
Persistence of Tembusu Virus in Culex tritaeniorhynchus in Yunnan Province, China

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Collection
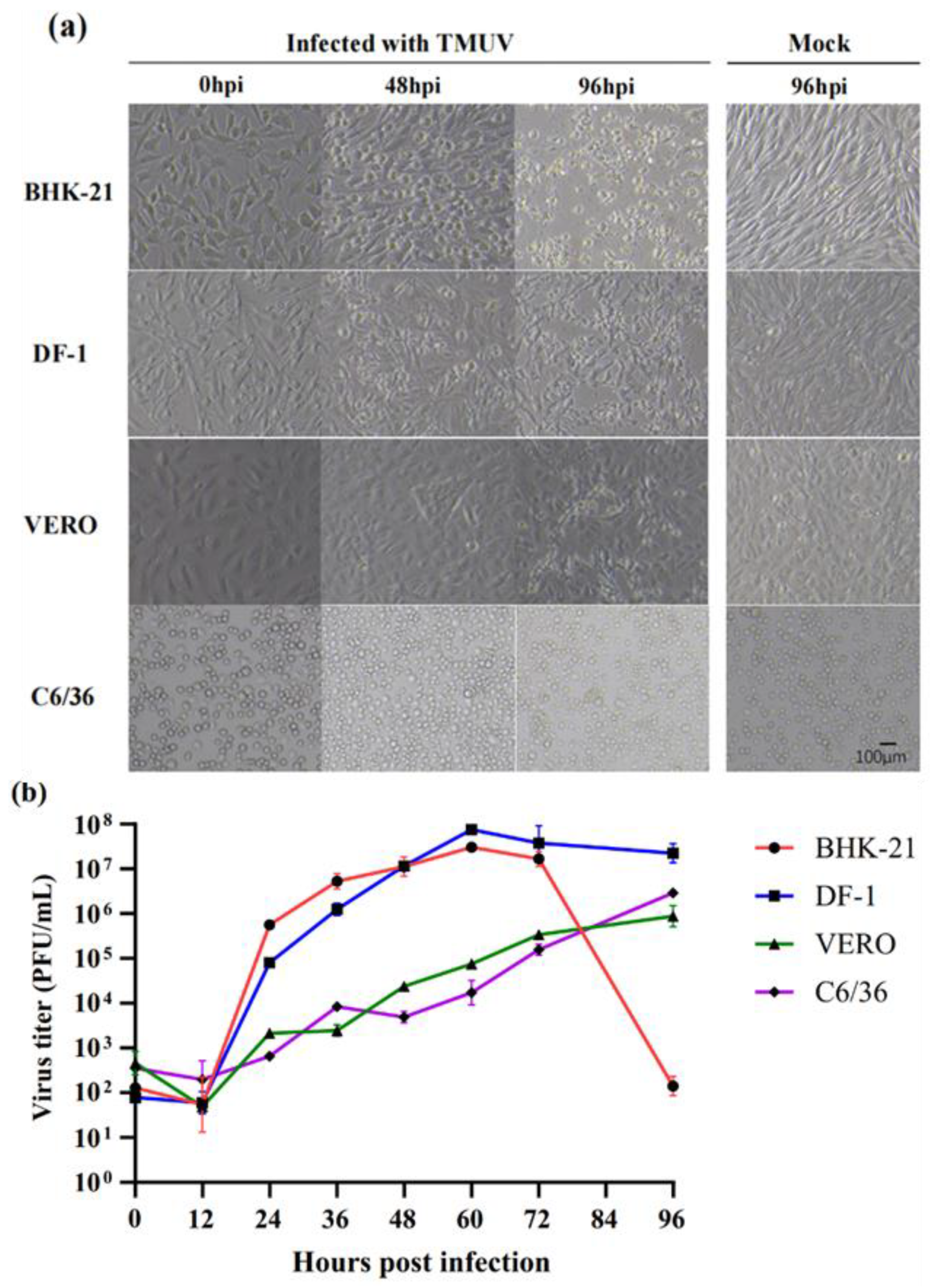
2.2. Cell Lines
2.3. Virus Isolation and RT-PCR Analysis
2.4. Viral Titer Determination
2.5. Virus Growth Kinetics
2.6. Virus Genome Sequencing
2.7. Sequence and Phylogenetic Analysis
2.8. Bayesian Phylogenetic Reconstruction and Time to the Most Recent Common Ancestry (tMRCA)
3. Results
3.1. Mosquito Collection
3.2. Virus Isolation and Identification
3.3. Virus Growth Kinetics
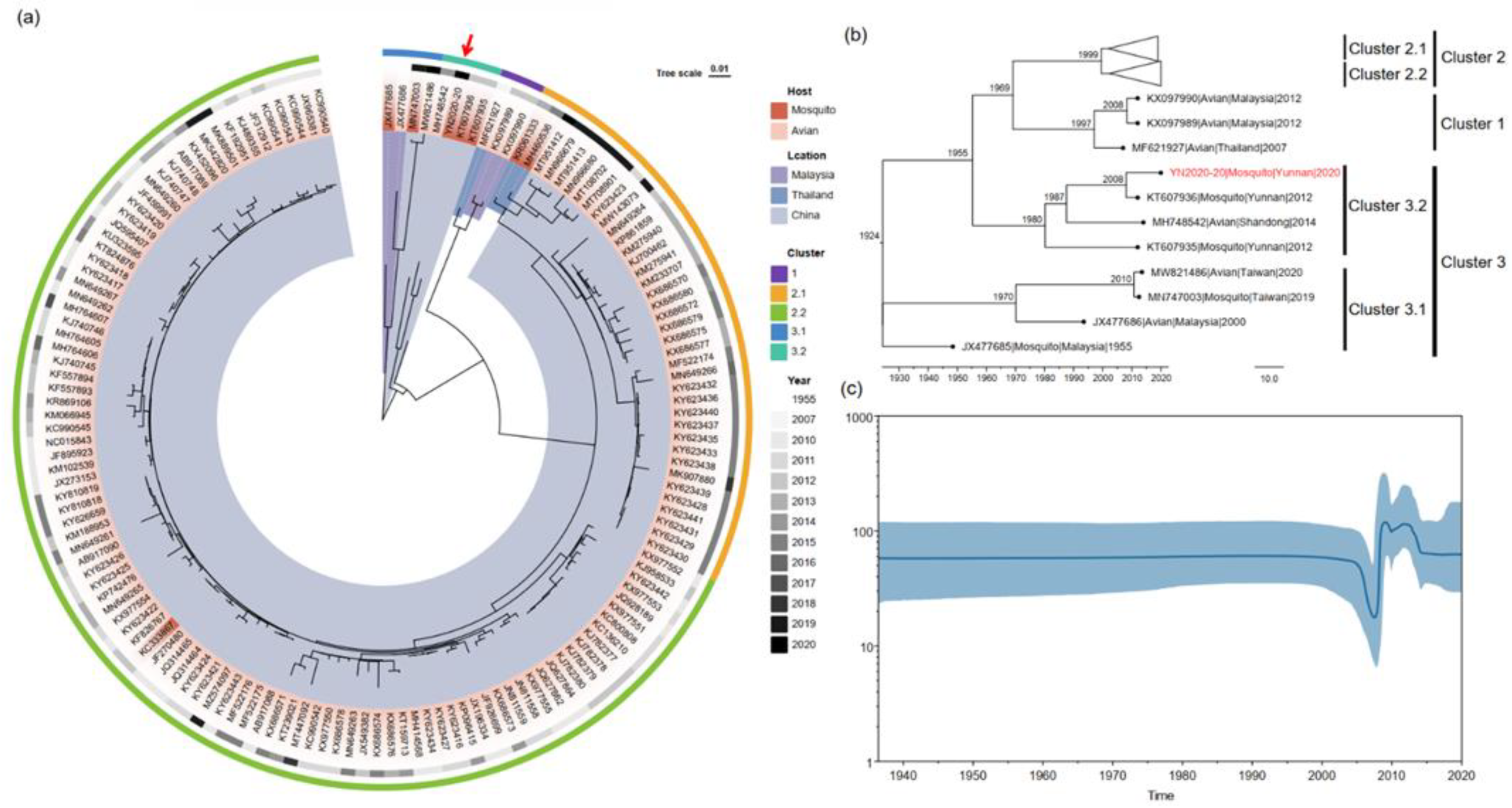
3.4. Phylogenetic Analysis
3.5. Estimation of Divergent Time of TMUVs
3.6. Genome Sequence Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Platt, G.S.; Way, H.J.; Bowen, E.T.; Simpson, D.I.; Hill, M.N.; Kamath, S.; Bendell, P.J.; Heathcote, O.H. Arbovirus infections in Sarawak, October 1968–February 1970 Tembusu and Sindbis virus isolations from mosquitoes. Ann. Trop. Med. Parasitol. 1975, 69, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.D.; Karabatsos, N.; Cropp, B.; Tagaki, M.; Tsuda, Y.; Ichinose, A.; Igarashi, A. Identification of a flavivirus isolated from mosquitos in Chiang Mai Thailand. Southeast Asian J. Trop. Med. Public Health 1999, 30, 161–165. [Google Scholar] [PubMed]
- Leake, C.J.; Ussery, M.A.; Nisalak, A.; Hoke, C.; Andre, R.; Burke, D. Virus isolations from mosquitoes collected during the 1982 Japanese encephalitis epidemic in northern Thailand. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Kono, Y.; Tsukamoto, K.; Abd Hamid, M.; Darus, A.; Lian, T.C.; Sam, L.S.; Yok, C.N.; Di, K.B.; Lim, K.T.; Yamaguchi, S.; et al. Encephalitis and retarded growth of chicks caused by Sitiawan virus, a new isolate belonging to the genus Flavivirus. Am. J. Trop. Med. Hyg. 2000, 63, 94–101. [Google Scholar] [CrossRef]
- Su, J.; Li, S.; Hu, X.; Yu, X.; Wang, Y.; Liu, P.; Lu, X.; Zhang, G.; Hu, X.; Liu, D.; et al. Duck egg-drop syndrome caused by BYD virus, a new Tembusu-related flavivirus. PLoS ONE 2011, 6, 18106. [Google Scholar] [CrossRef] [Green Version]
- Homonnay, Z.G.; Kovács, E.W.; Bányai, K.; Albert, M.; Fehér, E.; Mató, T.; Tatár-Kis, T.; Palya, V. Tembusu-like flavivirus (Perak virus) as the cause of neurological disease outbreaks in young Pekin ducks. Avian Pathol. 2014, 43, 552–560. [Google Scholar] [CrossRef]
