

The Brown Alga Bifurcaria bifurcata Presents an Anthelmintic Activity on All Developmental Stages of the Parasitic Nematode Heligmosomoides polygyrus bakeri
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algae Collection and Aqueous Extract Preparation
2.2. Preparation of Organic Extracts
2.3. Parasite Material
2.4. Hatching Test and Mortality after Hatching Test
2.5. Larval Development Assay (LDA)
2.6. Survival Test
2.7. Statistical Analyses
3. Results
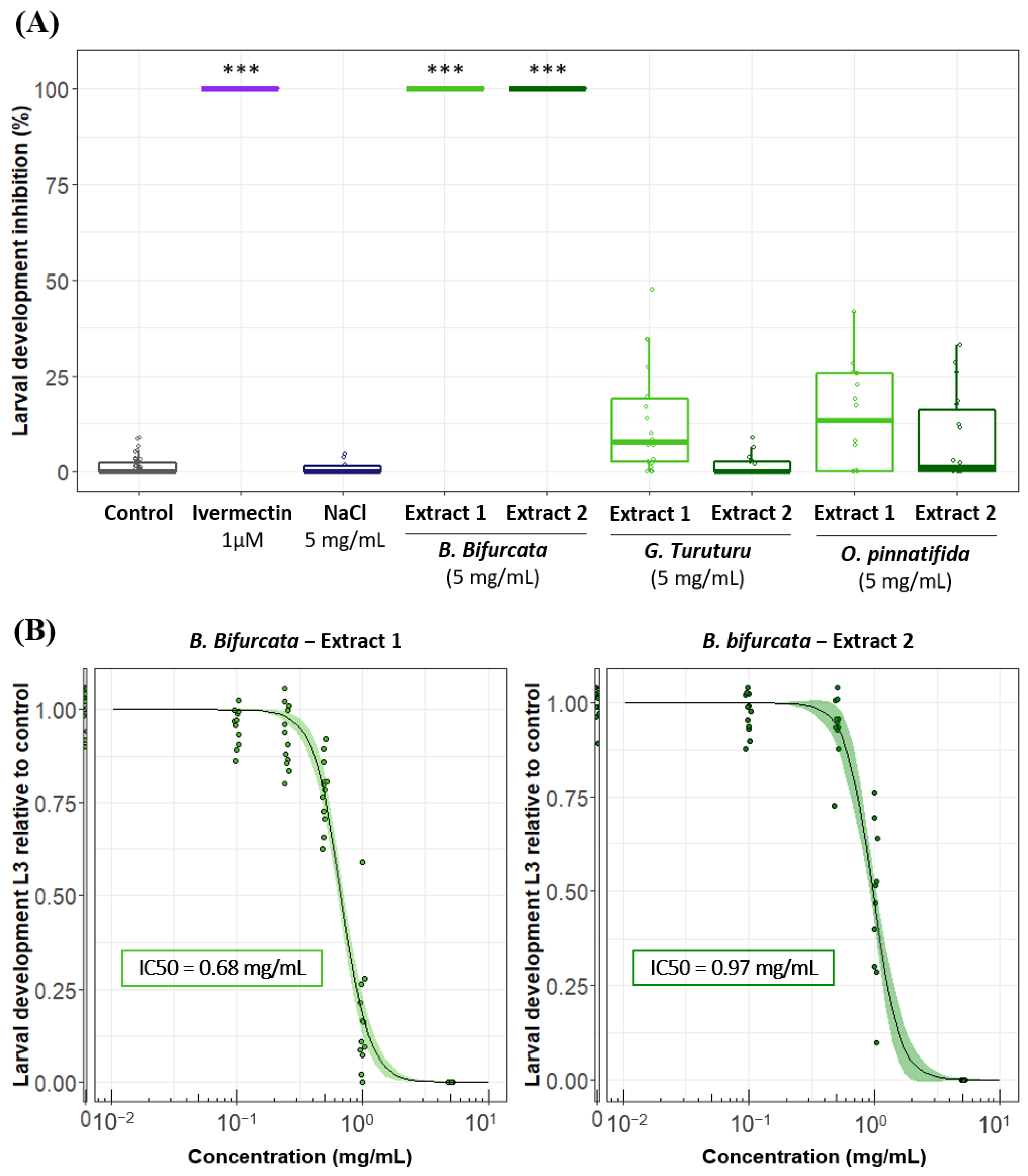
3.1. Larval Development Assay (LDA) Reveals Anthelmintic Activity of Aqueous Algal Extracts on H. polygyrus bakeri
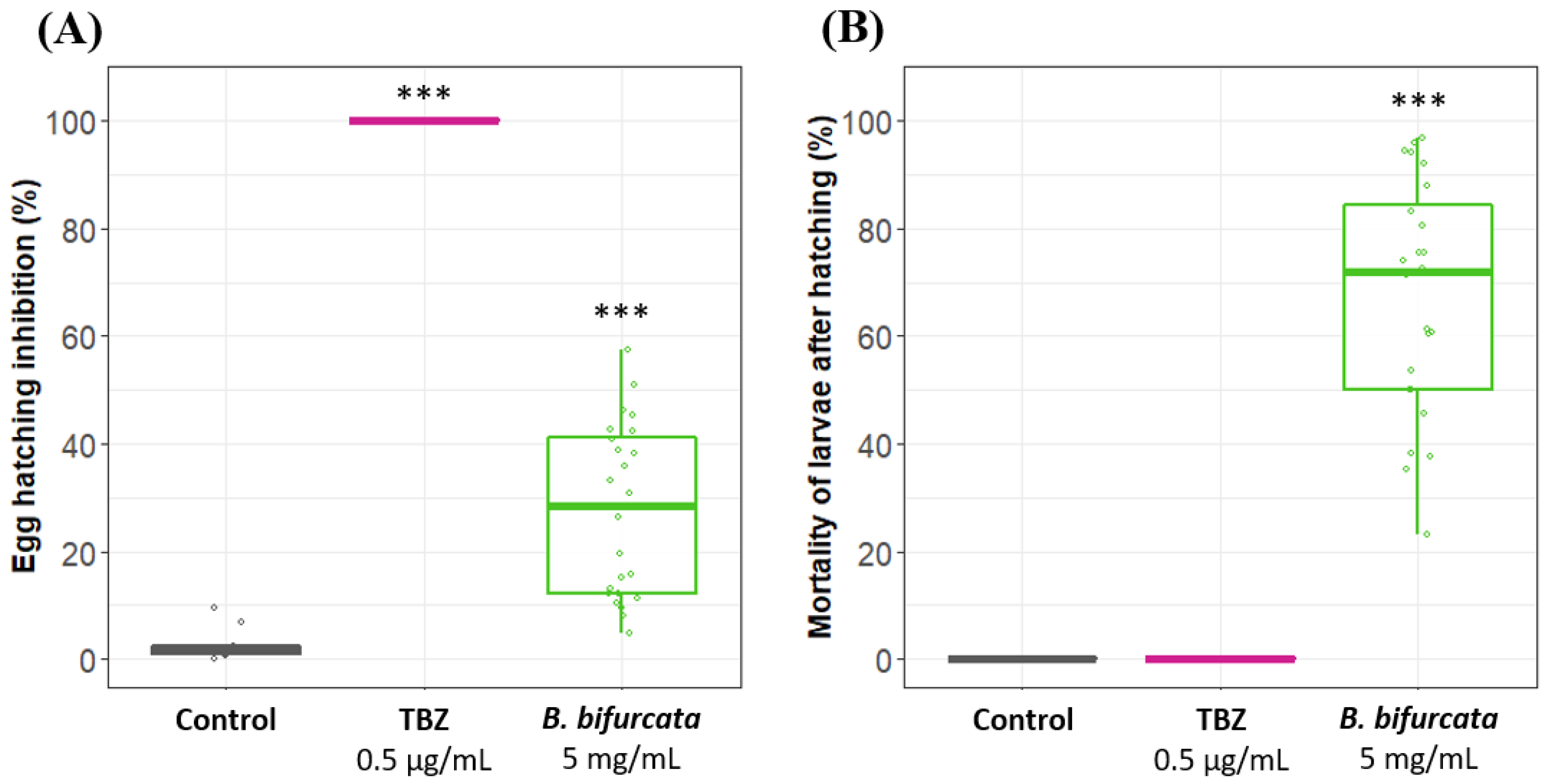
3.2. B. bifurcata Inhibits Egg Hatching and Has Nematicidal Activity on H. polygyrus bakeri Adult Worms
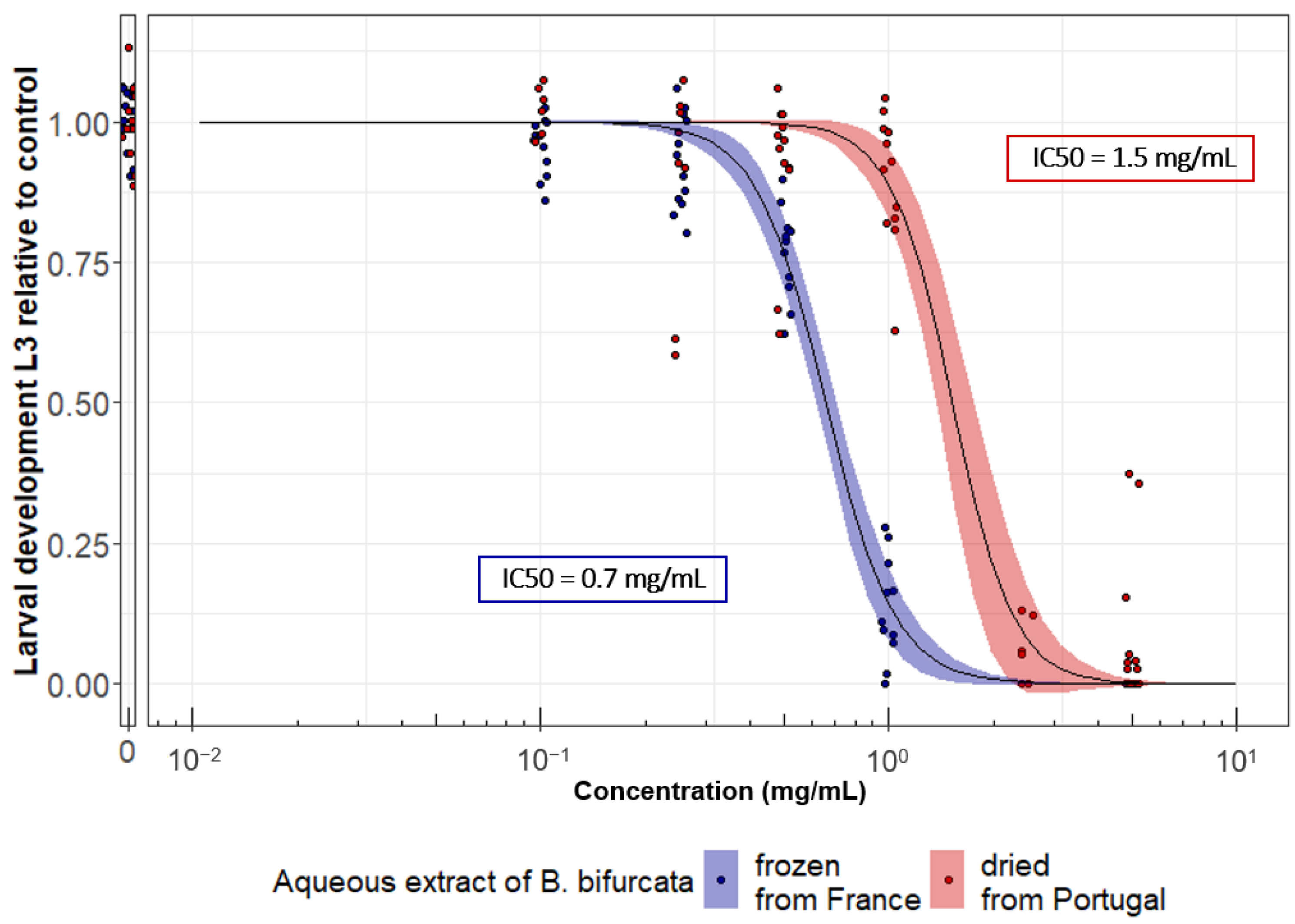
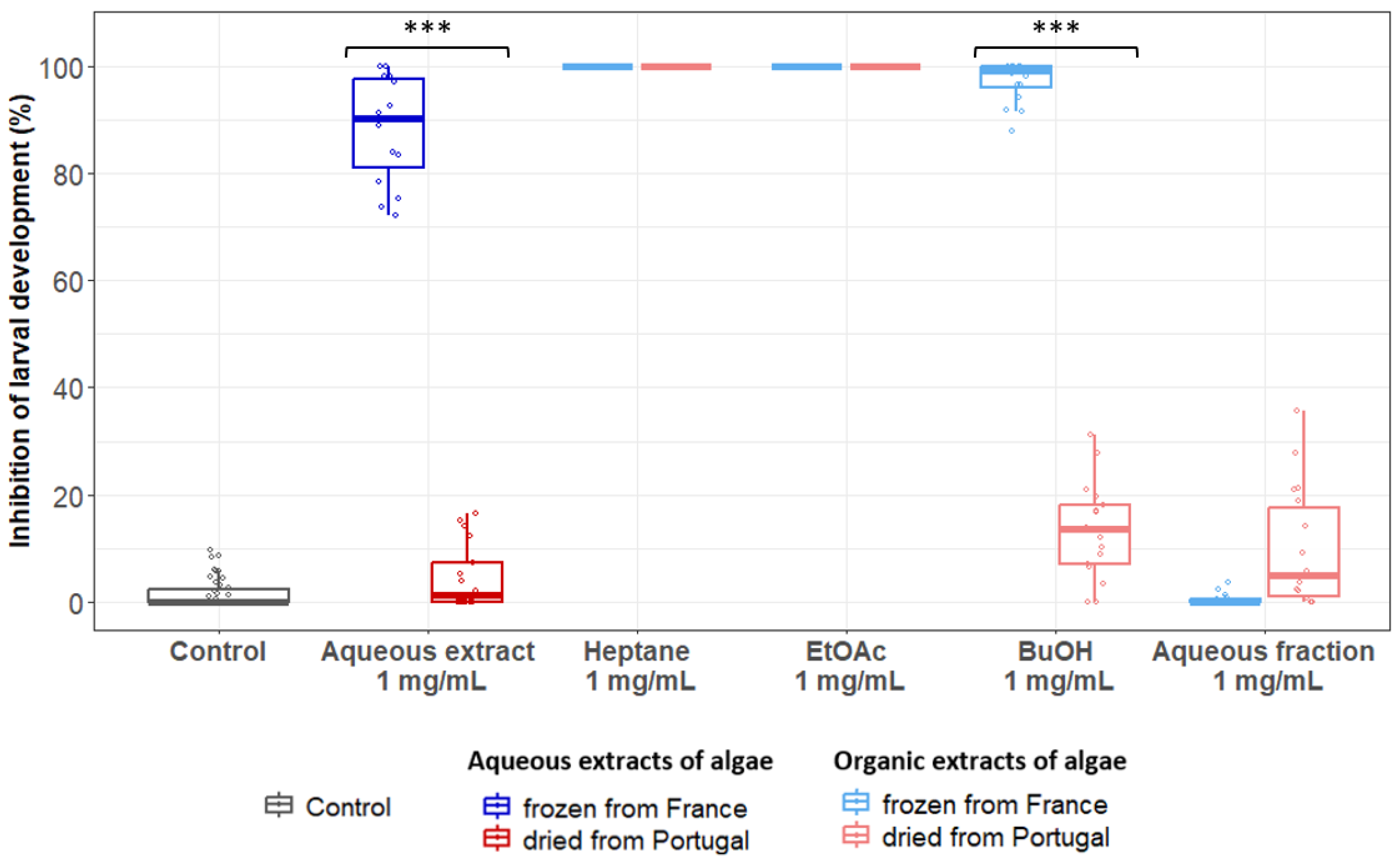
3.3. The Anthelmintic Activity of B. bifurcata Could Vary Depending on the Extraction Process and/or Geographical Origin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charlier, J.; van der Voort, M.; Kenyon, F.; Skuce, P.; Vercruysse, J. Chasing helminths and their economic impact on farmed ruminants. Trends Parasitol. 2014, 30, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Athanasiadou, S.; Kyriazakis, I. Plant secondary metabolites: Antiparasitic effects and their role in ruminant production systems. Proc. Nutr. Soc. 2004, 63, 631–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keiser, J.; Utzinger, J. Efficacy of current drugs against soil-transmitted helminth infections: Systematic review and meta-analysis. JAMA 2008, 299, 1937–1948. [Google Scholar] [CrossRef]
