
Geographical, Temporal and Host-Species Distribution of Potentially Human-Pathogenic Group B Streptococcus in Aquaculture Species in Southeast Asia
 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
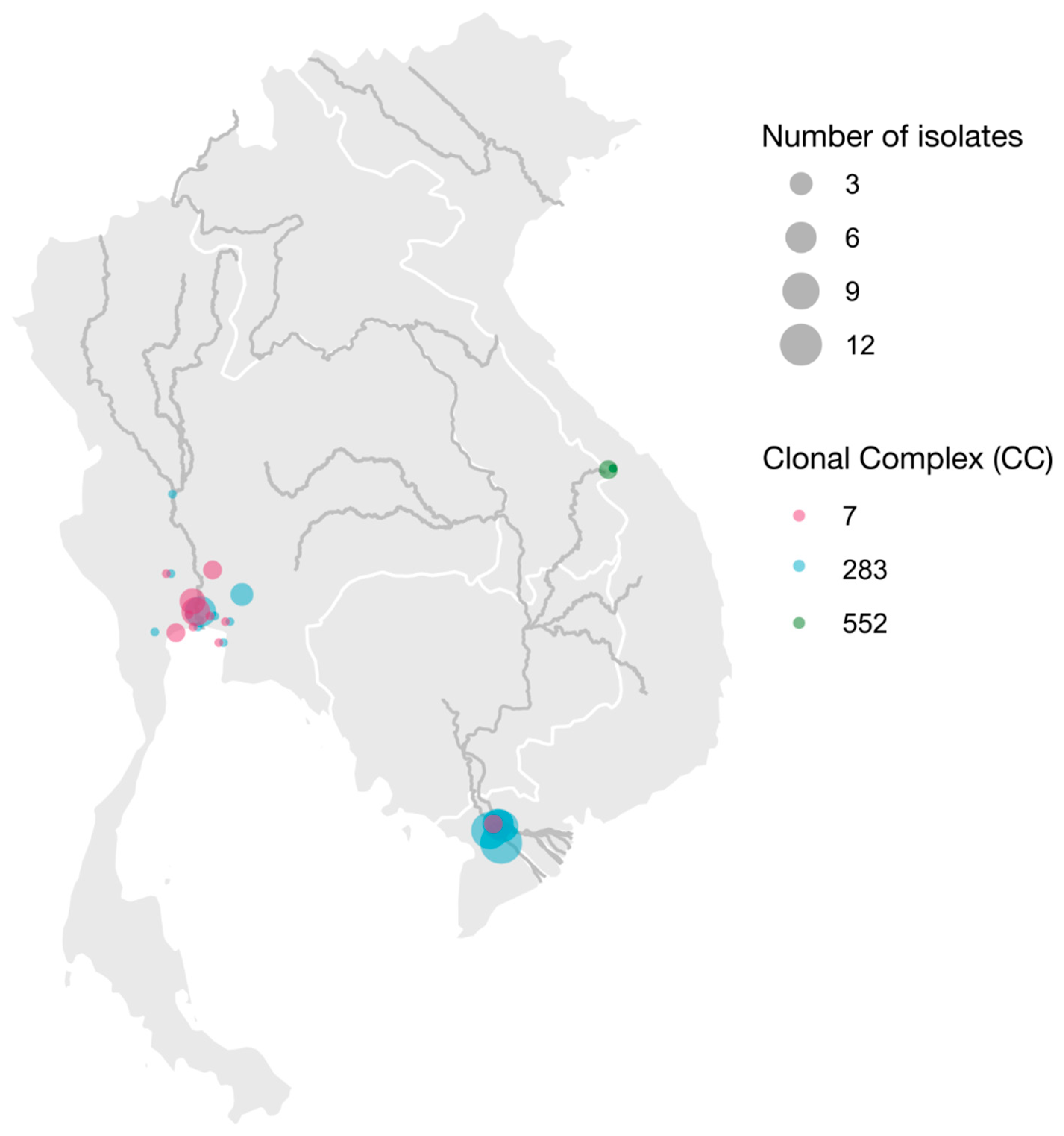
2.1. Samples and Isolates
2.2. Molecular Identification
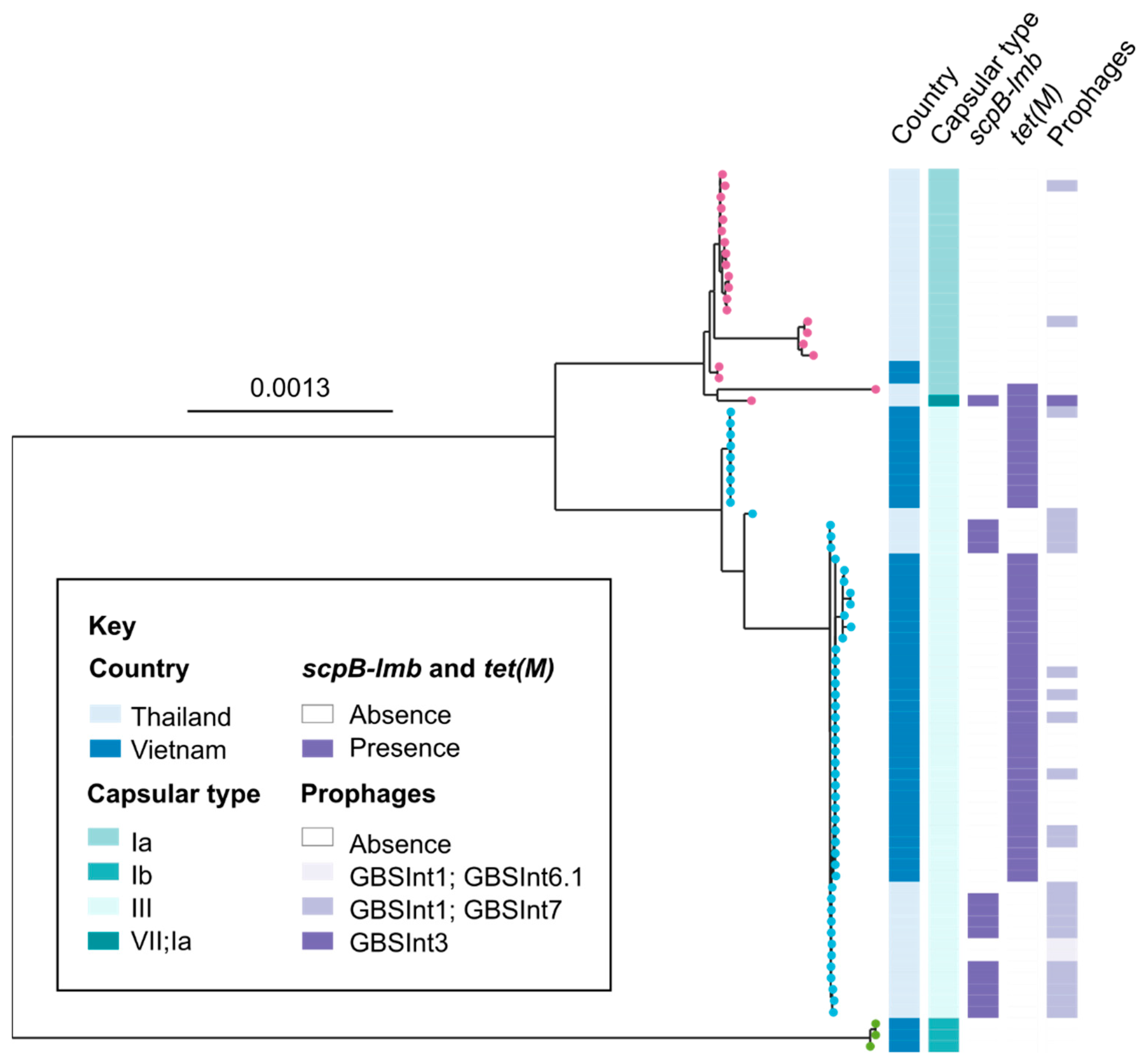
2.3. Genetic and Genomic Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Da Cunha, V.; Davies, M.R.; Douarre, P.E.; Rosinski-Chupin, I.; Margarit, I.; Spinali, S.; Perkins, T.; Lechat, P.; Dmytruk, N.; Sauvage, E.; et al. Streptococcus agalactiae clones infecting humans were selected and fixed through the extensive use of tetracycline. Nat. Commun. 2014, 5, 4544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, B.P.; Procter, S.R.; Paul, P.; Chandna, J.; Lewin, A.; Seedat, F.; Koukounari, A.; Dangor, Z.; Leahy, S.; Santhanam, S.; et al. Group B Streptococcus infection during pregnancy and infancy: Estimates of regional and global burden. Lancet Glob. Health 2022, 10, e807–e819. [Google Scholar] [CrossRef] [PubMed]
- Skoff, T.H.; Farley, M.M.; Petit, S.; Craig, A.S.; Schaffner, W.; Gershman, K.; Harrison, L.H.; Lynfield, R.; Mohle-Boetani, J.; Zansky, S.; et al. Increasing burden of invasive group B streptococcal disease in nonpregnant adults, 1990–2007. Clin. Infect. Dis. 2009, 49, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.M.; Shetty, N.; Guy, R.; Nyaga, V.N.; Bull, A.; Richards, M.J.; van der Kooi, T.I.; Koek, M.B.; De Almeida, M.; Roberts, S.A.; et al. Group B Streptococcus in surgical site and non-invasive bacterial infections worldwide: A systematic review and meta-analysis. Int. J. Infect. Dis. 2019, 83, 116–129. [Google Scholar] [CrossRef] [Green Version]
- Ip, M.; Cheuk, E.S.; Tsui, M.H.; Kong, F.; Leung, T.N.; Gilbert, G.L. Identification of a Streptococcus agalactiae serotype III subtype 4 clone in association with adult invasive disease in Hong Kong. J. Clin. Microbiol. 2006, 44, 4252–4254. [Google Scholar] [CrossRef] [Green Version]
- Barkham, T.; Zadoks, R.N.; Azmai, M.N.A.; Baker, S.; Bich, V.T.N.; Chalker, V.; Chau, M.L.; Dance, D.; Deepak, R.N.; van Doorn, H.R.; et al. One hypervirulent clone, sequence type 283, accounts for a large proportion of invasive Streptococcus agalactiae isolated from humans and diseased tilapia in Southeast Asia. PLoS Negl. Trop. Dis. 2019, 13, e0007421. [Google Scholar] [CrossRef] [Green Version]
