A Novel Bead-Based Immunoassay for the Measurement of Heat Shock Proteins 27 and 70



Abstract
:1. Introduction
2. Results
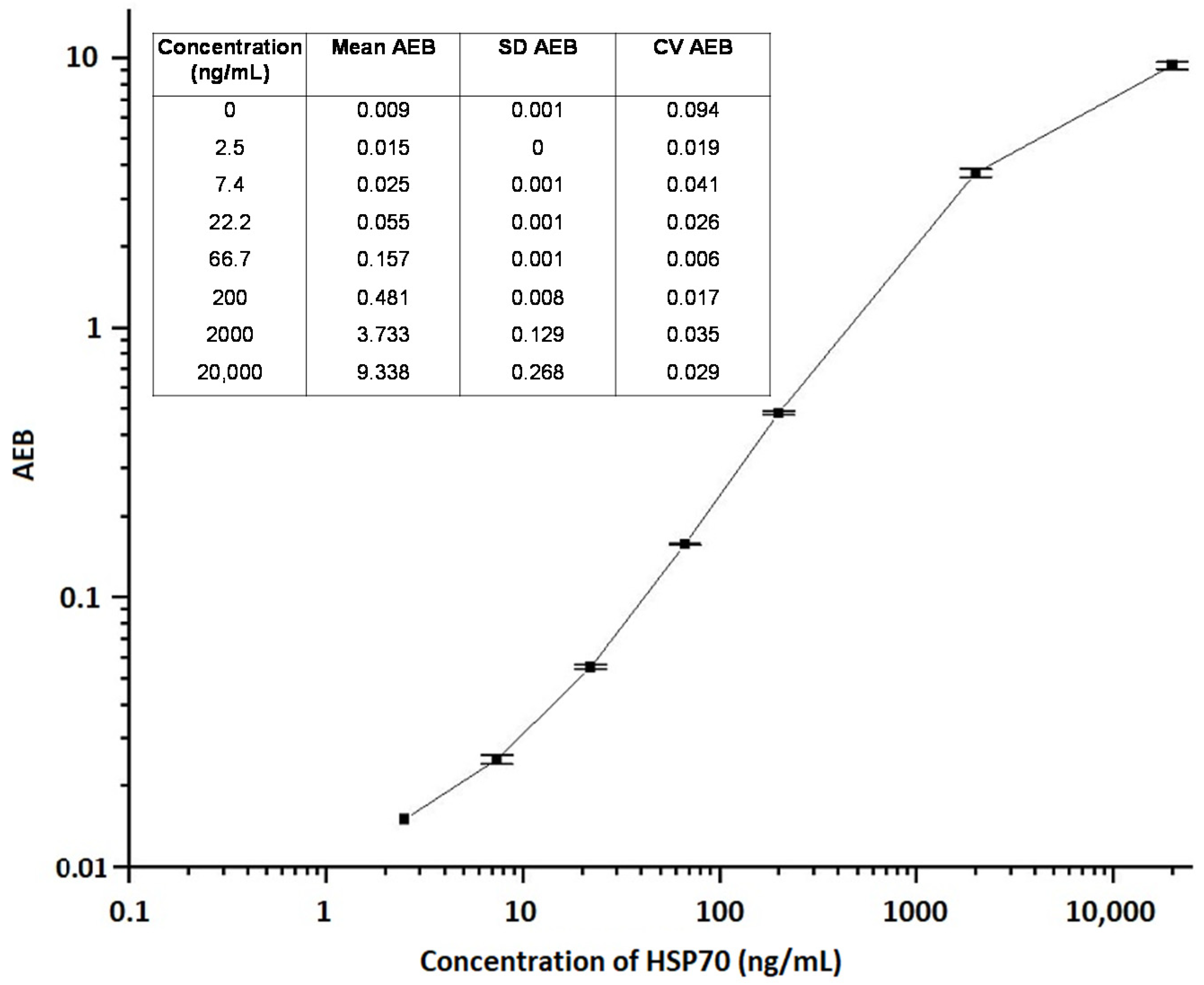
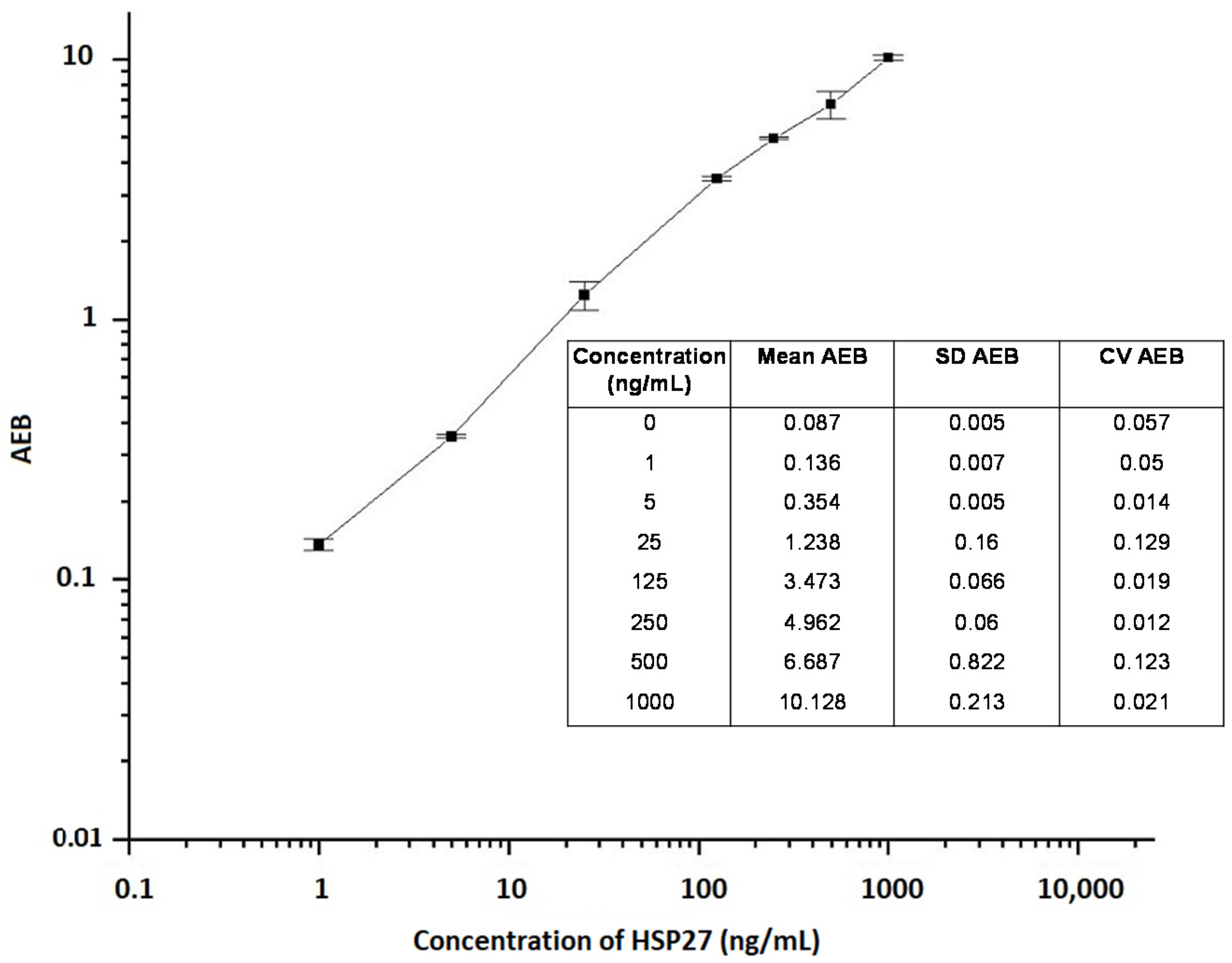
2.1. Assay Limit of Detection
2.2. Intra- and Inter-Assay Precision
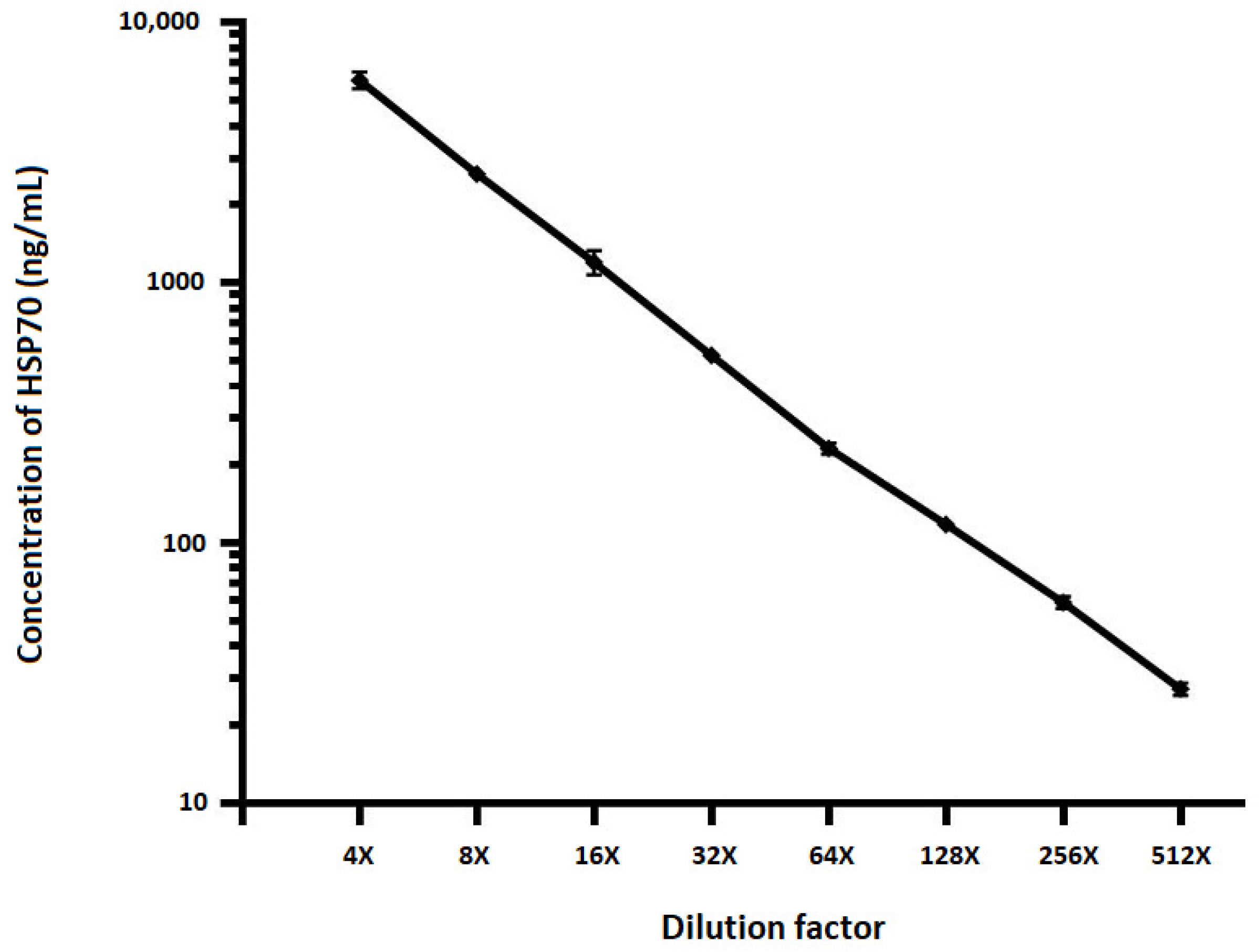
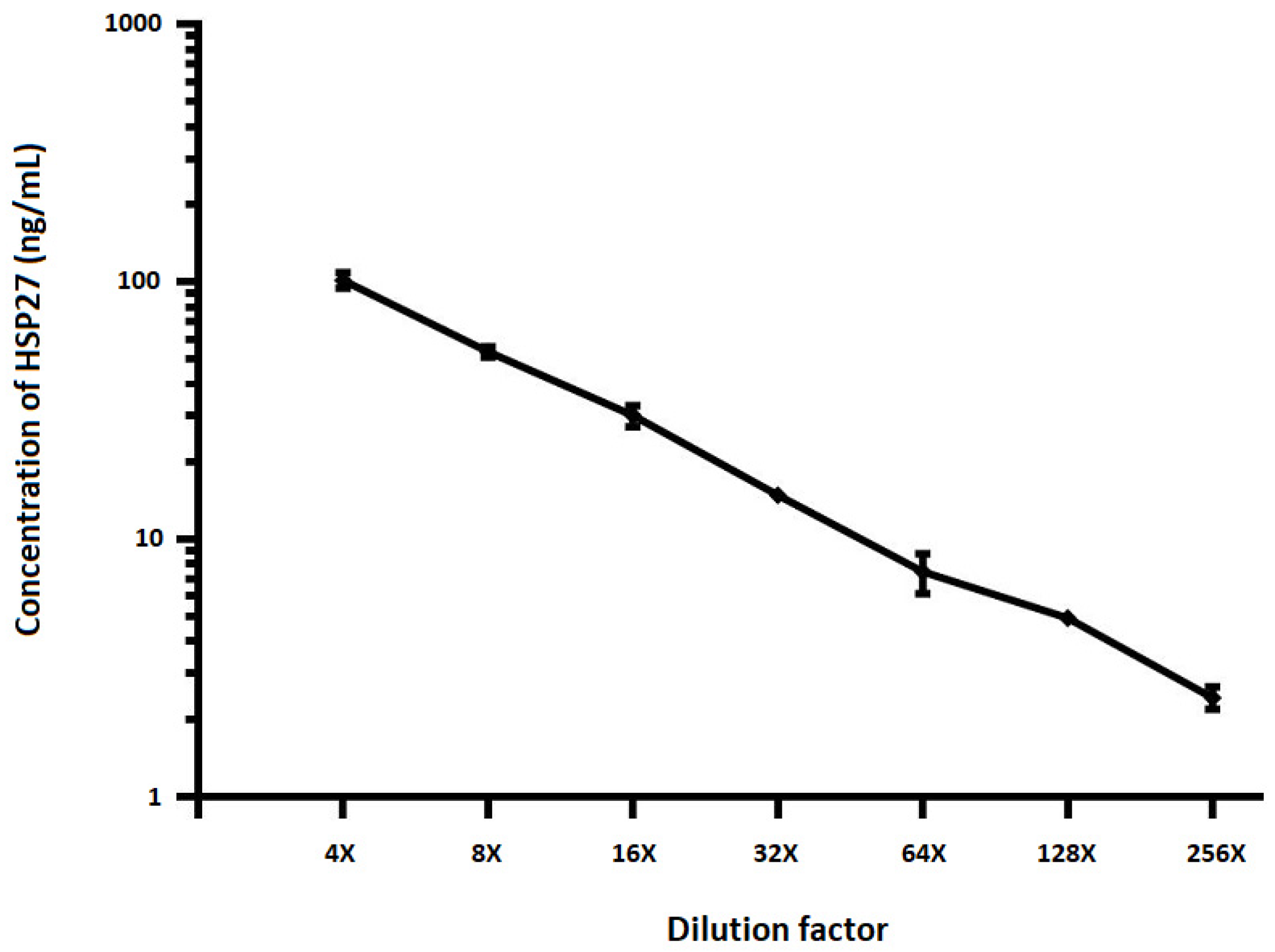
2.3. Dilutional Linearity
2.4. Admixture Linearity
2.5. Spike Recovery
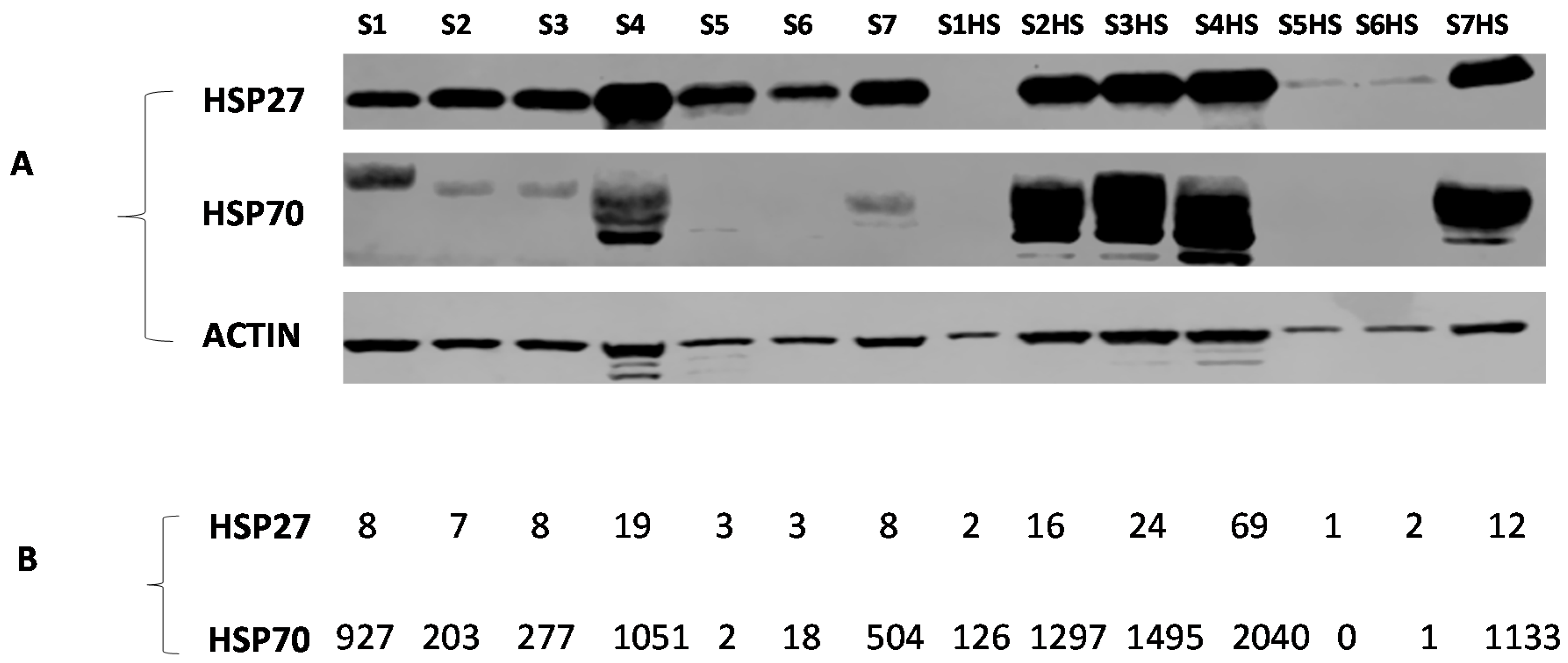
2.6. Cell Lysate Samples
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Detailed Procedures
4.2.1. Preparation of Paramagnetic Capture Beads
4.2.2. Biotinylation of Detection Antibody
4.2.3. Assay Validation Procedures
4.2.4. Cell Preparation
4.2.5. Preparation of Cell Lysates
4.2.6. SIMOA Assay for HSP Detection
4.2.7. Analysis of HSP by Western Blotting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Akerfelt, M.; Morimoto, R.I.; Sistonen, L. Heat shock factors: Integrators of cell stress, development and lifespan. Nat. Rev. Mol. Cell. Biol. 2010, 11, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Pinar, O.; Ozden, Y.A.; Omur, E.; Muhtesem, G. Heat Shock Proteins in Multiple Sclerosis. Adv. Exp. Med. Biol. 2017, 958, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vos, M.J.; Hageman, J.; Carra, S.; Kampinga, H.H. Structural and functional diversities between members of the human HSPB, HSPH, HSPA, and DNAJ chaperone families. Biochemistry 2008, 47, 7001–7011. [Google Scholar] [CrossRef] [PubMed]
- Kourtis, N.; Nikoletopoulou, V.; Tavernarakis, N. Small heat-shock proteins protect from heat-stroke-associated neurodegeneration. Nature 2012, 490, 213–218. [Google Scholar] [CrossRef]
- Morimoto, R.I.; Santoro, M.G. Stress-inducible responses and heat shock proteins: New pharmacologic targets for cytoprotection. Nat. Biotechnol. 1998, 16, 833–838. [Google Scholar] [CrossRef]