- Jayawardene, K.D.; Palombo, E.A.; Boag, P.R. Natural Products Are a Promising Source for Anthelmintic Drug Discovery. Biomolecules 2021, 11, 1457. [Google Scholar] [CrossRef] [PubMed]
- Taki, A.C.; Brkljaca, R.; Wang, T.; Koehler, A.V.; Ma, G.; Danne, J.; Ellis, S.; Hofmann, A.; Chang, B.C.H.; Jabbar, A.; et al. Natural Compounds from the Marine Brown Alga Caulocystis cephalornithos with Potent In Vitro-Activity against the Parasitic Nematode Haemonchus contortus. Pathogens 2020, 9, 550. [Google Scholar] [CrossRef] [PubMed]
- Berri, M.; Slugocki, C.; Olivier, M.; Helloin, E.; Jacques, I.; Salmon, H.; Demais, H.; Le Goff, M.; Collen, P.N. Marine-sulfated polysaccharides extract of Ulva armoricana green algae exhibits an antimicrobial activity and stimulates cytokine expression by intestinal epithelial cells. J. Appl. Phycol. 2016, 28, 2999–3008. [Google Scholar] [CrossRef]
- Moussavou, G.; Kwak, D.H.; Obiang-Obonou, B.W.; Maranguy, C.A.O.; Dinzouna-Boutamba, S.-D.; Lee, D.H.; Pissibanganga, O.G.M.; Ko, K.; Seo, J.I.; Choo, Y.K. Anticancer effects of different seaweeds on human colon and breast cancers. Mar. Drugs 2014, 12, 4898–4911. [Google Scholar] [CrossRef]
- Bonde, C.S.; Bornancin, L.; Lu, Y.; Simonsen, H.T.; Martínez-Valladares, M.; Peña-Espinoza, M.; Mejer, H.; Williams, A.R.; Thamsborg, S.M. Bio-Guided Fractionation and Molecular Networking Reveal Fatty Acids to Be Principal Anti-Parasitic Compounds in Nordic Seaweeds. Front. Pharmacol. 2021, 12, 674520. [Google Scholar] [CrossRef]
