
Evaluating the Diagnostic Potential of Chorismate Mutase Poly-Clonal Peptide Antibody for the Acanthamoeba Keratitis in an Animal Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Cell Cultivation
2.2. Encystation Assays
2.3. Immunocytochemistry
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Animals and Ethics Requirements
2.6. Establishment of an Animal Model for Acanthamoeba Keratitis
2.7. Detection of Antigen in AK Animal Model Using CM Antibody
2.8. Statistical Analysis
3. Results
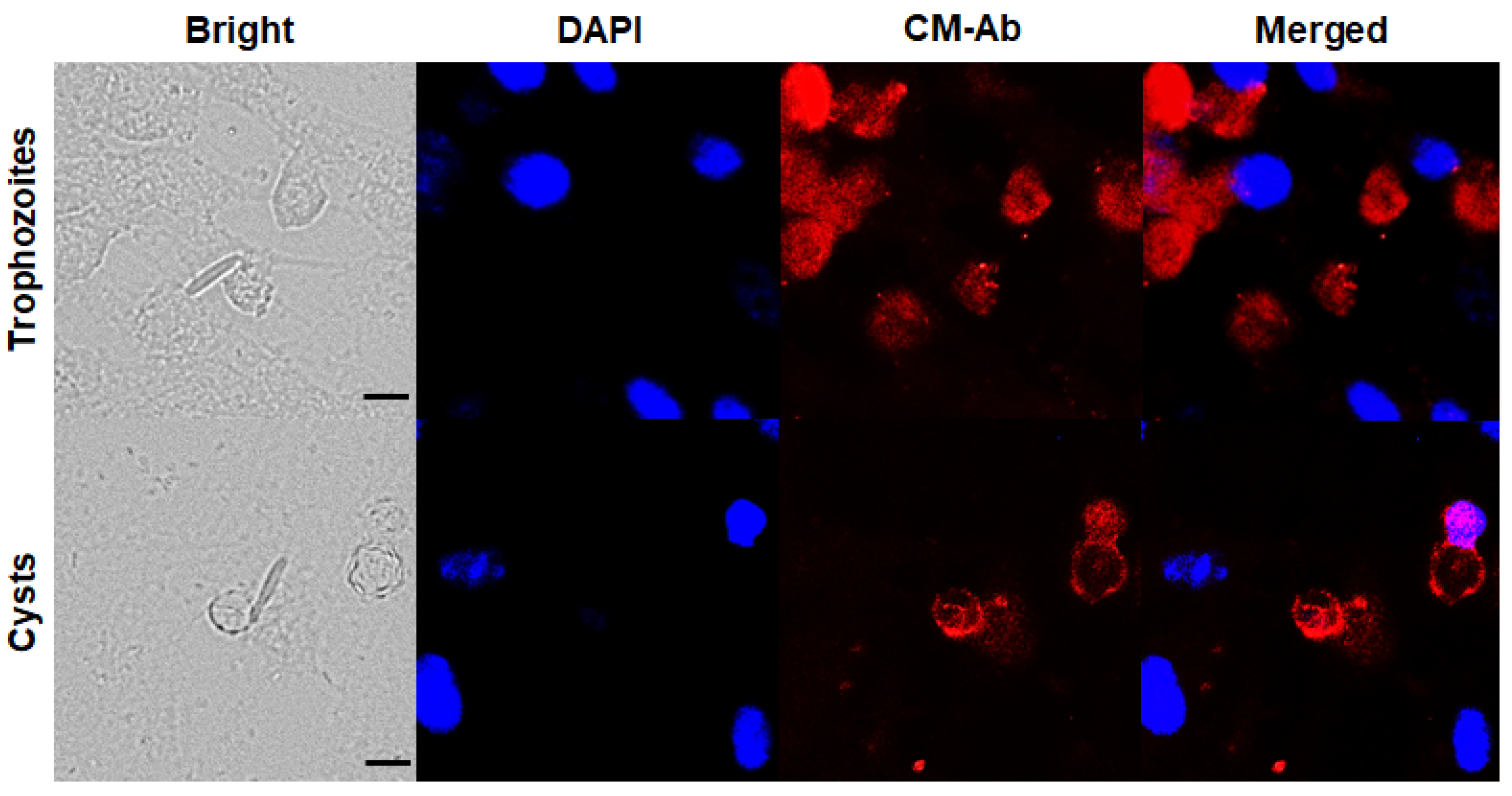
3.1. Confirmation of CM Antibody Specificity against Acanthamoeba Trophozoites and Cysts
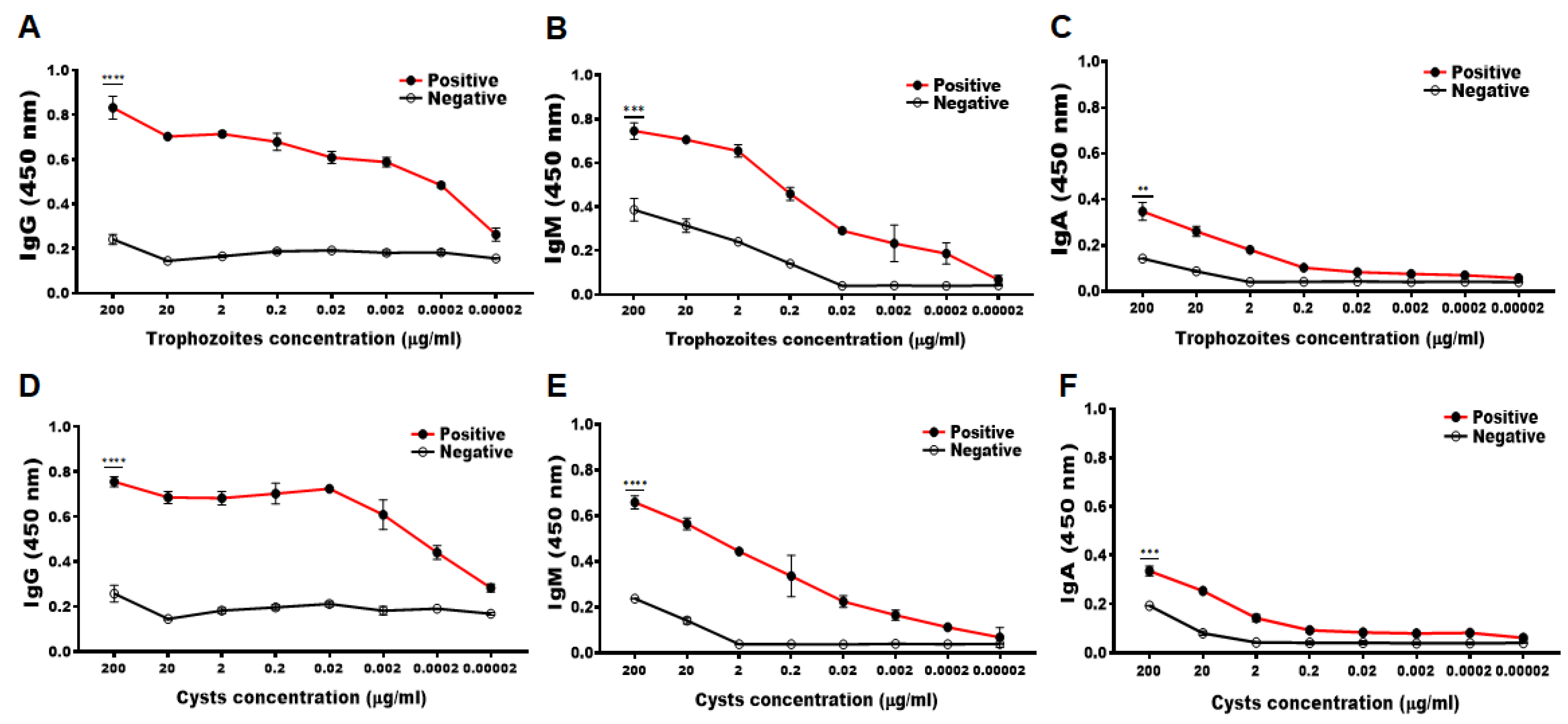
3.2. Determining the Minimum Detectable Concentration of Acanthamoeba Antigens by CM Antibody
3.3. Construction of the AK Animal Model
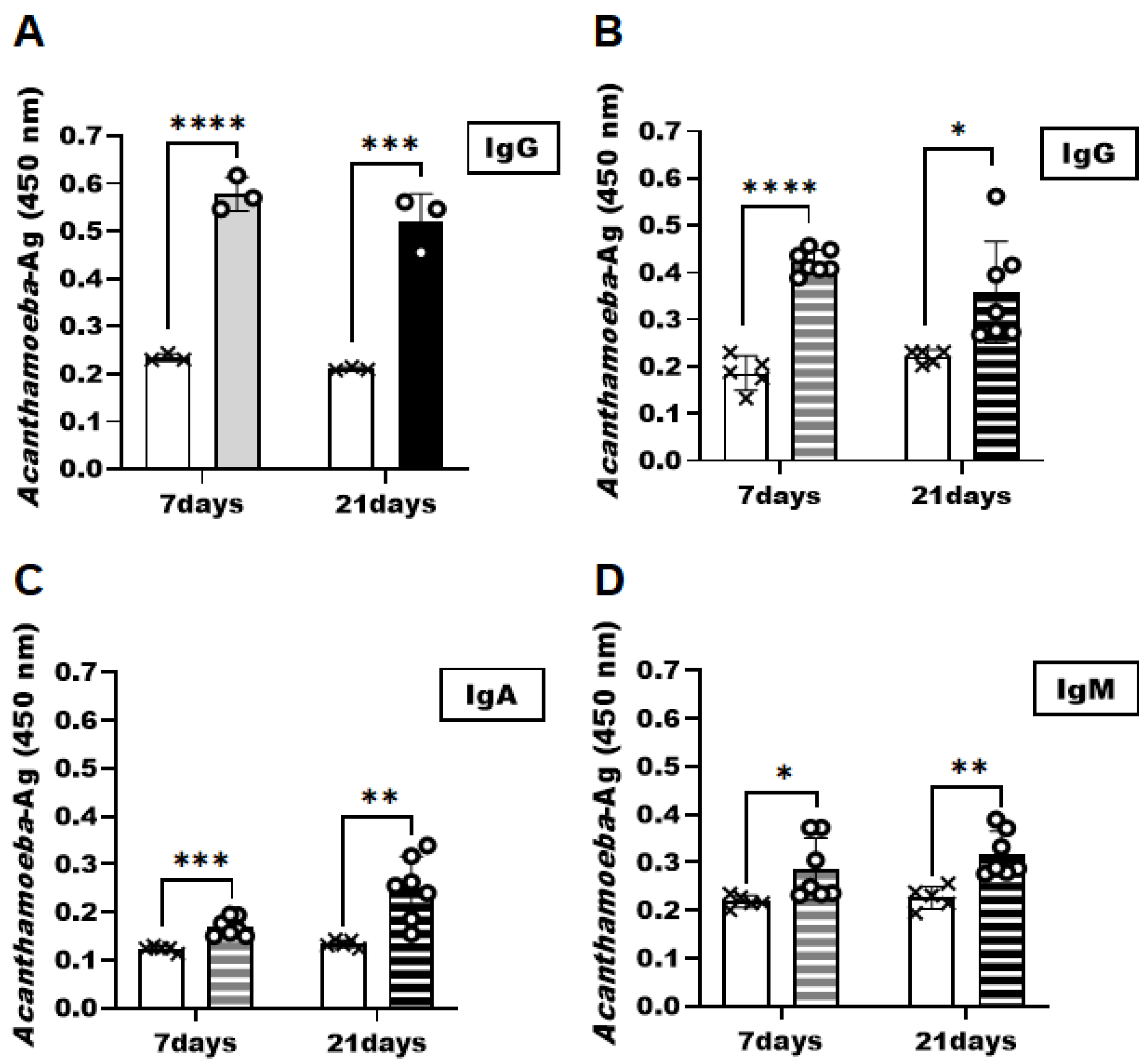
3.4. Detection of Acanthamoeba Antigens in the AK Animal Model
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cooper, E.; Cowmeadow, W.; Elsheikha, H. Should Veterinary Practitioners Be Concerned about Acanthamoeba Keratitis? Parasitologia 2021, 1, 12–19. [Google Scholar] [CrossRef]
- Dart, J.K.; Saw, V.P.; Kilvington, S. Acanthamoeba Keratitis: Diagnosis and Treatment Update 2009. Am. J. Ophthalmol. 2009, 148, 487–499.e2. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, R.; Khan, N.A. Biology and pathogenesis of Acanthamoeba. Parasit Vectors 2012, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Seal, D.V. Acanthamoeba keratitis update—Incidence, molecular epidemiology and new drugs for treatment. Eye 2003, 17, 893–905. [Google Scholar] [CrossRef] [Green Version]
