
Bacterial and Viral Pathogens with One Health Relevance in Invasive Raccoons (Procyon lotor, Linné 1758) in Southwest Germany
 ,
,
Abstract
:1. Introduction
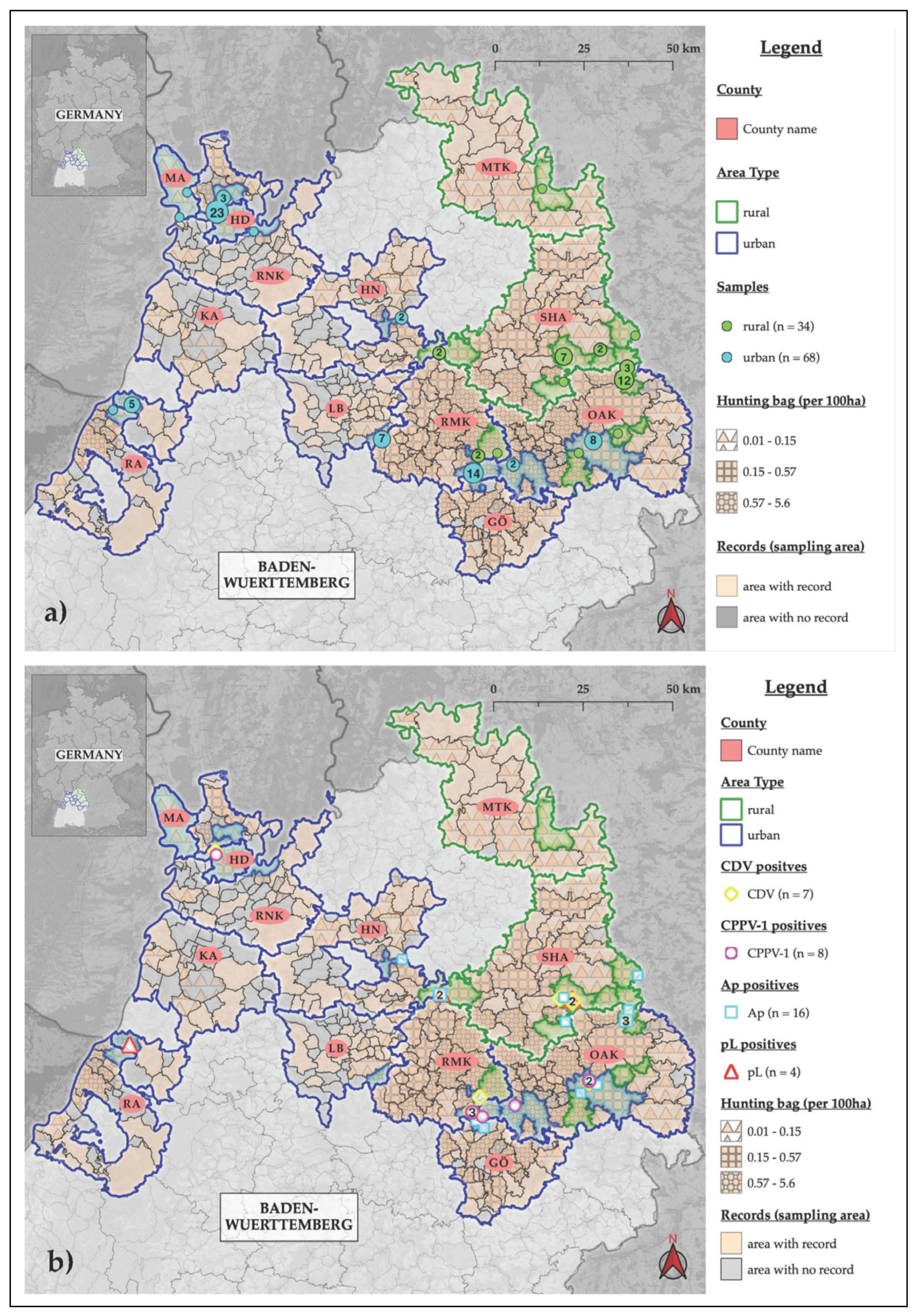
2. Results
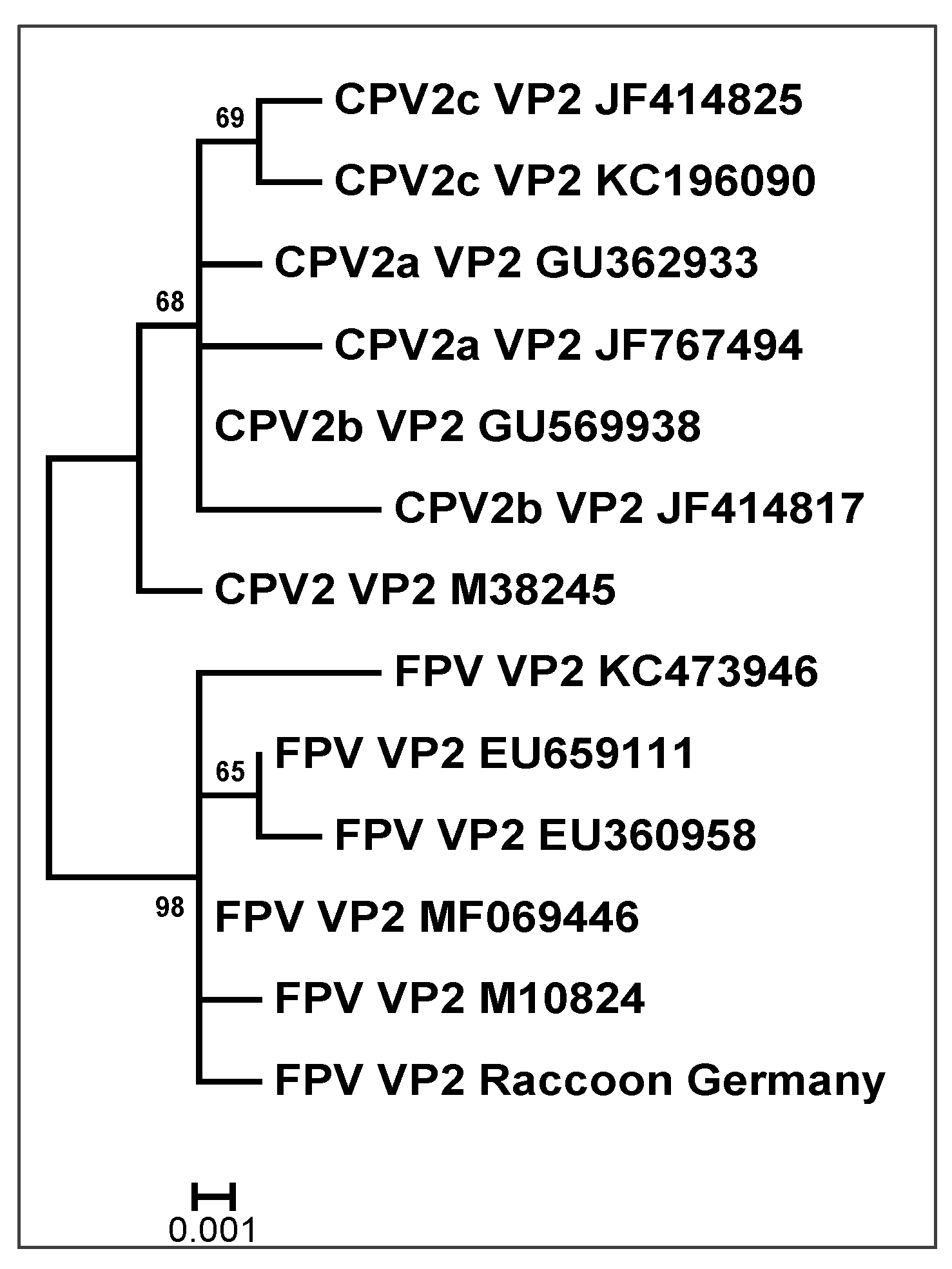
2.1. Carnivore Protoparvovirus-1
2.2. Canine Distemper Virus
2.3. Pathogenic Leptospira spp.
2.4. Anaplasma phagocytophilum
3. Discussion
3.1. Method Limitations
3.2. Carnivores Protoparvovirus-1
3.3. Canine Distemper Virus
3.4. Pathogenic Leptospira spp.
3.5. Anaplasma phagocytophilum
3.6. Lack of Detection for IAV and WNV
3.7. Raccoons and Pathogen Pollution
4. Materials and Methods
4.1. Sample Collection
4.2. Necropsy
4.3. RNA and DNA Extraction
4.4. PCR Analysis
4.5. Controls
4.6. Sequencing
4.7. Statistics and Graphics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yon, L.; Duff, J.P.; Agren, E.O.; Erdelyi, K.; Ferroglio, E.; Godfroid, J.; Hars, J.; Hestvik, G.; Horton, D.; Kuiken, T.; et al. Recent Changes in Infectious Diseases in European Wildlife. J. Wildl. Dis. 2019, 55, 3–43. [Google Scholar] [CrossRef] [PubMed]
- Tomassone, L.; Berriatua, E.; De Sousa, R.; Duscher, G.G.; Mihalca, A.D.; Silaghi, C.; Sprong, H.; Zintl, A. Neglected Vector-Borne Zoonoses in Europe: Into the Wild. Vet. Parasitol. 