- Samali, A.; Robertson, J.D.; Peterson, E.; Manero, F.; van Zeijl, L.; Paul, C.; Cotgreave, I.A.; Arrigo, A.P.; Orrenius, S. Hsp27 protects mitochondria of thermotolerant cells against apoptotic stimuli. Cell Stress Chaperones 2001, 6, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wang, J.; Zhou, Y.; Wang, Y.; Wang, S.; Zhang, W. Hsp70 promotes chemoresistance by blocking Bax mitochondrial translocation in ovarian cancer cells. Cancer Lett. 2012, 321, 137–143. [Google Scholar] [CrossRef]
- Garrido, C.; Brunet, M.; Didelot, C.; Zermati, Y.; Schmitt, E.; Kroemer, G. Heat shock proteins 27 and 70: Anti-apoptotic proteins with tumorigenic properties. Cell Cycle 2006, 5, 2592–2601. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Lee, J.J.; Seo, J.S. HSP70 deficiency results in activation of c-Jun N-terminal Kinase, extracellular signal-regulated kinase, and caspase-3 in hyperosmolarity-induced apoptosis. J. Biol. Chem. 2005, 280, 6634–6641. [Google Scholar] [CrossRef] [Green Version]
- Sabirzhanov, B.; Stoica, B.A.; Hanscom, M.; Piao, C.S.; Faden, A.I. Over-expression of HSP70 attenuates caspase-dependent and caspase-independent pathways and inhibits neuronal apoptosis. J. Neurochem. 2012, 123, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Meena, A.S.; Bhat, M.K. Hyperthermia-associated carboplatin resistance: Differential role of p53, HSF1 and Hsp70 in hepatoma cells. Cancer Sci. 2010, 101, 1186–1193. [Google Scholar] [CrossRef] [PubMed]
- Ravagnan, L.; Gurbuxani, S.; Susin, S.A.; Maisse, C.; Daugas, E.; Zamzami, N.; Mak, T.; Jaattela, M.; Penninger, J.M.; Garrido, C.; et al. Heat-shock protein 70 antagonizes apoptosis-inducing factor. Nat. Cell Biol. 2001, 3, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Voss, O.H.; Batra, S.; Kolattukudy, S.J.; Gonzalez-Mejia, M.E.; Smith, J.B.; Doseff, A.I. Binding of caspase-3 prodomain to heat shock protein 27 regulates monocyte apoptosis by inhibiting caspase-3 proteolytic activation. J. Biol. Chem. 2007, 282, 25088–25099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charette, S.J.; Lavoie, J.N.; Lambert, H.; Landry, J. Inhibition of Daxx-mediated apoptosis by heat shock protein 27. Mol. Cell Biol. 2000, 20, 7602–7612. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.K.; De los Rios, P.; Christen, P.; Lustig, A.; Goloubinoff, P. The kinetic parameters and energy cost of the Hsp70 chaperone as a polypeptide unfoldase. Nat. Chem. Biol. 2010, 6, 914–920. [Google Scholar] [CrossRef]