- Higa, T.; Kuniyoshi, M. Toxins associated with medicinal and edible seaweeds. J. Toxicol. Toxin Rev. 2000, 19, 119–137. [Google Scholar] [CrossRef]
- Orfanidis, S.; Breeman, A.M. Geographic variation in thermal traits in Digenea simplex and Champia parvula (Rhodophyta) in relation to present and glacial temperature regimes. J. Phycol. 1999, 35, 919–930. [Google Scholar] [CrossRef]
- Da Costa, E.; Melo, T.; Reis, M.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Polar lipids composition, antioxidant and anti-inflammatory activities of the atlantic red seaweed Grateloupia turuturu. Mar. Drugs 2021, 19, 414. [Google Scholar] [CrossRef] [PubMed]
- Plouguerne, E.; Hellio, C.; Deslandes, E.; Veron, B.; Stiger-Pouvreau, V. Anti-microfouling activities in extracts of two invasive algae: Grateloupia turuturu and Sargassum muticum. Bot. Mar. 2008, 51, 202–208. [Google Scholar] [CrossRef]
- Cardoso, I.; Cotas, J.; Rodrigues, A.; Ferreira, D.; Osório, N.; Pereira, L. Extraction and analysis of compounds with antibacterial potential from the red alga Grateloupia turuturu. J. Mar. Sci. Eng. 2019, 7, 220. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.; Fernandes, C.; Barros, L.; Ferreira, I.C.; Pereira, L.; Gonçalves, T. The antifungal activity of extracts of Osmundea pinnatifida, an edible seaweed, indicates its usage as a safe environmental fungicide or as a food additive preventing post-harvest fungal food contamination. Food Funct. 2018, 9, 6187–6195. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.; Pereira, L. Concise review of Osmundea pinnatifida (Hudson) Stackhouse. J. Appl. Phycol. 2020, 32, 2761–2771. [Google Scholar] [CrossRef]