- Kalimuddin, S.; Chen, S.L.; Lim, C.T.; Koh, T.H.; Tan, T.Y.; Kam, M.; Wong, C.W.; Mehershahi, K.S.; Chau, M.L.; Ng, L.C.; et al. 2015 epidemic of severe Streptococcus agalactiae Sequence Type 283 infections in Singapore associated with the consumption of raw freshwater fish: A detailed analysis of clinical, epidemiological, and bacterial sequencing data. Clin. Infect. Dis. 2017, 64 (Suppl. S2), S145–S152. [Google Scholar] [CrossRef] [Green Version]
- Wilder-Smith, E.; Chow, K.M.; Kay, R.; Ip, M.; Tee, N. Group B streptococcal meningitis in adults: Recent increase in Southeast Asia. Aust. N. Z. J. Med. 2000, 30, 462–465. [Google Scholar] [CrossRef]
- van Kassel, M.N.; Janssen, S.; Kofman, S.; Brouwer, M.C.; van de Beek, D.; Bijlsma, M.W. Prevalence of group B streptococcal colonization in the healthy non-pregnant population: A systematic review and meta-analysis. Clin. Microbiol. Infect. 2021, 27, 968–980. [Google Scholar] [CrossRef]
- Chau, M.L.; Chen, S.L.; Yap, M.; Hartantyo, S.H.; Chiew, P.K.; Fernandez, C.J.; Wong, W.K.; Fong, R.K.; Tan, W.L.; Tan, B.Z.; et al. Group B Streptococcus infections caused by improper sourcing and handling of fish for raw consumption, Singapore, 2015–2016. Emerg. Infect. Dis. 2017, 23, 2002. [Google Scholar] [CrossRef] [Green Version]
- Rajendram, P.; Kyaw, W.M.; Leo, Y.S.; Ho, H.; Chen, W.K.; Lin, R.; Pratim, P.; Badaruddin, H.; Ang, B.; Barkham, T.; et al. Group B Streptococcus Sequence Type 283 disease linked to consumption of raw fish, Singapore. Emerg. Infect. Dis. 2016, 22, 1974–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amborski, R.L.; Snider, T.G., 3rd; Thune, R.L.; Culley, D.D., Jr. A non-hemolytic, group B Streptococcus infection of cultured bullfrogs, Rana catesbeiana, in Brazil. J. Wildl. Dis. 1983, 19, 180–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.J.; Klesius, P.H.; Gilbert, P.M.; Shoemaker, C.A.; Al Sarawi, M.A.; Landsberg, J.; Duremdez, R.; Al Marzouk, A.; Al Zenki, S. Characterization of β-haemolytic Group B Streptococcus agalactiae in cultured seabream, Sparus auratus L., and wild mullet, Liza klunzingeri (Day), in Kuwait. J. Fish Dis. 2002, 25, 505–513. [Google Scholar] [CrossRef]