- Grosicki, S.; Bednarczyk, M.; Janikowska, G. Heat shock proteins as a new, promising target of multiple myeloma therapy. Expert Rev. Hematol. 2020, 13, 117–126. [Google Scholar] [CrossRef]
- Yun, C.W.; Kim, H.J.; Lim, J.H.; Lee, S.H. Heat Shock Proteins: Agents of Cancer Development and Therapeutic Targets in Anti-Cancer Therapy. Cells 2019, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Tukaj, S.; Kaminski, M. Heat shock proteins in the therapy of autoimmune diseases: Too simple to be true? Cell Stress Chaperones 2019, 24, 475–479. [Google Scholar] [CrossRef] [Green Version]
- Leishman, S.J.; Ford, P.J.; West, M.J.; Cullinan, M.P.; Seymour, G.J. Heat shock proteins: A double-edged sword linking periodontal and cardiovascular diseases. Future Cardiol. 2017, 13, 515–519. [Google Scholar] [CrossRef]
- Mittal, S.; Rajala, M.S. Heat shock proteins as biomarkers of lung cancer. Cancer Biol. Ther. 2020, 21, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Ban, H.S.; Han, T.S.; Hur, K.; Cho, H.S. Epigenetic Alterations of Heat Shock Proteins (HSPs) in Cancer. Int. J. Mol. Sci. 2019, 20, 4758. [Google Scholar] [CrossRef] [Green Version]
- Charmpilas, N.; Kyriakakis, E.; Tavernarakis, N. Small heat shock proteins in ageing and age-related diseases. Cell Stress Chaperones 2017, 22, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, K.J.; Ford, J.L.; Brunk, U.T. Heat shock proteins: Keys to healthy ageing? Redox Rep. 2009, 14, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Njemini, R.; Bautmans, I.; Onyema, O.O.; Van Puyvelde, K.; Demanet, C.; Mets, T. Circulating heat shock protein 70 in health, aging and disease. BMC Immunol. 2011, 12, 24. [Google Scholar] [CrossRef] [Green Version]
- Njemini, R.; Lambert, M.; Demanet, C.; Kooijman, R.; Mets, T. Basal and infection-induced levels of heat shock proteins in human aging. Biogerontology 2007, 8, 353–364. [Google Scholar] [CrossRef]