- Alves, C.; Pinteus, S.; Simões, T.; Horta, A.; Silva, J.; Tecelão, C.; Pedrosa, R. Bifurcaria bifurcata: A key macro-alga as a source of bioactive compounds and functional ingredients. Int. J. Food Sci. Technol. 2016, 51, 1638–1646. [Google Scholar] [CrossRef]
- Ainane, T.; Abourriche, A. Brown Seaweed Bifurcaria bifurcata: Bioguided Fractionation of Extracts by Antibacterial Activity and Cytotoxicity Test. Biosci. Biotechnol. Res. Asia 2014, 11, 1081–1085. [Google Scholar] [CrossRef]
- Horta, A.; Pinteus, S.; Alves, C.; Fino, N.; Silva, J.; Fernandez, S.; Rodrigues, A.; Pedrosa, R. Antioxidant and antimicrobial potential of the Bifurcaria bifurcata epiphytic bacteria. Mar. Drugs 2014, 12, 1676–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz, J.; Culioli, G.; Köck, M. Linear diterpenes from the marine brown alga Bifurcaria bifurcata: A chemical perspective. Phytochem. Rev. 2013, 12, 407–424. [Google Scholar] [CrossRef]
- Smyrniotopoulos, V.; Merten, C.; Kaiser, M.; Tasdemir, D. Bifurcatriol, a new antiprotozoal acyclic diterpene from the brown alga Bifurcaria bifurcata. Mar. Drugs 2017, 15, 245. [Google Scholar] [CrossRef] [Green Version]