2018, 251, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Vorou, R.M.; Papavassiliou, V.G.; Tsiodras, S. Emerging Zoonoses and Vector-Borne Infections Affecting Humans in Europe. Epidemiol. Infect. 2007, 135, 1231–1247. [Google Scholar] [CrossRef] [PubMed]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging Infectious Diseases of Wildlife–Threats to Biodiversity and Human Health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Kuiken, T. Implications of Transformative Changes for Research on Emerging Zoonoses. Ecohealth 2021, 18, 275–279. [Google Scholar] [CrossRef]
- Thompson, R.C. Parasite Zoonoses and Wildlife: One Health, Spillover and Human Activity. Int. J. Parasitol. 2013, 43, 1079–1088. [Google Scholar] [CrossRef]
- Fischer, M.L.; Sullivan, M.J.P.; Greiser, G.; Guerrero-Casado, J.; Heddergott, M.; Hohmann, U.; Keuling, O.; Lang, J.; Martin, I.; Michler, F.-U.; et al. Assessing and Predicting the Spread of Non-Native Raccoons in Germany Using Hunting Bag Data and Dispersal Weighted Models. Biol. Invasions 2016, 18, 57–71. [Google Scholar] [CrossRef]
- Fischer, M.L.; Hochkirch, A.; Heddergott, M.; Schulze, C.; Anheyer-Behmenburg, H.E.; Lang, J.; Michler, F.U.; Hohmann, U.; Ansorge, H.; Hoffmann, L.; et al. Historical Invasion Records Can Be Misleading: Genetic Evidence for Multiple Introductions of Invasive Raccoons (Procyon lotor) in Germany. PLoS ONE 2015, 10, e0125441. [Google Scholar] [CrossRef]
