell, S.L.; Kelley, S.T. Office space bacterial abundance and diversity in three metropolitan areas. PLoS ONE 2012, 7, e37849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.J.; Wu, F.H.; Hao, G.G.; Qi, Q.; Li, R.; Li, N.; Wei, L.M.; Chai, T.J. Bacillus subtilis Improves Immunity and Disease Resistance in Rabbits. Front. Immunol. 2017, 8, 354. [Google Scholar] [CrossRef] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Grady, K.L.; Sorensen, J.W.; Stopnisek, N.; Guittar, J.; Shade, A. Assembly and seasonality of core phyllosphere microbiota on perennial biofuel crops. Nat. Commun. 2019, 10, 4135. [Google Scholar] [CrossRef]
- Grobelak, A.; Hiller, J. Bacterial siderophores promote plant growth: Screening of catechol and hydroxamate siderophores. Int. J. Phytoremediat. 2017, 19, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Cernava, T.; Chen, X.Y.; Krug, L.; Li, H.X.; Yang, M.F.; Berg, G. The tea leaf microbiome shows specific responses to chemical pesticides and biocontrol applications. Sci. Total Environ. 2019, 667, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Prasanna, R.; Ranjan, K.; Velmourougane, K.; Ramakrishnan, B. Nutrients and host attributes modulate the abundance and functional traits of phyllosphere microbiome in rice. Microbiol. Res. 2017, 204, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Zhu, Y.G.; Wang, J.T.; Singh, B.; Han, L.L.; Shen, J.P.; Li, P.P.; Wang, G.B.; Wu, C.F.; Ge, A.H.; et al. Host selection shapes crop microbiome assembly and network complexity. New Phytol. 2021, 229, 1091–1104. [Google Scholar] [CrossRef]
- Muller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The Plant Microbiota: Systems-Level Insights and Perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Janakiev, T.; Dimkic, I.; Bojic, S.; Fira, D.; Stankovic, S.; Beric, T. Bacterial communities of plum phyllosphere and characterization of indigenous antagonistic Bacillus thuringiensis R3/3 isolate. J. Appl. Microbiol. 2020, 128, 528–543. [Google Scholar] [CrossRef]
- Toju, H.; Okayasu, K.; Notaguchi, M. Leaf-associated microbiomes of grafted tomato plants. Sci. Rep. 2019, 9, 1787. [Google Scholar] [CrossRef] [Green Version]
- McGarvey, J.A.; Franco, R.B.; Palumbo, J.D.; Hnasko, R.; Stanker, L.; Mitloehner, F.M. Bacterial population dynamics during the ensiling of Medicago sativa (alfalfa) and subsequent exposure to air. J. Appl. Microbiol. 2013, 114, 1661–1670. [Google Scholar] [CrossRef]
- Zheng, M.L.; Niu, D.Z.; Jiang, D.; Zuo, S.S.; Xu, C.C. Dynamics of microbial community during ensiling direct-cut alfalfa with and without LAB inoculant and sugar. J. Appl. Microbiol. 2017, 122, 1456–1470. [Google Scholar] [CrossRef]
- Andrews, J.H.; Harris, R.F. The Ecology and Biogeography of Microorganisms on Plant Surfaces. Annu. Rev. Phytopathol. 2000, 38, 145–180. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Kwon, S.W.; Kumutha, K.; Senthilkumar, M.; Ahmed, S.; Sa, T.; Anandham, R. Diversity of culturable methylotrophic bacteria in diffrent genotypes of groundnut and their potential for plant growth promotion. J. Biotechnol. 2018, 8, 275. [Google Scholar] [CrossRef]
- Meena, K.K.; Kumar, M.; Kalyuzhnaya, M.G.; Yandigeri, M.S.; Singh, D.P.; Saxena, A.K.; Arora, D.K. Epiphytic pink-pigmented methylotrophic bacteria enhance germination and seedling growth of wheat (Triticum aestivum) by producing phytohormone. Antonie van Leeuwenhoek 2012, 101, 777–786. [Google Scholar] [CrossRef]
- Wipat, A.; Harwood, C.R. The Bacillus subtilis genome sequence: The molecular blueprint of a soil bacterium. FEMS Microbiol. Ecol. 1999, 28, 1–9. [Google Scholar] [CrossRef]
