
Infectiousness of Asymptomatic Meriones shawi, Reservoir Host of Leishmania major
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sand Flies, Parasites and Rodents
2.2. Experimental Infections of Sand Flies
2.3. Experimental Infections of M. shawi
2.4. Xenodiagnoses
2.5. Tissue Sampling and Leishmania Detection Post-Mortem by qPCR
2.6. Statistical Analysis
2.7. Animal Experimentation Guidelines
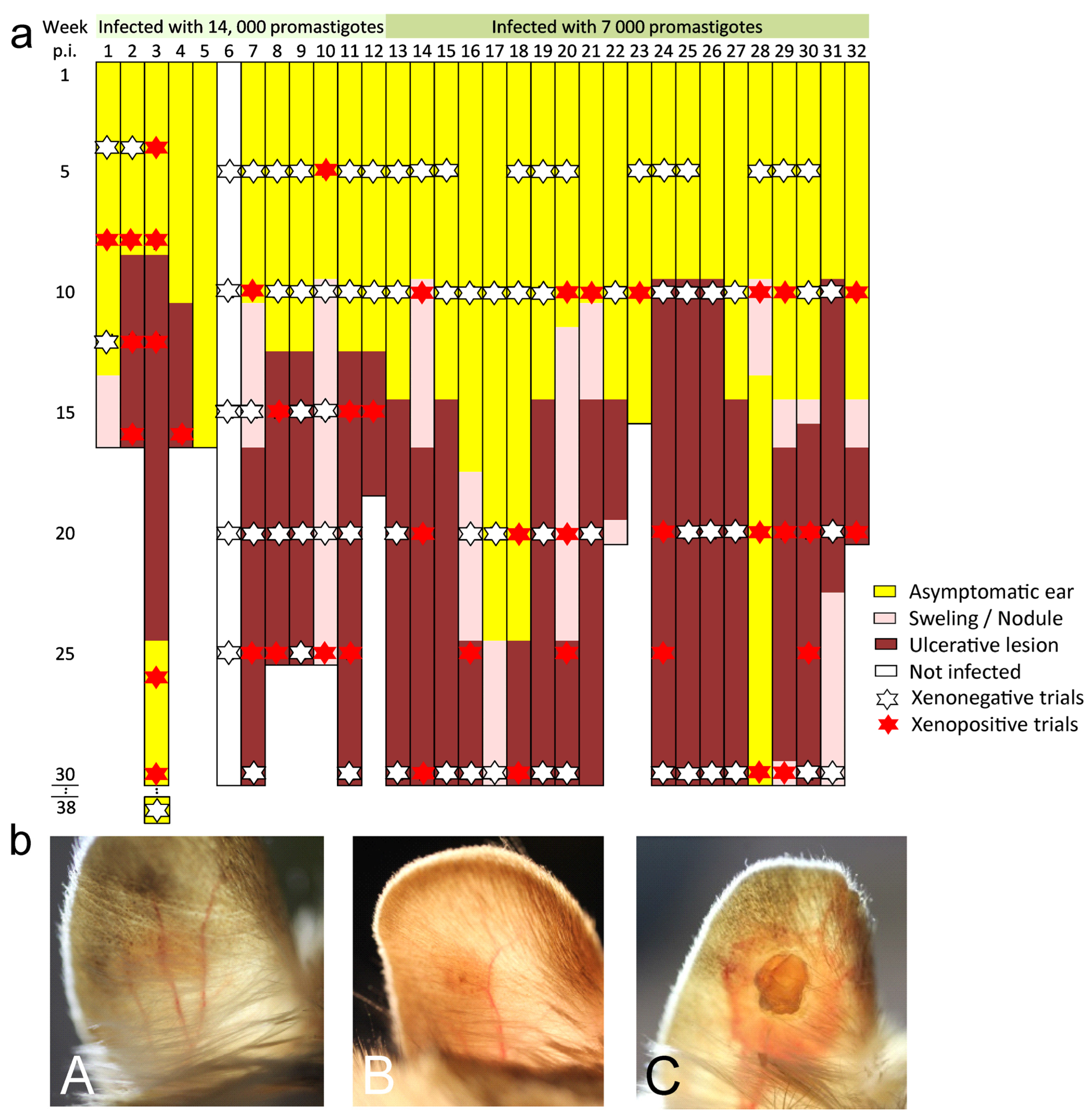
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Available online: https://www.who.int/health-topics/leishmaniasis (accessed on 11 April 2023).
