Correlation between Milk Bacteriology, Cytology and Mammary Tissue Histology in Cows: Cure from the Pathogen or Recovery from the Inflammation



Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals and Study Layout
4.2. Sample Collection and Analyses
4.2.1. Milk
4.2.2. Tissues
4.3. Bacteriology, Histology and PCR Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Auldist, M.J.; Coats, S.; Sutherland, B.J.; Mayes, J.J.; McDowell, G.H. Effects of somatic cell count and stage of lactation on raw milk composition and the yield and quality of Cheddar cheese. J. Dairy Res. 1996, 63, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Jaeggi, J.J.; Govindasamy-Lucey, S.; Berger, Y.M.; Johnson, M.E.; McKusick, B.C.; Thomas, D.L.; Wendorff, W.L. Hard ewe’s milk cheese manufactured from milk of three different groups of somatic cell counts. J. Dairy Sci. 2003, 86, 3082–3089. [Google Scholar] [CrossRef]
- Leitner, G.; Chaffer, M.; Shamay, A.; Shapiro, F.; Merin, U.; Ezra, E.; Saran, A.; Silanikove, N. Changes in milk composition as affected by subclinical mastitis in sheep. J. Dairy Sci. 2004, 87, 46–52. [Google Scholar] [CrossRef]
- Leitner, G.; Merin, U.; Silanikove, N. Changes in milk composition as affected by subclinical mastitis in goats. J. Dairy Sci. 2004, 87, 1719–1726. [Google Scholar] [CrossRef]
- Leitner, G.; Merin, U.; Silanikove, N. Effects of glandular bacterial infection and stage of lactation on milk quality: Comparison among cows, goats and sheep. Int. Dairy J. 2011, 21, 279–285. [Google Scholar] [CrossRef]
- Li, N.; Richoux, R.; Boutinaud, M.; Martin, P.; Gagnaire, V. Role of somatic cells on dairy processes and products: A review. Dairy Sci. Technol. 2014, 94, 517–538. [Google Scholar] [CrossRef] [Green Version]
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.; Hulland, C.; Ruegg, P.L. Characterization of clinical mastitis occurring in cows on 50 large dairy herds in Wisconsin. J. Dairy Sci. 2013, 96, 7538–7549. [Google Scholar] [CrossRef]
- Halasa, T.; Huijps, K.; Østerås, O.; Hogeveen, H. Economic effects of bovine mastitis and mastitis management: A review. Vet. Quart. 2007, 29, 18–31. [Google Scholar] [CrossRef]
- Gonçalves, J.L.; Kamphuis, C.; Martins, C.M.M.R.; Barreiro, J.R.; Tomazi, T.; Gameiro, A.H.; Hogeveen, H.; Dos Santos, M.V. Bovine subclinical mastitis reduces milk yield and economic return. Livest. Sci. 2018, 210, 25–32. [Google Scholar] [CrossRef]
- Schukken, Y.H.; Wilson, D.J.; Welcome, F.; Garrison-Tikofsky, L.; Gonzalez, R.N. Monitoring udder health and milk quality using somatic cell counts. Vet. Res. 2003, 34, 579–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitner, G.; Lavon, Y.; Matzrafi, Z.; Benun, O.; Bezman, D.; Merin, U. Pricing of cow’s milk in relation to bulk milk somatic cell count in the threshold range of 400 × 103 cells per milliliter. Isr. J. Vet. Med. 2016, 71, 10–15. [Google Scholar]
