
Migratory Wild Birds as Potential Long-Distance Transmitters of Toxoplasma gondii Infection

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population of Interest
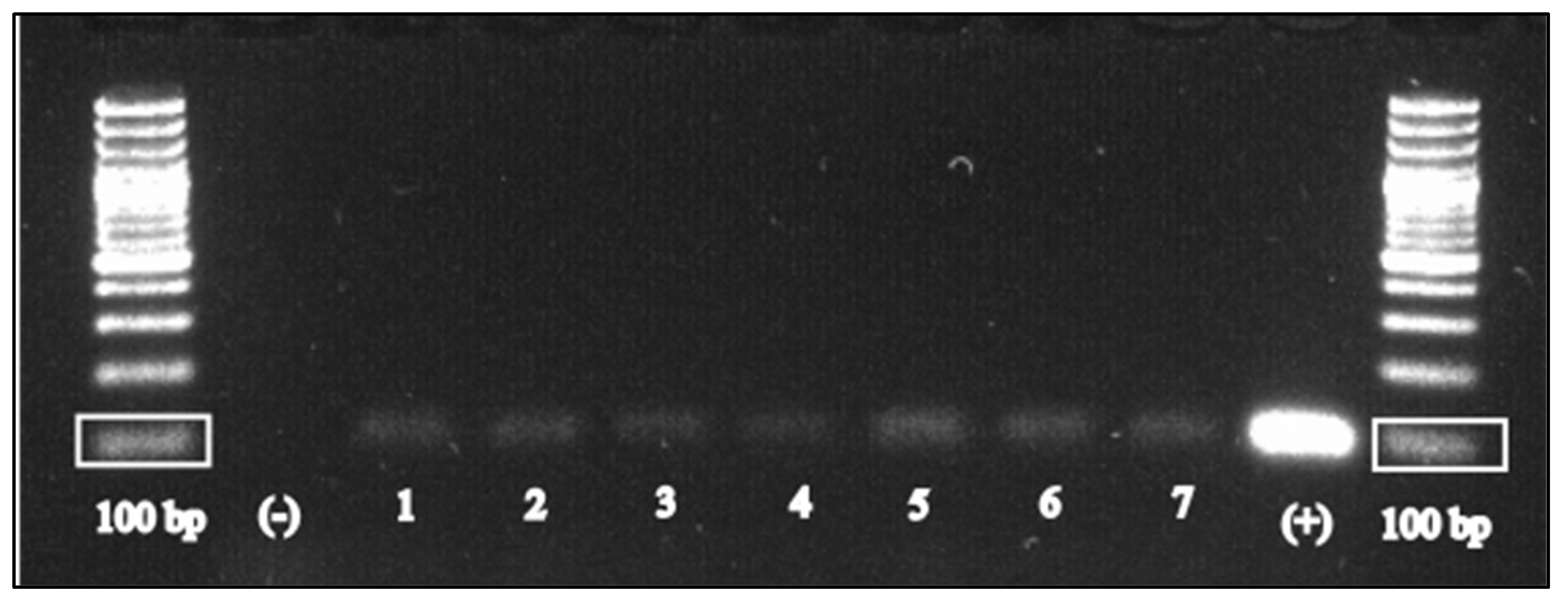
2.2. Molecular Detection of Toxoplasma gondii
2.3. Statistical Analyses

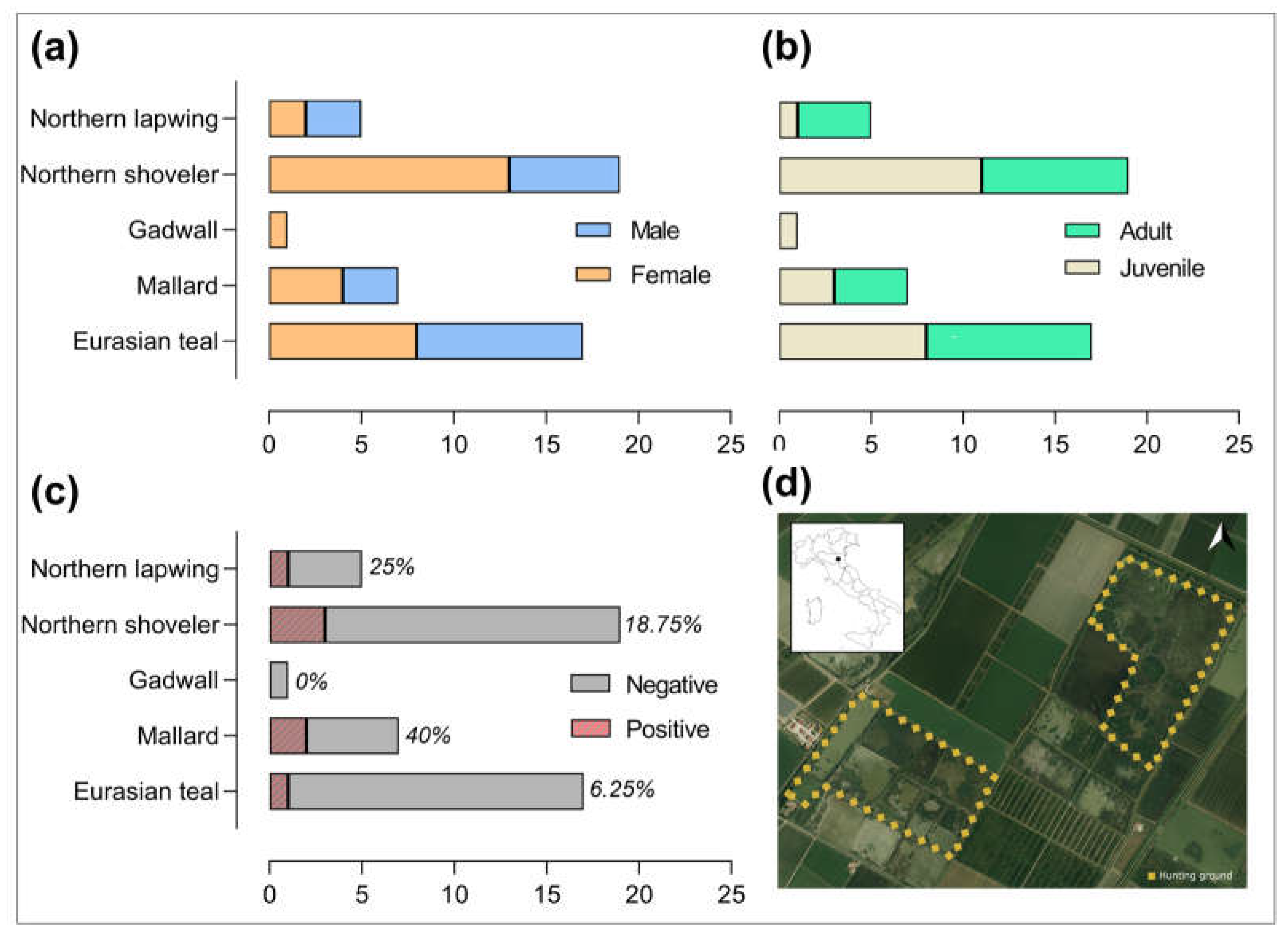
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dubey, J.P. The history and life cycle of Toxoplasma gondii. In Toxoplasma gondii; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–19. [Google Scholar]
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Shapiro, K.; Bahia-Oliveira, L.; Dixon, B.; Dumetre, A.; de Wit, L.A.; VanWormer, E.; Villena, I. Environmental transmission of Toxoplasma gondii: Oocysts in water, soil and food. Food Waterborne Parasitol. 2019, 15, e00049. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Long term persistence of Toxoplasma gondii in tissues of pigs inoculated with T. gondii oocysts and effect of freezing on viability of tissue cysts in pork. Am. J. Vet. Res. 1988, 49, 910–913. [Google Scholar] [PubMed]
- Akerstedt, J.; Lillehaug, A.; Larsen, I.-L.; Eide, N.E.; Arnemo, J.M.; Handeland, K. Serosurvey for canine distemper virus, canine adenovirus, Leptospira interrogans, and Toxoplasma gondii in free-ranging canids in Scandinavia and Svalbard. J. Wildl. Dis. 2010, 46, 474–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanet, S.; Sposimo, P.; Trisciuoglio, A.; Giannini, F.; Strumia, F.; Ferroglio, E. Epidemiology of Leishmania infantum, Toxoplasma gondii, and Neospora caninum in Rattus rattus in absence of domestic reservoir and definitive hosts. Vet. Parasitol. 2014, 199, 247–249. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Liang, L.J.; Zheng, K.Y.; Zhu, X.Q. Impact of environmental factors on the emergence, transmission and distribution of Toxoplasma gondii. Parasit Vectors 2016, 9, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenkel, J.K.; Hassanein, K.M.; Hassanein, R.S.; Brown, E.; Thulliez, P.; Quintero-Nunez, R. Transmission of Toxoplasma gondii in Panama City, Panama: A Five-Year Prospective Cohort Study of Children, Cats, Rodents, Birds, and Soil. Am. J. Trop. Med. 1995, 53, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. A review of toxoplasmosis in wild birds. Vet. Parasitol. 2002, 106, 121–153. [Google Scholar] [CrossRef]
- Wendte, J.M.; Gibson, A.K.; Grigg, M.E. Population genetics of Toxoplasma gondii: New perspectives from parasite genotypes in wildlife. Vet. Parasitol. 2011, 182, 96–111. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, K.; Largier, J.; Mazet, J.A.K.; Bernt, W.; Ell, J.R.; Melli, A.C.; Conrad, P.A. Surface properties of Toxoplasma gondii oocysts and surrogate microspheres. Appl. Environ. Microbiol. 2009, 75, 1185–1191. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.G.; Lapen, D.R.; Mitchell, G.W.; Provencher, J.F.; Wilson, S. Interaction of diet and habitat predicts Toxoplasma gondii infection rates in wild birds at a global scale. Glob. Ecol. Biogeogr. 2020, 29, 1189–1198. [Google Scholar] [CrossRef] [Green Version]
- Graziosi, G.; Catelli, E.; Fanelli, A.; Lupini, C. Infectious bursal disease virus in free-living wild birds: A systematic review and meta-analysis of its sero-viroprevalence on a global scale. Transbound. Emerg. Dis. 2021, 69, 2800–2815. [Google Scholar] [CrossRef] [PubMed]
- Graziosi, G.; Lupini, C.; Catelli, E. Disentangling the role of wild birds in avian metapneumovirus (aMPV) epidemiology: A systematic review and meta-analysis. Transbound. Emerg. Dis. 2022, 69, 3285–3299. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.A.; Delogu, M.; Facchini, M.; Di Trani, L.; Boni, A.; Cotti, C.; Graziosi, G.; Venturini, D.; Regazzi, D.; Ravaioli, V.; et al. Serologic Evidence of Occupational Exposure to Avian Influenza Viruses at the Wildfowl/Poultry/Human Interface. Microorganisms 2021, 9, 2153. [Google Scholar] [CrossRef] [PubMed]
