
Epidemiological Survey and Risk Factor Analysis of 14 Potential Pathogens in Golden Snub-Nosed Monkeys at Shennongjia National Nature Reserve, China
 , , ,
, , ,  , , ,
, , ,
Abstract
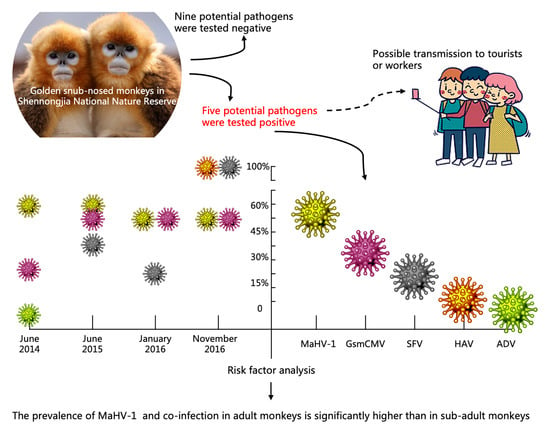
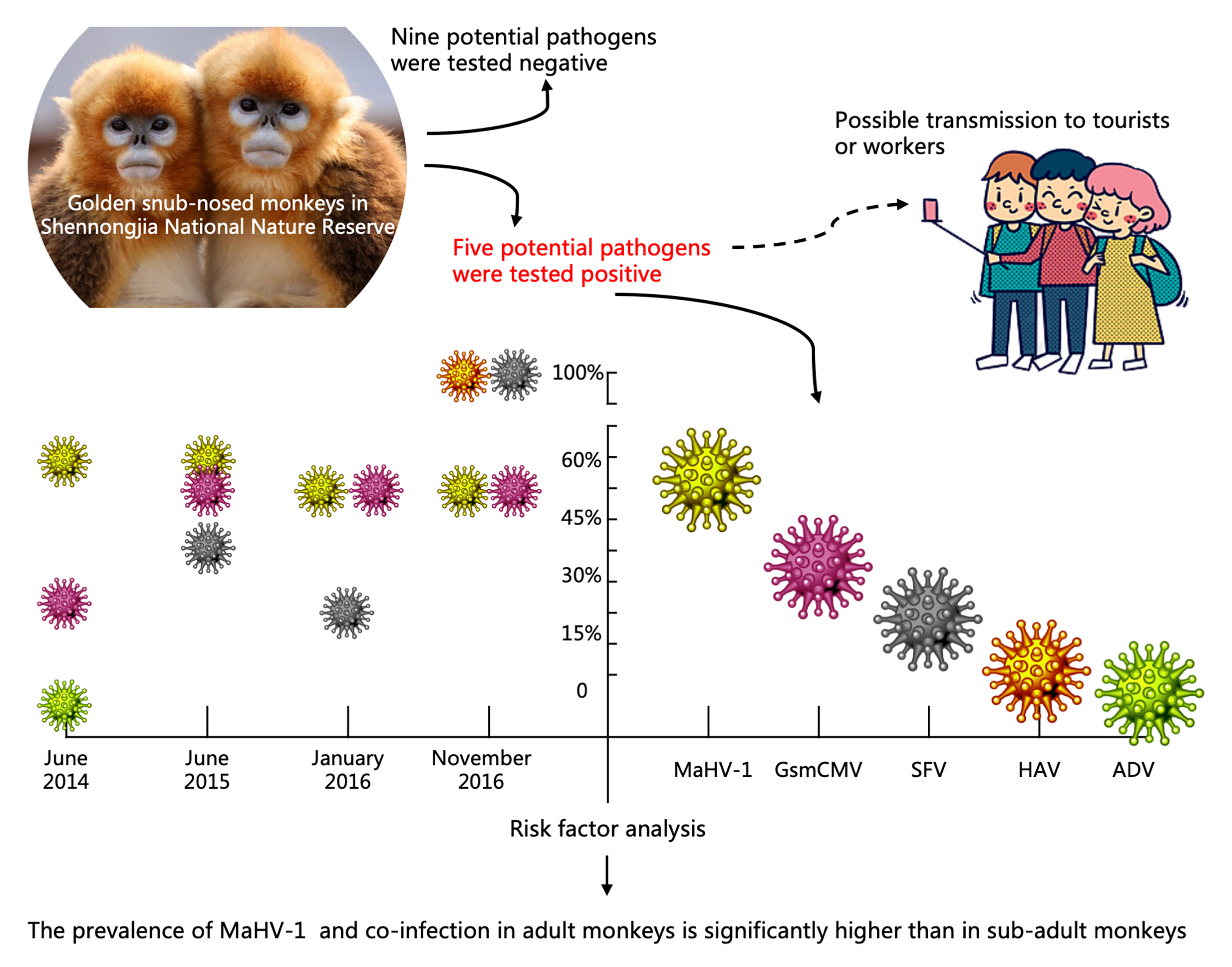
:
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
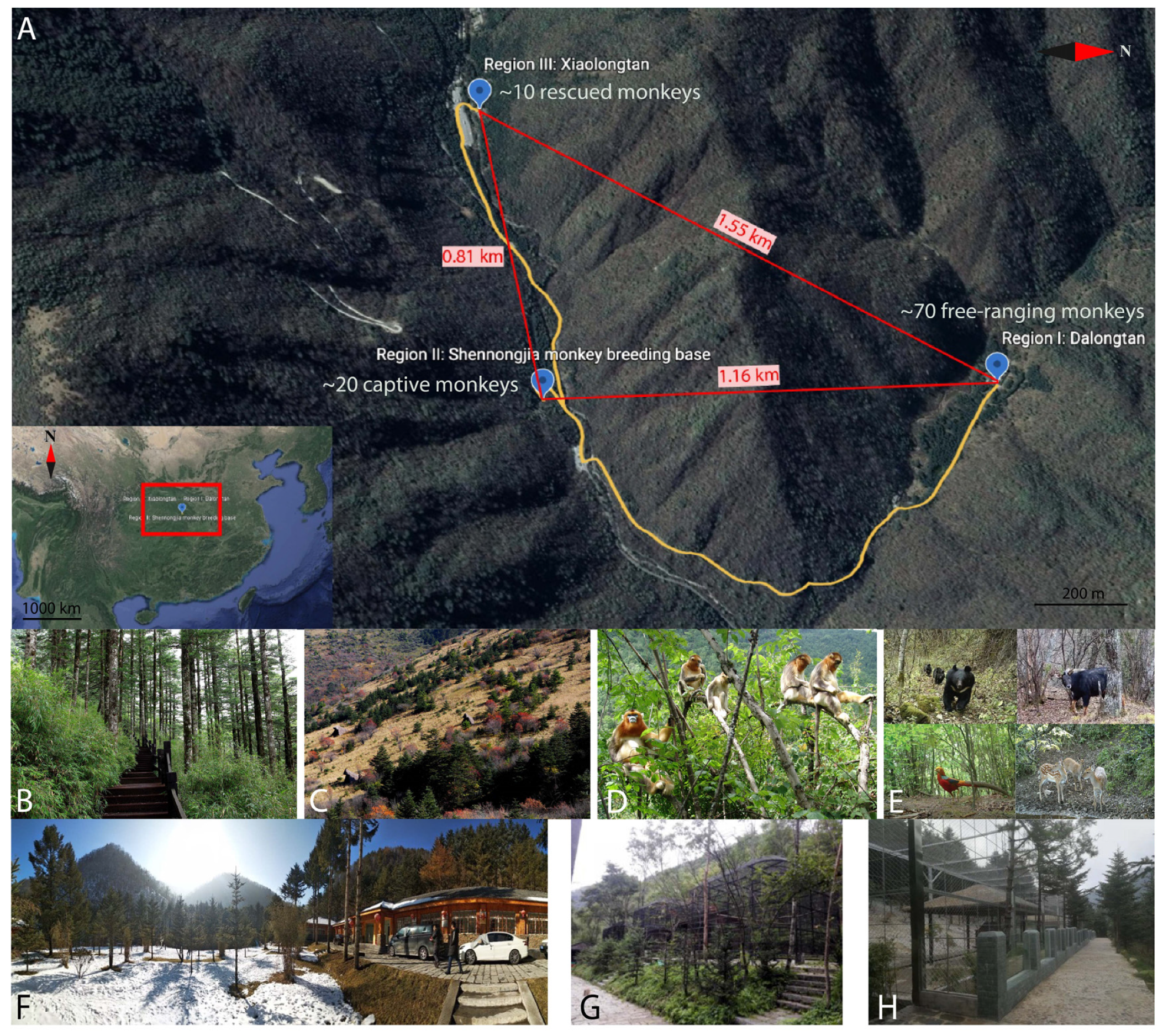
2.2. Study Area and Monkey Information
2.3. Sample Collection
2.4. Interferon-γ (IFN-γ) In Vitro Release Assay to Test TB
2.5. Detection of Serum Antibodies to 11 Viruses
2.6. PCR and Phylogenetic Analysis of Adenovirus and Rotavirus
2.7. Statistical Analysis
3. Results
3.1. Sample Information
3.2. Prevalence of the Investigated Pathogens at Sampling Periods
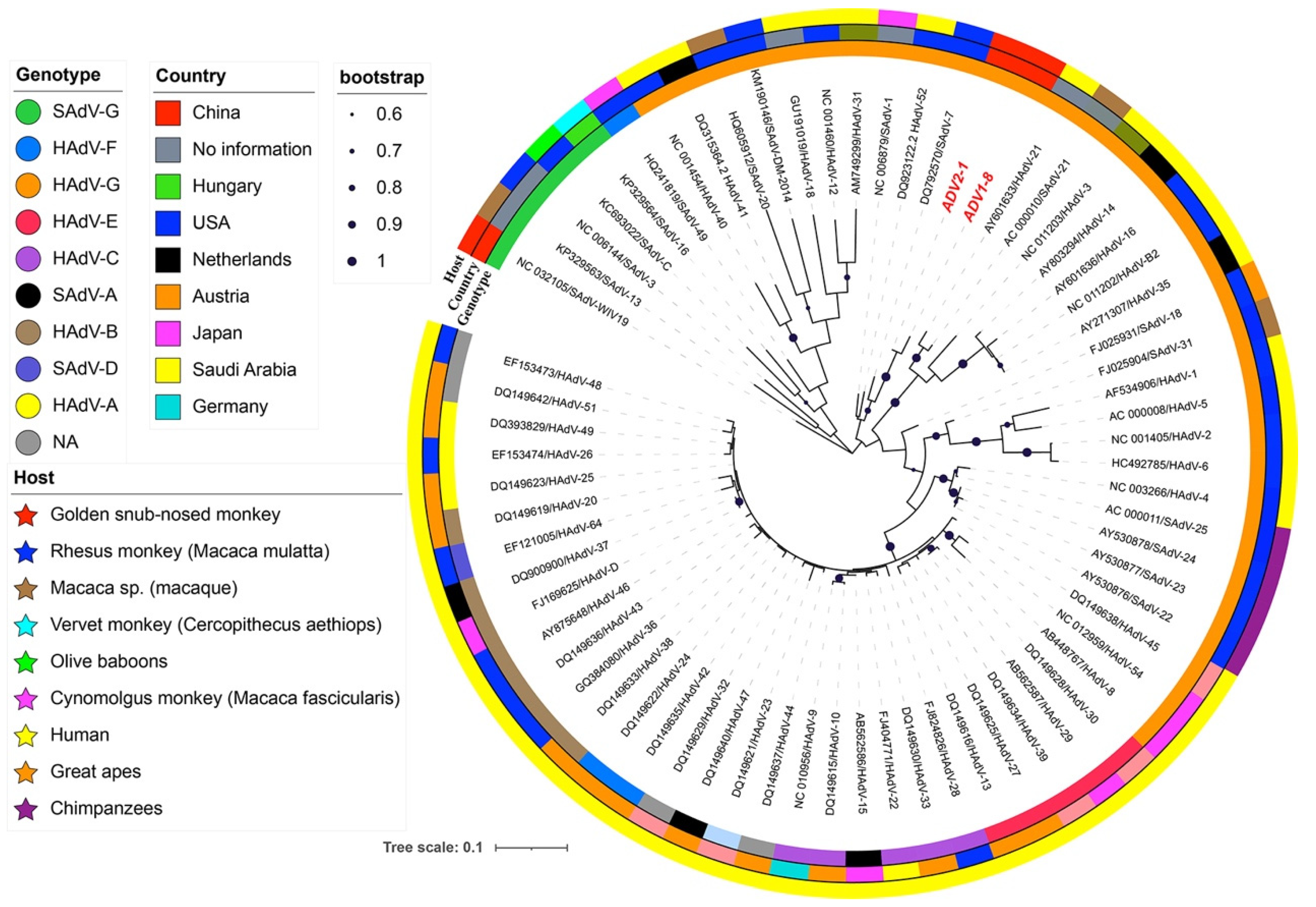
3.3. Genetic and Phylogenetic Analysis of Adenovirus
3.4. Risk Factor Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiang, Z.; Yu, Y.; Yang, M.; Yang, J.; Niao, M.; Li, M. Does flagship species tourism benefit conservation? A case study of the golden snub-nosed monkey in Shennongjia National Nature Reserve. Chin. Sci. Bull. 2011, 56, 2553–2558. [Google Scholar] [CrossRef] [Green Version]
- Long, Y.R.; Richardson, M. Rhinopithecus roxellana. The IUCN Red List of Threatened Species 2020. Available online: https://www.iucnredlist.org/species/19596/8985735 (accessed on 12 March 2023).
- Zhou, X.; Meng, X.; Liu, Z.; Chang, J.; Wang, B.; Li, M.; Wengel, P.O.; Tian, S.; Wen, C.; Wang, Z.; et al. Population Genomics Reveals Low Genetic Diversity and Adaptation to Hypoxia in Snub-Nosed Monkeys. Mol. Biol. Evol. 2016, 33, 2670–2681. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, Z.; Pan, H.; Qi, J.; Li, D.; Zhang, L.; Shen, Y.; Xiang, Z.; Li, M. Captivity Influences the Gut Microbiome of Rhinopithecus roxellana. Front. Microbiol. 2021, 12, 763022. [Google Scholar] [CrossRef]
- Luo, M.; Liu, Z.; Pan, H.; Zhao, L.; Li, M. Historical geographic dispersal of the golden snub-nosed monkey (Rhinopithecus roxellana) and the influence of climatic oscillations. Am. J. Primatol. 2012, 74, 91–101. [Google Scholar] [CrossRef]
- Smith, K.F.; Acevedo-Whitehouse, K.; Pedersen, A.B. The role of infectious diseases in biological conservation. Anim. Conserv. 2009, 12, 1–12. [Google Scholar] [CrossRef]
- Warit, S.; Billamas, P.; Makhao, N.; Jaitrong, S.; Juthayothin, T.; Yindeeyoungyeon, W.; Dokladda, K.; Smittipat, N.; Kemthong, T.; Meesawat, S.; et al. Detection of tuberculosis in cynomolgus macaques (Macaca fascicularis) using a supplementary Monkey Interferon Gamma Releasing Assay (mIGRA). Sci. Rep. 2020, 10, 16759. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, T.L.; Gillespie, T.R.; Rwego, I.B.; Estoff, E.L.; Chapman, C.A. Forest fragmentation as cause of bacterial transmission among nonhuman primates, humans, and livestock, Uganda. Emerg. Infect. Dis. 2008, 14, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Devaux, C.A.; Mediannikov, O.; Medkour, H.; Raoult, D. Infectious Disease Risk Across the Growing Human-Non Human Primate Interface: A Review of the Evidence. Front. Public Health 2019, 7, 305. [Google Scholar] [CrossRef] [Green Version]