- Andersen, S.; Dohoo, I.R.; Olde Riekerink, R.; Stryhn, H. Diagnosing intramammary infections: Evaluating expert opinions on the definition of intramammary infection using conjoint analysis. J. Dairy Sci. 2010, 93, 2966–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, A.J.; Leach, K.A.; Breen, J.E.; Green, L.E.; Green, M.J. Survey on the incidence and aetiology of mastitis on dairy farms in England and Wales. Vet. Rec. 2007, 160, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Koskinen, M.T.; Wellenberg, G.J.; Sampimon, O.C.; Holopainen, J.; Rothkamp, A.; Salmikivi, L.; Van Haeringen, W.A.; Lam, T.J.; Pyörälä, S. Field comparison of real-time polymerase chain reaction and bacterial culture for identification of bovine mastitis bacteria. J. Dairy Sci. 2010, 93, 5707–5715. [Google Scholar] [CrossRef] [Green Version]
- Chaffer, M.; Leitner, G.; Winkler, M.; Glickman, A.; Krifucks, O.; Ezra, E.; Saran, A. Coagulase-negative Staphylococci and mammary gland infections in cows. J. Vet. Med. B 1999, 46, 707–712. [Google Scholar] [CrossRef]
- Leitner, G. Goagulase-negative Staphylococci species and the dairy—significant IMI pathogens. In Seminar on Coagulase-Negative Staphylococci in the Bovine; Ghent University: Ghent, Belgium, 2010; pp. 23–24. [Google Scholar]
- Rovai, M.; Caja, G.; Salama, A.A.K.; Jubert, A.; Lázaro, B.; Lázaro, M.; Leitner, G. Identifying the major bacteria causing intramammary infections in individual milk samples of sheep and goats using traditional bacteria culturing and Real-time Polymerase Chain Reaction. J. Dairy Sci. 2014, 97, 5393–5400. [Google Scholar] [CrossRef] [Green Version]
- Taponen, S.; Salmikivi, L.; Simojoki, H.; Koskinen, M.T.; Pyörälä, S. Real-time polymerase chain reaction-based identification of bacteria in milk samples from bovine clinical mastitis with no growth in conventional culturing. J. Dairy Sci. 2009, 92, 2610–2617. [Google Scholar] [CrossRef] [Green Version]
- Rovai, M.; Caja, G.; Quevedo, J.M.; Manuelian, C.L.; Saldo, J.; Salama, A.A.; Torre, P.; Arana, I.; Such, X.; Leitner, G. Effect of subclinical intramammary infection on milk quality in dairy sheep: II. Matured-pressed cheese (Manchego) produced from milk of uninfected and infected glands and from their blends. Small Rumin. Res. 2015, 126, 59–67. [Google Scholar] [CrossRef]
- Rovai, M.; Rusek, N.; Caja, G.; Saldo, J.; Leitner, G. Effect of subclinical intrammamay infection on milk quality in dairy sheep: I. Fresh-soft cheese produced from milk of uninfected and infected glands and from their blends. Small Rumin. Res. 2015, 125, 127–136. [Google Scholar] [CrossRef]
- Blum, S.E.; Heller, E.D.; Leitner, G. Long term effects of Escherichia coli mastitis. Vet. J. 2014, 201, 72–77. [Google Scholar] [PubMed]