- Thontiravong, A.; Ninvilai, P.; Tunterak, W.; Nonthabenjawan, N.; Chaiyavong, S.; Angkabkingkaew, K.; Mungkundar, C.; Phuengpho, W.; Oraveerakul, K.; Amonsin, A. Tembusu-Related Flavivirus in Ducks, Thailand. Emerg. Infect. Dis. 2015, 21, 2164–2167. [Google Scholar] [CrossRef] [Green Version]
- Yan, P.; Zhao, Y.; Zhang, X.; Xu, D.; Dai, X.; Teng, Q.; Yan, L.; Zhou, J.; Ji, X.; Zhang, S.; et al. An infectious disease of ducks caused by a newly emerged Tembusu virus strain in mainland China. Virology 2011, 417, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Zhang, C.; Liu, Y.; Ye, W.; Han, J.; Ma, G.; Zhang, D.; Xu, F.; Gao, X.; Tang, Y.; et al. Tembusu virus in ducks, China. Emerg. Infect. Dis. 2011, 17, 1873–1875. [Google Scholar] [CrossRef]
- Liu, P.; Lu, H.; Li, S.; Moureau, G.; Deng, Y.-Q.; Wang, Y.; Zhang, L.; Jiang, T.; De Lamballerie, X.; Qin, C.-F.; et al. Genomic and antigenic characterization of the newly emerging Chinese duck egg-drop syndrome flavivirus: Genomic comparison with Tembusu and Sitiawan viruses. J. Gen. Virol. 2012, 93, 2158–2170. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Han, K.; Zhao, D.; Liu, Y.; Zhang, J.; Niu, H.; Zhang, K.; Zhu, J.; Wu, D.; Gao, L.; et al. Identification and molecular characterization of a novel flavivirus isolated from geese in China. Res. Vet. Sci. 2013, 94, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Diao, Y.; Yu, C.; Gao, X.; Ju, X.; Xue, C.; Liu, X.; Ge, P.; Qu, J.; Zhang, D. Characterization of a Tembusu virus isolated from naturally infected house sparrows (Passer domesticus) in Northern China. Transbound. Emerg. Dis. 2013, 60, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Chen, X.; Liu, W.; Fu, S.; Li, F.; Liang, G.; Yang, G.; Zheng, H.; Li, J.; Yin, Z.; et al. Emergence of Japanese encephalitis among adults 40 years of age or older in northern China: Epidemiological and clinical characteristics. Transbound. Emerg. Dis. 2021, 68, 3415–3423. [Google Scholar] [CrossRef]
- Gao, X.; Nasci, R.; Liang, G. The neglected arboviral infections in mainland China. PLoS Negl. Trop. Dis. 2010, 4, e624. [Google Scholar] [CrossRef]
- Yurayart, N.; Ninvilai, P.; Chareonviriyaphap, T.; Kaewamatawong, T.; Thontiravong, A.; Tiawsirisup, S. Pathogenesis of Thai duck Tembusu virus in BALB/c mice: Descending infection and neuroinvasive virulence. Transbound. Emerg. Dis. 2021, 68, 3529–3540. [Google Scholar] [CrossRef] [PubMed]