- Gallé, J.-B.; Attioua, B.; Kaiser, M.; Rusig, A.-M.; Lobstein, A.; Vonthron-Sénécheau, C. Eleganolone, a diterpene from the French marine alga Bifurcaria bifurcata inhibits growth of the human pathogens Trypanosoma brucei and Plasmodium falciparum. Mar. Drugs 2013, 11, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Palevich, N.; Maclean, P.H.; Choi, Y.-J.; Mitreva, M. Characterization of the complete mitochondrial genomes of two sibling species of parasitic roundworms, Haemonchus contortus and Teladorsagia circumcincta. Front. Genet. 2020, 11, 573395. [Google Scholar] [CrossRef] [PubMed]
- Wahid, F.N.; Behnke, J.M.; Conway, D.J. Factors affecting the efficacy of ivermectin against Heligmosomoides polygyrus (Nematospiroides dubius) in mice. Vet. Parasitol. 1989, 32, 325–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Githiori, J.B.; Höglund, J.; Waller, P.J.; Baker, R.L. Evaluation of anthelmintic properties of extracts from some plants used as livestock dewormers by pastoralist and smallholder farmers in Kenya against Heligmosomoides polygyrus infections in mice. Vet. Parasitol. 2003, 118, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.J.; Robertson, E.; Harcus, Y.; Grainger, J.R.; Coakley, G.; Smyth, D.J.; McSorley, H.J.; Maizels, R. Cultivation of Heligmosomoides polygyrus: An immunomodulatory nematode parasite and its secreted products. JoVE J. Vis. Exp. 2015, e52412. [Google Scholar]
- Taylor, M.; Hunt, K.; Goodyear, K. Anthelmintic resistance detection methods. Vet. Parasitol. 2002, 103, 183–194. [Google Scholar] [CrossRef]
- Santos, F.O.; Cerqueira, A.P.M.; Branco, A.; Batatinha, M.J.M.; Botura, M.B. Anthelmintic activity of plants against gastrointestinal nematodes of goats: A review. Parasitology 2019, 146, 1233–1246. [Google Scholar] [CrossRef]