- Wrońska, A.K.; Boguś, M.I. Heat shock proteins (HSP 90, 70, 60, and 27) in Galleria mellonella (Lepidoptera) hemolymph are affected by infection with Conidiobolus coronatus (Entomophthorales). PLoS ONE 2020, 15, e0228556. [Google Scholar] [CrossRef]
- Bolhassani, A.; Agi, E. Heat shock proteins in infection. Clin. Chim. Acta 2019, 498, 90–100. [Google Scholar] [CrossRef]
- Njemini, R.; Smitz, J.; Demanet, C.; Sosso, M.; Mets, T. Circulating heat shock protein 70 (Hsp70) in elderly members of a rural population from Cameroon: Association with infection and nutrition. Arch. Gerontol. Geriatr. 2011, 53, 359–363. [Google Scholar] [CrossRef] [Green Version]
- Njemini, R.; Bautmans, I.; Lambert, M.; Demanet, C.; Mets, T. Heat shock proteins and chemokine/cytokine secretion profile in ageing and inflammation. Mech. Ageing Dev. 2007, 128, 450–454. [Google Scholar] [CrossRef]
- Njemini, R.; Demanet, C.; Mets, T. Inflammatory status as an important determinant of heat shock protein 70 serum concentrations during aging. Biogerontology 2004, 5, 31–38. [Google Scholar] [CrossRef]
- Lopez-Martinez, G.; Elnitsky, M.A.; Benoit, J.B.; Lee, R.E., Jr.; Denlinger, D.L. High resistance to oxidative damage in the Antarctic midge Belgica antarctica, and developmentally linked expression of genes encoding superoxide dismutase, catalase and heat shock proteins. Insect Biochem. Mol. Biol. 2008, 38, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Zhao, H.; Wang, Y.; Liu, J.; Fei, D.; Yang, X.; Mu, M.; Xing, M. Elemental imbalance elicited by arsenic and copper exposures leads to oxidative stress and immunotoxicity in chicken gizzard, activating the protective effects of heat shock proteins. Environ. Sci. Pollut. Res. Int. 2019, 26, 36343–36353. [Google Scholar] [CrossRef]
- Sable, A.; Rai, K.M.; Choudhary, A.; Yadav, V.K.; Agarwal, S.K.; Sawant, S.V. Inhibition of Heat Shock proteins HSP90 and HSP70 induce oxidative stress, suppressing cotton fiber development. Sci. Rep. 2018, 8, 3620. [Google Scholar] [CrossRef]