- Pulmanausahakul, R.; Ketsuwan, K.; Jaimipuk, T.; Smith, D.R.; Auewarakul, P.; Songserm, T. Detection of antibodies to duck tembusu virus in human population with or without the history of contact with ducks. Transbound. Emerg. Dis. 2022, 69, 870–873. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Gao, X.; Diao, Y.; Feng, Q.; Chen, H.; Liu, X.; Ge, P.; Yu, C. Tembusu virus in human, China. Transbound. Emerg. Dis. 2013, 60, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Lu, H.; Li, S.; Wu, Y.; Gao, G.F.; Su, J. Duck egg drop syndrome virus: An emerging Tembusu-related flavivirus in China. Sci. China Life Sci. 2013, 56, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Sheng, Z.Z.; Huang, B.; Ma, X.; Li, Y.; Yuan, X.; Qin, Z.; Wang, D.; Chakravarty, S.; Li, F.; et al. Structural, antigenic, and evolutionary characterizations of the envelope protein of newly emerging Duck Tembusu Virus. PLoS ONE 2013, 8, 71319. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Pan, Y.; Guo, J.; Wang, D.; Tong, X.; Wang, Y.; Li, J.; Zhao, J.; Ji, Y.; Wu, Z.; et al. The Evolution, Genomic Epidemiology, and Transmission Dynamics of Tembusu Virus. Viruses 2022, 14, 1236. [Google Scholar] [CrossRef]
- Qiu, G.; Cui, Y.; Li, Y.; Li, Y.; Wang, Y. The spread of Tembusu virus in China from 2010 to 2019. Virus Res. 2021, 300, 198374. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Li, X.; Wang, Z.; Liu, X.; Dong, X.; Fu, R.; Su, X.; Xu, B.; Teng, Q.; Yuan, C.; et al. The emergence of a disease caused by a mosquito origin Cluster 3.2 Tembusu virus in chickens in China. Vet. Microbiol. 2022, 272, 109500. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, H.; Sun, X.; Fu, S.; Wang, H.; Feng, Y.; Wang, H.; Tang, Q.; Liang, G.D. Distribution of mosquitoes and mosquito-borne arboviruses in Yunnan province near the China-Myanmar-Laos border. Am. J. Trop. Med. Hyg. 2011, 84, 738–746. [Google Scholar] [CrossRef]
- Zhang, W.; Li, F.; Liu, A.; Lin, X.; Fu, S.; Song, J.; Liu, G.; Shao, N.; Tao, Z.; Wang, Q.; et al. Identification and genetic analysis of Kadipiro virus isolated in Shandong province, China. Virol. J. 2018, 15, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Xu, H.; Song, S.; Cheng, R.; Fan, N.; Fu, S.; Zhang, S.; Xu, Z.; He, Y.; Lei, W.; et al. Emergence of Zika Virus in Culex tritaeniorhynchus and Anopheles sinensis Mosquitoes in China. Virol. Sin. 2021, 36, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Kuno, G. Universal diagnostic RT-PCR protocol for arboviruses. J. Virol. Methods 1998, 72, 27–41. [Google Scholar] [CrossRef]