- Yurchenko, V.; Chistyakov, D.S.; Akhmadishina, L.V.; Lukashev, A.N.; Sádlová, J.; Strelkova, M.V. Revisiting epidemiology of leishmaniasis in Central Asia: Lessons learnt. Parasitology 2023, 150, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Aoun, K.; Bouratbine, A. Cutaneous Leishmaniasis in North Africa: A review. Parasite 2014, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Ashford, R.W. The leishmaniases as emerging and reemerging zoonoses. Int. J. Parasitol. 2000, 30, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Sadlova, J.; Vojtkova, B.; Hrncirova, K.; Lestinova, T.; Spitzova, T.; Becvar, T.; Votypka, J.; Bates, P.; Volf, P. Host competence of African rodents Arvicanthis neumanni, A. niloticus and Mastomys natalensis for Leishmania major. Int. J. Parasitol. Parasites Wildl. 2019, 8, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Ghawar, W.; Toumi, A.; Snoussi, M.-A.; Chlif, S.; Zaatour, A.; Boukthir, A.; Hamida, N.B.H.; Chernkhi, J.; Diouani, M.F.; Ben-Saleh, A. Leishmania major Infection among Psammomys obesus and Meriones shawi: Reservoirs of Zoonotic Cutaneous Leishmaniasis in Sidi Bouzid (Central Tunisia). Vector-Borne Zoonotic Dis. 2011, 11, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Fichet-Calvet, E.; Jomâa, I.; Ben Ismail, R.; Ashford, R.W. Leishmania major infection in the fat sand rat Psammomys obesus in Tunisia: Interaction of host and parasite populations. Ann. Trop. Med. Parasitol. 2003, 97, 593–603. [Google Scholar] [CrossRef]
- Akhavan, A.A.; Yaghoobi-Ershadi, M.R.; Khamesipour, A.; Mirhendi, H.; Alimohammadian, M.H.; Rassi, Y.; Arandian, M.H.; Jafari, R.; Abdoli, H.; Shareghi, N.; et al. Dynamics of Leishmania infection rates in Rhombomys opimus (Rodentia: Gerbillinae) population of an endemic focus of zoonotic cutaneous leishmaniasis in Iran. Bull. La Soc. Pathol. Exot. 2010, 103, 84–89. [Google Scholar] [CrossRef]
- Akhoundi, M.; Mohebali, M.; Asadi, M.; Mahmodi, M.R.; Amraei, K.; Mirzaei, A. Molecular characterization of Leishmania spp. in reservoir hosts in endemic foci of zoonotic cutaneous leishmaniasis in Iran. Folia Parasitol. 2013, 60, 218–224. [Google Scholar] [CrossRef]
- Singh, O.P.; Hasker, E.; Sacks, D.; Boelaert, M.; Sundar, S. Asymptomatic Leishmania infection: A new challenge for Leishmania control. Clin. Infect. Dis. 2014, 58, 1424–1429. [Google Scholar] [CrossRef]
- Michel, G.; Pomares, C.; Ferrua, B.; Marty, P. Importance of worldwide asymptomatic carriers of Leishmania infantum (L. chagasi) in human. Acta Trop. 2011, 119, 69–75. [Google Scholar] [CrossRef]
- Stauch, A.; Sarkar, R.R.; Picado, A.; Ostyn, B.; Sundar, S.; Rijal, S.; Boelaert, M.; Dujardin, J.-C.; Duerr, H.-P. Visceral leishmaniasis in the Indian subcontinent: Modelling epidemiology and control. PLoS Negl. Trop. Dis. 2011, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Quinnell, R.J.; Courtenay, O. Transmission, reservoir hosts and control of zoonotic visceral leishmaniasis. Parasitology 2009, 136, 1915–1934. [Google Scholar] [CrossRef] [PubMed]