- Michel, A.L.; Huchzermeyer, H.F. The zoonotic importance of Mycobacterium tuberculosis: Transmission from human to monkey. J. S. Afr. Vet. Assoc. 1998, 69, 64–65. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, W.L. CHAPTER 5—Common Viral Infections of Laboratory Primates. In The Laboratory Primate; Sonia, W.-C., Ed.; Academic Press: London, UK, 2005; pp. 75–89. [Google Scholar]
- Carbone, M.; Gazdar, A.; Butel, J.S. SV40 and human mesothelioma. Transl. Lung Cancer Res. 2020, 9, S47–S59. [Google Scholar] [CrossRef]
- Elmore, D.; Eberle, R. Monkey B virus (Cercopithecine herpesvirus 1). Comp. Med. 2008, 58, 11–21. [Google Scholar] [PubMed]
- Calattini, S.; Betsem, E.B.A.; Froment, A.; Mauclère, P.; Tortevoye, P.; Schmitt, C.; Njouom, R.; Saib, A.; Gessain, A. Simian foamy virus transmission from apes to humans, rural Cameroon. Emerg. Infect. Dis. 2007, 13, 1314–1320. [Google Scholar] [CrossRef]
- Khan, A.S.; Bodem, J.; Buseyne, F.; Gessain, A.; Johnson, W.; Kuhn, J.H.; Kuzmak, J.; Lindemann, D.; Linial, M.L.; Lochelt, M.; et al. Spumaretroviruses: Updated taxonomy and nomenclature. Virology 2018, 516, 158–164. [Google Scholar] [CrossRef]
- Buseyne, F.; Betsem, E.; Montange, T.; Njouom, R.; Bilounga Ndongo, C.; Hermine, O.; Gessain, A. Clinical Signs and Blood Test Results among Humans Infected with Zoonotic Simian Foamy Virus: A Case-Control Study. J. Infect. Dis. 2018, 218, 144–151. [Google Scholar] [CrossRef]
- Gessain, A.; Montange, T.; Betsem, E.; Bilounga Ndongo, C.; Njouom, R.; Buseyne, F. Case-Control Study of the Immune Status of Humans Infected with Zoonotic Gorilla Simian Foamy Viruses. J. Infect. Dis. 2020, 221, 1724–1733. [Google Scholar] [CrossRef]
- Pinto-Santini, D.M.; Stenbak, C.R.; Linial, M.L. Foamy virus zoonotic infections. Retrovirology 2017, 14, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posovszky, C.; Buderus, S.; Classen, M.; Lawrenz, B.; Keller, K.M.; Koletzko, S. Acute Infectious Gastroenteritis in Infancy and Childhood. Dtsch. Arztebl. Int. 2020, 117, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Caddy, S.; Papa, G.; Borodavka, A.; Desselberger, U. Rotavirus research: 2014–2020. Virus Res. 2021, 304, 198499. [Google Scholar] [CrossRef] [PubMed]
- Mo, J. Historical Investigation of Fowl Adenovirus Outbreaks in South Korea from 2007 to 2021: A Comprehensive Review. Viruses 2021, 13, 2256. [Google Scholar] [CrossRef]
- Assaf, B.T.; Knight, H.L.; Miller, A.D. rhesus cytomegalovirus (macacine herpesvirus 3)-associated facial neuritis in simian immunodeficiency virus-infected rhesus macaques (Macaca mulatta). Vet. Pathol. 2015, 52, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Yang, Y.; Tao, L.; Jianhua, H.U.; Gao, C. Serological survey of simian immunodeficiency virus and simian type D retrovirus infection in Shanghai area. Anim. Husb. Vet. Med. 2018, 52, 91–92. (In Chinese) [Google Scholar]
- Mahieux, R.; Gessain, A. HTLV-3/STLV-3 and HTLV-4 viruses: Discovery, epidemiology, serology and molecular aspects. Viruses 2011, 3, 1074–1090. [Google Scholar] [CrossRef] [PubMed]