- Lei, W.; Guo, X.; Fu, S.; Feng, Y.; Tao, X.; Song, J.; Yang, Z.; Zhou, H.; Liang, G. The genetic characteristics and evolution of Tembusu virus. Vet. Microbiol. 2017, 201, 32–41. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Burland, T.G. DNASTAR’s Lasergene sequence analysis software. Methods Mol. Biol. 2000, 132, 71–91. [Google Scholar]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.H.; Su, C.L.; Chang, M.C.; Hu, H.-C.; Yang, S.-L.; Shu, P.-Y. Genome Analysis of a Novel Tembusu Virus in Taiwan. Viruses 2020, 12, 567. [Google Scholar] [CrossRef]
- Tang, Y.; Diao, Y.; Chen, H.; Ou, Q.; Liu, X.; Gao, X.; Yu, C.; Wang, L. Isolation and genetic characterization of a tembusu virus strain isolated from mosquitoes in Shandong, China. Transbound. Emerg. Dis. 2015, 62, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Yurayart, N.; Ninvilai, P.; Chareonviriyaphap, T.; Kaewamatawong, T.; Thontiravong, A.; Tiawsirisup, S. Interactions of duck Tembusu virus with Aedes aegypti and Aedes albopictus mosquitoes: Vector competence and viral mutation. Acta Trop. 2021, 222, 106051. [Google Scholar] [CrossRef]
- Wang, C.; Yan, H. Diagnosis and control of a case of tembusovirus disease in Cherry Valley breeder ducks. Yunnan Anim. Husb. Vet. Med. 2020, 1, 18–20. [Google Scholar]
- Gutiérrez-Bugallo, G.; Piedra, L.A.; Rodriguez, M.; Bisset, J.A.; Lourenço-De-Oliveira, R.; Weaver, S.C.; Vasilakis, N.; Vega-Rúa, A. Vector-borne transmission and evolution of Zika virus. Nat. Ecol. Evol. 2019, 3, 561–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukhralia, S.; Verma, M.; Gopirajan, S.; Dhanaraj, P.S.; Lal, R.; Mehla, N.; Kant, C.R. From dengue to Zika: The wide spread of mosquito-borne arboviruses. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 3–14. [Google Scholar] [CrossRef]
- Huang, Y.S.; Higgs, S.; Vanlandingham, D.L.; Colpitts, T.M. Arbovirus-Mosquito Vector-Host Interactions and the Impact on Transmission and Disease Pathogenesis of Arboviruses. Front. Microbiol. 2019, 10, 22. [Google Scholar] [CrossRef]