- Beniklef, R.; Aoun, K.; Boudrissa, K.; Ben Abid, M.; Cherif, K.; Aissi, W.; Benrekta, S.; Boubidi, S.C.; Späth, G.F.; Bouratbine, A.; et al. Cutaneous Leishmaniasis in Algeria; Highlight on the Focus of M’Sila. Microorganisms 2021, 9, 962. [Google Scholar] [CrossRef] [PubMed]
- Volf, P.; Volfova, V. Establishment and maintenance of sand fly colonies. J. Vector Ecol. 2011, 36, S1–S9. [Google Scholar] [CrossRef] [PubMed]
- Dean, S.; Sunter, J.; Wheeler, R.J.; Hodkinson, I.; Gluenz, E.; Gull, K. A toolkit enabling efficient, scalable and reproducible gene tagging in trypanosomatids. Open Biol. 2015, 5, 140197. [Google Scholar] [CrossRef] [PubMed]
- Sadlova, J.; Myskova, J.; Lestinova, T.; Votypka, J.; Yeo, M.; Volf, P. Leishmania donovani development in Phlebotomus argentipes: Comparison of promastigote-and amastigote-initiated infections. Parasitology 2017, 144, 403–410. [Google Scholar] [CrossRef]
- Sadlova, J.; Seblova, V.; Votypka, J.; Warburg, A.; Volf, P. Xenodiagnosis of Leishmania donovani in BALB/c mice using Phlebotomus orientalis: A new laboratory model. Parasite Vector 2015, 8, 158. [Google Scholar] [CrossRef]
- Rodgers, M.R.; Popper, S.J.; Wirth, D.F. Amplification of kinetoplast DNA as a tool in the detection and diagnosis of Leishmania. Exp. Parasitol. 1990, 71, 267–275. [Google Scholar] [CrossRef]
- Pekár, S.; Brabec, M. Generalized estimating equations: A pragmatic and flexible approach to the marginal GLM modelling of correlated data in the behavioural sciences. Ethology 2018, 124, 86–93. [Google Scholar] [CrossRef]
- Halekoh, U.; Højsgaard, S.; Yan, J. The R package geepack for generalized estimating equations. J. Stat. Softw. 2006, 15, 1–11. [Google Scholar] [CrossRef]
- Fichet-Calvet, E. Meriones shawi Shaw’s Jird. In Mammals of Africa: Rodents, Hares and Rabbits; Happold, D.C.D., Ed.; Bloomsbury Publishing: London, UK, 2013; Volume 3, pp. 338–339. [Google Scholar]
- Samy, A.M.; Annajar, B.B.; Dokhan, M.R.; Boussaa, S.; Peterson, A.T. Coarse-resolution Ecology of Etiological Agent, Vector, and Reservoirs of Zoonotic Cutaneous Leishmaniasis in Libya. PLoS Negl. Trop. Dis. 2016, 10, e0004381. [Google Scholar] [CrossRef] [PubMed]
- Derbali, M.; Chelbi, I.; Ben Hadj Ahmed, S.; Zhioua, E. Leishmania major Yakimoff et Schokhor, 1914 (Kinetoplastida: Trypanosomatidae) chez Meriones shawi Duvernoy, 1842 (Rodentia: Gerbillidae): Persistance de l’infection du mérion et de son infectivité pour le phlébotome vecteur Phlebotomus (Phlebotomus) papatasi Scopoli, 1786 (Diptera: Psychodidae). Bull. La Soc. Pathol. Exot. 2012, 105, 399–402. [Google Scholar] [CrossRef]
- Ashford, R.W. Leishmaniasis reservoirs and their significance in control. Clin. Dermatol. 1996, 14, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Strelkova, M.V. Progress in studies on Central Asian foci of zoonotic cutaneous leishmaniasis: A review. Folia Parasitol. 1996, 43, 1–6. [Google Scholar]