- Ou, J.R.; Tan, M.S.; Xie, A.M.; Yu, J.T.; Tan, L. Heat shock protein 90 in Alzheimer's disease. Biomed. Res. Int. 2014, 2014, 796869. [Google Scholar] [CrossRef]
- Gezen-Ak, D.; Dursun, E.; Hanagasi, H.; Bilgic, B.; Lohman, E.; Araz, O.S.; Atasoy, I.L.; Alaylioglu, M.; Onal, B.; Gurvit, H.; et al. BDNF, TNFalpha, HSP90, CFH, and IL-10 serum levels in patients with early or late onset Alzheimer’s disease or mild cognitive impairment. J. Alzheimers Dis. 2013, 37, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Klucken, J.; Shin, Y.; Masliah, E.; Hyman, B.T.; McLean, P.J. Hsp70 Reduces alpha-Synuclein Aggregation and Toxicity. J. Biol. Chem. 2004, 279, 25497–25502. [Google Scholar] [CrossRef] [Green Version]
- Roodveldt, C.; Bertoncini, C.W.; Andersson, A.; van der Goot, A.T.; Hsu, S.T.; Fernandez-Montesinos, R.; de Jong, J.; van Ham, T.J.; Nollen, E.A.; Pozo, D.; et al. Chaperone proteostasis in Parkinson’s disease: Stabilization of the Hsp70/alpha-synuclein complex by Hip. EMBO J. 2009, 28, 3758–3770. [Google Scholar] [CrossRef] [Green Version]
- Iwaki, T.; Iwaki, A.; Tateishi, J.; Sakaki, Y.; Goldman, J.E. Alpha B-crystallin and 27-kd heat shock protein are regulated by stress conditions in the central nervous system and accumulate in Rosenthal fibers. Am. J. Pathol. 1993, 143, 487–495. [Google Scholar] [PubMed]
- Alexiou, G.A.; Karamoutsios, A.; Lallas, G.; Ragos, V.; Goussia, A.; Kyritsis, A.P.; Voulgaris, S.; Vartholomatos, G. Expression of heat shock proteins in brain tumors. Turk. Neurosurg. 2014, 24, 745–749. [Google Scholar] [CrossRef] [Green Version]
- Njemini, R.; Lambert, M.; Demanet, C.; Vanden Abeele, M.; Vandebosch, S.; Mets, T. The induction of heat shock protein 70 in peripheral mononuclear blood cells in elderly patients: A role for inflammatory markers. Hum. Immunol. 2003, 64, 575–585. [Google Scholar] [CrossRef]
- Njemini, R.; Lambert, M.; Demanet, C.; Mets, T. Heat shock protein 32 in human peripheral blood mononuclear cells: Effect of aging and inflammation. J. Clin. Immunol. 2005, 25, 405–417. [Google Scholar] [CrossRef]