- Mazzamuto, M.V.; Panzeri, M.; Bisi, F.; Wauters, L.A.; Preatoni, D.; Martinoli, A. When Management Meets Science: Adaptive Analysis for the Optimization of the Eradication of the Northern Raccoon (Procyon lotor). Biol. Invasions 2020, 22, 3119–3130. [Google Scholar] [CrossRef]
- Mackenstedt, U.; Jenkins, D.; Romig, T. The Role of Wildlife in the Transmission of Parasitic Zoonoses in Peri-Urban and Urban Areas. Int. J. Parasitol. Parasites Wildl. 2015, 17, 71–79. [Google Scholar] [CrossRef]
- Prange, S.; Gehrt, S.D.; Wiggers, E.P. Influences of Anthropogenic Resources on Raccoon (Procyon lotor) Movements and Spatial Distribution. J. Mamm. 2004, 85, 483–490. [Google Scholar] [CrossRef]
- Vos, A.; Ortmann, S.; Kretzschmar, A.S.; Köhnemann, B.; Michel, F. The Raccoon (Procyon lotor) as Potential Rabies Reservoir Species in Germany: A Risk Assessment. Berl. Und Münchner Tierärztliche Wochenschr. 2012, 125, 228–235. [Google Scholar] [CrossRef]
- Deutscher Jagdverband, E.V. Infografik_Jahresjagdstrecke_Waschbär_2020/2021. Dtsch. Jagdverband—Handb. 2022. Available online: https://www.jagdverband.de/sites/default/files/2022-02/2022-01_Infografik_Jahresjagdstrecke_Waschbaer_2020_2021.jpg (accessed on 26 January 2023).