- Fanselow, N.; Sirajuddin, N.; Yin, X.-T.; Huang, A.J.W.; Stuart, P.M. Acanthamoeba Keratitis, Pathology, Diagnosis and Treatment. Pathogens 2021, 10, 323. [Google Scholar] [CrossRef]
- Sharma, S.; Garg, P.; Rao, G.N. Patient characteristics, diagnosis, and treatment of non-contact lens related Acanthamoeba keratitis. Br. J. Ophthalmol. 2000, 84, 1103–1108. [Google Scholar] [CrossRef] [Green Version]
- Reverey, J.F.; Fromme, R.; Leippe, M.; Selhuber-Unkel, C. In vitro adhesion of Acanthamoeba castellanii to soft contact lenses depends on water content and disinfection procedure. Contact Lens Anterior Eye 2014, 37, 262–266. [Google Scholar] [CrossRef]
- Jones, D.B.; Visvesvara, G.S.; Robinson, N.M. Acanthamoeba polyphaga keratitis and Acanthamoeba uveitis associated with fetal meningoencephalitis. Trans. Ophthalmol Soc. UK 1975, 95, 221–232. [Google Scholar] [PubMed]
- Imam, A.M.; El Sheikh, M. Blindness due to Acanthamoeba: First case report from Sudan. Int. J. Health Sci. 2008, 2, 163–166. [Google Scholar]
- Lorenzo-Morales, J.; Martín-Navarro, C.M.; López-Arencibia, A.; Arnalich-Montiel, F.; Piñero, J.E.; Valladares, B. Acanthamoeba keratitis: An emerging disease gathering importance worldwide? Trends Parasitol. 2013, 29, 181–187. [Google Scholar] [CrossRef]
- Hammersmith, K.M. Diagnosis and management of Acanthamoeba keratitis. Curr. Opin. Ophthalmol. 2006, 17, 327–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, T.; Hadas, E.; Iwanicka, I. The duration of the cyst stage and the viability and virulence of Acanthamoeba isolates. Trop. Med. Parasitol. Off. Organ Dtsch. Trop. Ges. Dtsch. Ges. Fur. Tech. Zs. (GTZ) 1995, 46, 106–108. [Google Scholar]
