
Preventing Persistence of HPV Infection with Natural Molecules

 ,
,  , and
, and 
{kind=link}
Abstract
:1. Introduction
2. Methods
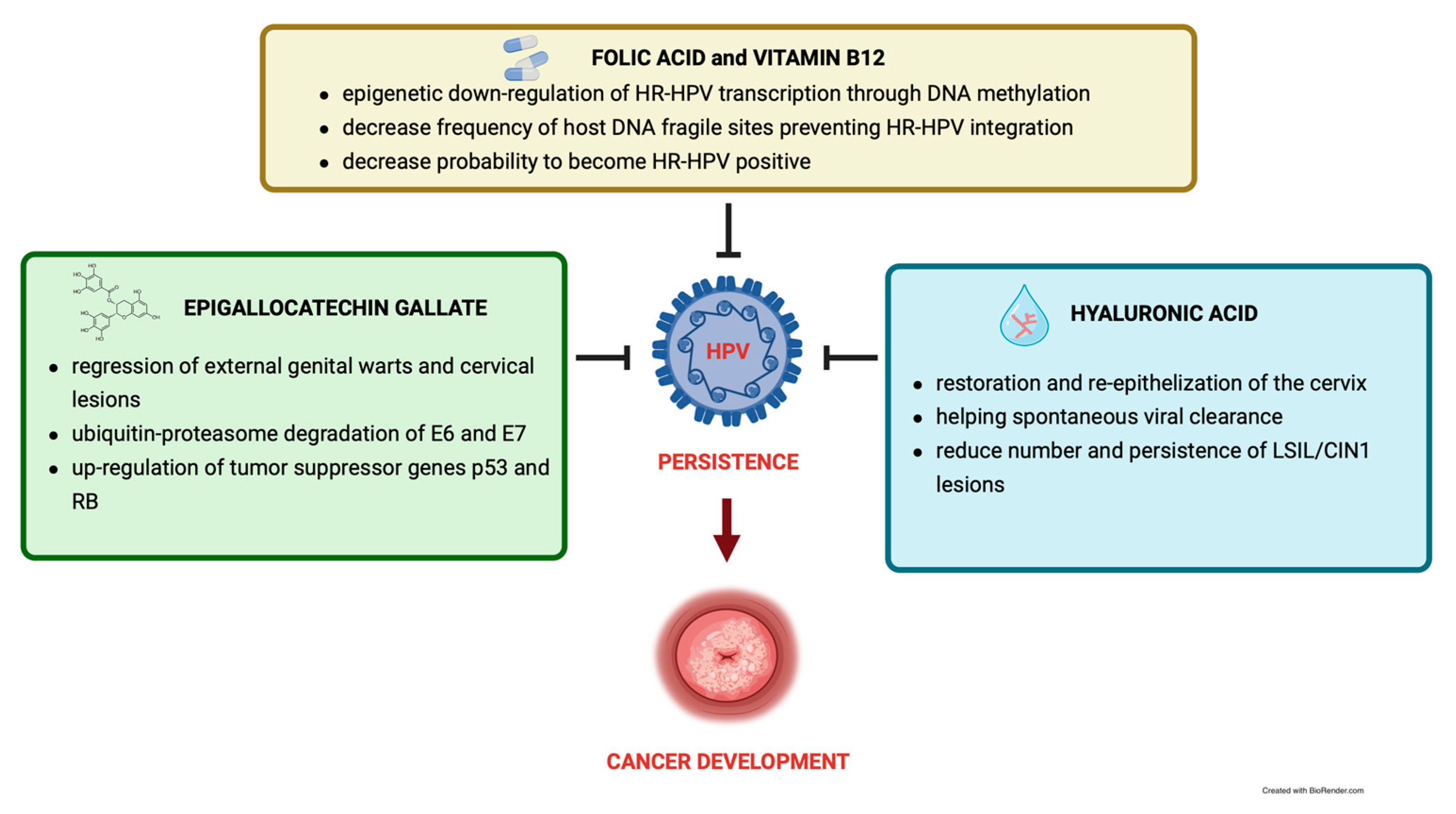
3. Epigallocatechin Gallate
4. Folic Acid and Vitamin B12
5. Hyaluronic Acid
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kombe Kombe, A.J.; Li, B.; Zahid, A.; Mengist, H.M.; Bounda, G.-A.; Zhou, Y.; Jin, T. Epidemiology and Burden of Human Papillomavirus and Related Diseases, Molecular Pathogenesis, and Vaccine Evaluation. Front. Public Health 2020, 8, 552028. [Google Scholar] [CrossRef] [PubMed]
- Chesson, H.W.; Dunne, E.F.; Hariri, S.; Markowitz, L.E. The Estimated Lifetime Probability of Acquiring Human Papillomavirus in the United States. Sex. Transm. Dis. 2014, 41, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Egawa, N.; Egawa, K.; Griffin, H.; Doorbar, J. Human Papillomaviruses; Epithelial Tropisms, and the Development of Neoplasia. Viruses 2015, 7, 3863–3890. [Google Scholar] [CrossRef] [Green Version]
- Van Doorslaer, K. Revisiting Papillomavirus Taxonomy: A Proposal for Updating the Current Classification in Line with Evolutionary Evidence. Viruses 2022, 14, 2308. [Google Scholar] [CrossRef]
- Garutti, P.; Montori, S.; Bazzan, E.; Tarabbia, C. Gender differences in the epidemiology and prevention of human papillomavirus (HPV) and HPV-related diseases. Ital. J. Gend.-Specif. Med. 2018, 4, 152–161. [Google Scholar]
- de Martel, C.; Plummer, M.; Vignat, J.; Franceschi, S. Worldwide Burden of Cancer Attributable to HPV by Site, Country and HPV Type. Int. J. Cancer 2017, 141, 664–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sanjosé, S.; Serrano, B.; Castellsagué, X.; Brotons, M.; Muñoz, J.; Bruni, L.; Bosch, F.X. Human Papillomavirus (HPV) and Related Cancers in the Global Alliance for Vaccines and Immunization (GAVI) Countries. A WHO/ICO HPV Information Centre Report. Vaccine 2012, 30 (Suppl. S4), D1–D83. [Google Scholar] [CrossRef]
- Cogliano, V.; Baan, R.; Straif, K.; Grosse, Y.; Secretan, B.; El Ghissassi, F. WHO International Agency for Research on Cancer Carcinogenicity of Human Papillomaviruses. Lancet Oncol. 2005, 6, 204. [Google Scholar] [CrossRef]
- Giuliano, A.R.; Nyitray, A.G.; Kreimer, A.R.; Pierce Campbell, C.M.; Goodman, M.T.; Sudenga, S.L.; Monsonego, J.; Franceschi, S. EUROGIN 2014 Roadmap: Differences in Human Papillomavirus Infection Natural History, Transmission and Human Papillomavirus-Related Cancer Incidence by Gender and Anatomic Site of Infection. Int. J. Cancer 2015, 136, 2752–2760. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, N.; Bosch, F.X.; de Sanjosé, S.; Herrero, R.; Castellsagué, X.; Shah, K.V.; Snijders, P.J.F.; Meijer, C.J.L.M.; International Agency for Research on Cancer Multicenter Cervical Cancer Study Group. Epidemiologic Classification of Human Papillomavirus Types Associated with Cervical Cancer. N. Engl. J. Med. 2003, 348, 518–527. [Google Scholar] [CrossRef] [Green Version]