- Sartori, M.; Nesci, A.; Formento, Á.; Etcheverry, M. Selection of potential biological control of Exserohilum turcicum with epiphytic microorganisms from maize. Rev. Argent. Microbiol. 2015, 47, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Innerebner, G.; Knief, C.; Vorholt, J.A. Protection of Arabidopsis thaliana against Leaf-Pathogenic Pseudomonas syringae by Sphingomonas Strains in a Controlled Model System. Appl. Environ. Microbiol. 2011, 77, 3202–3210. [Google Scholar] [CrossRef] [Green Version]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Thévenot-Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed. Sci. Technol. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, J.-B.; Kim, M.; Roh, E.; Jung, K.; Choi, M.; Oh, C.; Choi, J.; Yun, J.; Heu, S. Microbiota on spoiled vegetables and their characterization. J. Food Prot. 2013, 76, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Vega, H.; Arteaga-Garibay, R.I.; Maya-Lucas, O.; Gomez-Rodriguez, V.M.; Chavez-Diaz, I.F.; Ruvalcaba-Gomez, J.M.; Heredia-Nava, D.; Loperena-Martinez, R.; Zelaya-Molina, L.X. The Bacterial Community Associated with the Amarillo Zamorano Maize (Zea mays) Landrace Silage Process. Microorganisms 2020, 8, 1503. [Google Scholar] [CrossRef] [PubMed]
- Grube, M.; Schmid, F.; Berg, G. Black fungi and associated bacterial communities in the phyllosphere of grapevine. Fungal Biol. 2011, 115, 978–986. [Google Scholar] [CrossRef]
- Gu, L.K.; Bai, Z.H.; Jin, B.; Hu, Q.; Wang, H.L.; Zhuang, G.Q.; Zhang, H.X. Assessing the impact of fungicide enostroburin application on bacterial community in wheat phyllosphere. J. Environ. Sci. 2010, 22, 134–141. [Google Scholar] [CrossRef]
- Legein, M.; Smets, W.; Vandenheuvel, D.; Eilers, T.; Muyshondt, B.; Prinsen, E.; Samson, R.; Lebeer, S. Modes of Action of Microbial Biocontrol in the Phyllosphere. Front. Microbiol. 2020, 11, e1619. [Google Scholar] [CrossRef]
- Darlison, J.; Mogren, L.; Rosberg, A.K.; Gruden, M.; Minet, A.; Line, C.; Mieli, M.; Bengtsson, T.; Hakansson, A.; Uhlig, E.; et al. Leaf mineral content govern microbial community structure in the phyllosphere of spinach (Spinacia oleracea) and rocket (Diplotaxis tenuifolia). Sci. Total Environ. 2019, 675, 501–512. [Google Scholar] [CrossRef]
- Redford, A.J.; Fierer, N. Bacterial Succession on the Leaf Surface: A Novel System for Studying Successional Dynamics. Microb. Ecol. 2009, 58, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.J.; Hand, P.; Pink, D.; Whipps, J.M.; Bending, G.D. Both Leaf Properties and Microbe-Microbe Interactions Influence Within-Species Variation in Bacterial Population Diversity and Structure in the Lettuce (Lactuca Species) Phyllosphere. Appl. Environ. Microbiol. 2010, 76, 8117–8125. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Singh, D.; Lai-Hoe, A.; Rahim, R.A.; Ainuddin, A.N.; Chun, J.; Adams, J.M. Distinctive Phyllosphere Bacterial Communities in Tropical Trees. Microb. Ecol. 2012, 63, 674–681. [Google Scholar] [CrossRef]
- Chen, X.Y.L.; Wicaksono, W.A.; Berg, G.; Cernava, T. Bacterial communities in the plant phyllosphere harbour distinct responders to a broad-spectrum pesticide. Sci. Total Environ. 2021, 751, 141799. [Google Scholar] [CrossRef]
- Redford, A.J.; Bowers, R.M.; Knight, R.; Linhart, Y.; Fierer, N. The ecology of the phyllosphere: Geographic and phylogenetic variability in the distribution of bacteria on tree leaves. Environ. Microbiol. 2010, 12, 2885–2893. [Google Scholar] [CrossRef] [Green Version]
- Leff, J.W.; Del Tredici, P.; Friedman, W.E.; Fierer, N. Spatial structuring of bacterial communities within individual Ginkgo biloba trees. Environ. Microbiol. 2015, 17, 2352–2361. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef]
- Vacher, C.; Hampe, A.; Porté, A.J.; Sauer, U.; Compant, S.; Morris, C.E. The Phyllosphere: Microbial Jungle at the Plant–Climate Interface. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 1–24. [Google Scholar] [CrossRef]
