
Bioactive Antimicrobial Peptides from Food Proteins: Perspectives and Challenges for Controlling Foodborne Pathogens
 , , and
, , and
Abstract
:1. Introduction
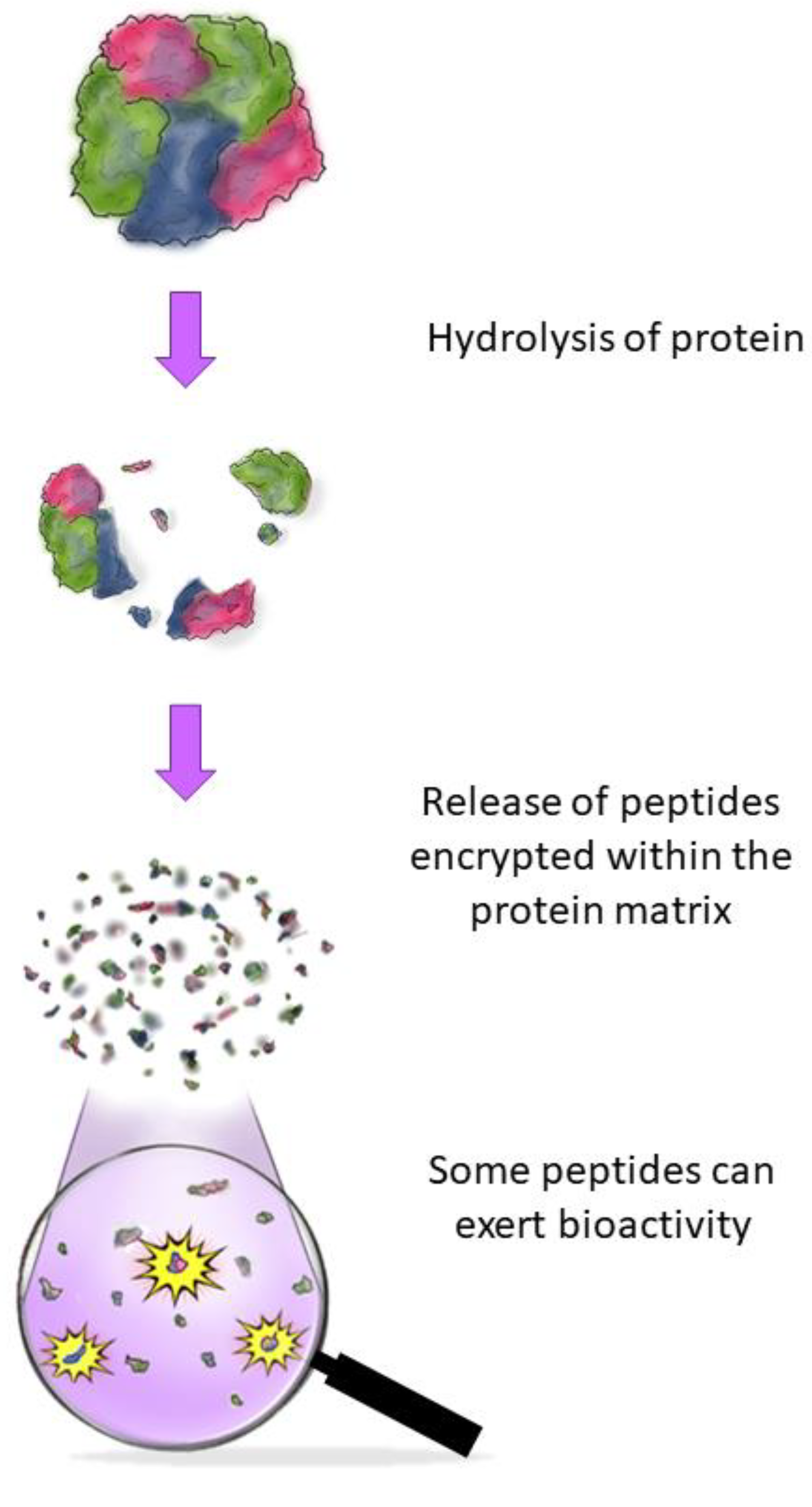
2. Production of BAPs and Hydrolysates
2.1. Sources of BAPs: A Route to Valorize By-Products
2.2. Encrypted Peptides and How to Find Them: The Importance of In Silico Approaches
3. The Antimicrobial Activity of Food-Sourced Peptides
3.1. Mechanism of Antimicrobial Activity of BAPs
Gut Microbiota Modulation and Immunomodulation as Mechanisms of Action: Gaps to Be Filled
4. The Endless Potential of a Vanguard: Lactic Acid Bacteria
5. Current Challenges to the Implementation of Bioactive Peptides in the Food Industry
6. Prospects
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toldrá, F.; Gallego, M.; Reig, M.; Aristoy, M.C.; Mora, L. Bioactive Peptides Generated in the Processing of Dry-Cured Ham. Food Chem. 2020, 321, 126689. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Kehinde, B.A.; Sharma, P.; Sharma, D.; Kaur, S. Recently Isolated Food-Derived Antihypertensive Hydrolysates and Peptides: A Review. Food Chem. 2021, 346, 128719. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.; de Souza Rocha, T.; Prudencio, S.H. Potential of Green and Roasted Coffee Beans and Spent Coffee Grounds to Provide Bioactive Peptides. Food Chem. 2021, 348, 129061. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, S.; Gulzar, N.; Sameen, A.; Huma, N.; Hayat, I.; Ijaz, R. Functional Role of Bioactive Peptides with Special Reference to Cheeses. Int. J. Dairy Technol. 2021, 74, 1–16. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM Structure of the 2019-NCoV Spike in the Prefusion Conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Shen, P.; Ding, K.; Wang, L.; Tian, J.; Huang, X.; Zhang, M.; Dang, X. In Vitro and in Vivo Antimicrobial Activity of Antimicrobial Peptide Jelleine-I against Foodborne Pathogen Listeria Monocytogenes. Int. J. Food Microbiol. 2023, 387, 110050. [Google Scholar] [CrossRef]
- Ma, X.; Cui, X.; Li, J.; Li, C.; Wang, Z. Peptides from Sesame Cake Reduce Oxidative Stress and Amyloid-β-Induced Toxicity by Upregulation of SKN-1 in a Transgenic Caenorhabditis Elegans Model of Alzheimer’s Disease. J. Funct. Foods 2017, 39, 287–298. [Google Scholar] [CrossRef]
- Zhang, Z.; He, S.; Cao, X.; Ye, Y.; Yang, L.; Wang, J.; Liu, H.; Sun, H. Potential Prebiotic Activities of Soybean Peptides Maillard Reaction Products on Modulating Gut Microbiota to Alleviate Aging-Related Disorders in D-Galactose-Induced ICR Mice. J. Funct. Foods 2020, 65, 103729. [Google Scholar] [CrossRef]
- Yardeni, T.; Tanes, C.E.; Bittinger, K.; Mattei, L.M.; Schaefer, P.M.; Singh, L.N.; Wu, G.D.; Murdock, D.G.; Wallace, D.C. Host Mitochondria Influence Gut Microbiome Diversity: A Role for ROS. Sci. Signal. 2019, 12, eaaw3159. [Google Scholar] [CrossRef]
- Wu, S.; Bekhit, A.E.-D.A.; Wu, Q.; Chen, M.; Liao, X.; Wang, J.; Ding, Y. Bioactive Peptides and Gut Microbiota: Candidates for a Novel Strategy for Reduction and Control of Neurodegenerative Diseases. Trends Food Sci. Technol. 2021, 108, 164–176. [Google Scholar] [CrossRef]
- Wang, S.; Sun-Waterhouse, D.; Neil Waterhouse, G.I.; Zheng, L.; Su, G.; Zhao, M. Effects of Food-Derived Bioactive Peptides on Cognitive Deficits and Memory Decline in Neurodegenerative Diseases: A Review. Trends Food Sci. Technol. 2021, 116, 712–732. [Google Scholar] [CrossRef]
- Vitetta, L.; Saltzman, E.; Nikov, T.; Ibrahim, I.; Hall, S. Modulating the Gut Micro-Environment in the Treatment of Intestinal Parasites. J. Clin. Med. 2016, 5, 102. [Google Scholar] [CrossRef] [Green Version]
- Swaggerty, C.; Callaway, T.; Kogut, M.; Piva, A.; Grilli, E. Modulation of the Immune Response to Improve Health and Reduce Foodborne Pathogens in Poultry. Microorganisms 2019, 7, 65. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Ko, G. Antiviral Effect of Vitamin A on Norovirus Infection via Modulation of the Gut Microbiome. Sci. Rep. 2016, 6, 25835. [Google Scholar] [CrossRef] [Green Version]
- Evangelista, A.G.; Danielski, G.M.; Corrêa, J.A.F.; Cavalari, C.M.d.A.; Souza, I.R.; Luciano, F.B.; de Macedo, R.E.F. Carnobacterium as a Bioprotective and Potential Probiotic Culture to Improve Food Quality, Food Safety, and Human Health–a Scoping Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–14. [Google Scholar] [CrossRef]
- Chauhan, V.; Kanwar, S.S. Bioactive Peptides. In Biotechnological Production of Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2020; pp. 107–137. ISBN 9780444643230. [Google Scholar]
- Etemadian, Y.; Ghaemi, V.; Shaviklo, A.R.; Pourashouri, P.; Sadeghi Mahoonak, A.R.; Rafipour, F. Development of Animal/Plant-Based Protein Hydrolysate and Its Application in Food, Feed and Nutraceutical Industries: State of the Art. J. Clean. Prod. 2021, 278, 123219. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial Peptides as Potential Alternatives to Antibiotics in Food Animal Industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef]
- Corrêa, J.A.F.; dos Santos, J.V.G.; Evangelista, A.G.; Pinto, A.C.S.M.; de Macedo, R.E.F.; Luciano, F.B. Combined Application of Phenolic Acids and Essential Oil Components against Salmonella Enteritidis and Listeria Monocytogenes in Vitro and in Ready-to-Eat Cooked Ham. LWT 2021, 149, 111881. [Google Scholar] [CrossRef]
- Mohammadrezaei, M.; Navidshad, B.; Gheisari, A.; Toghyani, M. Cottonseed Meal Bioactive Peptides as an Alternative to Antibiotic Growth Promoters in Broiler Chicks. Int. J. Pept. Res. Ther. 2021, 27, 329–340. [Google Scholar] [CrossRef]
- Pan, S.; Agyei, D.; Jeevanandam, J.; Danquah, M.K. Bio-Active Peptides: Role in Plant Growth and Defense. In Natural Bio-Active Compounds; Springer: Singapore, 2019; pp. 1–29. [Google Scholar]