- Parkin, D.M.; Bray, F. Chapter 2: The Burden of HPV-Related Cancers. Vaccine 2006, 24 (Suppl. S3), S11–S25. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Guideline for Screening and Treatment of Cervical Pre-Cancer Lesions for Cervical Cancer Prevention. In WHO Guidelines Approved by the Guidelines Review Committee, 2nd ed.; World Health Organization: Geneva, Switzerland, 2021; ISBN 978-92-4-003082-4. [Google Scholar]
- Ding, T.; Li, L.; Duan, R.; Chen, Y.; Yang, B.; Xi, M. Risk Factors Analysis of Recurrent Disease after Treatment with a Loop Electrosurgical Excision Procedure for High-Grade Cervical Intraepithelial Neoplasia. Int. J. Gynaecol. Obstet. 2023, 160, 538–547. [Google Scholar] [CrossRef] [PubMed]
- de Sanjosé, S.; Brotons, M.; Pavón, M.A. The Natural History of Human Papillomavirus Infection. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 47, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Gravitt, P.E.; Winer, R.L. Natural History of HPV Infection across the Lifespan: Role of Viral Latency. Viruses 2017, 9, 267. [Google Scholar] [CrossRef]
- Koshiol, J.; Lindsay, L.; Pimenta, J.M.; Poole, C.; Jenkins, D.; Smith, J.S. Persistent Human Papillomavirus Infection and Cervical Neoplasia: A Systematic Review and Meta-Analysis. Am. J. Epidemiol. 2008, 168, 123–137. [Google Scholar] [CrossRef]
- Gillison, M.L.; Koch, W.M.; Capone, R.B.; Spafford, M.; Westra, W.H.; Wu, L.; Zahurak, M.L.; Daniel, R.W.; Viglione, M.; Symer, D.E.; et al. Evidence for a Causal Association between Human Papillomavirus and a Subset of Head and Neck Cancers. J. Natl. Cancer Inst. 2000, 92, 709–720. [Google Scholar] [CrossRef] [Green Version]
- Haukioja, A.; Asunta, M.; Söderling, E.; Syrjänen, S. Persistent Oral Human Papillomavirus Infection Is Associated with Smoking and Elevated Salivary Immunoglobulin G Concentration. J. Clin. Virol. 2014, 61, 101–106. [Google Scholar] [CrossRef]
- Rositch, A.F.; Koshiol, J.; Hudgens, M.G.; Razzaghi, H.; Backes, D.M.; Pimenta, J.M.; Franco, E.L.; Poole, C.; Smith, J.S. Patterns of Persistent Genital Human Papillomavirus Infection among Women Worldwide: A Literature Review and Meta-Analysis. Int. J. Cancer 2013, 133, 1271–1285. [Google Scholar] [CrossRef] [Green Version]
- Xi, L.F.; Koutsky, L.A.; Castle, P.E.; Edelstein, Z.R.; Meyers, C.; Ho, J.; Schiffman, M. Relationship between Cigarette Smoking and Human Papilloma Virus Types 16 and 18 DNA Load. Cancer Epidemiol. Biomark. Prev. 2009, 18, 3490–3496. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.Y.; Kim, M.K.; Seo, S.; Lee, D.O.; Chung, Y.K.; Lim, M.C.; Kim, J.; Lee, C.W.; Park, S. Alcohol Consumption and Persistent Infection of High-Risk Human Papillomavirus. Epidemiol. Infect. 2015, 143, 1442–1450. [Google Scholar] [CrossRef]