- Fiderer, C.; Göttert, T.; Zeller, U. Spatial Interrelations between Raccoons (Procyon lotor), Red Foxes (Vulpes vulpes), and Ground-Nesting Birds in a Special Protection Area of Germany. Eur. J. Wildl. Res. 2019, 65, 14. [Google Scholar] [CrossRef]
- Salgado, I. Is the Raccoon (Procyon lotor) out of Control in Europe? Biodivers Conserv. 2018, 27, 2243–2256. [Google Scholar] [CrossRef]
- Beltrán-Beck, B.; García, F.J.; Gortázar, C. Raccoons in Europe: Disease Hazards due to the Establishment of an Invasive Species. Eur. J. Wildl. Res. 2012, 58, 5–15. [Google Scholar] [CrossRef]
- Wünschmann, A.; Lopez-Astacio, R.; Armién, A.G.; Reed, L.; Parrish, C.R. Parvovirus-Induced Encephalitis in a Juvenile Raccoon. J. Vet. Diagn. Invest. 2021, 33, 140–143. [Google Scholar] [CrossRef]
- Canuti, M.; Britton, A.P.; Graham, S.M.; Lang, A.S. Epidemiology and Molecular Characterization of Protoparvoviruses Infecting Wild Raccoons (Procyon lotor) in British Columbia, Canada. Virus Res. 2017, 242, 85–89. [Google Scholar] [CrossRef]
- Allison, A.B.; Kohler, D.J.; Fox, K.A.; Brown, J.D.; Gerhold, R.W.; Shearn-Bochsler, V.I.; Dubovi, E.J.; Parrish, C.R.; Holmes, E.C. Frequent Cross-Species Transmission of Parvoviruses Among Diverse Carnivore Hosts. J. Virol. 2013, 87, 2342–2347. [Google Scholar] [CrossRef]
- Hoelzer, K.; Parrish, C.R. The Emergence of Parvoviruses of Carnivores. Vet. Res. 2010, 41, 39. [Google Scholar] [CrossRef]
- Rentería-Solís, Z. Disease Occurrence in Free-Ranging Raccoons (Procyon lotor) from Rural and Urban Populations in North-Eastern Germany. Ph.D. Thesis, Freien Universität Berlin, Berlin, Germany, 2015. [Google Scholar]
- Wostenberg, D.J.; Walker, N.; Fox, K.A.; Spraker, T.R.; Piaggio, A.J.; Gilbert, A. Evidence of Two Cocirculating Canine Distemper Virus Strains in Mesocarnivores from Northern Colorado, USA. J. Wildl. Dis. 2018, 54, 534–543. [Google Scholar] [CrossRef]
- Martinez-Gutierrez, M.; Ruiz-Saenz, J. Diversity of Susceptible Hosts in Canine Distemper Virus Infection: A Systematic Review and Data Synthesis. BMC Vet. Res. 2016, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Rentería-Solís, Z.; Förster, C.; Aue, A.; Wittstatt, U.; Wibbelt, G.; König, M. Canine Distemper Outbreak in Raccoons Suggests Pathogen Interspecies Transmission Amongst Alien and Native Carnivores in Urban Areas from Germany. Vet. Microbiol. 