- León Madrazo, A.; Segura Campos, M.R. Review of Antimicrobial Peptides as Promoters of Food Safety: Limitations and Possibilities within the Food Industry. J. Food Saf. 2020, 40, e12854. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, Z.; Dai, Z.; Wang, G.; Wu, G. Protein Hydrolysates in Animal Nutrition: Industrial Production, Bioactive Peptides, and Functional Significance. J. Anim. Sci. Biotechnol. 2017, 8, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartal, C.; Kaplan Türköz, B.; Otles, S. Prediction, Identification and Evaluation of Bioactive Peptides from Tomato Seed Proteins Using in Silico Approach. J. Food Meas. Charact. 2020, 14, 1865–1883. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Quirós, A.; Amigo, L.; Recio, I. Identification of Bioactive Peptides after Digestion of Human Milk and Infant Formula with Pepsin and Pancreatin. Int. Dairy J. 2007, 17, 42–49. [Google Scholar] [CrossRef]
- Alnuaimi, A.; Fisayo Ajayi, F.; Hamdi, M.; Mudgil, P.; Kamal, H.; Yuen Gan, C.; Maqsood, S. A Comparative Analysis of Anti-Lipidemic Potential of Soybean (Glycine Max) Protein Hydrolysates Obtained from Different Ripening Stages: Identification, and Molecular Interaction Mechanisms of Novel Bioactive Peptides. Food Chem. 2023, 402, 134192. [Google Scholar] [CrossRef]
- Martini, S.; Conte, A.; Tagliazucchi, D. Effect of Ripening and in Vitro Digestion on the Evolution and Fate of Bioactive Peptides in Parmigiano-Reggiano Cheese. Int. Dairy J. 2020, 105, 104668. [Google Scholar] [CrossRef]
- Atanasova, J.; Dalgalarrondo, M.; Iliev, I.; Moncheva, P.; Todorov, S.D.; Ivanova, I.V. Formation of Free Amino Acids and Bioactive Peptides During the Ripening of Bulgarian White Brined Cheeses. Probiotics Antimicrob. Proteins 2021, 13, 261–272. [Google Scholar] [CrossRef]
- Gao, J.; Wang, Y.; Yan, Y.; Li, Z.; Chen, M. Protein Extraction from Excess Sludge by Alkali-Thermal Hydrolysis. Environ. Sci. Pollut. Res. 2020, 27, 8628–8637. [Google Scholar] [CrossRef]
- Pasupuleti, V.K.; Holmes, C.; Demain, A.L. Applications of Protein Hydrolysates in Biotechnology. In Protein Hydrolysates in Biotechnology; Springer: Dordrecht, The Netherlands, 2008; pp. 1–9. [Google Scholar]
- Connolly, A.; Cermeño, M.; Crowley, D.; O’Callaghan, Y.; O’Brien, N.M.; FitzGerald, R.J. Characterisation of the in Vitro Bioactive Properties of Alkaline and Enzyme Extracted Brewers’ Spent Grain Protein Hydrolysates. Food Res. Int. 2019, 121, 524–532. [Google Scholar] [CrossRef]
- Nasri, R.; Abdelhedi, O.; Nasri, M.; Jridi, M. Fermented Protein Hydrolysates: Biological Activities and Applications. Curr. Opin. Food Sci. 2022, 43, 120–127. [Google Scholar] [CrossRef]
- Rajendran, S.R.C.K.; Mohan, A.; Khiari, Z.; Udenigwe, C.C.; Mason, B. Yield, Physicochemical, and Antioxidant Properties of Atlantic Salmon Visceral Hydrolysate: Comparison of Lactic Acid Bacterial Fermentation with Flavourzyme Proteolysis and Formic Acid Treatment. J. Food Process. Preserv. 2018, 42, e13620. [Google Scholar] [CrossRef]
- Sanjukta, S.; Rai, A.K. Production of Bioactive Peptides during Soybean Fermentation and Their Potential Health Bene Fi Ts. Trends Food Sci. Technol. 2016, 50, 1–10. [Google Scholar] [CrossRef]
- Ulug, S.K.; Jahandideh, F.; Wu, J. Novel Technologies for the Production of Bioactive Peptides. Trends Food Sci. Technol. 2021, 108, 27–39. [Google Scholar] [CrossRef]
- Chew, L.Y.; Toh, G.T.; Ismail, A. Application of Proteases for the Production of Bioactive Peptides. In Enzymes in Food Biotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 247–261. [Google Scholar]
- Liu, C.-F.; Pan, T.-M. Recombinant Expression of Bioactive Peptide Lunasin in Escherichia Coli. Appl. Microbiol. Biotechnol. 2010, 88, 177–186. [Google Scholar] [CrossRef]
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive Peptides: Synthesis, Sources, Applications, and Proposed Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 1445. [Google Scholar] [CrossRef] [PubMed]
- Karami, Z.; Akbari-adergani, B.; Duangmal, K. Recent Development on Recovering Bioactive Peptides and Phenolic Compounds from Under-utilised By-products during Production of Certain Edible Oil Plants: Current Situation and Future Perspectives. Int. J. Food Sci. Technol. 2022, 57, 4894–4905. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Tagliazucchi, D.; Babini, E.; Sefora Rutella, G.; Taneyo Saa, D.L.; Gianotti, A. Bioactive Peptides from Vegetable Food Matrices: Research Trends and Novel Biotechnologies for Synthesis and Recovery. J. Funct. Foods 2016, 27, 549–569. [Google Scholar] [CrossRef]
- Ali, M.A.; Kamal, M.M.; Rahman, M.H.; Siddiqui, M.N.; Haque, M.A.; Saha, K.K.; Rahman, M.A. Functional Dairy Products as a Source of Bioactive Peptides and Probiotics: Current Trends and Future Prospectives. J. Food Sci. Technol. 2022, 59, 1263–1279. [Google Scholar] [CrossRef]
- Samtiya, M.; Samtiya, S.; Badgujar, P.C.; Puniya, A.K.; Dhewa, T.; Aluko, R.E. Health-Promoting and Therapeutic Attributes of Milk-Derived Bioactive Peptides. Nutrients 2022, 14, 3001. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Dhewa, T.; Moreno-Rojas, J.M. Potential Health Benefits of Plant Food-Derived Bioactive Components: An Overview. Foods 2021, 10, 839. [Google Scholar] [CrossRef]
- Maestri, E.; Marmiroli, M.; Marmiroli, N. Bioactive Peptides in Plant-Derived Foodstuffs. J. Proteomics 2016, 147, 140–155. [Google Scholar] [CrossRef] [PubMed]
- López-García, G.; Dublan-García, O.; Arizmendi-Cotero, D.; Oliván, L.M.G. Antioxidant and Antimicrobial Peptides Derived from Food Proteins. Molecules 2022, 27, 1343. [Google Scholar] [CrossRef] [PubMed]
- Ali Redha, A.; Valizadenia, H.; Siddiqui, S.A.; Maqsood, S. A State-of-Art Review on Camel Milk Proteins as an Emerging Source of Bioactive Peptides with Diverse Nutraceutical Properties. Food Chem. 2022, 373, 131444. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.; Chun, B.-S. Subcritical Water Hydrolysis for the Production of Bioactive Peptides from Tuna Skin Collagen. J. Supercrit. Fluids 2018, 141, 88–96. [Google Scholar] [CrossRef]