- Nour El-Deen, A.; Issa, A. Nematicidal properties of some algal aqueous extracts against root-knot nematode, Meloidogyne incognita in vitro. Egypt. J. Agronematol. 2016, 15, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Eguale, T.; Tilahun, G.; Debella, A.; Feleke, A.; Makonnen, E. Haemonchus contortus: In vitro and in vivo anthelmintic activity of aqueous and hydro-alcoholic extracts of Hedera helix. Exp. Parasitol. 2007, 116, 340–345. [Google Scholar] [CrossRef]
- Lone, B.; Chishti, M.; Bhat, F.; Tak, H.; Bandh, S.; Khan, A. Anthelmintic activities of aqueous and methanol extracts of Prunella vulgaris L. Nat. Prod. Chem. Res. 2017, 5, 269. [Google Scholar] [CrossRef]
- Michalak, I.; Tuhy, Ł.; Chojnacka, K. Seaweed extract by microwave assisted extraction as plant growth biostimulant. Open Chem. 2015, 13, 1183–1195. [Google Scholar] [CrossRef]
- Pais, A.C.; Saraiva, J.A.; Rocha, S.M.; Silvestre, A.J.; Santos, S.A. Current research on the bioprospection of linear diterpenes from Bifurcaria bifurcata: From extraction methodologies to possible applications. Mar. Drugs 2019, 17, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Extract | Concentration (mg/mL) | Geographical Location/Process | Mortality (% ± SD) | p-Value France (Frozen) vs. Portugal (Dried) |
|---|---|---|---|---|---|
| Control | - | 0 | - | 0.065 ± 0.34 | - |
| Bifurcaria bifurcata | Aqueous extract | 3 | France (frozen) | 12 ± 7.15 | 3.1 × 10−5 |
| Portugal (dried) | 0.318 ± 0.74 | ||||
| Heptane | 0.5 | France (frozen) | 100 ± 0.00 | 1 | |