- Rousseau, M.C.; Pereira, J.S.; Prado, J.C.; Villa, L.L.; Rohan, T.E.; Franco, E.L. Cervical Coinfection with Human Papillomavirus (HPV) Types as a Predictor of Acquisition and Persistence of HPV Infection. J. Infect. Dis. 2001, 184, 1508–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radley, D.; Saah, A.; Stanley, M. Persistent Infection with Human Papillomavirus 16 or 18 Is Strongly Linked with High-Grade Cervical Disease. Hum. Vaccin. Immunother. 2016, 12, 768–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, G.; Kreimer, A.R.; Viscidi, R.; Pawlita, M.; Fakhry, C.; Koch, W.M.; Westra, W.H.; Gillison, M.L. Case-Control Study of Human Papillomavirus and Oropharyngeal Cancer. N. Engl. J. Med. 2007, 356, 1944–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akagi, K.; Li, J.; Broutian, T.R.; Padilla-Nash, H.; Xiao, W.; Jiang, B.; Rocco, J.W.; Teknos, T.N.; Kumar, B.; Wangsa, D.; et al. Genome-Wide Analysis of HPV Integration in Human Cancers Reveals Recurrent, Focal Genomic Instability. Genome Res. 2014, 24, 185–199. [Google Scholar] [CrossRef] [Green Version]
- Travé, G.; Zanier, K. HPV-Mediated Inactivation of Tumor Suppressor P53. Cell Cycle 2016, 15, 2231–2232. [Google Scholar] [CrossRef] [Green Version]
- Münger, K.; Scheffner, M.; Huibregtse, J.M.; Howley, P.M. Interactions of HPV E6 and E7 Oncoproteins with Tumour Suppressor Gene Products. Cancer Surv. 1992, 12, 197–217. [Google Scholar]
- Stanley, M. Immune Responses to Human Papillomavirus. Vaccine 2006, 24 (Suppl. S1), S16–S22. [Google Scholar] [CrossRef]
- Westrich, J.A.; Warren, C.J.; Pyeon, D. Evasion of Host Immune Defenses by Human Papillomavirus. Virus Res. 2017, 231, 21–33. [Google Scholar] [CrossRef]
- Kristiansen, E.; Jenkins, A.; Holm, R. Coexistence of Episomal and Integrated HPV16 DNA in Squamous Cell Carcinoma of the Cervix. J. Clin. Pathol. 1994, 47, 253–256. [Google Scholar] [CrossRef] [Green Version]
- McBride, A.A.; Warburton, A. The Role of Integration in Oncogenic Progression of HPV-Associated Cancers. PLoS Pathog. 2017, 13, e1006211. [Google Scholar] [CrossRef] [Green Version]
- Alfaro, K.; Maza, M.; Cremer, M.; Masch, R.; Soler, M. Removing Global Barriers to Cervical Cancer Prevention and Moving towards Elimination. Nat. Rev. Cancer 2021, 21, 607–608. [Google Scholar] [CrossRef]
- Baethge, C.; Goldbeck-Wood, S.; Mertens, S. SANRA-a Scale for the Quality Assessment of Narrative Review Articles. Res. Integr. Peer Rev. 2019, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-Q.; Lu, J.-L.; Liang, Y.-R.; Li, Q.-S. Suppressive Effects of EGCG on Cervical Cancer. Molecules 2018, 23, 2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Major Constituent of Green Tea, Epigallocatechin-3-Gallate (EGCG), Inhibits the Growth of HPV18-Infected Keratinocytes by Stimulating Proteasomal Turnover of the E6 and E7 Oncoproteins—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/33920477/ (accessed on 5 February 2023).
- Qiao, Y.; Cao, J.; Xie, L.; Shi, X. Cell Growth Inhibition and Gene Expression Regulation by (-)-Epigallocatechin-3-Gallate in Human Cervical Cancer Cells. Arch. Pharm. Res. 2009, 32, 1309–1315. [Google Scholar] [CrossRef]
