
Increased Clinical Signs and Mortality in IFNAR(−/−) Mice Immunised with the Bluetongue Virus Outer-Capsid Proteins VP2 or VP5, after Challenge with an Attenuated Heterologous Serotype

 , ,
, ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Cultures and Viruses
2.3. Infection of Cell Cultures and Purification of Viruses Prior to Inoculation into IFNAR(−/−) Mice
2.4. Experimental Design
2.5. Blood Collection, RNA Extraction and Real-Time RT-PCR
3. Results
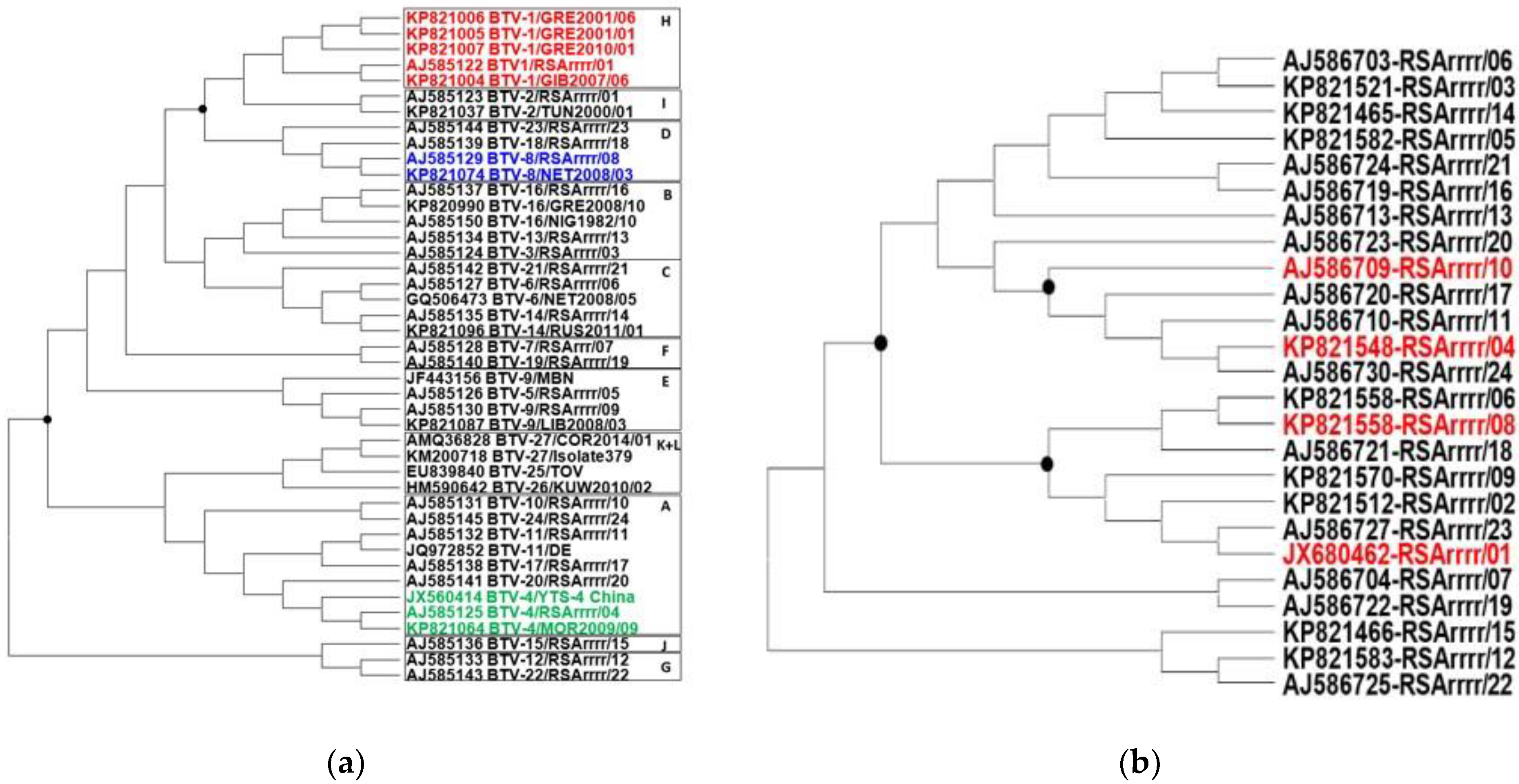
3.1. Phylogenetic Analysis and Group Design
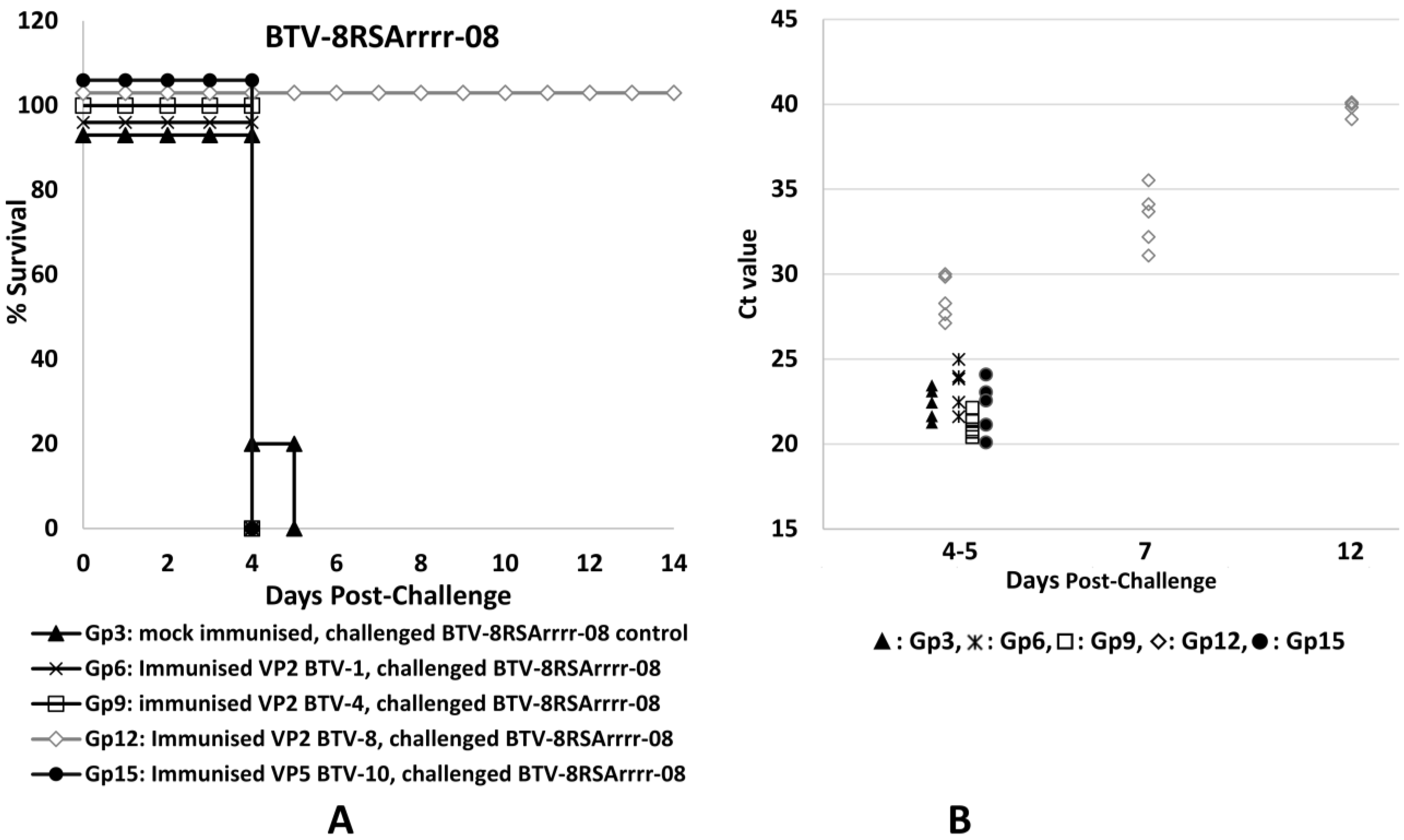
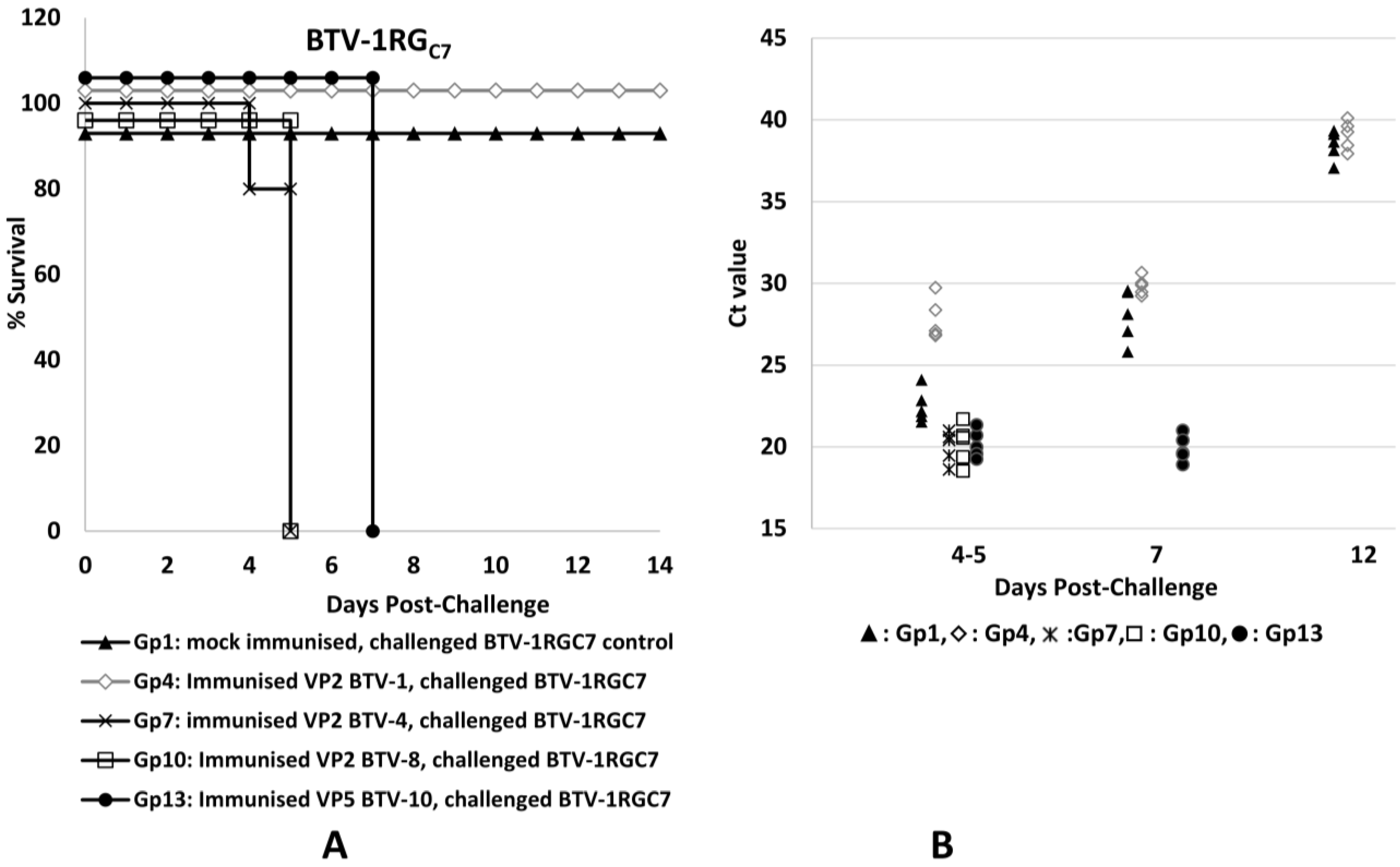
3.2. Clinical Observations
3.3. Real-Time PCR Assessment of BTV RNA in Mouse Blood (RNAemia)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greenspan, N.S.; Cavacini, L.A. Immunoglobulin Function. In Clinical Immunology; Rich, R.R., Fleisher, T.A., Shearer, W.T., Schroeder, H.W., Frew, A.J., Weyand, C.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 223–233. [Google Scholar]
- Utrilla-Trigo, S.; Jimenez-Cabello, L.; Calvo-Pinilla, E.; Marin-Lopez, A.; Lorenzo, G.; Sanchez-Cordon, P.; Moreno, S.; Benavides, J.; Gilbert, S.; Nogales, A.; et al. The Combined Expression of the Nonstructural Protein NS1 and the N-Terminal Half of NS2 (NS2(1-180)) by ChAdOx1 and MVA Confers Protection against Clinical Disease in Sheep upon Bluetongue Virus Challenge. J. Virol. 2022, 96, e0161421. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.C. T-cell-inducing vaccines—What’s the future. Immunology 2012, 135, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Dejnirattisai, W.; Jumnainsong, A.; Onsirisakul, N.; Fitton, P.; Vasanawathana, S.; Limpitikul, W.; Puttikhunt, C.; Edwards, C.; Duangchinda, T.; Supasa, S.; et al. Cross-reacting antibodies enhance dengue virus infection in humans. Science 2010, 328, 745–748. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, S.; Luedtke, A.; Langevin, E.; Zhu, M.; Bonaparte, M.; Machabert, T.; Savarino, S.; Zambrano, B.; Moureau, A.; Khromava, A.; et al. Effect of Dengue Serostatus on Dengue Vaccine Safety and Efficacy. N. Engl. J. Med. 2018, 379, 327–340. [Google Scholar] [CrossRef]