- Benskin, C.M.; Wilson, K.; Jones, K.; Hartley, I.R. Bacterial pathogens in wild birds: A review of the frequency and effects of infection. Biol. Rev. Camb. Philos. Soc. 2009, 84, 349–373. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.J.; Yabsley, M.J.; Hernandez, S.M. A Review of Pathogen Transmission at the Backyard Chicken-Wild Bird Interface. Front. Vet. Sci. 2020, 7, 539925. [Google Scholar] [CrossRef]
- Zhang, F.K.; Wang, H.J.; Qin, S.Y.; Wang, Z.D.; Lou, Z.L.; Zhu, X.Q.; Liu, Q. Molecular detection and genotypic characterization of Toxoplasma gondii in wild waterfowls in Jilin Province, Northeastern China. Parasitol. Int. 2015, 64, 576–578. [Google Scholar] [CrossRef]
- Murao, T.; Omata, Y.; Kano, R.; Murata, S.; Okada, T.; Konnai, S.; Asakawa, M.; Ohashi, K.; Onuma, M. Serological Survey of Toxoplasma gondii in Wild Waterfowl in Chukotka, Kamchatka, Russia and Hokkaido, Japan. J. Parasitol. 2008, 94, 830–833. [Google Scholar] [CrossRef]
- Guo, M.; Dubey, J.P.; Hill, D.; Buchanan, R.L.; Gamble, H.R.; Jones, J.L.; Pradhan, A.K. Prevalence and Risk Factors for Toxoplasma gondii Infection in Meat Animals and Meat Products Destined for Human Consumption. J. Food Prot. 2015, 78, 457–476. [Google Scholar] [CrossRef]
- Jones, C.D.; Okhravi, N.; Adamson, P.; Tasker, S.; Lightman, S. Comparison of PCR Detection Methods for B1, P30, and 18S rDNA Genes of T. gondii in Aqueous Humor. Investig. Ophtalmol. Vis. Sci. 2000, 41, 634–644. [Google Scholar]
- Team, Q.D.; QGIS Geographic Information System. Open Source Geospatial Foundation. 2016. Available online: http://qgis.org/ (accessed on 10 December 2022).