- Je, J.-Y.; Qian, Z.-J.; Byun, H.-G.; Kim, S.-K. Purification and Characterization of an Antioxidant Peptide Obtained from Tuna Backbone Protein by Enzymatic Hydrolysis. Process Biochem. 2007, 42, 840–846. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; Alashi, M.A.; Aluko, R.E.; FitzGerald, R.J. Peptide Identification in a Salmon Gelatin Hydrolysate with Antihypertensive, Dipeptidyl Peptidase IV Inhibitory and Antioxidant Activities. Food Res. Int. 2017, 100, 112–120. [Google Scholar] [CrossRef]
- Alemán, A.; Gómez-Guillén, M.C.; Montero, P. Identification of Ace-Inhibitory Peptides from Squid Skin Collagen after in Vitro Gastrointestinal Digestion. Food Res. Int. 2013, 54, 790–795. [Google Scholar] [CrossRef]
- Kobbi, S.; Bougatef, A.; Le Flem, G.; Balti, R.; Mickael, C.; Fertin, B.; Chaabouni, S.; Dhulster, P.; Nedjar, N. Purification and Recovery of RuBisCO Protein from Alfalfa Green Juice: Antioxidative Properties of Generated Protein Hydrolysate. Waste Biomass Valorization 2017, 8, 493–504. [Google Scholar] [CrossRef]
- Caballero, P.; Rodríguez-Morgado, B.; Macías, S.; Tejada, M.; Parrado, J. Obtaining Plant and Soil Biostimulants by Waste Whey Fermentation. Waste Biomass Valorization 2020, 11, 3281–3292. [Google Scholar] [CrossRef]
- Borrajo, P.; Pateiro, M.; Barba, F.J.; Mora, L.; Franco, D.; Toldrá, F.; Lorenzo, J.M. Antioxidant and Antimicrobial Activity of Peptides Extracted from Meat By-Products: A Review. Food Anal. Methods 2019, 12, 2401–2415. [Google Scholar] [CrossRef]
- Mariam, S.H.; Zegeye, N.; Aseffa, A.; Howe, R. Diffusible Substances from Lactic Acid Bacterial Cultures Exert Strong Inhibitory Effects on Listeria Monocytogenes and Salmonella Enterica Serovar Enteritidis in a Co-Culture Model. BMC Microbiol. 2017, 17, 35. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, K.; Pineda-hidalgo, K.V.; Rochín-medina, J.J. Fermentation of Spent Coffee Grounds by Bacillus Clausii Induces Release of Potentially Bioactive Peptides. LWT 2021, 138, 110685. [Google Scholar] [CrossRef]
- Ramírez, K.; Quintero-Soto, M.F.; Rochín-Medina, J.J. Enhancement of the Antioxidant and Antimicrobial Activities of Maize Wastewater by an Eco-Friendly Process. J. Food Meas. Charact. 2020, 14, 1682–1689. [Google Scholar] [CrossRef]
- Khubber, S.; Marti-Quijal, F.J.; Tomasevic, I.; Remize, F.; Barba, F.J. Lactic Acid Fermentation as a Useful Strategy to Recover Antimicrobial and Antioxidant Compounds from Food and By-Products. Curr. Opin. Food Sci. 2022, 43, 189–198. [Google Scholar] [CrossRef]
- Worsztynowicz, P.; Białas, W.; Grajek, W. Integrated Approach for Obtaining Bioactive Peptides from Whey Proteins Hydrolysed Using a New Proteolytic Lactic Acid Bacteria. Food Chem. 2020, 312, 126035. [Google Scholar] [CrossRef]
- López-Barrios, L.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bioactive Peptides and Hydrolysates from Pulses and Their Potential Use as Functional Ingredients. J. Food Sci. 2014, 79, 134–149. [Google Scholar] [CrossRef]
- Buitimea-Cantúa, N.E.; Antunes-Ricardo, M.; Gutiérrez-Uribe, J.A.; del Refugio Rocha-Pizaña, M.; de la Rosa-Millán, J.; Torres-Chávez, P.I. Protein-Phenolic Aggregates with Anti-Inflammatory Activity Recovered from Maize Nixtamalization Wastewaters (Nejayote). LWT 2020, 134, 109881. [Google Scholar] [CrossRef]
- Udenigwe, C.C. Bioinformatics Approaches, Prospects and Challenges of Food Bioactive Peptide Research. Trends Food Sci. Technol. 2014, 36, 137–143. [Google Scholar] [CrossRef]
- Corrêa, J.A.F.; Udenigwe, C.; Bittencourt Luciano, F. Avaliação in Vitro de Atividade Antimicrobiana de Peptídeos Anfipáticos Derivados de Proteínas Do Soro de Leite. Rev. Acadêmica Ciência Anim. 2018, 16, 1–6. [Google Scholar] [CrossRef]
- Brand, G.D.; Magalhães, M.T.Q.; Tinoco, M.L.P.; Aragão, F.J.L.; Nicoli, J.; Kelly, S.M.; Cooper, A.; Bloch, C. Probing Protein Sequences as Sources for Encrypted Antimicrobial Peptides. PLoS ONE 2012, 7, e45848. [Google Scholar] [CrossRef]
- Corrêa, J.A.F.; Evangelista, A.G.; de Melo Nazareth, T.; Luciano, F.B. Fundamentals on the Molecular Mechanism of Action of Antimicrobial Peptides. Materialia 2019, 8, 100494. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Virtual—A Novel Database of Food-Derived Peptides with In Silico-Predicted Biological Activity. Appl. Sci. 2022, 12, 7204. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP Database and Other Programs for Processing Bioactive Peptide Sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [CrossRef] [Green Version]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. CAMP R3: A Database on Sequences, Structures and Signatures of Antimicrobial Peptides. Nucleic Acids Res. 2016, 44, D1094–D1097. [Google Scholar] [CrossRef] [Green Version]
- Freitas, C.S.; Vericimo, M.A.; da Silva, M.L.; da Costa, G.C.V.; Pereira, P.R.; Paschoalin, V.M.F.; Del Aguila, E.M. Encrypted Antimicrobial and Antitumoral Peptides Recovered from a Protein-Rich Soybean (Glycine Max) by-Product. J. Funct. Foods 2019, 54, 187–198. [Google Scholar] [CrossRef]
- Porto, W.F.; Irazazabal, L.; Alves, E.S.F.; Ribeiro, S.M.; Matos, C.O.; Pires, Á.S.; Fensterseifer, I.C.M.; Miranda, V.J.; Haney, E.F.; Humblot, V.; et al. In Silico Optimization of a Guava Antimicrobial Peptide Enables Combinatorial Exploration for Peptide Design. Nat. Commun. 2018, 9, 1490. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Oh, D.-H. Integration of Nisin into Nanoparticles for Application in Foods. Innov. Food Sci. Emerg. Technol. 2016, 34, 376–384. [Google Scholar] [CrossRef]
- Huang, F.; Teng, K.; Liu, Y.; Wang, T.; Xia, T.; Yun, F.; Zhong, J. Nisin Z Attenuates Lipopolysaccharide-Induced Mastitis by Inhibiting the ERK1/2 and P38 Mitogen-Activated Protein Kinase Signaling Pathways. J. Dairy Sci. 2022, 105, 3530–3543. [Google Scholar] [CrossRef]
- Bennett, S.; Fliss, I.; Ben Said, L.; Malouin, F.; Lacasse, P. Efficacy of Bacteriocin-Based Formula for Reducing Staphylococci, Streptococci, and Total Bacterial Counts on Teat Skin of Dairy Cows. J. Dairy Sci. 2022, 105, 4498–4507. [Google Scholar] [CrossRef]