- Ikeda, Y.; Miyazaki, D.; Yakura, K.; Kawaguchi, A.; Ishikura, R.; Inoue, Y.; Mito, T.; Shiraishi, A.; Ohashi, Y.; Higaki, S.; et al. Assessment of Real-Time Polymerase Chain Reaction Detection of Acanthamoeba and Prognosis Determinants of Acanthamoeba Keratitis. Ophthalmology 2012, 119, 1111–1119. [Google Scholar] [CrossRef] [Green Version]
- Joseph, J.; Sharma, S.; Chaurasia, S. Case Report: Corneal Coinfection with Fungus and Amoeba: Report of Two Patients and Literature Review. Am. J. Trop. Med. Hyg. 2018, 99, 805–808. [Google Scholar] [CrossRef]
- Khan, N.A. Acanthamoeba: Biology and increasing importance in human health. FEMS Microbiol. Rev. 2006, 30, 564–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muiño, L.; Rodrigo, D.; Villegas, R.; Romero, P.; Peredo, D.E.; Vargas, R.A.; Liempi, D.; Osuna, A.; Jercic, M.I. Effectiveness of sampling methods employed for Acanthamoeba keratitis diagnosis by culture. Int. Ophthalmol. 2018, 39, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An update on Acanthamoeba keratitis: Diagnosis, pathogenesis and treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daas, L.; Szentmáry, N.; Eppig, T.; Langenbucher, A.; Hasenfus, A.; Roth, M.; Saeger, M.; Nolle, B.; Lippmann, B.; Bohringer, D.; et al. Das Deutsche Akanthamöbenkeratitis-Register. Der. Ophthalmol. 2015, 112, 752–763. [Google Scholar] [CrossRef]
- Pfister, D.R.; Cameron, J.D.; Krachmer, J.H.; Holland, E.J. Confocal Microscopy Findings of Acanthamoeba Keratitis. Am. J. Ophthalmol. 1996, 121, 119–128. [Google Scholar] [CrossRef]