- Adamska, M. Molecular detection of Toxoplasma gondii in natural surface water bodies in Poland. J. Water Health 2018, 16, 657–660. [Google Scholar] [CrossRef]
- Aramini, J.J.; Stephen, C.; Dubey, J.P.; Engelstoft, C.; Schwantje, H.; Ribble, C.S. Potential contamination of drinking water with Toxoplasma gondii oocysts. Epidemiol. Infect. 1999, 122, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Mancianti, F.; Nardoni, S.; Mugnaini, L.; Poli, A. Toxoplasma gondii in waterfowl: The first detection of this parasite in Anas crecca and Anas clypeata from Italy. J. Parasitol. 2013, 99, 561–563. [Google Scholar] [CrossRef] [PubMed]
- Nardoni, S.; Rocchigiani, G.; Varvaro, I.; Altomonte, I.; Ceccherelli, R.; Mancianti, F. Serological and Molecular Investigation on Toxoplasma gondii Infection in Wild Birds. Pathogens 2019, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puvanesuaran, V.R.; Noordin, R.; Balakrishnan, V. Isolation and genotyping of Toxoplasma gondii from free-range ducks in Malaysia. Avian Dis. 2013, 57, 128–132. [Google Scholar] [CrossRef]
- Aubert, D.; Ajzenberg, D.; Richomme, C.; Gilot-Fromont, E.; Terrier, M.E.; de Gevigney, C.; Game, Y.; Maillard, D.; Gibert, P.; Dardé, M.L.; et al. Molecular and biological characteristics of Toxoplasma gondii isolates from wildlife in France. Vet. Parasitol. 2010, 171, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Waldenström, J.; van Toor, M.; Lewis, N.; Lopes, S.; Javakhishvili, Z.; Muzika, D.; Fouchier, R.A.M.; Brouwer, A. Active wild bird surveillance of avian influenza viruses, a report. EFSA Support. Publ. 2022, 19, 7791E. [Google Scholar] [CrossRef]
- Fanelli, A.; Battisti, E.; Zanet, S.; Trisciuoglio, A.; Ferroglio, E. A systematic review and meta-analysis of Toxoplasma gondii in roe deer (Capreolus capreolus) and red deer (Cervus elaphus) in Europe. Zoonoses Public Health 2021, 68, 182–193. [Google Scholar] [CrossRef]
- Damek, F.; Swart, A.; Waap, H.; Jokelainen, P.; Le Roux, D.; Deksne, G.; Deng, H.; Schares, G.; Lunden, A.; Alvarez-Garcia, G.; et al. Systematic Review and Modelling of Age-Dependent Prevalence of Toxoplasma gondii in Livestock, Wildlife and Felids in Europe. Pathogens 2023, 12. [Google Scholar] [CrossRef]
- Schüz, E.; Berthold, P.; Gwinner, E.; Oelke, H. Grundriss der Vogelzugskunde; Verlag: Berlin, Germany, 1971. [Google Scholar]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H.; Grigg, M.E. Recent epidemiologic and clinical importance of Toxoplasma gondii infections in marine mammals: 2009–2020. Vet. Parasitol. 2020, 288, 109296. [Google Scholar] [CrossRef]
- Spina, F.; Volponi, S. Atlante Della Migrazione Degli Uccelli in Italia: Non-Passeriformi; Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA): Roma, Italy, 2008. [Google Scholar]
- Biancifiori, F.; Rondini, C.; Grelloni, V.; Frescura, T. Avian toxoplasmosis: Experimental infection of chicken and pigeon. Comp. Immunol. Microbiol. Infect. Dis. 1986, 9, 337–346. [Google Scholar] [CrossRef]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H.; Su, C. Epidemiologic significance of Toxoplasma gondii infections in turkeys, ducks, ratites and other wild birds: 2009-2020. Parasitology 2021, 148, 1–30. [Google Scholar] [CrossRef]
- Opsteegh, M.; Langelaar, M.; Sprong, H.; den Hartog, L.; De Craeye, S.; Bokken, G.; Ajzenberg, D.; Kijlstra, A.; van der Giessen, J. Direct detection and genotyping of Toxoplasma gondii in meat samples using magnetic capture and PCR. Int. J. Food Microbiol. 2010, 139, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Koethe, M.; Straubinger, R.K.; Pott, S.; Bangoura, B.; Geuthner, A.C.; Daugschies, A.; Ludewig, M. Quantitative detection of Toxoplasma gondii in tissues of experimentally infected turkeys and in retail turkey products by magnetic-capture PCR. Food Microbiol. 