- Chourasia, R.; Phukon, L.C.; Abedin, M.M.; Padhi, S.; Singh, S.P.; Rai, A.K. Whey Valorization by Microbial and Enzymatic Bioprocesses for the Production of Nutraceuticals and Value-Added Products. Bioresour. Technol. Rep. 2022, 19, 101144. [Google Scholar] [CrossRef]
- Svendsen, J.S.M.; Grant, T.M.; Rennison, D.; Brimble, M.A.; Svenson, J. Very Short and Stable Lactoferricin-Derived Antimicrobial Peptides: Design Principles and Potential Uses. Acc. Chem. Res. 2019, 52, 749–759. [Google Scholar] [CrossRef]
- Xiao, H.; Shao, F.; Wu, M.; Ren, W.; Xiong, X.; Tan, B.; Yin, Y. The Application of Antimicrobial Peptides as Growth and Health Promoters for Swine. J. Anim. Sci. Biotechnol. 2015, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Evangelista, A.G.; Corrêa, J.A.F.; Pinto, A.C.S.M.; Luciano, F.B. The Impact of Essential Oils on Antibiotic Use in Animal Production Regarding Antimicrobial Resistance—A Review. Crit. Rev. Food Sci. Nutr. 2021, 62, 5267–5283. [Google Scholar] [CrossRef]
- Wang, Y.; Shan, T.; Xu, Z.; Liu, J.; Feng, J. Effect of Lactoferrin on the Growth Performance, Intestinal Morphology, and Expression of PR-39 and Protegrin-1 Genes in Weaned Piglets1. J. Anim. Sci. 2006, 84, 2636–2641. [Google Scholar] [CrossRef]
- Tang, Z.; Yin, Y.; Zhang, Y.; Huang, R.; Sun, Z.; Li, T.; Chu, W.; Kong, X.; Li, L.; Geng, M.; et al. Effects of Dietary Supplementation with an Expressed Fusion Peptide Bovine Lactoferricin–Lactoferrampin on Performance, Immune Function and Intestinal Mucosal Morphology in Piglets Weaned at Age 21 D. Br. J. Nutr. 2008, 101, 998–1005. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Filho, J.G.; Rodrigues, J.M.; Valadares, A.C.F.; de Almeida, A.B.; Valencia-Mejia, E.; Fernandes, K.F.; Lemes, A.C.; Alves, C.C.F.; Sousa, H.A.d.F.; da Silva, E.R.; et al. Bioactive Properties of Protein Hydrolysate of Cottonseed Byproduct: Antioxidant, Antimicrobial, and Angiotensin-Converting Enzyme (ACE) Inhibitory Activities. Waste Biomass Valorization 2021, 12, 1395–1404. [Google Scholar] [CrossRef]
- Martín, I.; Rodríguez, A.; Alía, A.; Martínez-Blanco, M.; Lozano-Ojalvo, D.; Córdoba, J.J. Control of Listeria Monocytogenes Growth and Virulence in a Traditional Soft Cheese Model System Based on Lactic Acid Bacteria and a Whey Protein Hydrolysate with Antimicrobial Activity. Int. J. Food Microbiol. 2022, 361, 109444. [Google Scholar] [CrossRef]
- Demers-Mathieu, V.; Gauthier, S.F.; Britten, M.; Fliss, I.; Robitaille, G.; Jean, J. Antibacterial Activity of Peptides Extracted from Tryptic Hydrolyzate of Whey Protein by Nanofiltration. Int. Dairy J. 2013, 28, 94–101. [Google Scholar] [CrossRef]
- Mishra, B.; Felix, L.; Basu, A.; Kollala, S.S.; Chhonker, Y.S.; Ganesan, N.; Murry, D.J.; Mylonakis, E. Design and Evaluation of Short Bovine Lactoferrin-Derived Antimicrobial Peptides against Multidrug-Resistant Enterococcus Faecium. Antibiotics 2022, 11, 1085. [Google Scholar] [CrossRef]
- Memarpoor-Yazdi, M.; Asoodeh, A.; Chamani, J. A Novel Antioxidant and Antimicrobial Peptide from Hen Egg White Lysozyme Hydrolysates. J. Funct. Foods 2012, 4, 278–286. [Google Scholar] [CrossRef]
- Al-Mohammadi, A.-R.; Osman, A.; Enan, G.; Abdel-Shafi, S.; El-Nemer, M.; Sitohy, M.; Taha, M.A. Powerful Antibacterial Peptides from Egg Albumin Hydrolysates. Antibiotics 2020, 9, 901. [Google Scholar] [CrossRef] [PubMed]
- Zanutto-Elgui, M.R.; Vieira, J.C.S.; do Prado, D.Z.; Buzalaf, M.A.R.; Padilha, P.d.M.; Elgui de Oliveira, D.; Fleuri, L.F. Production of Milk Peptides with Antimicrobial and Antioxidant Properties through Fungal Proteases. Food Chem. 2019, 278, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Fialho, T.L.; Carrijo, L.C.; Magalhães Júnior, M.J.; Baracat-Pereira, M.C.; Piccoli, R.H.; de Abreu, L.R. Extraction and Identification of Antimicrobial Peptides from the Canastra Artisanal Minas Cheese. Food Res. Int. 2018, 107, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Castellano, P.; Mora, L.; Escudero, E.; Vignolo, G.; Aznar, R.; Toldrá, F. Antilisterial Peptides from Spanish Dry-Cured Hams: Purification and Identification. Food Microbiol. 2016, 59, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Lima, C.A.; Campos, J.F.; Filho, J.L.L.; Converti, A.; da Cunha, M.G.C.; Porto, A.L.F. Antimicrobial and Radical Scavenging Properties of Bovine Collagen Hydrolysates Produced by Penicillium Aurantiogriseum URM 4622 Collagenase. J. Food Sci. Technol. 2015, 52, 4459–4466. [Google Scholar] [CrossRef] [Green Version]
- Osman, A.; Goda, H.A.; Abdel-Hamid, M.; Badran, S.M.; Otte, J. Antibacterial Peptides Generated by Alcalase Hydrolysis of Goat Whey. LWT 2016, 65, 480–486. [Google Scholar] [CrossRef]
- Wald, M.; Schwarz, K.; Rehbein, H.; Bußmann, B.; Beermann, C. Detection of Antibacterial Activity of an Enzymatic Hydrolysate Generated by Processing Rainbow Trout By-Products with Trout Pepsin. Food Chem. 2016, 205, 221–228. [Google Scholar] [CrossRef]
- Gaspar, D.; Veiga, A.S.; Castanho, M.A.R.B. From Antimicrobial to Anticancer Peptides. A Review. Front. Microbiol. 2013, 4, 294. [Google Scholar] [CrossRef] [Green Version]
- Hoskin, D.W.; Ramamoorthy, A. Studies on Anticancer Activities of Antimicrobial Peptides. Biochim. Biophys. Acta Biomembr. 2008, 1778, 357–375. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, P.; Glossop, H.; Meikle, T.G.; Aburto-Medina, A.; Conn, C.E.; Sarojini, V.; Valery, C. Molecular Engineering of Antimicrobial Peptides: Microbial Targets, Peptide Motifs and Translation Opportunities. Biophys. Rev. 2021, 13, 35–69. [Google Scholar] [CrossRef]
- Lee, E.Y.; Lee, M.W.; Wong, G.C.L. Modulation of Toll-like Receptor Signaling by Antimicrobial Peptides. Semin. Cell Dev. Biol. 2019, 88, 173–184. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Reddy, D.; Reddy, R.; Sharda, A.; Bose, K.; Gupta, S. Incorporation of a Tag Helps to Overcome Expression Variability in a Recombinant Host. Biotechnol. Rep. 2016, 11, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Andoh, Y.; Kitou, S.; Okazaki, S. Difference in Molecular Mechanisms Governing Changes in Membrane Properties of Phospholipid Bilayers Induced by Addition of Nonionic and Zwitterionic Surfactants. J. Mol. Liq. 2018, 271, 933–941. [Google Scholar] [CrossRef]