- Szentmáry, N.; Daas, L.; Shi, L.; Laurik, K.L.; Lepper, S.; Milioti, G.; Seitz, B. Acanthamoeba keratitis—Clinical signs, differential diagnosis and treatment. J. Curr. Ophthalmol. 2018, 31, 16–23. [Google Scholar] [CrossRef]
- Yera, H.; Ok, V.; Kuet, F.L.K.; Dahane, N.; Ariey, F.; Hasseine, L.; Delaunay, P.; Martiano, D.; Marty, P.; Bourges, J.L. PCR and culture for diagnosis of Acanthamoeba keratitis. Br. J. Ophthalmol. 2020, 105, 1302–1306. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P.L.; Balczun, C. Failure of molecular diagnostics of a keratitis-inducing Acanthamoeba strain. Exp. Parasitol. 2017, 183, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Cerva, L. Acanthamoeba culbertsoni and Naegleria fowleri: Occurrence of antibodies in man. J. Hyg. Epidemiol. Microbiol. Immunol. 1989, 33, 99–103. [Google Scholar]
- Alouffi, A.S.; Dawoud, T.M.; Almaary, K.S.; Mubarak, A.S.; JarAllah, K.; Matin, A. First report of detection of IgA anti-Acanthamoeba antibodies among Saudi population and amoeba isolation from their surroundings. Trop. Biomed. 2021, 38, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, H.; Apte, S.; El-Agha, M.-S.H.; Li, L.; Hurt, M.; Howard, K.; Cavanagh, H.D.; McCulley, J.P.; Niederkorn, J.Y. Tear IgA and Serum IgG Antibodies Against Acanthamoeba in Patients With Acanthamoeba Keratitis. Cornea 2001, 20, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Greenman, J.; Topping, K.P.; Hough, V.C.; Temple, G.S.; Paget, T.A. Isolation of Acanthamoeba-Specific Antibodies from a Bacteriophage Display Library. J. Clin. Microbiol. 2000, 38, 2374–2377. [Google Scholar] [CrossRef]