- Lerche, N.W.; Switzer, W.M.; Yee, J.L.; Shanmugam, V.; Rosenthal, A.N.; Chapman, L.E.; Folks, T.M.; Heneine, W. Evidence of infection with simian type D retrovirus in persons occupationally exposed to nonhuman primates. J. Virol. 2001, 75, 1783–1789. [Google Scholar] [CrossRef] [Green Version]
- Gray, W.L. Simian Varicella Virus: Molecular Virology. In Varicella-Zoster Virus; Abendroth, A., Arvin, A.M., Moffat, J.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 291–308. [Google Scholar]
- Landolfo, S.; Gariglio, M.; Gribaudo, G.; Lembo, D. The human cytomegalovirus. Pharmacol. Ther. 2003, 98, 269–297. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Bailes, E.; Robertson, D.L.; Chen, Y.; Rodenburg, C.M.; Michael, S.F.; Cummins, L.B.; Arthur, L.O.; Peeters, M.; Shaw, G.M.; et al. Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature 1999, 397, 436–441. [Google Scholar] [CrossRef]
- Asikainen, K.; Vesanen, M.; Kuittinen, T.; Vaheri, A. Identification of human type D retrovirus as a contaminant in a neuroblastoma cell line. Arch. Virol. 1993, 129, 357–361. [Google Scholar] [CrossRef]
- Cai, Y.; Yu, H.; Liu, H.; Jiang, C.; Sun, L.; Niu, L.; Liu, X.; Li, D.; Li, J. Genome-wide screening of microsatellites in golden snub-nosed monkey (Rhinopithecus roxellana), for the development of a standardized genetic marker system. Sci. Rep. 2020, 10, 10614. [Google Scholar] [CrossRef]
- Wang, L.; Wu, J.; Liu, X.; Di, D.; Liang, Y.; Feng, Y.; Zhang, S.; Li, B.; Qi, X.-G. A high-quality genome assembly for the endangered golden snub-nosed monkey (Rhinopithecus roxellana). GigaScience 2019, 8, giz098. [Google Scholar] [CrossRef] [Green Version]
- Kuang, W.-M.; Ming, C.; Li, H.-P.; Wu, H.; Frantz, L.; Roos, C.; Zhang, Y.-P.; Zhang, C.-L.; Jia, T.; Yang, J.-Y.; et al. The Origin and Population History of the Endangered Golden Snub-Nosed Monkey (Rhinopithecus roxellana). Mol. Biol. Evol. 2019, 36, 487–499. [Google Scholar] [CrossRef]
- Xiang, Z.; Fan, P.; Chen, H.; Liu, R.; Zhang, B.; Yang, W.; Yao, H.; Grueter, C.C.; Garber, P.A.; Li, M. Routine allomaternal nursing in a free-ranging Old World monkey. Sci. Adv. 2019, 5, eaav0499. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.-M.-R.; Sha, J.C.M.; Kawazoe, T.; Dong, X. Sleeping site and tree selection by Sichuan snub-nosed monkeys (Rhinopithecus roxellana) in Baihe Nature Reserve, Sichuan, China. Am. J. Primatol. 2018, 80, e22936. [Google Scholar] [CrossRef]
- Qin, L.; Hai, Z.; Huanhong, J.; Shijun, Z.; Genggeng, G.; Wenwang, L.; Yiqing, M. Diagnosis of Streptococcus pneumoniae sepsis in golden monkey. J. Fourth Mil. Med. Univ. 2000, 691. (In Chinese) [Google Scholar]
- Jiabo, D.; Lili, N.; Qiang, W.; Bo, Z.; Xingming, Y.; Weigang, C. Diagnosis and Treatment of Golden Monkey Infected with Staphylococcus aureus. Sichuan J. Zool. 2008, 27, 436–439. (In Chinese) [Google Scholar]
- Wenwang, L. Diagnosis, Prevention and Control of Golden Monkeys Infectious Bacillary Dysentery. Prog. Vet. Med. 2009, 30, 115–116. (In Chinese) [Google Scholar] [CrossRef]