- Shai, Y. Mechanism of the Binding, Insertion and Destabilization of Phospholipid Bilayer Membranes by a-Helical Antimicrobial and Cell Non-Selective Membrane-Lytic Peptides. Biochim. Biophys. Acta Biomembr. 1999, 1462, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Liu, Q.; Wang, D.; Li, J. Characterization and Antimicrobial Mechanism of CF-14, a New Antimicrobial Peptide from the Epidermal Mucus of Catfish. Fish Shellfish. Immunol. 2019, 92, 881–888. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Hernández-Ledesma, B. Gastrointestinal Digestion of Food Proteins under the Effects of Released Bioactive Peptides on Digestive Health. Mol. Nutr. Food Res. 2020, 64, 2000401. [Google Scholar] [CrossRef]
- Liébana-García, R.; Olivares, M.; Bullich-Vilarrubias, C.; López-Almela, I.; Romaní-Pérez, M.; Sanz, Y. The Gut Microbiota as a Versatile Immunomodulator in Obesity and Associated Metabolic Disorders. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101542. [Google Scholar] [CrossRef]
- Meisel, H. Multifunctional Peptides Encrypted in Milk Proteins. BioFactors 2004, 21, 55–61. [Google Scholar] [CrossRef]
- Gauthier, S.F.; Pouliot, Y.; Saint-Sauveur, D. Immunomodulatory Peptides Obtained by the Enzymatic Hydrolysis of Whey Proteins. Int. Dairy J. 2006, 16, 1315–1323. [Google Scholar] [CrossRef]
- Chessa, C.; Bodet, C.; Jousselin, C.; Wehbe, M.; Lévêque, N.; Garcia, M. Antiviral and Immunomodulatory Properties of Antimicrobial Peptides Produced by Human Keratinocytes. Front. Microbiol. 2020, 11, 1155. [Google Scholar] [CrossRef] [PubMed]
- Théolier, J.; Hammami, R.; Labelle, P.; Fliss, I.; Jean, J. Isolation and Identification of Antimicrobial Peptides Derived by Peptic Cleavage of Whey Protein Isolate. J. Funct. Foods 2013, 5, 706–714. [Google Scholar] [CrossRef]
- Martín, R.; Langella, P. Emerging Health Concepts in the Probiotics Field: Streamlining the Definitions. Front. Microbiol. 2019, 10, 1047. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zeng, Y.; Wang, S.; Liu, H.; Zhang, D.; Zhang, W.; Wang, Y.; Ji, H. Swine-Derived Probiotic Lactobacillus Plantarum Inhibits Growth and Adhesion of Enterotoxigenic Escherichia Coli and Mediates Host Defense. Front. Microbiol. 2018, 9, 1364. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, S.; Abramov, A.Y. Mechanism of Oxidative Stress in Neurodegeneration. Oxid. Med. Cell. Longev. 2012, 2012, 428010. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The Role of Oxidative Stress in Neurodegenerative Diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Bevins, C.L.; Salzman, N.H. Paneth Cells, Antimicrobial Peptides and Maintenance of Intestinal Homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef]
- Zong, X.; Fu, J.; Xu, B.; Wang, Y.; Jin, M. Interplay between Gut Microbiota and Antimicrobial Peptides. Anim. Nutr. 2020, 6, 389–396. [Google Scholar] [CrossRef]
- Muniz, L.R.; Knosp, C.; Yeretssian, G. Intestinal Antimicrobial Peptides during Homeostasis, Infection, and Disease. Front. Immunol. 2012, 3, 310. [Google Scholar] [CrossRef] [Green Version]
- Cazorla, S.I.; Maldonado-Galdeano, C.; Weill, R.; De Paula, J.; Perdigón, G.D.V. Oral Administration of Probiotics Increases Paneth Cells and Intestinal Antimicrobial Activity. Front. Microbiol. 2018, 9, 736. [Google Scholar] [CrossRef]
- García-Díez, J.; Saraiva, C. Use of Starter Cultures in Foods from Animal Origin to Improve Their Safety. Int. J. Environ. Res. Public Health 2021, 18, 2544. [Google Scholar] [CrossRef]
- Kadyan, S.; Rashmi, H.M.; Pradhan, D.; Kumari, A.; Chaudhari, A.; Deshwal, G.K. Effect of Lactic Acid Bacteria and Yeast Fermentation on Antimicrobial, Antioxidative and Metabolomic Profile of Naturally Carbonated Probiotic Whey Drink. Lwt 2021, 142, 111059. [Google Scholar] [CrossRef]
- Giraffa, G.; Chanishvili, N.; Widyastuti, Y. Importance of Lactobacilli in Food and Feed Biotechnology. Res. Microbiol. 2010, 161, 480–487. [Google Scholar] [CrossRef]
- Raveschot, C.; Cudennec, B.; Coutte, F.; Flahaut, C.; Fremont, M.; Drider, D.; Dhulster, P. Production of Bioactive Peptides by Lactobacillus Species: From Gene to Application. Front. Microbiol. 2018, 9, 2354. [Google Scholar] [CrossRef] [Green Version]
- Amiri, S.; Aghamirzaei, M.; Mostashari, P.; Sarbazi, M.; Tizchang, S.; Madahi, H. The Impact of Biotechnology on Dairy Industry. In Microbial Biotechnology in Food and Health; Elsevier: Amsterdam, The Netherlands, 2021; pp. 53–79. [Google Scholar]
- Paul Ross, R.; Morgan, S.; Hill, C. Preservation and Fermentation: Past, Present and Future. Int. J. Food Microbiol. 2002, 79, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.M.; Jung, D.Y.-G.; Jin, D.Y.-Y.; Jayabalan, D.R.; Yang, D.S.H.; Suh, J.W. Bacteriocins as Food Preservatives: Challenges and Emerging Horizons. Crit. Rev. Food Sci. Nutr. 2017, 58, 2743–2767. [Google Scholar] [CrossRef]
- Acuña, L.; Corbalán, N.; Quintela-Baluja, M.; Barros-Velázquez, J.; Bellomio, A. Expression of the Hybrid Bacteriocin Ent35-MccV in Lactococcus Lactis and Its Use for Controlling Listeria Monocytogenes and Escherichia Coli in Milk. Int. Dairy J. 2020, 104, 104650. [Google Scholar] [CrossRef]
- Khelissa, S.; Chihib, N.-E.; Gharsallaoui, A. Conditions of Nisin Production by Lactococcus Lactis Subsp. Lactis and Its Main Uses as a Food Preservative. Arch. Microbiol. 2021, 203, 465–480. [Google Scholar] [CrossRef]
- Dal Bello, B.; Cocolin, L.; Zeppa, G.; Field, D.; Cotter, P.D.; Hill, C. Technological Characterization of Bacteriocin Producing Lactococcus Lactis Strains Employed to Control Listeria Monocytogenes in Cottage Cheese. Int. J. Food Microbiol. 2012, 153, 58–65. [Google Scholar] [CrossRef]
- Delorme, C. Safety Assessment of Dairy Microorganisms: Streptococcus Thermophilus. Int. J. Food Microbiol. 2008, 126, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Iyer, R.; Tomar, S.K.; Uma Maheswari, T.; Singh, R. Streptococcus Thermophilus Strains: Multifunctional Lactic Acid Bacteria. Int. Dairy J. 2010, 20, 133–141. [Google Scholar] [CrossRef]