- Hamel, R.; Phanitchat, T.; Wichit, S.; Vargas, R.E.M.; Jaroenpool, J.; Diagne, C.T.; Pompon, J.; Missé, D. New Insights into the Biology of the Emerging Tembusu Virus. Pathogens 2021, 10, 1010. [Google Scholar] [CrossRef]
- Campos, S.S.; Fernandes, R.S.; Dos Santos, A.A.C.; De Miranda, R.M.; Telleria, E.L.; Ferreira-De-Brito, A.; De Castro, M.G.; Failloux, A.-B.; Bonaldo, M.C.; Lourenço-De-Oliveira, R. Zika virus can be venereally transmitted between Aedes aegypti mosquitoes. Parasites Vectors 2017, 10, 605. [Google Scholar] [CrossRef] [Green Version]
- Thangamani, S.; Huang, J.; Hart, C.E.; Guzman, H.; Tesh, R.B. Vertical Transmission of Zika Virus in Aedes aegypti Mosquitoes. Am. J. Trop. Med. Hyg. 2016, 95, 1169–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira-de-Lima, V.H.; Lima-Camara, T.N. Natural vertical transmission of dengue virus in Aedes aegypti and Aedes albopictus: A systematic review. Parasites Vectors 2018, 11, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanisuriwong, J.; Yurayart, N.; Thontiravong, A.; Tiawsirisup, S. Vector competence of Culex tritaeniorhynchus and Culex quinquefasciatus (Diptera: Culicidae) for duck Tembusu virus transmission. Acta Trop. 2021, 214, 105785. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Cheng, G. Adaptive Evolution as a Driving Force of the Emergence and Re-Emergence of Mosquito-Borne Viral Diseases. Viruses 2022, 14, 435. [Google Scholar] [CrossRef]
- Fontes-Garfias, C.R.; Shan, C.; Luo, H.; Muruato, A.E.; Medeiros, D.B.; Mays, E.; Xie, X.; Zou, J.; Roundy, C.M.; Wakamiya, M.; et al. Functional Analysis of Glycosylation of Zika Virus Envelope Protein. Cell Rep. 2017, 21, 1180–1190. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Liang, T.; Lv, J.; Qu, S.; Meng, R.; Yang, B.; Feng, C.; Dai, W.; Wang, X.; Zhang, B.; et al. Substantial Attenuation of Virulence of Tembusu Virus Strain PS Is Determined by an Arginine at Residue 304 of the Envelope Protein. J. Virol. 2021, 95, e02331-20. [Google Scholar] [CrossRef]
- Sun, M.; Zhang, L.; Cao, Y.; Wang, J.; Yu, Z.; Sun, X.; Liu, F.; Li, Z.; Liu, P.; Su, J. Basic Amino Acid Substitution at Residue 367 of the Envelope Protein of Tembusu Virus Plays a Critical Role in Pathogenesis. J. Virol. 2020, 94, e02011-19. [Google Scholar] [CrossRef]