- Xu, L.; Ma, Z.; Li, Y.; Pang, Z.; Xiao, S. Antibody dependent enhancement: Unavoidable problems in vaccine development. Adv. Immunol. 2021, 151, 99–133. [Google Scholar]
- Hohdatsu, T.; Yamada, M.; Tominaga, R.; Makino, K.; Kida, K.; Koyama, H. Antibody-dependent enhancement of feline infectious peritonitis virus infection in feline alveolar macrophages and human monocyte cell line U937 by serum of cats experimentally or naturally infected with feline coronavirus. J. Vet. Med. Sci. 1998, 60, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.S.; Wheatley, A.K.; Kent, S.J.; DeKosky, B.J. Antibody-dependent enhancement and SARS-CoV-2 vaccines and therapies. Nat. Microbiol. 2020, 5, 1185–1191. [Google Scholar] [CrossRef]
- Martinelle, L.; Dal Pozzo, F.; Thys, C.; De Leeuw, I.; Van Campe, W.; De Clercq, K.; Thiry, E.; Saegerman, C. Assessment of cross-protection induced by a bluetongue virus (BTV) serotype 8 vaccine towards other BTV serotypes in experimental conditions. Vet. Res. 2018, 49, 63. [Google Scholar] [CrossRef] [Green Version]
- Howerth, E.W. Cytokine release and endothelial dysfunction: A perfect storm in orbivirus pathogenesis. Vet. Ital. 2015, 51, 275–281. [Google Scholar]
- Kim, H.W.; Canchola, J.G.; Brandt, C.D.; Pyles, G.; Chanock, R.M.; Jensen, K.; Parrott, R.H. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am. J. Epidemiol. 1969, 89, 422–434. [Google Scholar] [CrossRef]
- Graham, B.S. Vaccines against respiratory syncytial virus: The time has finally come. Vaccine 2016, 34, 3535–3541. [Google Scholar] [CrossRef] [Green Version]
- Toltzis, P. 50 Years Ago in The Journal of Pediatrics: Atypical Exanthem after Exposure to Natural Measles: Eleven Cases in Children Previously Inoculated with Killed Vaccine. J. Pediatr. 2018, 193, 84. [Google Scholar] [CrossRef]