2014, 174, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, E.; Fujii, K.; Ogawa, H.; Imai, K. First Detection of Influenza A Virus Genes from Wild Raccoons in Japan. Virus Genes 2018, 54, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Root, J.J.; Bosco-Lauth, A.M.; Bielefeldt-Ohmann, H.; Bowen, R.A. Experimental Infection of Peridomestic Mammals with Emergent H7N9 (A/Anhui/1/2013) Influenza A Virus: Implications for Biosecurity and Wet Markets. Virology 2016, 487, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.S.; Bentler, K.T.; Landolt, G.; Elmore, S.A.; Minnis, R.B.; Campbell, T.A.; Barras, S.C.; Root, J.J.; Pilon, J.; Pabilonia, K.; et al. Influenza Infection in Wild Raccoons. Emerg. Infect. Dis. 2008, 14, 1842–1848. [Google Scholar] [CrossRef]
- Keller, M.; Peter, N.; Holicki, C.M.; Schantz, A.V.; Ziegler, U.; Eiden, M.; Dorge, D.D.; Vilcinskas, A.; Groschup, M.H.; Klimpel, S. SARS-CoV-2 and West Nile Virus Prevalence Studies in Raccoons and Raccoon Dogs from Germany. Viruses 2022, 14, 2559. [Google Scholar] [CrossRef]
- Root, J.J.; Bosco-Lauth, A.M. West Nile Virus Associations in Wild Mammals: An Update. Viruses 2019, 11, 459. [Google Scholar] [CrossRef]
- Root, J.J.; Bentler, K.T.; Nemeth, N.M.; Gidlewski, T.; Spraker, T.R.; Franklin, A.B. Experimental Infection of Raccoons (Procyon lotor) with West Nile Virus. Am. J. Trop. Med. Hyg. 2010, 83, 803–807. [Google Scholar] [CrossRef]
- Straub, M.H.; Church, M.; Glueckert, E.; Foley, J.E. Raccoons (Procyon lotor) and Striped Skunks (Mephitis mephitis) as Potential Reservoirs of Leptospira spp. in California. Vector Borne Zoonotic Dis. 2020, 20, 418–426. [Google Scholar] [CrossRef]
- Pedersen, K.; Anderson, T.D.; Maison, R.M.; Wiscomb, G.W.; Pipas, M.J.; Sinnett, D.R.; Baroch, J.A.; Gidlewski, T. Leptospira Antibodies Detected in Wildlife in the USA and the US Virgin Islands. J. Wildl. Dis. 2018, 54, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Andre, M.R. Diversity of Anaplasma and Ehrlichia/Neoehrlichia Agents in Terrestrial Wild Carnivores Worldwide: Implications for Human and Domestic Animal Health and Wildlife Conservation. Front. Vet. Sci. 2018, 5, 293. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, J.; Buńkowska-Gawlik, K.; Adamczyk, M.; Gajda, E.; Merta, D.; Popiołek, M.; Perec-Matysiak, A. The Occurrence of Anaplasmataceae in European Populations of Invasive Carnivores. Ticks Tick Borne Dis. 2018, 9, 934–937. [Google Scholar] [CrossRef] [PubMed]
- Bundesinstitut für Bau-Stadt- und Raumforschung (BBSR). Raumabgrenzungen und Raumtypen des BBSR. Available online: https://www.bbsr.bund.de/BBSR/DE/forschung/raumbeobachtung/Raumabgrenzungen/deutschland/gemeinden/Raumtypen2010_vbg/Raumtypen2010_LageSied.html#doc2826692bodyText2 (accessed on 26 January 2023).