- Duremdez, R.; Al-Marzouk, A.; Qasem, J.A.; Al-Harbi, A.; Gharabally, H. Isolation of Streptococcus agalactiae from cultured silver pomfret, Pampus argenteus (Euphrasen), in Kuwait. J. Fish Dis. 2004, 27, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Hernández, E.; Figueroa, J.; Iregui, C. Streptococcosis on a red tilapia, Oreochromis sp., farm: A case study. J. Fish Dis. 2009, 32, 247–252. [Google Scholar] [CrossRef]
- Zamri-Saad, M.; Amal, M.N.; Siti-Zahrah, A. Pathological changes in red tilapias (Oreochromis spp.) naturally infected by Streptococcus agalactiae. J. Comp. Pathol. 2010, 143, 227–229. [Google Scholar] [CrossRef]
- Delannoy, C.M.; Crumlish, M.; Fontaine, M.C.; Pollock, J.; Foster, G.; Dagleish, M.P.; Turnbull, J.F.; Zadoks, R.N. Human Streptococcus agalactiae strains in aquatic mammals and fish. BMC Microbiol. 2013, 13, 41. [Google Scholar] [CrossRef] [Green Version]
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Jones, N.; Bohnsack, J.F.; Takahashi, S.; Oliver, K.A.; Chan, M.S.; Kunst, F.; Glaser, P.; Rusniok, C.; Crook, D.W.; Harding, R.M.; et al. Multilocus sequence typing system for group B Streptococcus. J. Clin. Microbiol. 2003, 41, 2530–2536. [Google Scholar] [CrossRef] [Green Version]
- Tiruvayipati, S.; Tang, W.Y.; Barkham, T.M.S.; Chen, S.L. GBS-SBG—GBS Serotyping by genome sequencing. Microb. Genom. 2021, 7, 2021–2106. [Google Scholar] [CrossRef]
- Kong, F.; Gowan, S.; Martin, D.; James, G.; Gilbert, G.L. Serotype identification of group B streptococci by PCR and sequencing. J. Clin. Microbiol. 2002, 40, 216–226. [Google Scholar] [CrossRef] [Green Version]
- Barony, G.M.; Tavares, G.C.; Pereira, F.L.; Carvalho, A.F.; Dorella, F.A.; Leal, C.A.G.; Figueiredo, H.C.P. Large-scale genomic analyses reveal the population structure and evolutionary trends of Streptococcus agalactiae strains in Brazilian fish farms. Sci. Rep. 2017, 7, 13538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, M.; Delamare-Deboutteville, J.; Bowater, R.O.; Walker, M.J.; Beatson, S.; Zakour, N.L.B.; Barnes, A.C. Microevolution of Streptococcus agalactiae ST-261 from Australia indicates dissemination via imported tilapia and ongoing adaptation to marine hosts or environment. Appl. Environ. Microbiol. 