- Qi, M.; Wang, Q.; Tong, S.; Zhao, G.; Hu, C.; Chen, Y.; Li, X.; Yang, W.; Zhao, Y.; Platto, S.; et al. Identification of Atypical Enteropathogenic Escherichia coli O98 from Golden Snub-Nosed Monkeys with Diarrhea in China. Front. Vet. Sci. 2017, 4, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linlin, J.; Tongshan, Z.; Jianghai, L.; Zidong, S. Common Types of Parasitic Diseases of Sichuan Golden Monkey and Control Measures. Graziery Vet. Sci. (Electron. Version) 2021, 140–141. (In Chinese) [Google Scholar]
- He, W.; Lu, H.; Zhao, K.; Song, D.; Gai, X.; Gao, F. Complete genome sequence of a coxsackievirus B3 isolated from a Sichuan snub-nosed monkey. J. Virol. 2012, 86, 13134–13135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.; Wu, L.J.; Yang, X.L.; Li, B.; Zhang, W.; Lei, Y.S.; Li, Y.; Yang, G.X.; Chen, J.; Chen, G.; et al. Isolation and characterization of adenoviruses infecting endangered golden snub-nosed monkeys (Rhinopithecus roxellana). Virol. J. 2016, 13, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, M.A.; Yee, J.; Bouley, D.M.; Moorhead, R.; Lerche, N.W. Diagnosis of tuberculosis in macaques, using whole-blood in vitro interferon-gamma (PRIMAGAM) testing. Comp. Med. 2004, 54, 86–92. [Google Scholar] [PubMed]
- Wang, Q.; Jiang, Y.; Tong, S.; Hao, L.; Hu, C.; Chen, Y.; Li, X.; Ren, X.; Chen, H.; Guo, A. Establishment and preliminary application of an IFN-γ in vitro release assay to detect macaque tuberculosis. Chin. J. Vet. Sci. 2015, 35, 1488–1494. (In Chinese) [Google Scholar] [CrossRef]
- Yan, H.; Nguyen, T.A.; Phan, T.G.; Okitsu, S.; Li, Y.; Ushijima, H. Development of RT-multiplex PCR assay for detection of adenovirus and group A and C rotaviruses in diarrheal fecal specimens from children in China. Kansenshogaku Zasshi 2004, 78, 699–709. [Google Scholar] [CrossRef] [Green Version]
- Khamrin, P.; Okame, M.; Thongprachum, A.; Nantachit, N.; Nishimura, S.; Okitsu, S.; Maneekarn, N.; Ushijima, H. A single-tube multiplex PCR for rapid detection in feces of 10 viruses causing diarrhea. J. Virol. Methods 2011, 173, 390–393. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A language and environment for statistical computing. MSOR Connect. 2013, 1, 201. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: New York, NY, USA, 2018. [Google Scholar]
- Zhao, G.; Qi, M.; Wang, Q.; Hu, C.; Li, X.; Chen, Y.; Yang, J.; Yu, H.; Chen, H.; Guo, A. Gut microbiome variations in Rhinopithecus roxellanae caused by changes in the environment. BMC Genom. 2023, 24, 62. [Google Scholar] [CrossRef]
- Barkati, S.; Taher, H.B.; Beauchamp, E.; Yansouni, C.P.; Ward, B.J.; Libman, M.D. Decision Tool for Herpes B Virus Antiviral Prophylaxis after Macaque-Related Injuries in Research Laboratory Workers. Emerg. Infect. Dis. 2019, 25, e190045. [Google Scholar] [CrossRef] [Green Version]
- Wenling, W.; Wenjie, Q.; Jingyuan, L.; Haijun, D.; Li, Z.; Yang, Z.; Guoxing, W.; Yang, P.; Baoying, H.; Zhaomin, F.; et al. Notes from the Field: First Human Infection Case of Monkey B Virus Identified in China, 2021. China CDC Wkly. 2021, 3, 632–633. [Google Scholar] [CrossRef]
- Hui, L. Preliminary report of B virus in golden monkey. Anim. Husb. Vet. Med. 1993, 222. (In Chinese) [Google Scholar]