- Turner, M.L.; Cockerell, E.J.; Brereton, H.M.; Badenoch, P.R.; Tea, M.; Coster, D.J.; Williams, K.A. Antigens of selected Acanthamoeba species detected with monoclonal antibodies. Int. J. Parasitol. 2005, 35, 981–990. [Google Scholar] [CrossRef]
- Kim, M.-J.; Ham, A.-J.; Park, A.-Y.; Sohn, H.-J.; Shin, H.-J.; Quan, F.-S.; Kong, H.-H.; Moon, E.-K. Detection of Acanthamoeba from Acanthamoeba Keratitis Mouse Model Using Acanthamoeba-Specific Antibodies. Microorganisms 2022, 10, 1711. [Google Scholar] [CrossRef]
- Lee, H.-A.; Chu, K.-B.; Kim, M.-J.; Quan, F.-S.; Kong, H.-H.; Moon, E.-K. Chorismate mutase peptide antibody enables specific detection of Acanthamoeba. PLoS ONE 2021, 16, e0250342. [Google Scholar] [CrossRef]
- Bowers, B.; Korn, E.D. The fine structure of Acanthamoeba castellanii (neff strain). J. Cell Biol. 1969, 41, 786–805. [Google Scholar] [CrossRef]
- Kang, H.; Sohn, H.-J.; Park, A.-Y.; Ham, A.-J.; Lee, J.-H.; Oh, Y.-H.; Chwae, Y.-J.; Kim, K.; Park, S.; Yang, H.; et al. Establishment of an Acanthamoeba keratitis mouse model confirmed by amoebic DNA amplification. Sci. Rep. 2021, 11, 4183. [Google Scholar] [CrossRef]
- Kim, M.-J.; Lee, H.-A.; Quan, F.-S.; Kong, H.-H.; Moon, E.-K. Characterization of a Peptide Antibody Specific to the Adenylyl Cyclase-Associated Protein of Acanthamoeba castellanii. Korean J. Parasitol. 2022, 60, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Quan, F.-S.; Kong, H.-H.; Kim, J.-H.; Moon, E.-K. Specific Detection of Acanthamoeba species using Polyclonal Peptide Antibody Targeting the Periplasmic Binding Protein of A. castellanii. Korean J. Parasitol. 2022, 60, 143–147. [Google Scholar] [CrossRef] [PubMed]