- Akhavan, A.A.; Mirhendi, H.; Khamesipour, A.; Alimohammadian, M.H.; Rassi, Y.; Bates, P.; Kamhawi, S.; Valenzuela, J.G.; Arandian, M.H.; Abdoli, H.; et al. Leishmania species: Detection and identification by nested PCR assay from skin samples of rodent reservoirs. Exp. Parasitol. 2010, 126, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Kamhawi, S. The biological and immunomodulatory properties of sand fly saliva and its role in the establishment of Leishmania infections. Microbes Infect. 2000, 2, 1765–1773. [Google Scholar] [CrossRef]
- Lestinova, T.; Rohousova, I.; Sima, M.; de Oliveira, C.I.; Volf, P. Insights into the sand fly saliva: Blood-feeding and immune interactions between sand flies, hosts, and Leishmania. PLoS Negl. Trop. Dis. 2017, 11, e0005600. [Google Scholar] [CrossRef]
- Kimblin, N.; Peters, N.; Debrabant, A.; Secundino, N.; Egen, J.; Lawyer, P.; Fay, M.P.; Kamhawi, S.; Sacks, D. Quantification of the infectious dose of Leishmania major transmitted to the skin by single sand flies. Proc. Natl. Acad. Sci. USA 2008, 105, 10125–10130. [Google Scholar] [CrossRef]
- Maia, C.; Seblova, V.; Sadlova, J.; Votypka, J.; Volf, P. Experimental transmission of Leishmania infantum by two major vectors: A comparison between a viscerotropic and a dermotropic strain. PLoS Negl. Trop. Dis. 2011, 5, e1181. [Google Scholar] [CrossRef]
- Belkaid, Y.; Mendez, S.; Lira, R.; Kadambi, N.; Milon, G.; Sacks, D. Natural Model of Leishmania major Infection Reveals a Prolonged “Silent” Phase of Parasite Amplification in the Skin Before the Onset of Lesion Formation and Immunity. J. Immunol. 2000, 165, 969–977. [Google Scholar] [CrossRef]
- Côrtes, D.F.; Carneiro, M.B.; Santos, L.M.; Souza, T.C.; Maioli, T.U.; Duz, A.L.; Ramos-Jorge, M.L.; Afonso, L.C.; Carneiro, C.; Vieira, L.Q. Low and high-dose intradermal infection with Leishmania major and Leishmania amazonensis in C57BL/6 mice. Mem. Inst. Oswaldo Cruz 2010, 105, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Vojtkova, B.; Spitzova, T.; Votypka, J.; Lestinova, T.; Kominkova, I.; Hajkova, M.; Santos-Mateus, D.; Miles, M.A.; Volf, P.; Sadlova, J. Central Asian Rodents as Model Animals for Leishmania major and Leishmania donovani Research. Microorganisms 2020, 8, 1440. [Google Scholar] [CrossRef] [PubMed]
- Doehl, J.S.P.; Bright, Z.; Dey, S.; Davies, H.; Magson, J.; Brown, N.; Romano, A.; Dalton, J.E.; Pinto, A.I.; Pitchford, J.W.; et al. Skin parasite landscape determines host infectiousness in visceral leishmaniasis. Nat. Comm. 2017, 28, 57. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, M.; González, E.; Martín-Martín, I.; Hernández, S.; Molina, R. Could wild rabbits (Oryctolagus cuniculus) be reservoirs for Leishmania infantum in the focus of Madrid, Spain? Vet. Parasitol. 2014, 202, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Molina, R.; Jiménez, M.I.; Cruz, I.; Iriso, A.; Martín-Martín, I.; Sevillano, O.; Melero, S.; Bernal, J. The hare (Lepus granatensis) as potential sylvatic reservoir of Leishmania infantum in Spain. Vet. Parasitol. 2012, 190, 268–271. [Google Scholar] [CrossRef]