- Renye, J.A.; Somkuti, G.A.; Garabal, J.I.; Steinberg, D.H. Bacteriocin Production by Streptococcus Thermophilus in Complex Growth Media. Biotechnol. Lett. 2016, 38, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.; Wang, Y.; Cheng, Q.; Imran, M. Lactobacillus Acidophilus Bacteriocin, from Production to Their Application: An Overview. African J. Biotechnol. 2010, 9, 2843–2850. [Google Scholar]
- Uzuner, A.E.; Kinik, O.; Korel, F.; Yildiz, G.; Yerlikaya, O. Usage of Rice Milk in Probiotic Yoghurt Production. Carpathian J. Food Sci. Technol. 2016, 8, 5–25. [Google Scholar]
- Hati, S.; Patel, N.; Sakure, A.; Mandal, S. Influence of Whey Protein Concentrate on the Production of Antibacterial Peptides Derived from Fermented Milk by Lactic Acid Bacteria. Int. J. Pept. Res. Ther. 2018, 24, 87–98. [Google Scholar] [CrossRef]
- Atanasova, J.; Moncheva, P.; Ivanova, I. Proteolytic and Antimicrobial Activity of Lactic Acid Bacteria Grown in Goat Milk. Biotechnol. Biotechnol. Equip. 2014, 28, 1073–1078. [Google Scholar] [CrossRef] [Green Version]
- Muhialdin, B.J.; Algboory, H.L. Identification of Low Molecular Weight Antimicrobial Peptides from Iraqi Camel Milk Fermented with Lactobacillus Plantarum. PharmaNutrition 2018, 6, 69–73. [Google Scholar] [CrossRef]
- Tenea, G.N.; Olmedo, D.; Ortega, C. Peptide-Based Formulation from Lactic Acid Bacteria Impairs the Pathogen Growth in Ananas Comosus (Pineapple). Coatings 2020, 10, 457. [Google Scholar] [CrossRef]
- Gupta, R.; Srivastava, S. Antifungal Effect of Antimicrobial Peptides (AMPs LR14) Derived from Lactobacillus Plantarum Strain LR/14 and Their Applications in Prevention of Grain Spoilage. Food Microbiol. 2014, 42, 1–7. [Google Scholar] [CrossRef]
- Weerkamp, A.H.; Klijn, N.; Neeter, R.; Smit, G. Properties of Mesophilic Lactic Acid Bacteria from Raw Milk and Naturally Fermented Raw Milk Products. Netherlands Milk Dairy J. 1996, 50, 319–332. [Google Scholar]
- Slattery, L.; O’Callaghan, J.; Fitzgerald, G.F.; Beresford, T.; Ross, R.P. Invited Review: Lactobacillus Helveticus—A Thermophilic Dairy Starter Related to Gut Bacteria. J. Dairy Sci. 2010, 93, 4435–4454. [Google Scholar] [CrossRef] [Green Version]
- Papathanasopoulos, M.A.; Krier, F.; Revol-Junelles, A.-M.; Lefebvre, G.; Le Caer, J.P.; von Holy, A.; Hastings, J.W. Multiple Bacteriocin Production by Leuconostoc MesenteroidesTA33a and Other Leuconostoc/Weissella Strains. Curr. Microbiol. 1997, 35, 331–335. [Google Scholar] [CrossRef]
- Kingcha, Y.; Tosukhowong, A.; Zendo, T.; Roytrakul, S.; Luxananil, P.; Chareonpornsook, K.; Valyasevi, R.; Sonomoto, K.; Visessanguan, W. Anti-Listeria Activity of Pediococcus Pentosaceus BCC 3772 and Application as Starter Culture for Nham, a Traditional Fermented Pork Sausage. Food Control 2012, 25, 190–196. [Google Scholar] [CrossRef]
- Vera Pingitore, E.; Todorov, S.D.; Sesma, F.; Gombossy de Melo Franco, B.D. Application of Bacteriocinogenic Enterococcus Mundtii CRL35 and Enterococcus Faecium ST88Ch in the Control of Listeria Monocytogenes in Fresh Minas Cheese. Food Microbiol. 2012, 32, 38–47. [Google Scholar] [CrossRef]
- Pinto, A.; Barbosa, J.; Albano, H.; Isidro, J.; Teixeira, P. Screening of Bacteriocinogenic Lactic Acid Bacteria and Their Characterization as Potential Probiotics. Microorganisms 2020, 8, 393. [Google Scholar] [CrossRef] [Green Version]
- Siroli, L.; Patrignani, F.; Serrazanetti, D.I.; Tabanelli, G.; Montanari, C.; Gardini, F.; Lanciotti, R. Lactic Acid Bacteria and Natural Antimicrobials to Improve the Safety and Shelf-Life of Minimally Processed Sliced Apples and Lamb’s Lettuce. Food Microbiol. 2015, 47, 74–84. [Google Scholar] [CrossRef]
- Lafta, H.; Jarallah, E.M.; Darwash, A. Antibacterial Activity of Fermented Camel Milk Using Two Lactic Acid Bacteria. J. Babylon Univ. Pure Appl. Sci. 2014, 22, 2377–2382. [Google Scholar]
- Mal, G.; Sena, D.S.; Jain, V.K.; Sahani, M.S. Therapeutic Value of Camel Milk as a Nutritional Supplement for Multiple Drug Resistant (MDR) Tuberculosis Patients. Isr. J. Vet. Med. 2006, 61, 88–91. [Google Scholar]
- Soleymanzadeh, N.; Mirdamadi, S.; Kianirad, M. Antioxidant Activity of Camel and Bovine Milk Fermented by Lactic Acid Bacteria Isolated from Traditional Fermented Camel Milk (Chal). Dairy Sci. Technol. 2016, 96, 443–457. [Google Scholar] [CrossRef] [Green Version]
- Ayyash, M.; Al-Nuaimi, A.K.; Al-Mahadin, S.; Liu, S.-Q. In Vitro Investigation of Anticancer and ACE-Inhibiting Activity, α-Amylase and α-Glucosidase Inhibition, and Antioxidant Activity of Camel Milk Fermented with Camel Milk Probiotic: A Comparative Study with Fermented Bovine Milk. Food Chem. 2018, 239, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Ejtahed, H.S.; Niasari Naslaji, A.; Mirmiran, P.; Zraif Yeganeh, M.; Hedayati, M.; Azizi, F.; Moosavi Movahedi, A. Effect of Camel Milk on Blood Sugar and Lipid Profile of Patients With Type 2 Diabetes: A Pilot Clinical Trial. Int. J. Endocrinol. Metab. 2014, 13, e21160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benkerroum, N.; Mekkaoui, M.; Bennani, N.; Hidane, K. Antimicrobial Activity of Camel’s Milk against Pathogenic Strains of Escherichia Coli and Listeria Monocytogenes. Int. J. Dairy Technol. 2004, 57, 39–43. [Google Scholar] [CrossRef]
- Abdel-Hamid, M.; Romeih, E.; Saporito, P.; Osman, A.; Mateiu, R.V.; Mojsoska, B.; Jenssen, H. Camel Milk Whey Hydrolysate Inhibits Growth and Biofilm Formation of Pseudomonas Aeruginosa PAO1 and Methicillin-Resistant Staphylococcus Aureus. Food Control 2020, 111, 107056. [Google Scholar] [CrossRef]
- Izquierdo-González, J.J.; Amil-Ruiz, F.; Zazzu, S.; Sánchez-Lucas, R.; Fuentes-Almagro, C.A.; Rodríguez-Ortega, M.J. Proteomic Analysis of Goat Milk Kefir: Profiling the Fermentation-Time Dependent Protein Digestion and Identification of Potential Peptides with Biological Activity. Food Chem. 2019, 295, 456–465. [Google Scholar] [CrossRef]