2018, 84, e00859–e00918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.J.; Bohnsack, J.F.; Klesius, P.H.; Whiting, A.A.; Garcia, J.C.; Shoemaker, C.A.; Takahashi, S. Phylogenetic relationships among Streptococcus agalactiae isolated from piscine, dolphin, bovine and human sources: A dolphin and piscine lineage associated with a fish epidemic in Kuwait is also associated with human neonatal infections in Japan. J. Med. Microbiol. 2008, 57 Pt 11, 1369–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, C.A.G.; Queiroz, G.A.; Pereira, F.L.; Tavares, G.C.; Figueiredo, H.C.P. Streptococcus agalactiae Sequence Type 283 in farmed fish, Brazil. Emerg. Infect. Dis. 2019, 25, 776–779. [Google Scholar] [CrossRef] [Green Version]
- Rosinski-Chupin, I.; Sauvage, E.; Mairey, B.; Mangenot, S.; Ma, L.; Da Cunha, V.; Rusniok, C.; Bouchier, C.; Barbe, V.; Glaser, P. Reductive evolution in Streptococcus agalactiae and the emergence of a host adapted lineage. BMC Genom. 2013, 14, 252. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.J.; Klesius, P.H.; Pasnik, D.J.; Bohnsack, J.F. Human Streptococcus agalactiae isolate in Nile tilapia (Oreochromis niloticus). Emerg. Infect. Dis. 2009, 15, 774–776. [Google Scholar] [CrossRef]
- Aiewsakun, P.; Ruangchai, W.; Thawornwattana, Y.; Jaemsai, B.; Mahasirimongkol, S.; Homkaew, A.; Suksomchit, P.; Dubbs, P.; Palittapongarnpim, P. Genomic epidemiology of Streptococcus agalactiae ST283 in Southeast Asia. Sci. Rep. 2022, 12, 4185. [Google Scholar] [CrossRef]
- Kayansamruaj, P.; Pirarat, N.; Kondo, H.; Hirono, I.; Rodkhum, C. Genomic comparison between pathogenic Streptococcus agalactiae isolated from Nile tilapia in Thailand and fish-derived ST7 strains. Infect. Genet. Evol. 2015, 36, 307–314. [Google Scholar] [CrossRef]
- Tint, K.K.; Ngin, K.; Sapari, A.; Souliphone, K.; Suwannapoom, S.; Viron, J.G.; Thanh, V.T.P.; Chumchuen, S.V. Fish trade practices: Southeast Asian perspective. In Fish for the People; Southeast Asian Fisheries Development Center: Bangkok, Thailand, 2020; Volume 18, pp. 9–20. [Google Scholar]
- FAO. Risk Profile—Group B Streptococcus (GBS)/Streptococcus Agalactiae Sequence Type (ST) 283 in Freshwater Fish; FAO: Bangkok, Thailand, 2021. [Google Scholar]