- Rea, I.M.; McNerlan, S.; Pockley, A.G. Serum heat shock protein and anti-heat shock protein antibody levels in aging. Exp. Gerontol. 2001, 36, 341–352. [Google Scholar] [CrossRef]
- Njemini, R.; Abeele, M.V.; Demanet, C.; Lambert, M.; Vandebosch, S.; Mets, T. Age-Related Decrease in the Inducibility of Heat-Shock Protein 70 in Human Peripheral Blood Mononuclear Cells. J. Clin. Immunol. 2002, 22, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Njemini, R.; Demanet, C.; Mets, T. Aging-related differences in basal heat shock protein 70 levels in lymphocytes are linked to altered frequencies of lymphocyte subsets. Aging Cell 2008, 7, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Welch, W.J. Heat shock proteins functioning as molecular chaperones: Their roles in normal and stressed cells. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1993, 339, 327–333. [Google Scholar] [CrossRef]
- Lachno, D.R.; Emerson, J.K.; Vanderstichele, H.; Gonzales, C.; Martenyi, F.; Konrad, R.J.; Talbot, J.A.; Lowe, S.L.; Oefinger, P.E.; Dean, R.A. Validation of a multiplex assay for simultaneous quantification of amyloid-beta peptide species in human plasma with utility for measurements in studies of Alzheimer’s disease therapeutics. J. Alzheimers Dis. 2012, 32, 905–918. [Google Scholar] [CrossRef]
- Okereke, O.I.; Xia, W.; Irizarry, M.C.; Sun, X.; Qiu, W.Q.; Fagan, A.M.; Mehta, P.D.; Hyman, B.T.; Selkoe, D.J.; Grodstein, F. Performance characteristics of plasma amyloid-beta 40 and 42 assays. J. Alzheimers Dis. 2009, 16, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Lachno, D.R.; Hanlon, D.; Shepro, A.; Jeromin, A.; Gemani, D.; Talbot, J.A.; Racke, M.M.; Dage, J.L.; Dean, R.A. A digital enzyme-linked immunosorbent assay for ultrasensitive measurement of amyloid-beta 1–42 peptide in human plasma with utility for studies of Alzheimer’s disease therapeutics. Alzheimer Res. Ther. 2016, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Kocsis, J.; Prohaszka, Z.; Biro, A.; Fust, G.; Banhegyi, D. Elevated levels of antibodies against 70 kDa heat shock proteins in the sera of patients with HIV infection. J. Med. Virol. 2003, 71, 480–482. [Google Scholar] [CrossRef]