- Hayes, M.; Ross, R.P.; Fitzgerald, G.F.; Hill, C.; Stanton, C. Casein-Derived Antimicrobial Peptides Generated by Lactobacillus Acidophilus DPC6026. Appl. Environ. Microbiol. 2006, 72, 2260–2264. [Google Scholar] [CrossRef] [Green Version]
- Villegas, J.M.; Picariello, G.; Mamone, G.; Espeche Turbay, M.B.; Savoy de Giori, G.; Hebert, E.M. Milk-Derived Angiotensin-I-Converting Enzymeinhibitory Peptides Generated by Lactobacillus Delbrueckii Subsp. Lactis CRL 581. Peptidomics 2014, 1, 22–29. [Google Scholar] [CrossRef]
- Agyei, D.; Danquah, M.K. Industrial-Scale Manufacturing of Pharmaceutical-Grade Bioactive Peptides. Biotechnol. Adv. 2011, 29, 272–277. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Peñas, E.; Frias, J. Bioactive Peptides in Fermented Foods. In Fermented Foods in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2017; pp. 23–47. [Google Scholar]
- Chalamaiah, M.; Keskin Ulug, S.; Hong, H.; Wu, J. Regulatory Requirements of Bioactive Peptides (Protein Hydrolysates) from Food Proteins. J. Funct. Foods 2019, 58, 123–129. [Google Scholar] [CrossRef]
- Witkowska, A.M.; Hickey, D.K.; Alonso-Gomez, M.; Wilkinson, M.G. The Microbiological Quality of Commercial Herb and Spice Preparations Used in the Formulation of a Chicken Supreme Ready Meal and Microbial Survival Following a Simulated Industrial Heating Process. Food Control 2011, 22, 616–625. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, F.; Carrasco, E.; Valero, A. Impact of Dehydration and Drying Operations on the Microbial Ecology of Foods. In Quantitative Microbiology in Food Processing; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 160–175. [Google Scholar]
- Udenigwe, C.C.; Aluko, R.E. Food Protein-Derived Bioactive Peptides: Production, Processing, and Potential Health Benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef]
- Martín, M.-H.J.; Ángel, M.-M.M.; Aarón, S.-L.J.; Israel, B.-G. Protein Hydrolysates as Biostimulants of Plant Growth and Development. In Biostimulants: Exploring Sources and Applications; Springer: Singapore, 2022; pp. 141–175. [Google Scholar]
- Zhao, X.; Zhang, X.; Liu, D. Collagen Peptides and the Related Synthetic Peptides: A Review on Improving Skin Health. J. Funct. Foods 2021, 86, 104680. [Google Scholar] [CrossRef]
- Cai, G.; Moffitt, K.; Navone, L.; Zhang, Z.; Robins, K.; Speight, R. Valorisation of Keratin Waste: Controlled Pretreatment Enhances Enzymatic Production of Antioxidant Peptides. J. Environ. Manag. 2022, 301, 113945. [Google Scholar] [CrossRef]
- Ashaolu, T.J. Nanoemulsions for Health, Food, and Cosmetics: A Review. Environ. Chem. Lett. 2021, 19, 3381–3395. [Google Scholar] [CrossRef]
- Silveira, R.F.; Roque-Borda, C.A.; Vicente, E.F. Antimicrobial Peptides as a Feed Additive Alternative to Animal Production, Food Safety and Public Health Implications: An Overview. Anim. Nutr. 2021, 7, 896–904. [Google Scholar] [CrossRef]
- Nazeer, N.; Uribe-Diaz, S.; Rodriguez-Lecompte, J.C.; Ahmed, M. Antimicrobial Peptides as an Alternative to Relieve Antimicrobial Growth Promoters in Poultry. Br. Poult. Sci. 2021, 62, 672–685. [Google Scholar] [CrossRef]
- Deng, Z.; Yu, H.; Yang, Z.; Hu, L.; Liu, Q.; Wang, Y.; Wei, H.-K.; Peng, J. Gly-Pro-Ala Peptide and FGSHF3 Exert Protective Effects in DON-Induced Toxicity and Intestinal Damage via Decreasing Oxidative Stress. Food Res. Int. 2021, 139, 109840. [Google Scholar] [CrossRef]
- Taroncher, M.; Rodríguez-Carrasco, Y.; Aspevik, T.; Kousoulaki, K.; Barba, F.J.; Ruiz, M.-J. Cytoprotective Effects of Fish Protein Hydrolysates against H2O2-Induced Oxidative Stress and Mycotoxins in Caco-2/TC7 Cells. Antioxidants 2021, 10, 975. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium Toxins in Cereals: Occurrence, Legislation, Factors Promoting the Appearance and Their Management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.N.; Tabassum, M.; Banik, M.; Daayf, F.; Fernando, W.G.D.; Harris, L.J.; Sura, S.; Wang, X. Naturally Occurring Fusarium Species and Mycotoxins in Oat Grains from Manitoba, Canada. Toxins 2021, 13, 670. [Google Scholar] [CrossRef]
- Tkaczewska, J. Peptides and Protein Hydrolysates as Food Preservatives and Bioactive Components of Edible Films and Coatings—A Review. Trends Food Sci. Technol. 2020, 106, 298–311. [Google Scholar] [CrossRef]


{kind=link}
| BAP/Hydrolysate | Treatment/Evaluation Method | Observed Effects | Reference |
|---|---|---|---|
| Enzymatic hydrolyzed cottonseed meal | In vivo evaluation of supplementation in feed for broiler chicks (15 or 20 g/kg) | Decreased population of Escherichia coli and increased Lactobacilli counts in ileum; positive effects on zootechnical parameters. | [21] |
| Enzymatic hydrolyzed cottonseed meal | Challenge in vitro test with Colletotrichum gloeosporioides, E. coli O157:H7, and Staphylococcus aureus. | Inhibition of C. gloeosporioides and S. aureus growth; no inhibition on E. coli was observed. | [81] |
| Whey protein hydrolysate | Challenge test with Listeria monocytogenes; evaluation in soft cheese-based agar (5 μg/plate), combined or not with LAB strains. | In combination with LAB strains, reduction of L. monocytogenes was achieved, although no inhibition was observed when alone. | [82] |
| Anionic peptide-enriched extract derived from whey proteins | Challenge test with Listeria innocua and L. monocytogenes; application in reconstituted Cheddar cheese (10 or 20 mg/g) incorporated with lactococci. | Higher anti-listerial activity under higher temperatures and/or low salt content; L. monocytogenes more susceptible than L. innocua; no inhibition on LAB. | [83] |
| Encrypted peptides recovered from soybean meal by-product | In vitro evaluation of aqueous extract fractions towards Gram-positive and -negative pathogens; in silico prediction of the antimicrobial sequences. | Inhibition of S. aureus, Acinetobacter genomospecies, Aeromonas hydrophila, E. coli, Salmonella enterica, and Vibrio parahaemolyticus; 83 peptide sequences classified as AMP candidates. | [70] |