- Gradoni, L.; Pozio, E.; Gramiccia, M.; Maroli, M.; Bettini, S. Leishmaniasis in Tuscany (Italy): VII. Studies on the role of the black rat, Rattus rattus, in the epidemiology of visceral leishmaniasis. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 427–431. [Google Scholar] [CrossRef]
- De Oliveira, A.R.; Pinheiro, G.R.G.; Tinoco, H.P.; Loyola, M.E.; Coelho, C.M.; Dias, E.S.; Michalsky Monteiro, E.; de Oliveira Lara e Silva, F.; Pessanha, A.T.; Maia Souza, A.M.; et al. Competence of non-human primates to transmit Leishmania infantum to the invertebrate vector Lutzomyia longipalpis. PLoS Negl. Trop. Dis. 2019, 13, e0007313. [Google Scholar] [CrossRef]
- Mol, J.P.S.; Soave, S.A.; Turchetti, A.P.; Pinheiro, G.R.G.; Pessanha, A.T.; Malta, M.C.C.; Tinoco, H.P.; Figueiredo, L.A.; Gontijo, N.F.; Paixão, T.A.; et al. Transmissibility of Leishmania infantum from maned wolves (Chrysocyon brachyurus) and bush dogs (Speothos venaticus) to Lutzomyia longipalpis. Vet. Parasitol. 2015, 212, 86–91. [Google Scholar] [CrossRef]
- Travi, B.L.; Osorio, Y.; Guarın, N.; Cadena, H. Leishmania (Leishmania) chagasi: Clinical and Parasitological Observations in Experimentally Infected Didelphis marsupialis, Reservoir of New World Visceral Leishmaniasis. Exp. Parasitol. 1998, 88, 73–75. [Google Scholar] [CrossRef]
- Courtenay, O.; Peters, N.C.; Rogers, M.E.; Bern, C. Combining epidemiology with basic biology of sand flies, parasites, and hosts to inform leishmaniasis transmission dynamics and control. PLoS Pathog. 2017, 13, e1006571. [Google Scholar] [CrossRef]
- Ashford, R.W.; Kohestany, K.A.; Karimzad, M.A. Cutaneous leishmaniasis in Kabul: Observations on a ‘prolonged epidemic’. Ann. Trop. Med. Parasitol. 1992, 86, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.A.; Herrer, A. Detection of Leishmania braziliensis by xenodiagnosis. Trans. R. Soc. Trop. Med. Hyg. 1972, 66, 798–799. [Google Scholar] [CrossRef] [PubMed]
- Svobodová, M.; Votýpka, J.; Nicolas, L.; Volf, P. Leishmania tropica in the black rat (Rattus rattus): Persistence and transmission from asymptomatic host to sand fly vector Phlebotomus sergenti. Microbes Infect. 2003, 5, 361–364. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Infective Dose | Week p.i. | Meriones No. | IE | CE | IE-LN | CE-LN | FP | HP | Tail | Spleen | Liver |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 14 × 103 | 16 | 1 | 8.30 × 106 | 0 | <1 × 102 | 0 | 0 | 0 | 0 | <1 × 102 | <1 × 102 |
| 14 × 103 | 16 | 2 | 2.75 × 105 | 0 | 0 | 0 | 0 | 0 | <1 × 102 | 0 | 0 |
| 14 × 103 | 40 | 3 * | 0 | 0 | <1 × 102 | 0 | 0 | 0 | 0 | <1 × 102 | 0 |
| 14 × 103 | 16 | 4 | 3.43 × 106 | 0 | <1 × 102 | 0 | 0 | 0 | 0 | 0 | 0 |
| 14 × 103 | 16 | 5 | 2.13 × 104 | 0 | <1 × 102 | 0 | 0 | 0 | 0 | <1 × 102 | 0 |