- Polack, F.P. Atypical measles and enhanced respiratory syncytial virus disease (ERD) made simple. Pediatr. Res. 2007, 62, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Mohd Jaafar, F.; Belhouchet, M.; Belaganahalli, M.; Tesh, R.B.; Mertens, P.P.; Attoui, H. Full-genome characterisation of Orungo, Lebombo and Changuinola viruses provides evidence for co-evolution of orbiviruses with their arthropod vectors. PLoS ONE 2014, 9, e86392. [Google Scholar] [CrossRef] [Green Version]
- Maan, S.; Maan, N.S.; Samuel, A.R.; Rao, S.; Attoui, H.; Mertens, P.P.C. Analysis and phylogenetic comparisons of full-length VP2 genes of the 24 bluetongue virus serotypes. J. Gen. Virol. 2007, 88 Pt 2, 621–630. [Google Scholar] [CrossRef]
- Wu, W.; Celma, C.C.; Kerviel, A.; Roy, P. Mapping the pH Sensors Critical for Host Cell Entry by a Complex Nonenveloped Virus. J. Virol. 2019, 93, e01897-18. [Google Scholar] [CrossRef] [Green Version]
- Attoui, H.; Mertens, P.P.C.; Becnel, J.; Belaganahalli, M.; Bergoin, M.; Brussaard, C.P.; Chappell, J.D.; Ciarlet, M.; del Vas, M.; Dermody, T.S.; et al. Reoviridae. In Virus Taxonomy: The Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier-Academic Press: Amsterdam, The Netherlands, 2012; pp. 541–637. [Google Scholar]
- Matthijnssens, J.; Attoui, H.; Banyai, K.; Brussaard, C.P.D.; Danthi, P.; Del Vas, M.; Dermody, T.S.; Duncan, R.; Fang, Q.; Johne, R.; et al. ICTV Virus Taxonomy Profile: Sedoreoviridae 2022. J. Gen. Virol. 2022, 103, 001782. [Google Scholar] [CrossRef]
- Cowley, J.A.; Gorman, B.M. Cross-neutralization of genetic reassortants of bluetongue virus serotypes 20 and 21. Vet. Microbiol. 1989, 19, 37–51. [Google Scholar] [CrossRef]
- Mertens, P.P.; Pedley, S.; Cowley, J.; Burroughs, J.N.; Corteyn, A.H.; Jeggo, M.H.; Jennings, D.M.; Gorman, B.M. Analysis of the roles of bluetongue virus outer capsid proteins VP2 and VP5 in determination of virus serotype. Virology 1989, 170, 561–565. [Google Scholar] [CrossRef]
- DeMaula, C.D.; Heidner, H.W.; Rossitto, P.V.; Pierce, C.M.; MacLachlan, N.J. Neutralization determinants of United States bluetongue virus serotype ten. Virology 1993, 195, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Belbis, G.; Zientara, S.; Breard, E.; Sailleau, C.; Caignard, G.; Vitour, D.; Attoui, H. Bluetongue Virus: From BTV-1 to BTV-27. Adv. Virus Res. 2017, 99, 161–197. [Google Scholar] [PubMed]