- Palang, I.; Hirono, I.; Senapin, S.; Sirimanapong, W.; Withyachumnarnkul, B.; Vanichviriyakit, R. Cytotoxicity of Streptococcus agalactiae secretory protein on tilapia cultured cells. J. Fish Dis. 2020, 43, 1229–1236. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [Green Version]
- Inouye, M.; Dashnow, H.; Raven, L.A.; Schultz, M.B.; Pope, B.J.; Tomita, T.; Zobel, J.; Holt, K.E. SRST2: Rapid genomic surveillance for public health and hospital microbiology labs. Genome Med. 2014, 6, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Metcalf, B.J.; Chochua, S.; Gertz, R.E., Jr.; Hawkins, P.A.; Ricaldi, J.; Li, Z.; Walker, H.; Tran, T.; Rivers, J.; Mathis, S.; et al. Short-read whole genome sequencing for determination of antimicrobial resistance mechanisms and capsular serotypes of current invasive Streptococcus agalactiae recovered in the USA. Clin. Microbiol. Infect. 2017, 23, 574.e7–574.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheppard, A.E.; Vaughan, A.; Jones, N.; Turner, P.; Turner, C.; Efstratiou, A.; Patel, D.; Walker, A.S.; Berkley, J.A.; Crook, D.W.; et al. Capsular typing method for Streptococcus agalactiae using whole-genome sequence data. J. Clin. Microbiol. 2016, 54, 1388–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crestani, C.; Forde, T.L.; Zadoks, R.N. Development and application of a prophage integrase typing scheme for Group B Streptococcus. Front. Microbiol. 2020, 11, 1993. [Google Scholar] [CrossRef] [PubMed]
- Tonkin-Hill, G.; MacAlasdair, N.; Ruis, C.; Weimann, A.; Horesh, G.; Lees, J.A.; Gladstone, R.A.; Lo, S.; Beaudoin, C.; Floto, R.A.; et al. Producing polished prokaryotic pangenomes with the Panaroo pipeline. Genome Biol. 2020, 21, 180. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Dance, D.; Zadoks, R.N.; Luangraj, M.; Simpson, A.; Chen, S.L.; Barkham, T. Community-acquired Group B streptococcal meningitis in adults. J. Infect. 2020, 81, 147–178. [Google Scholar] [CrossRef]
- Paveenkittiporn, W.; Ungcharoen, R.; Kerdsin, A. Streptococcus agalactiae infections and clinical relevance in adults, Thailand. Diagn. Microbiol. Infect. Dis. 2020, 97, 115005. [Google Scholar] [CrossRef]