- Lee, M.-H.; Rostal, M.K.; Hughes, T.; Sitam, F.; Lee, C.-Y.; Japning, J.; Harden, M.E.; Griffiths, A.; Basir, M.; Wolfe, N.D.; et al. Macacine Herpesvirus 1 in Long-Tailed Macaques, Malaysia, 2009–2011. Emerg. Infect. Dis. 2015, 21, 1107–1113. [Google Scholar] [CrossRef]
- Wisely, S.M.; Sayler, K.A.; Anderson, C.J.; Boyce, C.L.; Klegarth, A.R.; Johnson, S.A. Macacine Herpesvirus 1 Antibody Prevalence and DNA Shedding among Invasive Rhesus Macaques, Silver Springs State Park, Florida, USA. Emerg. Infect. Dis. 2018, 24, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, M.R.; Yee, J.; Barry, P.; Spinner, A.; Roberts, J.A.; Cabello, P.H.; Leite, J.P.; Lerche, N.W. Prevalence of antibodies to selected viruses in a long-term closed breeding colony of rhesus macaques (Macaca mulatta) in Brazil. Am. J. Primatol. 2003, 59, 123–128. [Google Scholar] [CrossRef]
- Weigler, B.J. Biology of B virus in macaque and human hosts: A review. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1992, 14, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Kozak, R.A.; Rutherford, C.; Richard-Greenblatt, M.; Chau, N.Y.E.; Cabrera, A.; Biondi, M.; Borlang, J.; Day, J.; Osiowy, C.; Ramachandran, S.; et al. Development and Evaluation of a Molecular Hepatitis A Virus Assay for Serum and Stool Specimens. Viruses 2022, 14, 159. [Google Scholar] [CrossRef]
- Balayan, M.S. Natural hosts of hepatitis A virus. Vaccine 1992, 10 (Suppl. S1), S27–S31. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.M.; Gupta, S.P. Chapter 10—Studies on Picornaviral Proteases and Their Inhibitors. In Viral Proteases and Their Inhibitors; Gupta, S.P., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 263–315. [Google Scholar]
- Shevtsova, Z.V.; Korzaia, L.I.; Lomovskaia, I.B.; Doroshenko, N.V.; Zairov, G.K. An outbreak of hepatitis A in imported rhesus monkeys. Vopr. Virusol. 1989, 34, 346–351. [Google Scholar] [PubMed]
- Emerson, S.U.; Tsarev, S.A.; Govindarajan, S.; Shapiro, M.; Purcell, R.H. A simian strain of hepatitis A virus, AGM-27, functions as an attenuated vaccine for chimpanzees. J. Infect. Dis. 1996, 173, 592–597. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.G.; Otto, G.; Colby, L.A. Chapter 28—Selected Zoonoses. In Laboratory Animal Medicine, 3rd ed.; Fox, J.G., Anderson, L.C., Otto, G.M., Pritchett-Corning, K.R., Whary, M.T., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 1313–1370. [Google Scholar]
- Bozkurt, H.; Phan-Thien, K.Y.; van Ogtrop, F.; Bell, T.; McConchie, R. Outbreaks, occurrence, and control of norovirus and hepatitis a virus contamination in berries: A review. Crit. Rev. Food Sci. Nutr. 2021, 61, 116–138. [Google Scholar] [CrossRef]
- Yeargin, T.; Gibson, K.E. Key characteristics of foods with an elevated risk for viral enteropathogen contamination. J. Appl. Microbiol. 2019, 126, 996–1010. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Ma, Y.; Gao, Y.; You, H. Epidemiology of Adenovirus Pneumonia and Risk Factors for Bronchiolitis Obliterans in Children during an Outbreak in Jilin, China. Front. Pediatr. 2021, 9, 722885. [Google Scholar] [CrossRef]
- Roy, S.; Sandhu, A.; Medina, A.; Clawson, D.S.; Wilson, J.M. Adenoviruses in fecal samples from asymptomatic rhesus macaques, United States. Emerg. Infect. Dis. 2012, 18, 1081–1088. [Google Scholar] [CrossRef]