- Schwartz-Cornil, I.; Mertens, P.P.; Contreras, V.; Hemati, B.; Pascale, F.; Breard, E.; Mellor, P.S.; MacLachlan, N.J.; Zientara, S. Bluetongue virus: Virology, pathogenesis and immunity. Vet. Res. 2008, 39, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drew, C.P.; Heller, M.C.; Mayo, C.; Watson, J.L.; Maclachlan, N.J. Bluetongue virus infection activates bovine monocyte-derived macrophages and pulmonary artery endothelial cells. Vet. Immunol. Immunopathol. 2010, 136, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Darpel, K.E.; Batten, C.A.; Veronesi, E.; Shaw, A.E.; Anthony, S.; Bachanek-Bankowska, K.; Kgosana, L.; bin-Tarif, A.; Carpenter, S.; Muller-Doblies, U.U.; et al. Clinical signs and pathology shown by British sheep and cattle infected with bluetongue virus serotype 8 derived from the 2006 outbreak in northern Europe. Vet. Rec. 2007, 161, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Huismans, H.; van der Walt, N.T.; Cloete, M.; Erasmus, B.J. Isolation of a capsid protein of bluetongue virus that induces a protective immune response in sheep. Virology 1987, 157, 172–179. [Google Scholar] [CrossRef]
- Bonneau, K.R.; DeMaula, C.D.; Mullens, B.A.; MacLachlan, N.J. Duration of viraemia infectious to Culicoides sonorensis in bluetongue virus-infected cattle and sheep. Vet. Microbiol. 2002, 88, 115–125. [Google Scholar] [CrossRef]
- Di Gialleonardo, L.; Migliaccio, P.; Teodori, L.; Savini, G. The length of BTV-8 viraemia in cattle according to infection doses and diagnostic techniques. Res. Vet. Sci. 2011, 91, 316–320. [Google Scholar] [CrossRef]
- De Clercq, K.; Mertens, P.; De Leeuw, I.; Oura, C.; Houdart, P.; Potgieter, A.C.; Maan, S.; Hooyberghs, J.; Batten, C.; Vandemeulebroucke, E.; et al. Emergence of bluetongue serotypes in Europe, part 2: The occurrence of a BTV-11 strain in Belgium. Transbound. Emerg. Dis. 2009, 56, 355–361. [Google Scholar] [CrossRef]