- Bundesinstitut für Bau-Stadt- und Raumforschung (BBSR). Raumtypen 2010: Besiedelung und Lage. Available online: https://www.bbsr.bund.de/BBSR/DE/forschung/raumbeobachtung/Raumabgrenzungen/deutschland/gemeinden/Raumtypen2010_vbg/raumTypenBesiedlLage_2020.csv?__blob=publicationFile&v=2 (accessed on 26 January 2023).
- Decaro, N.; Elia, G.; Martella, V.; Desario, C.; Campolo, M.; Trani, L.D.; Tarsitano, E.; Tempesta, M.; Buonavoglia, C. A Real-Time PCR Assay for Rapid Detection and Quantitation of Canine Parvovirus Type 2 in the Feces of Dogs. Vet. Microbiol. 2005, 105, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Elia, G.; Decaro, N.; Martella, V.; Cirone, F.; Lucente, M.S.; Lorusso, E.; Di Trani, L.; Buonavoglia, C. Detection of Canine Distemper Virus in Dogs by Real-Time RT-PCR. J. Virol. Methods 2006, 136, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.S.; Costa, P.; Rocha, T.; Amaro, A.; Vieira, M.L.; Ahmed, A.; Thompson, G.; Hartskeerl, R.A.; Inácio, J. Direct Detection and Differentiation of Pathogenic Leptospira Species Using a Multi-Gene Targeted Real Time PCR Approach. PLoS ONE 2014, 9, e112312. [Google Scholar] [CrossRef]
- Courtney, J.W.; Kostelnik, L.M.; Zeidner, N.S.; Massung, R.F. Multiplex Real-Time PCR for Detection of Anaplasma Phagocytophilum and Borrelia Burgdorferi. J. Clin. Microbiol. 2004, 42, 3164–3168. [Google Scholar] [CrossRef]
- Sainz, Á.; Roura, X.; Miró, G.; Estrada-Peña, A.; Kohn, B.; Harrus, S.; Solano-Gallego, L. Guideline for Veterinary Practitioners on Canine Ehrlichiosis and Anaplasmosis in Europe. Parasit Vectors 2015, 8, 01–20. [Google Scholar] [CrossRef]
- Overzier, E.; Pfister, K.; Herb, I.; Mahling, M.; Böck, G., Jr.; Silaghi, C. Detection of Tick-Borne Pathogens in Roe Deer (Capreolus capreolus), in Questing Ticks (Ixodes ricinus), and in Ticks Infesting Roe Deer in Southern Germany. Ticks Tick Borne Dis. 2013, 4, 320–328. [Google Scholar] [CrossRef]
- Hamir, A.N. Pathology of Neurologic Disorders of Raccoons (Procyon lotor). J. Vet. Diagn. Invest. 2011, 23, 873–884. [Google Scholar] [CrossRef]
- Williams, E.S. Canine Distemper. In Infectious Diseases of Wild Mammals; Williams, E.S., Barker, I.K., Eds.; Iowa State University Press: Ames, IA, USA, 2001; Volume 3, pp. 50–59. [Google Scholar]
- Giacinti, J.A.; Pearl, D.L.; Ojkic, D.; Jardine, C.M. Comparison of Two Surveillance Components for Investigating the Epidemiology of Canine Distemper Virus in Raccoons (Procyon lotor). J. Wildl. Dis. 2021, 57, 104–115. [Google Scholar] [CrossRef]
- Michler, F.U. Säugetierkundliche Freilandforschung zur Populationsbiologie des Waschbären (Procyon lotor Linnaeus, 1758) in einem naturnahen Tieflandbuchenwald im Müritz-Nationalpark (Mecklenburg-Vorpommern). Ph.D. Thesis, Technische Universität Dresden, Dresden, Germany, 2016. [Google Scholar]