- Sun, X.; Sun, M.; Zhang, L.; Yu, Z.; Li, J.; Xie, W.; Su, J. Amino Acid Substitutions in NS5 Contribute Differentially to Tembusu Virus Attenuation in Ducklings and Cell Cultures. Viruses 2021, 13, 921. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genomic Region | Cluster 3 | Cluster 1 | Cluster 2 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| YN12193 | SD14 | YN12115 | MM1775 | Sitiawan | TP1906 | NTUC225/20 | \ | \ | |
| ORF | 97.7% | 94.0% | 93.4% | 89.7% | 88.3% | 87.6% | 87.4% | 89.8–90.0% | 89.0–89.9% |
| (99.4%) | (97.7%) | (98.3%) | (97.6%) | (97.3%) | (97.3%) | (97.2%) | (96.5–96.8%) | (95.9–97.6%) | |
| C | 98.6% | 93.9% | 96.1% | 93.6% | 91.7% | 91.7% | 91.4% | 93.1–93.3% | 90.6–93.1% |
| (99.2%) | (95.8%) | (97.5%) | (96.7%) | (95.8%) | (95.8%) | (95.8%) | (95.0–95.8%) | (91.7–95.0%) | |
| prM | 97.6% | 94.0% | 93.2% | 91.6% | 86.4% | 87.2% | 87.0% | 89.2–90.6% | 88.0–89.8% |
| (100%) | (96.4%) | (98.8%) | (100%) | (97.0%) | (98.8%) | (98.2%) | (97.6–97.6%) | (95.8–98.2%) | |
| E | 97.9% | 93.4% | 94.0% | 89.2% | 87.3% | 86.4% | 86.1% | 89.1–89.7% | 87.6–89.6% |
| (99.6%) | (95.2%) | (97.4%) | (97.2%) | (97.0%) | (97.0%) | (96.8%) | (95.6–96.2%) | (94.6–96.8%) | |
| NS1 | 97.1% | 94.2% | 92.5% | 89.4% | 87.9% | 88.2% | 87.8% | 89.2–89.7% | 86.7–88.8% |
| (98.0%) | (98.0%) | (96.9%) | (96.0%) | (94.9%) | (95.7%) | (95.5%) | (94.6–95.5%) | (88.9–94.6%) | |
| NS2A | 98.1% | 92.8% | 91.6% | 88.5% | 89.3% | 86.6% | 86.6% | 87.8–88.0% | 86.6–88.7% |
| (99.1%) | (96.0%) | (96.0%) | (94.7%) | (94.3%) | (94.3%) | (94.3%) | (92.1–93.8%) | (91.6–93.8%) | |
| NS2B | 97.5% | 94.7% | 93.4% | 89.3% | 89.3% | 86.3% | 87.5% | 90.3–90.6% | 88.5–91.3% |
| (99.2%) | (99.2%) | (100%) | (96.9%) | (96.9%) | (93.9%) | (96.2%) | (98.5–99.2%) | (95.4–98.5%) | |
| NS3 | 97.3% | 94.3% | 93.4% | 90.6% | 89.4% | 89.4% | 89.3% | 90.5–90.9% | 89.8–91.0% |
| (99.5%) | (99.4%) | (99.2%) | (99.4%) | (98.9%) | (98.7%) | (98.7%) | (98.4–98.5%) | (97.9–98.9%) | |
| NS4A | 98.1% | 92.6% | 92.9% | 88.9% | 87.0% | 87.0% | 86.2% | 88.4–88.9% | 84.9–88.1% |
| (99.2%) | (96.8%) | (97.6%) | (99.2%) | (97.6%) | (97.6%) | (96.8%) | (95.2–96.0%) | (94.4–97.6%) | |
| 2K | 97.1% | 92.8% | 94.2% | 85.5% | 85.5% | 87.0% | 87.0% | 84.1–88.4% | 88.4–94.2% |
| (100%) | (95.7%) | (100%) | (95.7%) | (95.7%) | (95.7%) | (95.7%) | (100–100%) | (100–100%) | |
| NS4B | 97.4% | 93.4% | 92.4% | 89.5% | 87.1% | 86.0% | 85.7% | 88.5–89.8% | 85.6–89.9% |