- Fay, P.C.; Attoui, H.; Batten, C.; Mohd Jaafar, F.; Lomonossoff, G.P.; Daly, J.M.; Mertens, P.P.C. Bluetongue virus outer-capsid protein VP2 expressed in Nicotiana benthamiana raises neutralising antibodies and a protective immune response in IFNAR (−/−) mice. Vaccine X 2019, 2, 100026. [Google Scholar] [CrossRef]
- Fay, P.C.; Mohd Jaafar, F.; Batten, C.; Attoui, H.; Saunders, K.; Lomonossoff, G.P.; Reid, E.; Horton, D.; Maan, S.; Haig, D.; et al. Serological Cross-Reactions between Expressed VP2 Proteins from Different Bluetongue Virus Serotypes. Viruses 2021, 13, 1455. [Google Scholar] [CrossRef]
- Sato, M.; Maeda, N.; Yoshida, H.; Urade, M.; Saito, S. Plaque formation of herpes virus hominis type 2 and rubella virus in variants isolated from the colonies of BHK21/WI-2 cells formed in soft agar. Arch. Virol. 1977, 53, 269–273. [Google Scholar] [CrossRef]
- Attoui, H.; Monsion, B.; Klonjkowski, B.; Zientara, S.; Mertens, P.P.C.; Mohd Jaafar, F. Identification of the Genome Segments of Bluetongue Virus Type 26/Type 1 Reassortants Influencing Horizontal Transmission in a Mouse Model. Viruses 2021, 13, 2208. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Thuenemann, E.C. Virus-Like Particle Production Using Cowpea Mosaic Virus-Based Vectors. Ph.D. Thesis, University of East Anglia, Norwich, UK, 2010. [Google Scholar]
- Mohd Jaafar, F.; Belhouchet, M.; Vitour, D.; Adam, M.; Breard, E.; Zientara, S.; Mertens, P.P.; Attoui, H. Immunisation with bacterial expressed VP2 and VP5 of bluetongue virus (BTV) protect alpha/beta interferon-receptor knock-out (IFNAR(−/−)) mice from homologous lethal challenge. Vaccine 2014, 32, 4059–4067. [Google Scholar] [CrossRef]
- Hofmann, M.; Griot, C.; Chaignat, V.; Perler, L.; Thur, B. Bluetongue disease reaches Switzerland. Schweiz. Arch. Tierheilkd. 2008, 150, 49–56. [Google Scholar] [CrossRef]
- Hofmann, M.A.; Renzullo, S.; Mader, M.; Chaignat, V.; Worwa, G.; Thuer, B. Genetic characterization of toggenburg orbivirus, a new bluetongue virus, from goats, Switzerland. Emerg. Infect. Dis. 2008, 14, 1855–1861. [Google Scholar] [CrossRef]
- Singh, K.P.; Maan, S.; Samuel, A.R.; Rao, S.; Meyer, A.J.; Mertens, P.P. Phylogenetic analysis of bluetongue virus genome segment 6 (encoding VP5) from different serotypes. Vet. Ital. 2004, 40, 479–483. [Google Scholar]