| Bovine lactoferrin-derived AMPs | Short (~12 residues-long) AMPs derived from the protein were designed and evaluated towards Enterococcus faecium in vivo and ex vivo. | Designed AMPs showed high antimicrobial activity on free cells and biofilm, low mammalian cytotoxicity, and membrane-activating mechanisms. | [84] |
| Hen egg-white lysozyme-derived AMPs (enzymatic hydrolysis) | Evaluation through a radial diffusion assay. | Antibacterial activity against Leuconostoc mesenteroides and E. coli, the latter showing greater susceptibility. | [85] |
| Egg albumin hydrolysates (enzymatic hydrolysis) | Evaluation of the antimicrobial activity of hydrolysates through the disc diffusion and tube dilution method. | Antibacterial activity against L. monocytogenes, Bacillus cereus, S. aureus, Salmonella Typhimurium, Streptococcus pyogenes, Klebsiella oxytoca, Pseudomonas aeruginosa, Bacillus subtilis, Listeria ivanovii, and E. coli. | [86] |
| Goat- and bovine-milk-derived BAPs (enzymatic hydrolysis) | After fungal proteolysis, evaluation through disk diffusion towards bacterial and fungal microorganisms. | Antimicrobial activity towards L. monocytogenes, S. aureus, E. coli, S. enterica, P. aeruginosa, Fusarium oxysporum, Penicillium expansum, and Candida albicans; no inhibition against Aspergillus fumigatus was observed. | [87] |
| Water-soluble AMPs recovered from the ripened Brazilian Canastra artisanal Minas cheese | Evaluation of the promoted inhibition on E. coli, comparing different ripening stages and cheese producers; identification of peptide sequences. | Observed variations influenced by temperature, pH, and other manufacturing characteristics; identification of six validated AMPs, 8–14 residues long, derived from caseins. | [88] |
| Peptide-rich fractions extracted from Spanish dry-cured ham | 128 fractions chromatographically purified were evaluated through agar-well-diffusion assay for the inhibition of L. monocytogenes and L. innocua; peptidomic study on the naturally generated BAPs. | Two fractions showed inhibitory effects towards Listeria strains; identification of 105 BAPs in the two bioactive fractions, 10 with anti-listerial activity. | [89] |
| Bovine collagen hydrolysates (enzymatic hydrolysis) | Antimicrobial activity was evaluated for the hydrolysates (0.5 to 5 mg/mL); peptide profiling of hydrolysates. | Hydrolysates showed inhibitory activity towards E. coli, S. aureus, and B. subtilis; no inhibition was achieved against E. faecalis, P. aeruginosa, or Klebsiella pneumoniae. Identification of several peptides with low molar mass (<2 kDa). | [90] |
| Goat-whey hydrolysates (enzymatic hydrolysis) | Evaluation of the antimicrobial activity of hydrolysates through disc diffusion method; peptide profiling of fractions. | The hydrolysate showed bactericidal effects towards B. cereus, Salmonella Typhimurium, and E. coli; and bacteriostatic activity against S. aureus. Two peptides accounted for the bioactivity. | [91] |
| Rainbow trout by-product hydrolysates (enzymatic hydrolysis) | Assessment of inhibitory activity against several bacterial strains. | Inhibitory activity was detected towards all tested strains, with the highest activity against Flavobacterium species; prolonged lag phase of bacterial growth. | [92] |
| Genus/Species | Studied Food Product | Mechanism of AMP Production | References |
|---|---|---|---|
| Lactococcus | |||
| Lactococcus lactis lactis | Skim milk | Ribosomal synthesis of bacteriocins | [123,124] |
| Lactococcus lactis lactis | Cottage cheese | Protein hydrolysis, releasing BAPs | [125] |
| Streptococcus | |||
| Streptococcus thermophilus | Milk, yogurt, soft and hard cheeses | Ribosomal synthesis of bacteriocins | [126,127,128] |
| Lactobacillus | |||
| Lactobacillus acidophilus | Cheese, yogurt | Protein hydrolysis, releasing free amino acids and BAPs | [119,129] |
| Lactobacillus gasseri | Yogurt | Protein hydrolysis, releasing free amino acids and BAPs | [119,130] |
| Lactobacillus helveticus | Skim milk supplemented with whey protein | Protein hydrolysis, releasing free BAPs | [131] |
| Lactobacillus delbrueckii bulgaricus | Skimmed goat milk | Protein hydrolysis, releasing free amino acids and BAPs | [132] |
| Lactiplantibacillus | |||
| Lactiplantibacillus plantarum | Fermented camel milk | Protein hydrolysis, releasing free amino acids and BAPs | [119,133] |
| Lactiplantibacillus plantarum | Pineapple | Protein hydrolysis, releasing free amino acids and BAPs | [134] |
| Lactiplantibacillus plantarum | Wheat grain | Protein hydrolysis, releasing free amino acids and BAPs | [135] |
| Leuconostoc | |||
| Leuconostoc mesenteroides cremoris | Cheese, butter, heavy cream | Ribosomal synthesis of bacteriocins | [136,137,138] |
| Pediococcus | |||
| Pediococcus pentosaceus | Fermented pork sausage | Production of pediocin PA-1/AcH by protein hydrolysis | [139] |
| Enterococcus | |||
| Enterococcus faecium | Minas cheese | Ribosomal synthesis of bacteriocins | [140] |
| Enterococcus mundtii | Minas cheese | Ribosomal synthesis of bacteriocins | [140] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corrêa, J.A.F.; de Melo Nazareth, T.; Rocha, G.F.d.; Luciano, F.B. Bioactive Antimicrobial Peptides from Food Proteins: Perspectives and Challenges for Controlling Foodborne Pathogens. Pathogens 2023, 12, 477. https://doi.org/10.3390/pathogens12030477
Corrêa JAF, de Melo Nazareth T, Rocha GFd, Luciano FB. Bioactive Antimicrobial Peptides from Food Proteins: Perspectives and Challenges for Controlling Foodborne Pathogens. Pathogens. 2023; 12(3):477. https://doi.org/10.3390/pathogens12030477
Chicago/Turabian StyleCorrêa, Jessica Audrey Feijó, Tiago de Melo Nazareth, Giovanna Fernandes da Rocha, and Fernando Bittencourt Luciano. 2023. "Bioactive Antimicrobial Peptides from Food Proteins: Perspectives and Challenges for Controlling Foodborne Pathogens" Pathogens 12, no. 3: 477. https://doi.org/10.3390/pathogens12030477