| 14 × 103 | 30 | 6 ** | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 14 × 103 | 30 | 7 | 3.29 × 109 | <1 × 102 | 0 | 0 | 0 | <1 × 102 | <1 × 102 | 0 | 0 |
| 14 × 103 | 25 | 8 | 1.41 × 105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 14 × 103 | 25 | 9 | 7.42 × 108 | <1 × 102 | 0 | 0 | <1 × 102 | <1 × 102 | <1 × 102 | 0 | <1 × 102 |
| 14 × 103 | 25 | 10 | 1.08 × 104 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 14 × 103 | 30 | 11 | 4.78 × 107 | <1 × 102 | 0 | 0 | 0 | <1 × 102 | <1 × 102 | 0 | 0 |
| 14 × 103 | 18 | 12 | 3.94 × 105 | 0 | <1 × 102 | 0 | <1 × 102 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 13 | 2.19 × 107 | <1 × 102 | 0 | 0 | 0 | <1 × 102 | <1 × 102 | 0 | <1 × 102 |
| 7 × 103 | 30 | 14 | 5.86 × 105 | <1 × 102 | 0 | 0 | 0 | 0 | <1 × 102 | <1 × 102 | <1 × 102 |
| 7 × 103 | 30 | 15 | 1.76 × 105 | 0 | <1 × 102 | 0 | 0 | 0 | <1 × 102 | 0 | 0 |
| 7 × 103 | 30 | 16 | 2.43 × 107 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 17 | 3.74 × 104 | 0 | 0 | 0 | <1 × 102 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 18 | 1.83 × 106 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 19 | 9.81 × 107 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 20 | 5.45 × 104 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 21 | 5.52 × 104 | 0 | 0 | 0 | <1 × 102 | 0 | <1 × 102 | <1 × 102 | 0 |
| 7 × 103 | 20 | 22 | 1.14 × 105 | <1 × 102 | 1.75 × 106 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 × 103 | 15 | 23 | 2.66 × 106 | <1 × 102 | <1 × 102 | 0 | 0 | 0 | 0 | <1 × 102 | 0 |
| 7 × 103 | 30 | 24 | 4.68 × 106 | 0 | 0 | 0 | 0 | <1 × 102 | 0 | 0 | 0 |
| 7 × 103 | 30 | 25 | 1.67 × 108 | 0 | 0 | 0 | <1 × 102 | 0 | 0 | 0 | <1 × 102 |
| 7 × 103 | 30 | 26 | 1.32 × 105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 27 | 7.46 × 103 | 0 | 0 | 0 | <1 × 102 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 28 | 1.56 × 103 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 29 | 3.35 × 105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 × 103 | 30 | 30 | 3.46 × 10 | 0 | 0 | 0 | 0 | <1 × 102 | 1.14 × 103 | 0 | 0 |
| 7 × 103 | 30 | 31 | 7.06 × 102 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadlova, J.; Vojtkova, B.; Lestinova, T.; Becvar, T.; Frynta, D.; Benallal, K.E.; Mekarnia, N.; Harrat, Z.; Volf, P. Infectiousness of Asymptomatic Meriones shawi, Reservoir Host of Leishmania major. Pathogens 2023, 12, 614. https://doi.org/10.3390/pathogens12040614
Sadlova J, Vojtkova B, Lestinova T, Becvar T, Frynta D, Benallal KE, Mekarnia N, Harrat Z, Volf P. Infectiousness of Asymptomatic Meriones shawi, Reservoir Host of Leishmania major. Pathogens. 2023; 12(4):614. https://doi.org/10.3390/pathogens12040614
Chicago/Turabian StyleSadlova, Jovana, Barbora Vojtkova, Tereza Lestinova, Tomas Becvar, Daniel Frynta, Kamal Eddine Benallal, Nalia Mekarnia, Zoubir Harrat, and Petr Volf. 2023. "Infectiousness of Asymptomatic Meriones shawi, Reservoir Host of Leishmania major" Pathogens 12, no. 4: 614. https://doi.org/10.3390/pathogens12040614