- Vasikasin, V.; Changpradub, D. Clinical manifestations and prognostic factors for Streptococcus agalactiae bacteremia among nonpregnant adults in Thailand. J. Infect. Chemother. 2021, 27, 967–971. [Google Scholar] [CrossRef]
- Sørensen, U.B.; Poulsen, K.; Ghezzo, C.; Margarit, I.; Kilian, M. Emergence and global dissemination of host-specific Streptococcus agalactiae clones. MBio 2010, 1, e00178–e00210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, A.; Berg, B. Wastewater and aquatic biotopes are potential reservoirs of group B streptococci. Dan. Vet. J. 1982, 65, 197–200. [Google Scholar]
- Kayansamruaj, P.; Soontara, C.; Unajak, S.; Dong, H.T.; Rodkhum, C.; Kondo, H.; Hirono, I.; Areechon, N. Comparative genomics inferred two distinct populations of piscine pathogenic Streptococcus agalactiae, serotype Ia ST7 and serotype III ST283, in Thailand and Vietnam. Genomics 2019, 111, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Jafar, Q.A.; Sameer, A.Z.; Salwa, A.M.; Samee, A.A.; Ahmed, A.M.; Al-Sharifi, F. Molecular investigation of Streptococcus agalactiae isolates from environmental samples and fish specimens during a massive fish kill in Kuwait Bay. Pak. J. Biol. Sci. 2008, 11, 2500–2504. [Google Scholar]
- Niu, G.; Khattiya, R.; Zhang, T.; Boonyayatra, S.; Wongsathein, D. Phenotypic and genotypic characterization of Streptococcus spp. isolated from tilapia (Oreochromis spp.) cultured in river-based cage and earthen ponds in Northern Thailand. J. Fish Dis. 2020, 43, 391–398. [Google Scholar] [CrossRef]
- Verner-Jeffreys, D.W.; Wallis, T.J.; Cejas, I.C.; Ryder, D.; Haydon, D.J.; Domazoro, J.F.; Dontwi, J.; Field, T.R.; Adjei-Boteng, D.; Wood, G.; et al. Streptococcus agalactiae Multilocus sequence type 261 is associated with mortalities in the emerging Ghanaian tilapia industry. J. Fish Dis. 2018, 41, 175–179. [Google Scholar] [CrossRef]
- Li, C.; Sapugahawatte, D.N.; Yang, Y.; Wong, K.T.; Lo, N.W.S.; Ip, M. Multidrug-resistant Streptococcus agalactiae strains found in human and fish with high penicillin and cefotaxime non-susceptibilities. Microorganisms 2020, 8, 1055. [Google Scholar] [CrossRef]
- Chu, C.; Huang, P.Y.; Chen, H.M.; Wang, Y.H.; Tsai, I.A.; Lu, C.C.; Chen, C.C. Genetic and pathogenic difference between Streptococcus agalactiae serotype Ia fish and human isolates. BMC Microbiol. 2016, 16, 175. [Google Scholar] [CrossRef] [Green Version]