- Sukmak, M.; Wajjwalku, W.; Ostner, J.; Schulke, O. A first report of non-invasive adenovirus detection in wild Assamese macaques in Thailand. Primates 2017, 58, 307–313. [Google Scholar] [CrossRef]
- de Melo Silva, J.; Pinheiro-Silva, R.; Costa de Oliveira, R.; de Castro Alves, C.E.; Barbosa, A.N.; Pontes, G.S. Prevalence and Recurrence Rates of Cytomegalovirus Infection among Patients with Hematological Diseases in the Western Brazilian Amazon: A Cross-Sectional Study. Front. Public Health 2021, 9, 692226. [Google Scholar] [CrossRef]
- Ambagala, A.P.; Marsh, A.; Chan, J.; Pilon, R.; Fournier, J.; Mazzulli, T.; Sandstrom, P.; Willer, D.O.; MacDonald, K.S. Isolation and characterization of cynomolgus macaque (Macaca fascicularis) cytomegalovirus (CyCMV). Virology 2011, 412, 125–135. [Google Scholar] [CrossRef]
- Barry, P.A.; Lockridge, K.M.; Salamat, S.; Tinling, S.P.; Yue, Y.; Zhou, S.S.; Gospe, S.M., Jr.; Britt, W.J.; Tarantal, A.F. Nonhuman primate models of intrauterine cytomegalovirus infection. ILAR J. 2006, 47, 49–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.L.; Tarantal, A.F.; Zhou, S.S.; Borowsky, A.D.; Barry, P.A. A recombinant rhesus cytomegalovirus expressing enhanced green fluorescent protein retains the wild-type phenotype and pathogenicity in fetal macaques. J. Virol. 2002, 76, 9493–9504. [Google Scholar] [CrossRef] [Green Version]
- Anoh, A.E.; Murthy, S.; Akoua-Koffi, C.; Couacy-Hymann, E.; Leendertz, F.H.; Calvignac-Spencer, S.; Ehlers, B. Cytomegaloviruses in a Community of Wild Nonhuman Primates in Taï National Park, Côte D’Ivoire. Viruses 2017, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Vogel, P.; Weigler, B.J.; Kerr, H.; Hendrickx, A.G.; Barry, P.A. Seroepidemiologic studies of cytomegalovirus infection in a breeding population of rhesus macaques. Lab. Anim. Sci. 1994, 44, 25–30. [Google Scholar] [PubMed]
- Blewett, E.L.; Lewis, J.; Gadsby, E.L.; Neubauer, S.R.; Eberle, R. Isolation of cytomegalovirus and foamy virus from the drill monkey (Mandrillus leucophaeus) and prevalence of antibodies to these viruses amongst wild-born and captive-bred individuals. Arch. Virol. 2003, 148, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Mouinga-Ondémé, A.; Kazanji, M. Simian foamy virus in non-human primates and cross-species transmission to humans in Gabon: An emerging zoonotic disease in central Africa? Viruses 2013, 5, 1536–1552. [Google Scholar] [CrossRef] [Green Version]
- Engel, G.A.; Small, C.T.; Soliven, K.; Feeroz, M.M.; Wang, X.; Kamrul Hasan, M.; Oh, G.; Rabiul Alam, S.M.; Craig, K.L.; Jackson, D.L.; et al. Zoonotic simian foamy virus in Bangladesh reflects diverse patterns of transmission and co-infection. Emerg. Microbes Infect. 2013, 2, e58. [Google Scholar] [CrossRef] [PubMed]