- Hayem, G.; De Bandt, M.; Palazzo, E.; Roux, S.; Combe, B.; Eliaou, J.F.; Sany, J.; Kahn, M.F.; Meyer, O. Anti-heat shock protein 70 kDa and 90 kDa antibodies in serum of patients with rheumatoid arthritis. Ann. Rheum. Dis. 1999, 58, 291–296. [Google Scholar] [CrossRef]
- Avrameas, S.; Dighiero, G.; Lymberi, P.; Guilbert, B. Studies on natural antibodies and autoantibodies. Ann. Immunol. 1983, 134, 103–113. [Google Scholar]
- Njemini, R.; Demanet, C.; Met, T. Determination of intracellular heat shock protein 70 using a newly developed cell lysate immunometric assay. J. Immunol. Methods 2003, 274, 271–279. [Google Scholar]
- Costa, O.R.; Verhaeghen, K.; Roels, S.; Stange, G.; Ling, Z.; Pipeleers, D.; Gorus, F.K.; Martens, G.A. An analytical comparison of three immunoassay platforms for subpicomolar detection of protein biomarker GAD65. PLoS ONE 2018, 13, e0193670. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HSP70 Concentration (ng/mL) | |||||||
|---|---|---|---|---|---|---|---|
| Theoretical Concentration | 20,000 | 2000 | 200 | 66.7 | 22.2 | 7.4 | 2.5 |
| Assay 1 | 18,416 | 2215 | 185 | 63.6 | 16.9 | 7.2 | 2.0 |
| Assay 2 | 19,979 | 1862 | 158 | 47.5 | 18.4 | 6.6 | 3.3 |
| Assay 3 | 19,229 | 3034 | 205 | 73.1 | 18.8 | 7.6 | 2.4 |
| Assay 4 | 21,281 | 1932 | 200 | 68.4 | 23.1 | 7.3 | 2.3 |
| Assay 5 | 16,617 | 1903 | 217 | 70.7 | 21.7 | 8.4 | 1.3 |
| Assay 6 | 17,793 | 2553 | 195 | 56.1 | 19.3 | 5.8 | 3.0 |
| Assay 7 | 17,431 | 1981 | 199 | 54.7 | 20.8 | 6.6 | 1.9 |
| Assay 8 | 18,432 | 2533 | 211 | 63.7 | 24.4 | 8.4 | 1.5 |
| Inter-assay means | 18,647 | 2251 | 196 | 62.2 | 20.4 | 7.2 | 2.2 |
| SD | 1487 | 419 | 18.3 | 8.8 | 2.5 | 0.9 | 0.7 |
| CV% | 8.0 | 18.6 | 9.3 | 14.2 | 12.4 | 12.5 | 31.2 |
| HSP27 Concentration (ng/mL) | |||||||
|---|---|---|---|---|---|---|---|
| Theoretical Concentration | 1000 | 500 | 250 | 125 | 25 | 5 | 1 |