- Xia, X.; Wu, W.; Cui, Y.; Roy, P.; Zhou, Z.H. Bluetongue virus capsid protein VP5 perforates membranes at low endosomal pH during viral entry. Nat. Microbiol. 2021, 6, 1424–1432. [Google Scholar] [CrossRef]
- Hassan, S.H.; Wirblich, C.; Forzan, M.; Roy, P. Expression and functional characterization of bluetongue virus VP5 protein: Role in cellular permeabilization. J. Virol. 2001, 75, 8356–8367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maan, S.; Maan, N.S.; Nomikou, K.; Veronesi, E.; Bachanek-Bankowska, K.; Belaganahalli, M.N.; Attoui, H.; Mertens, P.P. Complete genome characterisation of a novel 26th bluetongue virus serotype from Kuwait. PLoS ONE 2011, 6, e26147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, P.; Bishop, D.H.; LeBlois, H.; Erasmus, B.J. Long-lasting protection of sheep against bluetongue challenge after vaccination with virus-like particles: Evidence for homologous and partial heterologous protection. Vaccine 1994, 12, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Utrilla-Trigo, S.; Calvo-Pinilla, E.; Jimenez-Cabello, L.; Ortego, J.; Nogales, A. Inhibition of Orbivirus Replication by Aurintricarboxylic Acid. Int. J. Mol. Sci. 2020, 21, 7294. [Google Scholar] [CrossRef]
- Calvo-Pinilla, E.; de la Poza, F.; Gubbins, S.; Mertens, P.P.; Ortego, J.; Castillo-Olivares, J. Antiserum from mice vaccinated with modified vaccinia Ankara virus expressing African horse sickness virus (AHSV) VP2 provides protection when it is administered 48h before, or 48h after challenge. Antiviral Res. 2015, 116, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Pinilla, E.; Rodriguez-Calvo, T.; Sevilla, N.; Ortego, J. Heterologous prime boost vaccination with DNA and recombinant modified vaccinia virus Ankara protects IFNAR(−/−) mice against lethal bluetongue infection. Vaccine 2009, 28, 437–445. [Google Scholar] [CrossRef]