- Jones-Engel, L.; Engel, G.A.; Heidrich, J.; Chalise, M.; Poudel, N.; Viscidi, R.; Barry, P.A.; Allan, J.S.; Grant, R.; Kyes, R. Temple monkeys and health implications of commensalism, Kathmandu, Nepal. Emerg. Infect. Dis. 2006, 12, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Smiley Evans, T.; Barry, P.A.; Gilardi, K.V.; Goldstein, T.; Deere, J.D.; Fike, J.; Yee, J.; Ssebide, B.J.; Karmacharya, D.; Cranfield, M.R.; et al. Optimization of a Novel Non-invasive Oral Sampling Technique for Zoonotic Pathogen Surveillance in Nonhuman Primates. PLoS Negl. Trop. Dis. 2015, 9, e0003813. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Monkey NO. | Sampling Time | Growth Patterns | Sex | Age * |
|---|---|---|---|---|
| 1 | June 2014 | Captive | M | Sub-adult |
| 2 | June 2014 | Captive | M | Sub-adult |
| 3 | June 2014 | Captive | M | Adult |
| 4 | June 2014 | Captive | M | Adult |
| 5 | June 2014 | Captive | M | Adult |
| 6 | June 2014 | Captive | M | Adult |
| 7 | June 2014 | Captive | F | Sub-adult |
| 8 | June 2014 | Captive | F | Sub-adult |
| 9 | June 2014 | Captive | F | Sub-adult |
| 10 | June 2014 | Captive | F | Adult |
| 11 | June 2015 | Captive | F | Adult |
| 12 | June 2015 | Captive | F | Adult |
| 13 | June 2015 | Captive | F | Adult |
| 14 | June 2015 | Free ranging | M | Adult |
| 15 | June 2015 | Free ranging | M | Adult |
| 16 | June 2015 | Free ranging | M | Adult |
| 17 | June 2015 | Free ranging | M | Adult |
| 18 | June 2015 | Free ranging | M | Adult |
| 19 | June 2015 | Free ranging | F | Sub-adult |
| 20 | June 2015 | Free ranging | F | Sub-adult |
| 21 | January 2016 | Free ranging | F | Sub-adult |
| 22 | January 2016 | Free ranging | F | Adult |
| 23 | January 2016 | Free ranging | F | Adult |
| 24 | January 2016 | Free ranging | F | Adult |
| 25 | November 2016 | Free ranging | F | Adult |
| 26 | November 2016 | Free ranging | F | Adult |
| Target Viruses | Target Genes | Sequence (5’ to 3’) | Annealing Temperature | Length of Products | References |
|---|---|---|---|---|---|
| Adenovirus | hexon | TTCCCCATGGCICAYAACAC | 50 °C | 453 bp | [44] |
| CCCTGGTAKCCRATRTTGTA | |||||
| Rotavirus | VP7 | AAAGGATGGCCAACAGGATCAGT | 52 °C | 569 bp | [45] |
| GTATARAAHACTTGCCACCAT |
| Jun. 2014 (n = 10) | Jun. 2015 (n = 10) | Jan. 2016 (n = 4) | Nov. 2016 (n = 2) | Total (n = 26) | |
|---|---|---|---|---|---|
| Pathogens | TP% (95% CI: lower, upper) | TP% (95% CI: lower, upper) | TP% (95% CI: lower, upper) | TP% (95% CI: lower, upper) | TP% (95% CI: lower, upper) |
| MaHV-1 | 60.0 (26.2, 87.8) | 60.0 (26.2, 87.8) | 50.0 (6.8, 93.2) | 50.0 (1.3, 98.7) | 57.7 (36.9,76.6) |
| CV | 0.0 (0.0, 30.8) | 0.0 (0.0, 30.8) | 0.0 (0.0, 60.2) | 0.0 (0.0, 84.2) | 0.0 (0.0, 13.2) |
| HAV | 0.0 (0.0, 30.8) | 0.0 (0.0, 30.8) | 0.0 (0.0, 60.2) | 100.0 (15.8, 100) | 7.7 (0.0, 84.2) |
| Measles | 0.0 (0.0, 30.8) | 0.0 (0.0, 30.8) | 0.0 (0.0, 60.2) | 0.0 (0.0, 84.2) | 0.0 (0.0, 13.2) |
| M.tb | 0.0 (0.0, 30.8) | 0.0 (0.0, 30.8) | 0.0 (0.0, 60.2) | 0.0 (0.0, 84.2) | 0.0 (0.0, 13.2) |
| GsmCMV | 20.0 (2.5, 55.6) | 50.0 (6.8, 93.2) | 50.0 (6.8, 93.2) | 50.0 (1.3, 98.7) | 38.5 (20.2, 59.4) |
| SFV | 0.0 (0.0, 30.8) | 40.0 (12.2, 73.8) | 25.0 (0.6, 80.7) | 100.0 (15.8, 100) | 26.9 (11.6, 47.8) |
| SIV | 0.0 (0.0, 30.8) | 0.0 (0.0, 30.8) | 0.0 (0.0, 60.2) | 0.0 (0.0, 84.2) | 0.0 (0.0, 13.2) |
| SRV | 0.0 (0.0, 30.8) | 0.0 (0.0, 30.8) | 0.0 (0.0, 60.2) | 0.0 (0.0, 84.2) | 0.0 (0.0, 13.2) |