| (99.2%) | (96.9%) | (98.0%) | (97.6%) | (98.0%) | (97.6%) | (97.2%) | (94.9–96.1%) | (91.3–98.0%) | |
| NS5 | 98.0% | 94.5% | 93.9% | 89.6% | 88.6% | 87.5% | 87.2% | 90.1–90.5% | 89.5–90.9% |
| (99.9%) | (99.0%) | (99.2%) | (97.8%) | (98.1%) | (98.1%) | (98.0%) | (97.2–97.9%) | (97.5–98.5%) | |
| GENOMIC REGION | SITES | CLUSTER 1 | CLUSTER 2 | CLUSTER 3 2 |
|---|---|---|---|---|
| C | 34 | M (100%) 1 | T (100%) | T (100%) |
| 40 | K (100%) | R (97.81%); T (2.19%) | R (100%) | |
| 61 | K (100%) | R (100%) | K (100%) | |
| 76 | N (100%) | N (98.54%); D (1.46%) | S (100%) | |
| E | 2 | S (100%) | S (100%) | N (100%) |
| 52 | D (100%) | E (100%) | E (100%) | |
| 72 | S (100%) | S (100%) | P (100%) | |
| 83 | S (100%) | P (100%) | P (100%) | |
| 89 | D (100%) | E (98.54%); G (0.73%); K (0.73%) | D (87.50%) *; N (12.50%) | |
| 157 | A (100%) | A (97.81%); V (2.19%) | V (100%) | |
| 180 | L (100%) | M (100%) | L (100%) | |
| 185 | S (100%) | T (98.54%); A (1.46%) | A (62.50%) *; S (25.00%); T (12.50%) | |
| 236 | K (100%) | K (100%) | R (100%) | |
| 312 | A (100%) | V (95.62%); A (4.38%) | A (100%) | |
| 332 | S (100%) | T (97.81%); A (1.46%); I (0.73%) | S (100%) | |
| 358 | V (100%) | V (100%) | V (87.50%%); I (12.50%) ** | |
| M | 22 | L (100%) | L (99.27%); N (0.73%) | M (87.50%) *; T (12.50%) |
| 24 | I (100%) | V (100%) | I (87.50%) *; V (12.50%) | |
| 29 | T (100%) | A (100%) | T (100%) | |
| 60 | H (100%) | Y (97.08%); H (2.92%) | Y (100%) | |
| 115 | I (100%) | T (100%) | T (100%) | |
| 150 | S (100%) | S (99.27%); G (0.73%) | G (87.50%) *; S (12.50%) | |
| NS1 | 2 | V (100%) | T (100%) | M (100%) |
| 21 | V (100%) | V (100%) | I (87.50%) *; V (12.50%) | |
| 41 | R (100%) | R (99.27%); K (0.73%) | K (100%) | |
| 51 | K (100%) | E (89.78%); V (10.22%) | E (100%) | |
| 83 | G (100%) | A (100%) | G (50.00%) *; S (50.00%) | |
| 99 | R (100%) | K (99.27%); R (0.73%) | R (100%) | |
| 108 | E (100%) | D (98.54%); G (1.46%) | E (100%) | |
| 113 | F (100%) | Y (100%) | F (75.00%); I (12.50%); L (12.50%) ** | |
| 139 | K (100%) | K (100%) | R (100%) | |
| 274 | K (100%) | V (98.54%); A (1.46%) | K (100%) | |
| 351 | M (100%) | M (100%) | V (100%) | |
| NS2A | 31 | P (100%) | P (100%) | S (100%) |
| 35 | S (100%) | S (100%) | P (100%) | |
| 53 | E (100%) | D (100%) | D (100%) | |
| 68 | R (100%) | S (98.54%); N (1.46%) | S (100%) | |
| 122 | D (100%) | N (100%) | N (100%) | |