2015, 52, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Bachand, N.; Ravel, A.; Leighton, P.; Stephen, C.; Ndao, M.; Avard, E.; Jenkins, E. Serological and molecular detection of Toxoplasma gondii in terrestrial and marine wildlife harvested for food in Nunavik, Canada. Parasites Vectors 2019, 12, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Nie, L.B.; Zhang, N.Z.; Zou, F.C.; Zhu, X.Q.; Cong, W. First genetic characterization of Toxoplasma gondii infection in poultry meat intended for human consumption in eastern China. Infect. Genet. Evol. 2017, 55, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Work, T.M.; Verma, S.K.; Su, C.; Medeiros, J.; Kaiakapu, T.; Kwok, O.C.; Dubey, J.P. Toxoplasma gondii antibody prevalence and two new genotypes of the parasite in endangered hawaiian geese (Nene: Branta sandvicensis). J. Wildl. Dis. 2016, 52, 253–257, 255. [Google Scholar] [CrossRef] [PubMed]
- Skorpikova, L.; Reslova, N.; Lorencova, A.; Plhal, R.; Drimaj, J.; Kamler, J.; Slany, M. Molecular detection of Toxoplasma gondii in feathered game intended for human consumption in the Czech Republic. Int. J. Food Microbiol. 2018, 286, 75–79. [Google Scholar] [CrossRef]
- Karakavuk, M.; Aldemir, D.; Mercier, A.; Atalay Şahar, E.; Can, H.; Murat, J.B.; Döndüren, Ö.; Can, Ş.; Özdemir, H.G.; Değirmenci Döşkaya, A.; et al. Prevalence of toxoplasmosis and genetic characterization of Toxoplasma gondii strains isolated in wild birds of prey and their relation with previously isolated strains from Turkey. PLoS ONE 2018, 13, e0196159. [Google Scholar] [CrossRef] [Green Version]
- Opsteegh, M.; Teunis, P.; Züchner, L.; Koets, A.; Langelaar, M.; van der Giessen, J. Low predictive value of seroprevalence of Toxoplasma gondii in cattle for detection of parasite DNA. Int. J. Parasitol. 2011, 41, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Sandström, C.A.; Buma, A.G.; Hoye, B.J.; Prop, J.; van der Jeugd, H.; Voslamber, B.; Madsen, J.; Loonen, M.J. Latitudinal variability in the seroprevalence of antibodies against Toxoplasma gondii in non-migrant and Arctic migratory geese. Vet. Parasitol. 2013, 194, 9–15, Epub 2012 Dec 20. [Google Scholar] [CrossRef] [PubMed]
- Vitaliano, S.N.; Mineo, T.W.P.; André, M.R.; Machado, R.Z.; Mineo, J.R.; Werther, K.; Donato Castellane, P. Experimental infection of Crested Caracara (Caracara plancus) with Toxoplasma gondii simulating natural conditions. Vet. Parasitol. 2010, 172, 71–75. [Google Scholar] [CrossRef]
- Ross, R.D.; Stec, L.A.; Werner, J.C.; Blumenkranz, M.S.; Glazer, L.; Williams, G.A. Presumed acquired ocular toxoplasmosis in deer hunters. Retina 2001, 21, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Conrady, C.D.; Besirli, C.G.; Baumal, C.R.; Kovach, J.L.; Etzel, J.D.; Tsui, J.C.; Elner, S.G.; Johnson, M.W. Ocular Toxoplasmosis after Exposure to Wild Game. Ocul. Immunol. Inflamm. 2022, 30, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.L. Documented Outbreaks of Toxoplasmosis: Transmission of Toxoplasma gondii to Humans. J. Food Prot. 1993, 56, 630–639. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dini, F.M.; Graziosi, G.; Lupini, C.; Catelli, E.; Galuppi, R. Migratory Wild Birds as Potential Long-Distance Transmitters of Toxoplasma gondii Infection. Pathogens 2023, 12, 478. https://doi.org/10.3390/pathogens12030478
Dini FM, Graziosi G, Lupini C, Catelli E, Galuppi R. Migratory Wild Birds as Potential Long-Distance Transmitters of Toxoplasma gondii Infection. Pathogens. 2023; 12(3):478. https://doi.org/10.3390/pathogens12030478
Chicago/Turabian StyleDini, Filippo Maria, Giulia Graziosi, Caterina Lupini, Elena Catelli, and Roberta Galuppi. 2023. "Migratory Wild Birds as Potential Long-Distance Transmitters of Toxoplasma gondii Infection" Pathogens 12, no. 3: 478. https://doi.org/10.3390/pathogens12030478