| STLV-1 | 0.0 (0.0, 30.8) | 0.0 (0.0, 30.8) | 0.0 (0.0, 60.2) | 0.0 (0.0, 84.2) | 0.0 (0.0, 13.2) |
| SV40 | 0.0 (0.0, 30.8) | 0.0 (0.0, 30.8) | 0.0 (0.0, 60.2) | 0.0 (0.0, 84.2) | 0.0 (0.0, 13.2) |
| SVV | 0.0 (0.0, 30.8) | 0.0 (0.0, 30.8) | 0.0 (0.0, 60.2) | 0.0 (0.0, 84.2) | 0.0 (0.0, 13.2) |
| Dec. 2014 | Jun. 2015 | Jan. 2016 | Total | |
|---|---|---|---|---|
| Pathogens | % (95% CI: lower, upper) | % (95% CI: lower, upper) | % (95% CI: lower, upper) | % (95% CI: lower, upper) |
| Adenovirus | 2.0 (0.6, 7.1) | 0 (0, 4.1) | 0 (0, 3.9) | 0.7 (0.2, 2.5) |
| Rotavirus | 0 (0, 3.8) | 0 (0, 4.1) | 0 (0, 3.9) | 0 (0, 0.13) |
| Variables | MaHV-1 | GsmCMV | SFV | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Crude OR | 95% CI | p value | Crude OR | 95% CI | p value | Crude OR | 95% CI | p value | ||
| Age | Adult vs. Sub-adult | 16.8 | 1.6, 176.2 | 0.005 | 5.4 | 0.5, 53.9 | 0.106 | - | - | - |
| Sex | Male vs. Female | 0.9 | 0.2, 4.3 | 0.850 | 0.9 | 0.2, 4.6 | 0.899 | 5.4 | 0.5, 53.9 | 0.106 |
| Growth patterns | Captive vs. Free-ranging | 0.4 | 0.1, 1.9 | 0.231 | 1.0 | 0.2, 4.9 | 1.0 | 1.5 | 0.3, 8.5 | 0.658 |
| Seasons | Summer vs. Winter | 1.5 | 0.2, 9.4 | 0.665 | 0.5 | 0.1, 3.4 | 0.512 | 0.3 | 0.0, 1.7 | 0.162 |
| Variables | MaHV-1 | GsmCMV | SFV | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Adjusted OR | 95% CI | p value | Adjusted OR | 95% CI | p value | Adjusted OR | 95% CI | p value | ||
| Age | Adult vs. Sub-adult | 33.0 | 2.5, 443.6 | 0.002 | 5.4 | 0.5, 53.9 | 0.106 | - | - | - |
| Sex | Male vs. Female | - | - | - | - | - | - | 5.4 | 0.5, 53.9 | 0.106 |
| Seasons | Summer vs. Winter | 5.5 | 0.6, 49.5 | 0.128 | - | - | - | - | - | - |
| Variables | Category | Crude OR (95% CI) | p Value | Adjusted OR (95% CI) | p Value |
|---|---|---|---|---|---|
| Sex | Female vs. Male | 0.6 (0.1, 3.6) | 0.524 | - | - |
| Age | Adult vs. Sub-adult | 21.3 (2.4, 191.6) | 0.003 | 21.3 (2.4, 191.6) | 0.003 |
| Growth pattern | Captive vs. Free-ranging | 0.3 (0.04, 1.9) | 0.179 | - | - |
| Season | Summer vs. Winter | 0.5 (0.04, 4.9) | 0.503 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, M.; Wang, Q.; Wang, Y.; Chen, Y.; Hu, C.; Yang, W.; Wu, F.; Huang, T.; Dawood, A.S.; Zubair, M.; et al. Epidemiological Survey and Risk Factor Analysis of 14 Potential Pathogens in Golden Snub-Nosed Monkeys at Shennongjia National Nature Reserve, China. Pathogens 2023, 12, 483. https://doi.org/10.3390/pathogens12030483
Qi M, Wang Q, Wang Y, Chen Y, Hu C, Yang W, Wu F, Huang T, Dawood AS, Zubair M, et al. Epidemiological Survey and Risk Factor Analysis of 14 Potential Pathogens in Golden Snub-Nosed Monkeys at Shennongjia National Nature Reserve, China. Pathogens. 2023; 12(3):483. https://doi.org/10.3390/pathogens12030483
Chicago/Turabian StyleQi, Mingpu, Qiankun Wang, Yu Wang, Yingyu Chen, Changmin Hu, Wanji Yang, Feng Wu, Tianpeng Huang, Ali Sobhy Dawood, Muhammad Zubair, and et al. 2023. "Epidemiological Survey and Risk Factor Analysis of 14 Potential Pathogens in Golden Snub-Nosed Monkeys at Shennongjia National Nature Reserve, China" Pathogens 12, no. 3: 483. https://doi.org/10.3390/pathogens12030483