- Lavon, Y.; Ezra, E.; Leitner, G.; Wolfenson, D. Association of conception rate with the pattern and level of somatic cell count elevation relative to time of insemination in dairy cows. J. Dairy Sci. 2011, 94, 4538–4545. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Leitner, G.; Lavon, Y. The disruptive effects of mastitis on reproduction and fertility in dairy cows. Ital. J. Anim. Sci. 2015, 14–4125, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Leitner, G.; Lavon, Y.; Matzrafi, Z.; Benun, O.; Bezman, D.; Merin, U. Somatic cell counts, chemical composition and coagulation properties of goat and sheep bulk tank milk. Int. Dairy J. 2016, 58, 9–13. [Google Scholar] [CrossRef]
- Merin, U.; Fleminger, G.; Komanovsky, J.; Silanikove, N.; Bernstein, S.; Leitner, G. Subclinical udder infection with Streptococcus dysgalactiae impair milk coagulation properties: Emerging role of proteose-peptones. Dairy Sci. Technol. 2008, 88, 407–419. [Google Scholar] [CrossRef]
- Blum, S.E.; Goldstone, R.J.; Connolly, J.P.R.; Répérant-Ferter, M.; Germon, P.; Inglis, N.F.; Krifucks, O.; Mathur, S.; Manson, E.; Mclean, K.; et al. Postgenomics characterization of an essential genetic determinant of mammary pathogenic Escherichia coli. mBio 2018, 9, e00423-18. [Google Scholar]
- Leitner, G.; Eligulashvily, R.; Krifucks, O.; Perl, S.; Saran, A. Immune cell differentiation in mammary gland tissues and milk of cows chronically infected with Staphylococcus aureus. J. Vet. Med. B 2003, 50, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Leitner, G.; Silanikove, N.; Jacobi, S.; Weisblit, L.; Bernstein, S.; Merin, U. The influence of milk storage time on the farm and in dairy silos on its quality for cheese production. Int. Dairy J. 2008, 18, 109–113. [Google Scholar] [CrossRef]
- Oliver, S.P.; Gonzalez, R.N.; Hogan, J.S.; Jayarao, B.M.; Owens, W.E. Microbiological Procedures for the Diagnosis of Bovine Udder Infection and Determination of Milk Quality, 4th ed.; The National Mastitis Council, Inc.: Verona, WI, USA, 2004. [Google Scholar]


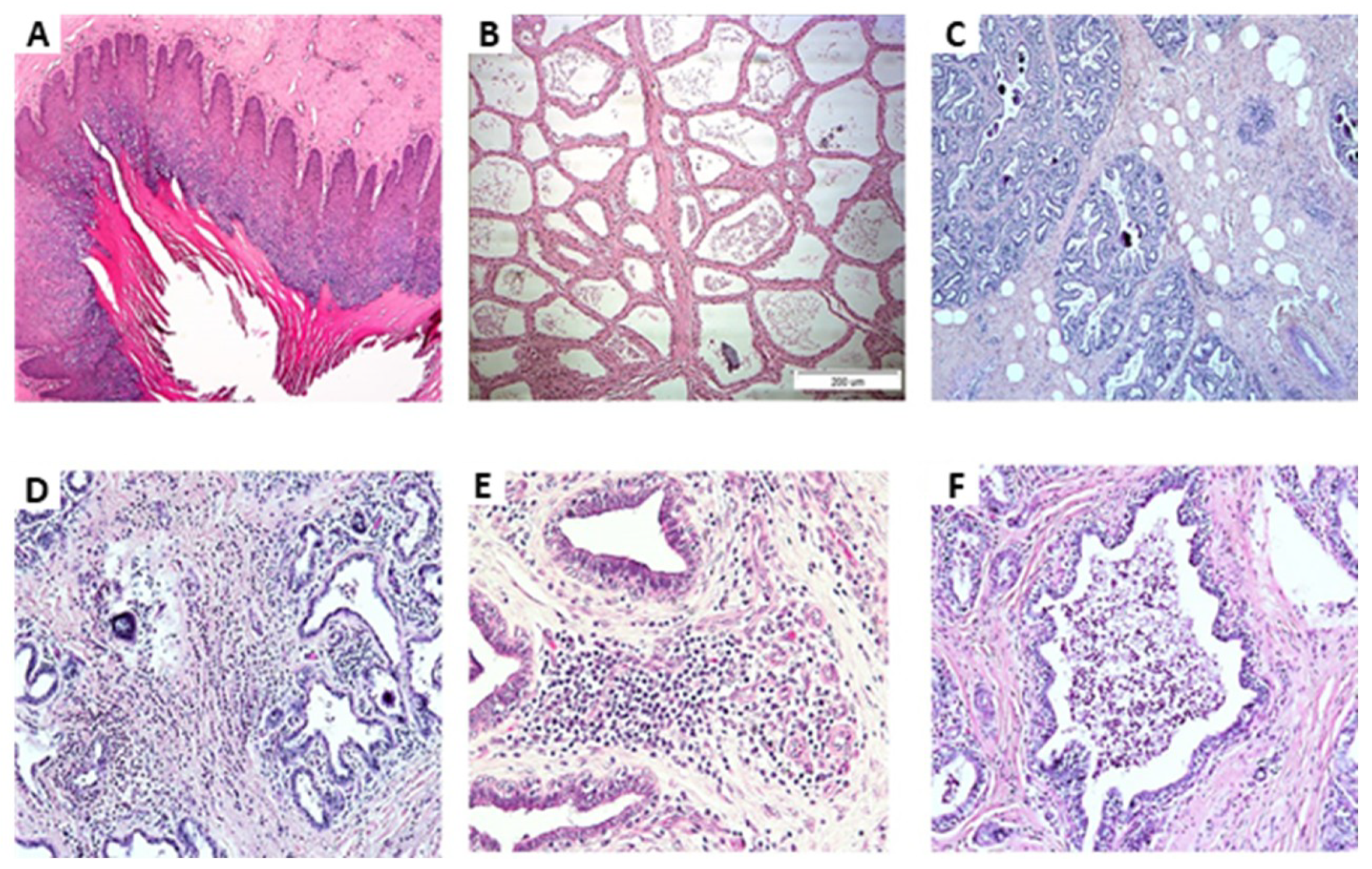{kind=link}
| Variable | No Inflammation and NBF (n = 33) | Inflammation and NBF (n = 26) | Inflammation and Isolation of CNS (n = 7) | Inflammation and Isolation of E. coli (n = 13) | Inflammation and Isolation of Strep. (n = 2) |
|---|---|---|---|---|---|
| SCC (× 103) | 185 ± 34 a | 1152 ± 140 b | 656 ± 175 ab | 1440 ± 677 b | 600 ± 2 ab |
| CD18+ (%) | 48.9 ± 3.7 a | 78.5 ± 2.5 b | 74.9 ± 4.5 b | 78.3 ± 2.5 b | 72.0 ± 8.7 b |
| PMN (%) | 25.9 ± 2.9 | 34.4 ± 4.3 | 36.9 ± 10.5 | 38.1 ± 6.1 | 15.5 ± 9.2 |
| CD14+ (%) | 14.3 ± 2.2 | 29.9 ± 4.1 | 18.3 ± 7.5 | 17.7 ± 4.0 | 30.0 ± 10.9 |
| CD4+ (%) | 3.1 ± 0.5 | 4.6 ± 0.8 | 4.4 ± 1.0 | 7.2 ± 1.2 | 5.0 ± 1.1 |
| CD8+ (%) | 4.9 ± 0.9 | 4.6 ± 1.2 | 5.2 ± 1.4 | 7.4 ± 1.3 | 3.5 ± 0.5 |
| Fat (g/L) | 33.9 ± 2.9 | 37.7 ± 2.7 | 36.3 ± 5.2 | 35.3 ± 3.3 | 46.9 ± 6.5 |
| Protein (g/L) | 40.0 ± 0.9 | 36.2 ± 1.1 | 38.3 ± 1.8 | 39.6 ± 0.9 | 30.4 ± 4.4 |