- Zou, C.; Liu, H.; Feugang, J.M.; Hao, Z.; Chow, H.-H.S.; Garcia, F. Green Tea Compound in Chemoprevention of Cervical Cancer. Int. J. Gynecol. Cancer 2010, 20, 617–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Tyagi, S.; Bhui, K.; Prasad, S.; Shukla, Y. Regulation of Cell Growth through Cell Cycle Arrest and Apoptosis in HPV 16 Positive Human Cervical Cancer Cells by Tea Polyphenols. Investig. New Drugs 2010, 28, 216–224. [Google Scholar] [CrossRef]
- Mantovani, F.; Banks, L. The Human Papillomavirus E6 Protein and Its Contribution to Malignant Progression. Oncogene 2001, 20, 7874–7887. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, M.; Noguchi, M.; Nakao, Y.; Pater, A.; Iwasaka, T. The Tea Polyphenol, (-)-Epigallocatechin Gallate Effects on Growth, Apoptosis, and Telomerase Activity in Cervical Cell Lines. Gynecol. Oncol. 2004, 92, 197–204. [Google Scholar] [CrossRef]
- Jacob, J.; Cabarcas, S.; Veras, I.; Zaveri, N.; Schramm, L. The Green Tea Component EGCG Inhibits RNA Polymerase III Transcription. Biochem. Biophys. Res. Commun. 2007, 360, 778–783. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarty, S.; Ganguli, A.; Das, A.; Nag, D.; Chakrabarti, G. Epigallocatechin-3-Gallate Shows Anti-Proliferative Activity in HeLa Cells Targeting Tubulin-Microtubule Equilibrium. Chem. Biol. Interact. 2015, 242, 380–389. [Google Scholar] [CrossRef]
- Jordan, M.A.; Wilson, L. Microtubules as a Target for Anticancer Drugs. Nat. Rev. Cancer 2004, 4, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Nusri, Q.E.-A.; Begum, S.; Javed, E.; Rizvi, T.A.; Hussain, A. (-)-Epigallocatechin-3-Gallate Induces Apoptosis and Inhibits Invasion and Migration of Human Cervical Cancer Cells. Asian Pac. J. Cancer Prev. 2012, 13, 4815–4822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roomi, M.W.; Monterrey, J.C.; Kalinovsky, T.; Rath, M.; Niedzwiecki, A. In Vitro Modulation of MMP-2 and MMP-9 in Human Cervical and Ovarian Cancer Cell Lines by Cytokines, Inducers and Inhibitors. Oncol. Rep. 2010, 23, 605–614. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.-P.; LeCouter, J. The Biology of VEGF and Its Receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Singh, R.; Bhui, K.; Tyagi, S.; Mahmood, Z.; Shukla, Y. Tea Polyphenols Induce Apoptosis through Mitochondrial Pathway and by Inhibiting Nuclear Factor-KappaB and Akt Activation in Human Cervical Cancer Cells. Oncol. Res. 2011, 19, 245–257. [Google Scholar] [CrossRef]
- Al-Hazzani, A.A.; Alshatwi, A.A. Catechin Hydrate Inhibits Proliferation and Mediates Apoptosis of SiHa Human Cervical Cancer Cells. Food Chem. Toxicol. 2011, 49, 3281–3286. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, N.-D.; Zhou, F.; Shen, T.; Duan, T.; Zhou, J.; Shi, Y.; Zhu, X.-Q.; Shen, H.-M. (-)-Epigallocatechin-3-Gallate Induces Non-Apoptotic Cell Death in Human Cancer Cells via ROS-Mediated Lysosomal Membrane Permeabilization. PLoS ONE 2012, 7, e46749. [Google Scholar] [CrossRef]