| Assay 1 | 1107 | 531 | 260 | 145 | 44 | 6.4 | 1.3 |
| Assay 2 | 1107 | 515 | 265 | 134 | 31 | 5.2 | 1.2 |
| Assay 3 | 1093 | 469 | 252 | 147 | 29 | 5.5 | 1.2 |
| Assay 4 | 1058 | 549 | 277 | 144 | 28 | 5.1 | 0.9 |
| Assay 5 | 1083 | 484 | 262 | 143 | 27 | 5.4 | 1.1 |
| Assay 6 | 1001 | 429 | 261 | 148 | 29 | 5.5 | 1.1 |
| Assay 7 | 1074 | 584 | 264 | 125 | 27 | 5.1 | 0.9 |
| Inter-assay means | 1075 | 509 | 263 | 141 | 31 | 5.5 | 1.1 |
| SD | 37 | 52 | 7.5 | 8.4 | 6.0 | 0.5 | 0.2 |
| CV% | 3.4 | 10 | 2.8 | 5.9 | 20 | 8.3 | 14 |
| Parameter | HSP70 Concentration (ng/mL) a | |||
|---|---|---|---|---|
| High Control | Low Control | High Panel | Low Panel | |
| Intra-assay means (n = 6) | ||||
| Assay 1 | 2359 | 88 | 1884 | 66 |
| Assay 2 | 2174 | 80 | 1627 | 55 |
| Assay 3 | 2435 | 57 | 1434 | 42 |
| Assay 4 | 2061 | 77 | 2014 | 55 |
| Intra-assay CV% | 1 | 3 | 4 | 4 |
| 7 | 9 | 7 | 16 | |
| 4 | 4 | 6 | 8 | |
| 8 | 4 | 1 | 18 | |
| Inter-assay means (n = 4) | 2257 | 75 | 1740 | 55 |
| Inter-assay CV% | 8 | 18 | 15 | 18 |
| Parameter | High (2000 ng/mL) | Low (66.7 ng/mL) | ||
|---|---|---|---|---|
| Measured Concentration (ng/mL) | % Recovery | Measured Concentration (ng/mL) | % Recovery | |
| Intra-assay means (n = 3) | ||||
| Sample 1 | 1560 | 78 | 48 | 72 |
| Sample 2 | 1138 | 57 | 44 | 66 |
| Sample 3 | 1892 | 95 | 60 | 90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Njemini, R.; Verhaeghen, K.; Mets, T.; Weets, I.; Bautmans, I. A Novel Bead-Based Immunoassay for the Measurement of Heat Shock Proteins 27 and 70. Pathogens 2020, 9, 863. https://doi.org/10.3390/pathogens9110863
Njemini R, Verhaeghen K, Mets T, Weets I, Bautmans I. A Novel Bead-Based Immunoassay for the Measurement of Heat Shock Proteins 27 and 70. Pathogens. 2020; 9(11):863. https://doi.org/10.3390/pathogens9110863
Chicago/Turabian StyleNjemini, Rose, Katrijn Verhaeghen, Tony Mets, Ilse Weets, and Ivan Bautmans. 2020. "A Novel Bead-Based Immunoassay for the Measurement of Heat Shock Proteins 27 and 70" Pathogens 9, no. 11: 863. https://doi.org/10.3390/pathogens9110863