- Jabbar, T.K.; Calvo-Pinilla, E.; Mateos, F.; Gubbins, S.; Bin-Tarif, A.; Bachanek-Bankowska, K.; Alpar, O.; Ortego, J.; Takamatsu, H.H.; Mertens, P.P.; et al. Protection of IFNAR (−/−) mice against bluetongue virus serotype 8, by heterologous (DNA/rMVA) and homologous (rMVA/rMVA) vaccination, expressing outer-capsid protein VP2. PLoS ONE 2013, 8, e60574. [Google Scholar] [CrossRef]
- Mohd Jaafar, F.; Monsion, B.; Belhouchet, M.; Mertens, P.P.C.; Attoui, H. Inhibition of Orbivirus Replication by Fluvastatin and Identification of the Key Elements of the Mevalonate Pathway Involved. Viruses 2021, 13, 1437. [Google Scholar] [CrossRef]
- Jeggo, M.H.; Gumm, I.D.; Taylor, W.P. Clinical and serological response of sheep to serial challenge with different bluetongue virus types. Res. Vet. Sci. 1983, 34, 205–211. [Google Scholar] [CrossRef]
- Batten, C.A.; Maan, S.; Shaw, A.E.; Maan, N.S.; Mertens, P.P. A European field strain of bluetongue virus derived from two parental vaccine strains by genome segment reassortment. Virus Res. 2008, 137, 56–63. [Google Scholar] [CrossRef]
- Maan, S.; Maan, N.S.; van Rijn, P.A.; van Gennip, R.G.; Sanders, A.; Wright, I.M.; Batten, C.; Hoffmann, B.; Eschbaumer, M.; Oura, C.A.; et al. Full genome characterisation of bluetongue virus serotype 6 from The Netherlands 2008 and comparison to other field and vaccine strains. PLoS ONE 2010, 5, e10323. [Google Scholar] [CrossRef]
- Rojas, J.M.; Barba-Moreno, D.; Avia, M.; Sevilla, N.; Martin, V. Vaccination With Recombinant Adenoviruses Expressing the Bluetongue Virus Subunits VP7 and VP2 Provides Protection Against Heterologous Virus Challenge. Front. Vet. Sci. 2021, 8, 645561. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Immunisation | Challenge | |
|---|---|---|---|
| Day 0 (Prime) | Day 14 (Boost) | Day 28 | |
| Group 1 (n = 5) | mock-immunised | mock-immunised | BTV-1RGC7 (control) |
| Group 2 (n = 5) | mock-immunised | mock-immunised | BTV-4RSArrrr/04 (control) |
| Group 3 (n = 5) | mock-immunised | mock-immunised | BTV-8RSArrrr/08 (control) |
| Group 4 (n = 5) | rVP2 of BTV-1 | rVP2 of BTV-1 | BTV-1RGC7 (homologous) |
| Group 5 (n = 5) | rVP2 of BTV-1 | rVP2 of BTV-1 | BTV-4RSArrrr/04 (heterologous) |