- Song, J.Y.; Han, J.H.; Song, Y.; Lee, J.H.; Choi, S.Y.; Park, Y.M. Epigallocatechin-3-Gallate Can Prevent Type 2 Human Papillomavirus E7 from Suppressing Interferon-Stimulated Genes. Int. J. Mol. Sci. 2021, 22, 2418. [Google Scholar] [CrossRef]
- Tatti, S.; Stockfleth, E.; Beutner, K.R.; Tawfik, H.; Elsasser, U.; Weyrauch, P.; Mescheder, A. Polyphenon E: A New Treatment for External Anogenital Warts. Br. J. Dermatol. 2010, 162, 176–184. [Google Scholar] [CrossRef]
- Tzellos, T.G.; Sardeli, C.; Lallas, A.; Papazisis, G.; Chourdakis, M.; Kouvelas, D. Efficacy, Safety and Tolerability of Green Tea Catechins in the Treatment of External Anogenital Warts: A Systematic Review and Meta-Analysis. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 345–353. [Google Scholar] [CrossRef]
- Tatti, S.; Swinehart, J.M.; Thielert, C.; Tawfik, H.; Mescheder, A.; Beutner, K.R. Sinecatechins, a Defined Green Tea Extract, in the Treatment of External Anogenital Warts: A Randomized Controlled Trial. Obstet. Gynecol. 2008, 111, 1371–1379. [Google Scholar] [CrossRef]
- Stockfleth, E.; Beti, H.; Orasan, R.; Grigorian, F.; Mescheder, A.; Tawfik, H.; Thielert, C. Topical Polyphenon E in the Treatment of External Genital and Perianal Warts: A Randomized Controlled Trial. Br. J. Dermatol. 2008, 158, 1329–1338. [Google Scholar] [CrossRef]
- Gross, G.; Meyer, K.-G.; Pres, H.; Thielert, C.; Tawfik, H.; Mescheder, A. A Randomized, Double-Blind, Four-Arm Parallel-Group, Placebo-Controlled Phase II/III Study to Investigate the Clinical Efficacy of Two Galenic Formulations of Polyphenon E in the Treatment of External Genital Warts. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Ahn, W.-S.; Yoo, J.; Huh, S.-W.; Kim, C.-K.; Lee, J.-M.; Namkoong, S.-E.; Bae, S.-M.; Lee, I.P. Protective Effects of Green Tea Extracts (Polyphenon E and EGCG) on Human Cervical Lesions. Eur. J. Cancer Prev. 2003, 12, 383–390. [Google Scholar] [CrossRef]
- Butterworth, C.E.; Hatch, K.D.; Macaluso, M.; Cole, P.; Sauberlich, H.E.; Soong, S.J.; Borst, M.; Baker, V.V. Folate Deficiency and Cervical Dysplasia. JAMA 1992, 267, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Butterworth, C.E.; Hatch, K.D.; Gore, H.; Mueller, H.; Krumdieck, C.L. Improvement in Cervical Dysplasia Associated with Folic Acid Therapy in Users of Oral Contraceptives. Am. J. Clin. Nutr. 1982, 35, 73–82. [Google Scholar] [CrossRef]
- Weinstein, S.J.; Ziegler, R.G.; Selhub, J.; Fears, T.R.; Strickler, H.D.; Brinton, L.A.; Hamman, R.F.; Levine, R.S.; Mallin, K.; Stolley, P.D. Elevated Serum Homocysteine Levels and Increased Risk of Invasive Cervical Cancer in US Women. Cancer Causes Control 2001, 12, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Kwanbunjan, K.; Saengkar, P.; Cheeramakara, C.; Thanomsak, W.; Benjachai, W.; Laisupasin, P.; Buchachart, K.; Songmuaeng, K.; Boontaveeyuwat, N. Low Folate Status as a Risk Factor for Cervical Dysplasia in Thai Women. Nutr. Res. 2005, 25, 641–654. [Google Scholar] [CrossRef]