- Rico, L.; Ogaya, R.; Terradas, J.; Peñuelas, J.; Papen, H. Community structures of N2-fixing bacteria associated with the phyllosphere of a Holm oak forest and their response to drought. Plant Biol. 2014, 16, 586–593. [Google Scholar] [CrossRef]
- Xu, J.; Kloepper, J.W.; Huang, P.; McInroy, J.A.; Hu, C.H. Isolation and characterization of N2-fixing bacteria from giant reed and switchgrass for plant growth promotion and nutrient uptake. J. Basic Microbiol. 2018, 58, 459–471. [Google Scholar] [CrossRef]
- Paul, K.; Saha, C.; Nag, M.; Mandal, D.; Naiya, H.; Sen, D.; Mitra, S.; Kumar, M.; Bose, D.; Mukherjee, G.; et al. A Tripartite Interaction among the Basidiomycete Rhodotorula mucilaginosa, N2-Fixing Endobacteria, and Rice Improves Plant Nitrogen Nutrition. Plant Cell 2019, 32, 486–507. [Google Scholar] [CrossRef] [PubMed]
- Kaymak, H.C.; Aksoy, A.; Kotan, R. Inoculation With N2-Fixing Plant Growth Promoting Rhizobacteria to Reduce Nitrogen Fertilizer Requirement of Lettuce. Acta Sci. Pol. Hortorum Cultus 2020, 19, 23–35. [Google Scholar] [CrossRef]
- Vissers, M.; Giffel, M.; Driehuis, F.; Jong, P.D.; Lankveld, J. Minimizing the level of Bacillus cereus spores in farm tank milk. J. Dairy Sci. 2007, 90, 3286–3293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayton, P.K. Biological diversity: The coexistence of species on changing landscapes. J. Exp. Mar. Biol. Ecol. 1995, 191, 127–128. [Google Scholar] [CrossRef]





| Site | Variety | Season | OTUs | Shannon | Chao | Ace | Coverage |
|---|---|---|---|---|---|---|---|
| Guiyang | Algonguin | Spring | 592 | 6.35 bcd | 563.03 ab | 549.21 b | 0.999 |
| Summer | 717 | 4.99 ab | 952.40 ab | 992.39 ab | 0.997 | ||
| Autumn | 740 | 6.77 a | 1005.85 ab | 1047.94 ab | 0.998 | ||
| Winter | 690 | 7.10 abc | 991.58 ab | 1015.89 ab | 0.998 | ||
| Xinjiang Daye | Spring | 1384 | 6.65 ab | 1353.79 a | 1399.40 a | 0.997 | |
| Summer | 692 | 5.10 bcd | 956.60 ab | 986.77 ab | 0.997 | ||
| Autumn | 637 | 6.88 ab | 913.76 ab | 948.01 ab | 0.998 | ||
| Winter | 734 | 6.72 a | 1077.28 ab | 1112.48 ab | 0.998 | ||
| Tongren | Algonguin | Spring | 277 | 5.18 cd | 292.41 b | 292.21 b | 0.998 |
| Summer | 552 | 4.14 abc | 917.46 ab | 914.48 ab | 0.998 | ||
| Autumn | 696 | 6.45 ab | 1022.10 ab | 1057.75 ab | 0.998 | ||
| Winter | 434 | 4.84 d | 628.20 ab | 650.52 ab | 0.998 | ||
| Xinjiang Daye | Spring | 595 | 5.66 abcd | 605.82 ab | 615.97 ab | 0.999 | |
| Summer | 682 | 4.72 abc | 964.86 ab | 986.18 ab | 0.997 | ||
| Autumn | 674 | 6.72 ab | 939.24 ab | 940.96 ab | 0.998 | ||
| Winter | 585 | 5.89 ab | 782.16 ab | 797.57 ab | 0.998 |
| Variables | Bray–Curtis Dissimilarities | ||
|---|---|---|---|
| R2 (%) | p (>F) | ||
| Single factor | Season | 27.6 | 0.001 *** |
| Site | 4.0 | 0.004 ** | |
| Variety | NS | NS | |
| Second-order interaction | Season * site | 42.9 | 0.001 *** |
| Season * variety | 34.7 | 0.001 *** | |
| Site * variety | NS | NS | |
| Third-order interaction | Season * site * variety | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Peng, C.; Sun, W.; Dong, R.; Hao, J. Effects of Variety, Plant Location, and Season on the Phyllosphere Bacterial Community Structure of Alfalfa (Medicago sativa L.). Microorganisms 2022, 10, 2023. https://doi.org/10.3390/microorganisms10102023
Zhang M, Peng C, Sun W, Dong R, Hao J. Effects of Variety, Plant Location, and Season on the Phyllosphere Bacterial Community Structure of Alfalfa (Medicago sativa L.). Microorganisms. 2022; 10(10):2023. https://doi.org/10.3390/microorganisms10102023
Chicago/Turabian StyleZhang, Mingzhu, Chao Peng, Wentao Sun, Rui Dong, and Jun Hao. 2022. "Effects of Variety, Plant Location, and Season on the Phyllosphere Bacterial Community Structure of Alfalfa (Medicago sativa L.)" Microorganisms 10, no. 10: 2023. https://doi.org/10.3390/microorganisms10102023