- de Lacerda, A.G.; Lira, M. Acanthamoeba keratitis: A review of biology, pathophysiology and epidemiology. Ophthalmic Physiol. Opt. 2020, 41, 116–135. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, R.; Aqeel, Y.; Khan, N.A. The use of dimethyl sulfoxide in contact lens disinfectants is a potential preventative strategy against contracting Acanthamoeba keratitis. Contact Lens Anterior Eye 2016, 39, 389–393. [Google Scholar] [CrossRef]
- Niederkorn, J.Y.; Alizadeh, H.; Leher, H.; McCulley, J.P. The pathogenesis of Acanthamoeba keratitis. Microbes Infect. 1999, 1, 437–443. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-J.; Jo, H.-J.; Sohn, H.-J.; Shin, H.-J.; Quan, F.-S.; Kong, H.-H.; Moon, E.-K. Evaluating the Diagnostic Potential of Chorismate Mutase Poly-Clonal Peptide Antibody for the Acanthamoeba Keratitis in an Animal Model. Pathogens 2023, 12, 526. https://doi.org/10.3390/pathogens12040526
Kim M-J, Jo H-J, Sohn H-J, Shin H-J, Quan F-S, Kong H-H, Moon E-K. Evaluating the Diagnostic Potential of Chorismate Mutase Poly-Clonal Peptide Antibody for the Acanthamoeba Keratitis in an Animal Model. Pathogens. 2023; 12(4):526. https://doi.org/10.3390/pathogens12040526
Chicago/Turabian StyleKim, Min-Jeong, Hye-Jeong Jo, Hae-Jin Sohn, Ho-Joon Shin, Fu-Shi Quan, Hyun-Hee Kong, and Eun-Kyung Moon. 2023. "Evaluating the Diagnostic Potential of Chorismate Mutase Poly-Clonal Peptide Antibody for the Acanthamoeba Keratitis in an Animal Model" Pathogens 12, no. 4: 526. https://doi.org/10.3390/pathogens12040526