- Piyathilake, C.J.; Badiga, S.; Paul, P.; Vijayaraghavan, K.; Vedantham, H.; Sudula, M.; Sowjanya, P.; Ramakrishna, G.; Shah, K.V.; Partridge, E.E.; et al. Indian Women with Higher Serum Concentrations of Folate and Vitamin B12 Are Significantly Less Likely to Be Infected with Carcinogenic or High-Risk (HR) Types of Human Papillomaviruses (HPVs). Int. J. Women’s Health 2010, 2, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shulpekova, Y.; Nechaev, V.; Kardasheva, S.; Sedova, A.; Kurbatova, A.; Bueverova, E.; Kopylov, A.; Malsagova, K.; Dlamini, J.C.; Ivashkin, V. The Concept of Folic Acid in Health and Disease. Molecules 2021, 26, 3731. [Google Scholar] [CrossRef]
- Abike, F.; Engin, A.B.; Dunder, I.; Tapisiz, O.L.; Aslan, C.; Kutluay, L. Human Papilloma Virus Persistence and Neopterin, Folate and Homocysteine Levels in Cervical Dysplasias. Arch. Gynecol. Obstet. 2011, 284, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Kwaśniewska, A.; Tukendorf, A.; Goździcka-Józefiak, A.; Semczuk-Sikora, A.; Korobowicz, E. Content of Folic Acid and Free Homocysteine in Blood Serum of Human Papillomavirus-Infected Women with Cervical Dysplasia. Eur. J. Gynaecol. Oncol. 2002, 23, 311–316. [Google Scholar] [PubMed]
- Mason, J.B.; Levesque, T. Folate: Effects on Carcinogenesis and the Potential for Cancer Chemoprevention. Oncology (Williston Park) 1996, 10, 1727–1736, 1742–1743; discussion 1743–1744. [Google Scholar] [PubMed]
- Van Dyke, T.; Finlay, C.; Levine, A.J. A Comparison of Several Lines of Transgenic Mice Containing the SV40 Early Genes. Cold Spring Harb. Symp. Quant. Biol. 1985, 50, 671–678. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Vogelstein, B. A Technique for Radiolabeling DNA Restriction Endonuclease Fragments to High Specific Activity. Anal. Biochem. 1983, 132, 6–13. [Google Scholar] [CrossRef]
- Bailey, L.B.; Gregory, J.F. Folate Metabolism and Requirements. J. Nutr. 1999, 129, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Herbert, V. The Role of Vitamin B12 and Folate in Carcinogenesis. Adv. Exp. Med. Biol. 1986, 206, 293–311. [Google Scholar] [CrossRef]
- Pirisi, L.; Yasumoto, S.; Feller, M.; Doniger, J.; DiPaolo, J.A. Transformation of Human Fibroblasts and Keratinocytes with Human Papillomavirus Type 16 DNA. J. Virol. 1987, 61, 1061–1066. [Google Scholar] [CrossRef] [Green Version]
- Kwaśniewska, A.; Tukendorf, A.; Semczuk, M. Folate Deficiency and Cervical Intraepithelial Neoplasia. Eur. J. Gynaecol. Oncol. 1997, 18, 526–530. [Google Scholar]
- Kim, Y.I.; Giuliano, A.; Hatch, K.D.; Schneider, A.; Nour, M.A.; Dallal, G.E.; Selhub, J.; Mason, J.B. Global DNA Hypomethylation Increases Progressively in Cervical Dysplasia and Carcinoma. Cancer 1994, 74, 893–899. [Google Scholar] [CrossRef]