| Group 6 (n = 5) | rVP2 of BTV-1 | rVP2 of BTV-1 | BTV-8RSArrrr/08 (heterologous) |
| Group 7 (n = 5) | rVP2 of BTV-4 | rVP2 of BTV-4 | BTV-1RGC7 (heterologous) |
| Group 8 (n = 5) | rVP2 of BTV-4 | rVP2 of BTV-4 | BTV-4RSArrrr/04 (homologous) |
| Group 9 (n = 5) | rVP2 of BTV-4 | rVP2 of BTV-4 | BTV-8RSArrrr/08 (heterologous) |
| Group 10 (n = 5) | rVP2 of BTV-8 | rVP2 of BTV-8 | BTV-1RGC7 (heterologous) |
| Group 11 (n = 5) | rVP2 of BTV-8 | rVP2 of BTV-8 | BTV-4RSArrrr/04 (heterologous) |
| Group 12 (n = 5) | rVP2 of BTV-8 | rVP2 of BTV-8 | BTV-8RSArrrr/08 (homologous) |
| Group 13 (n = 5) | rVP5 of BTV-10 | rVP5 of BTV-10 | BTV-1RGC7 (heterologous) |
| Group 14 (n = 5) | rVP5 of BTV-10 | rVP5 of BTV-10 | BTV-4RSArrrr/04 (heterologous) |
| Group 15 (n = 5) | rVP5 of BTV-10 | rVP5 of BTV-10 | BTV-8RSArrrr/08 (heterologous) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attoui, H.; Mohd Jaafar, F.; Monsion, B.; Klonjkowski, B.; Reid, E.; Fay, P.C.; Saunders, K.; Lomonossoff, G.; Haig, D.; Mertens, P.P.C. Increased Clinical Signs and Mortality in IFNAR(−/−) Mice Immunised with the Bluetongue Virus Outer-Capsid Proteins VP2 or VP5, after Challenge with an Attenuated Heterologous Serotype. Pathogens 2023, 12, 602. https://doi.org/10.3390/pathogens12040602
Attoui H, Mohd Jaafar F, Monsion B, Klonjkowski B, Reid E, Fay PC, Saunders K, Lomonossoff G, Haig D, Mertens PPC. Increased Clinical Signs and Mortality in IFNAR(−/−) Mice Immunised with the Bluetongue Virus Outer-Capsid Proteins VP2 or VP5, after Challenge with an Attenuated Heterologous Serotype. Pathogens. 2023; 12(4):602. https://doi.org/10.3390/pathogens12040602
Chicago/Turabian StyleAttoui, Houssam, Fauziah Mohd Jaafar, Baptiste Monsion, Bernard Klonjkowski, Elizabeth Reid, Petra C. Fay, Keith Saunders, George Lomonossoff, David Haig, and Peter P. C. Mertens. 2023. "Increased Clinical Signs and Mortality in IFNAR(−/−) Mice Immunised with the Bluetongue Virus Outer-Capsid Proteins VP2 or VP5, after Challenge with an Attenuated Heterologous Serotype" Pathogens 12, no. 4: 602. https://doi.org/10.3390/pathogens12040602