- Pu, W.; Wang, Y.; Yang, N.; Guo, G.; Li, H.; Li, Q.; Rehman, N.U.; Zheng, L.; Wang, P.; Han, S.; et al. Investigation of Streptococcus agalactiae using pcsB-based LAMP in milk, tilapia and vaginal swabs in Haikou, China. J. Appl. Microbiol. 2020, 128, 784–793. [Google Scholar] [CrossRef]
- Legario, F.S.; Choresca, C.H., Jr.; Turnbull, J.F.; Crumlish, M. Isolation and molecular characterization of streptococcal species recovered from clinical infections in farmed Nile tilapia (Oreochromis niloticus) in the Philippines. J. Fish Dis. 2020, 43, 1431–1442. [Google Scholar] [CrossRef]
- Richards, V.P.; Velsko, I.M.; Alam, T.; Zadoks, R.N.; Manning, S.D.; Bitar, P.D.P.; Hasler, H.B.; Crestani, C.; Springer, G.; Probert, B.; et al. Population gene introgression and high genome plasticity for the zoonotic pathogen Streptococcus agalactiae. Mol. Biol. Evol. 2019, 36, 2572–2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schar, D.; Klein, E.Y.; Laxminarayan, R.; Gilbert, M.; Van Boeckel, T.P. Global trends in antimicrobial use in aquaculture. Sci. Rep. 2020, 10, 21878. [Google Scholar] [CrossRef] [PubMed]
- Schar, D.; Zhao, C.; Wang, Y.; Larsson, D.G.J.; Gilbert, M.; Van Boeckel, T.P. Twenty-year trends in antimicrobial resistance from aquaculture and fisheries in Asia. Nat. Commun. 2021, 12, 5384. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirimanapong, W.; Phước, N.N.; Crestani, C.; Chen, S.; Zadoks, R.N. Geographical, Temporal and Host-Species Distribution of Potentially Human-Pathogenic Group B Streptococcus in Aquaculture Species in Southeast Asia. Pathogens 2023, 12, 525. https://doi.org/10.3390/pathogens12040525
Sirimanapong W, Phước NN, Crestani C, Chen S, Zadoks RN. Geographical, Temporal and Host-Species Distribution of Potentially Human-Pathogenic Group B Streptococcus in Aquaculture Species in Southeast Asia. Pathogens. 2023; 12(4):525. https://doi.org/10.3390/pathogens12040525
Chicago/Turabian StyleSirimanapong, Wanna, Nguyễn Ngọc Phước, Chiara Crestani, Swaine Chen, and Ruth N. Zadoks. 2023. "Geographical, Temporal and Host-Species Distribution of Potentially Human-Pathogenic Group B Streptococcus in Aquaculture Species in Southeast Asia" Pathogens 12, no. 4: 525. https://doi.org/10.3390/pathogens12040525