- Duthie, S.J.; Hawdon, A. DNA Instability (Strand Breakage, Uracil Misincorporation, and Defective Repair) Is Increased by Folic Acid Depletion in Human Lymphocytes in Vitro. FASEB J. 1998, 12, 1491–1497. [Google Scholar] [CrossRef] [PubMed]
- Thorland, E.C.; Myers, S.L.; Persing, D.H.; Sarkar, G.; McGovern, R.M.; Gostout, B.S.; Smith, D.I. Human Papillomavirus Type 16 Integrations in Cervical Tumors Frequently Occur in Common Fragile Sites. Cancer Res. 2000, 60, 5916–5921. [Google Scholar] [PubMed]
- Xiao, S.; Tang, Y.-S.; Kusumanchi, P.; Stabler, S.P.; Zhang, Y.; Antony, A.C. Folate Deficiency Facilitates Genomic Integration of Human Papillomavirus Type 16 DNA In Vivo in a Novel Mouse Model for Rapid Oncogenic Transformation of Human Keratinocytes. J. Nutr. 2018, 148, 389–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piyathilake, C.J.; Henao, O.L.; Macaluso, M.; Cornwell, P.E.; Meleth, S.; Heimburger, D.C.; Partridge, E.E. Folate Is Associated with the Natural History of High-Risk Human Papillomaviruses. Cancer Res. 2004, 64, 8788–8793. [Google Scholar] [CrossRef] [Green Version]
- Piyathilake, C.J.; Macaluso, M.; Chambers, M.M.; Badiga, S.; Siddiqui, N.R.; Bell, W.C.; Edberg, J.C.; Partridge, E.E.; Alvarez, R.D.; Johanning, G.L. Folate and Vitamin B12 May Play a Critical Role in Lowering the HPV 16 Methylation-Associated Risk of Developing Higher Grades of CIN. Cancer Prev. Res. 2014, 7, 1128–1137. [Google Scholar] [CrossRef] [Green Version]
- Yenigul, N.N.; Yazıcı Yılmaz, F.; Ayhan, I. Can Serum Vitamin B12 and Folate Levels Predict HPV Penetration in Patients with ASCUS? Nutr. Cancer 2021, 73, 602–608. [Google Scholar] [CrossRef]
- Balazs, E.A. Viscoelastic Properties of Hyaluronic Acid and Biological Lubrication. Univ. Mich. Med. Cent. J. 1968, 255–259. [Google Scholar]
- Lokeshwar, V.B.; Selzer, M.G. Differences in Hyaluronic Acid-Mediated Functions and Signaling in Arterial, Microvessel, and Vein-Derived Human Endothelial Cells. J. Biol. Chem. 2000, 275, 27641–27649. [Google Scholar] [CrossRef] [Green Version]
- Day, A.J.; de la Motte, C.A. Hyaluronan Cross-Linking: A Protective Mechanism in Inflammation? Trends Immunol. 2005, 26, 637–643. [Google Scholar] [CrossRef]
- Woodman, C.B.J.; Collins, S.I.; Young, L.S. The Natural History of Cervical HPV Infection: Unresolved Issues. Nat. Rev. Cancer 2007, 7, 11–22. [Google Scholar] [CrossRef]
- Angelou, K.; Grigoriadis, T.; Diakosavvas, M.; Zacharakis, D.; Athanasiou, S. The Genitourinary Syndrome of Menopause: An Overview of the Recent Data. Cureus 2020, 12, e7586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essendoubi, M.; Gobinet, C.; Reynaud, R.; Angiboust, J.F.; Manfait, M.; Piot, O. Human Skin Penetration of Hyaluronic Acid of Different Molecular Weights as Probed by Raman Spectroscopy. Skin Res. Technol. 2016, 22, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Galia, T.L.; Micali, A.; Puzzolo, D.; Cancellieri, F. Oral Low-Molecular Weight Hyaluronic Acid in the Treatment of Atrophic Vaginitis. Int. J. Clin. Med. 2014, 5, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Song, L.; Zou, Y.; Sun, D.; Wang, L.; Yu, Z.; Guo, J. Role of Hyaluronic Acids and Potential as Regenerative Biomaterials in Wound Healing. ACS Appl. Bio Mater. 2021, 4, 311–324. [Google Scholar] [CrossRef]