| 123 | I (100%) | M (97.81%); I (2.19%) | M (100%) | |
| 150 | L (100%) | F (100%) | L (100%) | |
| 157 | S (100%) | L (98.54%); V (1.46%) | S (100%) | |
| 187 | S (100%) | S (100%) | N (100%) | |
| NS2B | 95 | L (100%) | L (100%) | F (100%) |
| NS3 | 15 | R (100%) | R (100%) | K (100%) |
| 258 | I (100%) | I (100%) | V (100%) | |
| 324 | E (100%) | E (99.27%); D (0.73%) | D (100%) | |
| 591 | I (100%) | I (98.54%); T (0.73%); V (0.73%) | T (100%) | |
| NS4A | 70 | L (100%) | F (99.27%); L (0.73%) | F (100%) |
| 89 | A (100%) | T (100%) | A (87.50%); V (12.50%) ** | |
| 96 | M (100%) | I (100%) | V (100%) | |
| 110 | I (66.67%); F (33.33%) | V (99.27%); I (0.73%) | I (100%) | |
| NS4B | 14 | A (100%) | S (100%) | S (100%) |
| 22 | D (100%) | D (89.78%); N (1.46%); E (8.76%) | D (50.00%); N (25.00%); C (12.50%); S (12.50%) **; | |
| 60 | I (100%) | I (100%) | V (100%) | |
| 118 | V (100%) | I (100%) | I (100%) | |
| 183 | M (100%) | V (99.27%); M (0.73%) | V (75.00%) *; A (25.00%) | |
| NS5 | 133 | Y (100%) | H (98.54%); Q (1.46%) | H (100%) |
| 150 | A (100%) | A (100%) | S (100%) | |
| 188 | M (100%) | M (100%) | T (100%) | |
| 232 | G (100%) | S (100%) | S (87.50%) *; T (12.50%) | |
| 246 | R (100%) | K (99.27%); R (0.73%) | R (100%) | |
| 288 | K (100%) | R (100%) | K (100%) | |
| 388 | S (100%) | G (99.27%); S (0.73%) | G (100%) | |
| 390 | D (100%) | N (100%) | N (100%) | |
| 448 | R (100%) | K (100%) | K (100%) | |
| 638 | E (100%) | E (100%) | E (87.50%); G (12.50%) ** | |
| 790 | I (100%) | V (100%) | V (100%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, D.; Wu, C.; Wang, R.; Yao, X.; Nie, K.; Lv, Q.; Fu, S.; Yin, Q.; Su, W.; Li, F.; et al. Persistence of Tembusu Virus in Culex tritaeniorhynchus in Yunnan Province, China. Pathogens 2023, 12, 490. https://doi.org/10.3390/pathogens12030490
Hu D, Wu C, Wang R, Yao X, Nie K, Lv Q, Fu S, Yin Q, Su W, Li F, et al. Persistence of Tembusu Virus in Culex tritaeniorhynchus in Yunnan Province, China. Pathogens. 2023; 12(3):490. https://doi.org/10.3390/pathogens12030490
Chicago/Turabian StyleHu, Danhe, Chao Wu, Ruichen Wang, Xiaohui Yao, Kai Nie, Quan Lv, Shihong Fu, Qikai Yin, Wenzhe Su, Fan Li, and et al. 2023. "Persistence of Tembusu Virus in Culex tritaeniorhynchus in Yunnan Province, China" Pathogens 12, no. 3: 490. https://doi.org/10.3390/pathogens12030490