| % casein | 76.2 ± 0.3 | 71.9 ± 1.6 | 73.8 ± 1.0 | 73.8 ± 0.4 | 72.1 ± 2.5 |
| Lactose (g/L) | 47.7 ± 0.8 a | 33.2 ± 2.4 c | 43.4 ± 2.8 ab | 44.0 ± 1.0 ab | 38.7 ± 4.2 bc |
| RCT (sec) | 2429 ± 218 b | 4723 ± 156 a | 3157 ± 683 b | 4414 ± 299 a | >5000 a |
| CF (V) | 9.70 ± 1.0 a | 0.77 ± 0.27 c | 6.48 ± 2.44 ab | 2.38 ± 0.93 bc | 0 c |
| No. | Milk | Tissues | |||
|---|---|---|---|---|---|
| Gland’s class | Bacteriology | RT-CR | Edge of Nipple | Lobules | |
| No inflammation and NBF | 33 | 0 | 1 (Strep.) | 4 (3-Strep., 1-CNS) | 3 (CNS) |
| Inflammation | 26 | 0 | 1 (E. coli) | 9 (CNS) | 4 (CNS) |
| Inflammation and isolation of CNS | 7 | 7 | 5 (CNS) | 5 (CNS) | 5 (CNS) |
| Inflammation and isolation of E. coli | 3 | 3 | 1 (E. coli) | 1 (E. coli) | 2 (E. coli) |
| Inflammation and post isolation of E. coli | 10 | 0 | 0 | 1 (Strep.) | 1 (CNS) |
| Inflammation and isolation of Strep. | 2 | 2 | 1 (Strep.) | 1 (CNS) | 2 (CNS) |
| Gland’s Class | No. | Edge of the Nipple | Lobules |
|---|---|---|---|
| No inflammation and NBF | 33 | Normal | 50% normal production 50% connective tissue with increased fat globules |
| Inflammation | 26 | 75% Normal 25% Proliferation of blood vessels | 25% normal 75% Infiltrations of mononuclear cells and neutrophils in the connective tissue were observed but not in the lobules |
| Inflammation and isolation of CNS | 7 | Normal | 75% glands were identified with infiltration of mononuclear cells and neutrophils in the connective tissue but not in the lobules |
| Inflammation and isolation of E. coli | 3 | Normal | High number of neutrophils in the lobules |
| Inflammation and post-isolation of E. coli | 10 | Normal | Glands were identified with infiltration of mononuclear cells and neutrophils in the connective tissue but not in the lobules |
| Inflammation and isolation of Strep. | 2 | Normal | glands were identified with infiltration of mononuclear cells and neutrophils in the connective tissue but not in the lobules |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leitner, G.; Blum, S.E.; Krifuks, O.; Edery, N.; Merin, U. Correlation between Milk Bacteriology, Cytology and Mammary Tissue Histology in Cows: Cure from the Pathogen or Recovery from the Inflammation. Pathogens 2020, 9, 364. https://doi.org/10.3390/pathogens9050364
Leitner G, Blum SE, Krifuks O, Edery N, Merin U. Correlation between Milk Bacteriology, Cytology and Mammary Tissue Histology in Cows: Cure from the Pathogen or Recovery from the Inflammation. Pathogens. 2020; 9(5):364. https://doi.org/10.3390/pathogens9050364
Chicago/Turabian StyleLeitner, Gabriel, Shlomo E. Blum, Oloeg Krifuks, Nir Edery, and Uzi Merin. 2020. "Correlation between Milk Bacteriology, Cytology and Mammary Tissue Histology in Cows: Cure from the Pathogen or Recovery from the Inflammation" Pathogens 9, no. 5: 364. https://doi.org/10.3390/pathogens9050364