- Gao, F.; Yang, C.X.; Mo, W.; Liu, Y.W.; He, Y.Q. Hyaluronan Oligosaccharides Are Potential Stimulators to Angiogenesis via RHAMM Mediated Signal Pathway in Wound Healing. Clin. Investig. Med. 2008, 31, E106–E116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riccio, M.; Marchesini, A.; Senesi, L.; Skrami, E.; Gesuita, R.; De Francesco, F. Managing Pathologic Scars by Injecting Auto-Cross-Linked Hyaluronic Acid: A Preliminary Prospective Clinical Study. Aesthetic Plast. Surg. 2019, 43, 480–489. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, F.; Riccio, M.; Jimi, S. Contribution of Topical Agents Such as Hyaluronic Acid and Silver Sulfadiazine to Wound Healing and Management of Bacterial Biofilm. Medicina 2022, 58, 835. [Google Scholar] [CrossRef]
- Li, X.; Lv, H.-F.; Zhao, R.; Ying, M.-F.; Samuriwo, A.T.; Zhao, Y.-Z. Recent Developments in Bio-Scaffold Materials as Delivery Strategies for Therapeutics for Endometrium Regeneration. Mater. Today Bio 2021, 11, 100101. [Google Scholar] [CrossRef]
- Nyman, E.; Henricson, J.; Ghafouri, B.; Anderson, C.D.; Kratz, G. Hyaluronic Acid Accelerates Re-Epithelialization and Alters Protein Expression in a Human Wound Model. Plast. Reconstr. Surg. Glob. Open 2019, 7, e2221. [Google Scholar] [CrossRef]
- Ruiz-Huertas, P.; Borrego-Luque, A.; Toledano-Valero, P.; Manzotti, C.; Rollón-Mayordomo, Á. Oral Florid Papillomatosis: Topical Treatment with 5% Imiquimod in Orabase. Clin. Exp. Dent. Res. 2022, 8, 858–862. [Google Scholar] [CrossRef]
- Riemma, G.; Schettino, M.T.; Munno, G.M.; Fasulo, D.D.; Sandullo, L.; Amabile, E.; La Verde, M.; Torella, M. Echinacea Angustifolia and Echinacea Purpurea Supplementation Combined with Vaginal Hyaluronic Acid to Boost the Remission of Cervical Low-Grade Squamous Intraepithelial Lesions (L-SILs): A Randomized Controlled Trial. Medicina 2022, 58, 646. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laganà, A.S.; Chiantera, V.; Gerli, S.; Proietti, S.; Lepore, E.; Unfer, V.; Carugno, J.; Favilli, A. Preventing Persistence of HPV Infection with Natural Molecules. Pathogens 2023, 12, 416. https://doi.org/10.3390/pathogens12030416
Laganà AS, Chiantera V, Gerli S, Proietti S, Lepore E, Unfer V, Carugno J, Favilli A. Preventing Persistence of HPV Infection with Natural Molecules. Pathogens. 2023; 12(3):416. https://doi.org/10.3390/pathogens12030416
Chicago/Turabian StyleLaganà, Antonio Simone, Vito Chiantera, Sandro Gerli, Sara Proietti, Elisa Lepore, Vittorio Unfer, Jose Carugno, and Alessandro Favilli. 2023. "Preventing Persistence of HPV Infection with Natural Molecules" Pathogens 12, no. 3: 416. https://doi.org/10.3390/pathogens12030416