| Portugal (dried) | 99.8 ± 0.51 | ||||
| EtOAc | 1 | France (frozen) | 73.2 ± 17.90 | 1 | |
| Portugal (dried) | 55.3 ± 31.20 | ||||
| BuOH | 3 | France (frozen) | 2.49 ± 3.42 | 1 | |
| Portugal (dried) | 3.54 ± 4.07 | ||||
| Residual aqueous fraction | 3 | France (frozen) | 0 ± 0.00 | nd | |
| Portugal (dried) | 0 ± 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miclon, M.; Courtot, É.; Guégnard, F.; Lenhof, O.; Boudesocque-Delaye, L.; Matard-Mann, M.; Nyvall Collén, P.; Castagnone-Sereno, P.; Neveu, C. The Brown Alga Bifurcaria bifurcata Presents an Anthelmintic Activity on All Developmental Stages of the Parasitic Nematode Heligmosomoides polygyrus bakeri. Pathogens 2023, 12, 540. https://doi.org/10.3390/pathogens12040540
Miclon M, Courtot É, Guégnard F, Lenhof O, Boudesocque-Delaye L, Matard-Mann M, Nyvall Collén P, Castagnone-Sereno P, Neveu C. The Brown Alga Bifurcaria bifurcata Presents an Anthelmintic Activity on All Developmental Stages of the Parasitic Nematode Heligmosomoides polygyrus bakeri. Pathogens. 2023; 12(4):540. https://doi.org/10.3390/pathogens12040540
Chicago/Turabian StyleMiclon, Morgane, Élise Courtot, Fabrice Guégnard, Océane Lenhof, Leslie Boudesocque-Delaye, Maria Matard-Mann, Pi Nyvall Collén, Philippe Castagnone-Sereno, and Cédric Neveu. 2023. "The Brown Alga Bifurcaria bifurcata Presents an Anthelmintic Activity on All Developmental Stages of the Parasitic Nematode Heligmosomoides polygyrus bakeri" Pathogens 12, no. 4: 540. https://doi.org/10.3390/pathogens12040540