- Nemeth, N.M.; Oesterle, P.T.; Campbell, G.D.; Ojkic, D.; Jardine, C.M. Comparison of Reverse-Transcription Real-Time PCR and Immunohistochemistry for the Detection of Canine Distemper Virus Infection in Raccoons in Ontario, Canada. J. Vet. Diagn. Invest. 2018, 30, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Silaghi, C.; Santos, A.S.; Gomes, J.; Christova, I.; Matei, I.A.; Walder, G.; Domingos, A.; Bell-Sakyi, L.; Sprong, H.; von Loewenich, F.D.; et al. Guidelines for the Direct Detection of Anaplasma spp. in Diagnosis and Epidemiological Studies. Vector Borne Zoonotic Dis. 2017, 17, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Härtwig, V.; von Loewenich, F.D.; Schulze, C.; Straubinger, R.K.; Daugschies, A.; Dyachenko, V. Detection of Anaplasma Phagocytophilum in Red Foxes (Vulpes vulpes) and Raccoon Dogs (Nyctereutes procyonoides) from Brandenburg, Germany. Ticks Tick Borne Dis. 2014, 5, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Levett, P.N. Leptospirosis. Clin. Microbiol. Rev. 2001, 14, 296–326. [Google Scholar] [CrossRef] [PubMed]
- Frölich, K.; Streich, W.J.; Fickel, J.; Jung, S.; Truyen, U.; Hentschke, J.; Dedek, J.; Prager, D.; Latz, N. Epizootiologic Investigations of Parvovirus Infections in Free-Ranging Carnivores from Germany. J. Wildl. Dis. 2005, 41, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Barker, I.K.; Parrish, C.R. Parvovirus Infections. In Infectious Diseases of Wild Mammals, 3rd ed.; Williams, E.S., Barker, I.K., Eds.; Iowa State University Press: Ames, IA, USA, 2001; pp. 131–146. [Google Scholar]
- Decaro, N.; Buonavoglia, C. Canine Parvovirus—A Review of Epidemiological and Diagnostic Aspects, with Emphasis on Type 2c. Vet. Microbiol. 2012, 155, 1–12. [Google Scholar] [CrossRef]
- Junge, R.E.; Bauman, K.; King, M.; Gompper, M.E. A Serologic Assessment of Exposure to Viral Pathogens and Leptospira in an Urban Raccoon (Procyon lotor) Population Inhabiting a Large Zoological Park. J. Zoo Wildl. Med. 2007, 38, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Nikolin, V.M.; Wibbelt, G.; Michler, F.U.; Wolf, P.; East, M.L. Susceptibility of Carnivore Hosts to Strains of Canine Distemper Virus from Distinct Genetic Lineages. Vet. Microbiol. 2012, 156, 45–53. [Google Scholar] [CrossRef]
- Michler, F.U.; Köhnemann, B.A.; Roth, M.; Speck, S.; Fickel, J.; Wibbelt, G. Todesursachen sendermarkierter Waschbären (Procyon lotor L., 1758) im Müritz-Nationalpark (Mecklenburg-Vorpommern). Beiträge Jagd Wildforschung 2009, 34, 339–355. [Google Scholar]
- Anheyer-Behmenburg, H.E. Untersuchungen zum Vorkommen von Zoonoseerregern und dem kaninen Staupevirus in der Waschbärpopulation Niedersachsen (2011–2013). Ph.D. Thesis, Tierärztliche Hochschule Hannover, Hannover, Germany, 2013. [Google Scholar]
- Danner, K.J. State Rabies and Epidemiology Centre Freiburg, Baden-Württemberg, Germany. Pers. Commun. 2022. [Google Scholar]
- Blazey, B. Chemisches und Veterinäruntersuchungsamt Stuttgart, Baden-Württemberg, Germany. Pers. Commun. 2022. [Google Scholar]
- Dupouey, J.; Faucher, B.; Edouard, S.; Richet, H.; Kodjo, A.; Drancourt, M.; Davoust, B. Human Leptospirosis: An Emerging Risk in Europe? Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Grimm, K.; Rivera, N.A.; Fredebaugh-Siller, S.; Weng, H.Y.; Warner, R.E.; Maddox, C.W.; Mateus-Pinilla, N.E. Evidence of Leptospira Serovars in Wildlife and Leptospiral DNA in Water Sources in a Natural Area in East-Central Illinois, USA. J. Wildl. Dis. 2020, 56, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Duncan, C.; Krafsur, G.; Podell, B.; Baeten, L.A.; LeVan, I.; Charles, B.; Ehrhart, E.J. Leptospirosis and Tularaemia in Raccoons (Procyon lotor) of Larimer County, Colorado. Zoonoses Public Health 2012, 59, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Jaarsma, R.I.; Sprong, H.; Takumi, K.; Kazimirova, M.; Silaghi, C.; Mysterud, A.; Rudolf, I.; Beck, R.; Földvári, G.; Tomassone, L.; et al. Anaplasma Phagocytophilum Evolves in Geographical and Biotic Niches of Vertebrates and Ticks. Parasit. Vectors 2019, 12, 328. [Google Scholar] [CrossRef]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; van Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; van Maanen, C.; Butler, C.M.; Földvári, G.; et al. Circulation of Four Anaplasma Phagocytophilum Ecotypes in Europe. Parasit. Vectors 2014, 7, 365. [Google Scholar] [CrossRef]
- Müller, M. Untersuchung zur Prävalenz von Frühsommer-Meningoenzephalitis-Viren, von Borrelia burgdorferi sensu lato und Anaplasma phagocytophilum in Zecken der Art Ixodes ricinus in Rheinland-Pfalz und im Saarland (Deutschland). Ph.D. Thesis, Universität des Saarlandes, Homburg/Saar, Germany, 2017. [Google Scholar]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma Phagocytophilum—A Widespread Multi-Host Pathogen with Highly Adaptive Strategies. Front. Cell Infect. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef]
- Leonhard, S. Untersuchungen zur Häufigkeit von Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum und Babesia spp. in Ixodes ricinus aus Bayern und Baden-Württemberg. Ph.D. Thesis, Ludwig-Maximilians-University, München, Germany, 2005. [Google Scholar]
- Mitchell, M.A.; Hungerford, L.L.; Nixon, C.; Esker, T.; Sullivan, J.; Koerkenmeier, R.; Dubey, J.P. Serologic Survey for Selected Infectious Disease Agents in Raccoons from Illinois. J. Wildl. Dis. 1999, 35, 347–355. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Kuiken, T.; Niqueux, E.; Staubach, C.; Terregino, C.; Guajardo, I.M.; Baldinelli, F. Avian Influenza Overview November 2019–February 2020. EFSA J. 2020, 18, 6096. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Kuiken, T.; Smietanka, K.; Staubach, C.; Guajardo, M.; Baldinelli, F. Avian influenza Overview August–November 2019. EFSA J. 2019, 17, e05988. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, E.; Staubach, C.; Terregino, C.; Baldinelli, F. Avian Influenza Overview August–December 2020. EFSA J. 2020, 18, e06379. [Google Scholar] [CrossRef] [PubMed]
- Robert-Koch-Institute. Infektionsepidemiologisches Jahrbuch meldepflichtiger Krankheiten für 2019; Robert-Koch-Institute: Berlin, Germany, 2020.
- Robert-Koch-Institute. Infektionsepidemiologisches Jahrbuch meldepflichtiger Krankheiten für 2020; Robert-Koch-Institute: Berlin, Germany, 2021.
- Root, J.J.; Bentler, K.T.; Shriner, S.A.; Mooers, N.L.; VanDalen, K.K.; Sullivan, H.J.; Franklin, A.B. Ecological Routes of Avian Influenza Virus Transmission to a Common Mesopredator: An Experimental Evaluation of Alternatives. PLoS ONE 2014, 9, e102964. [Google Scholar] [CrossRef] [PubMed]
- Fiero, B.C.; Verts, B.J. Comparison of Techniques for Estimating Age of Raccoons. J. Mamm. 1986, 67, 392–395. [Google Scholar] [CrossRef]
- Grau, G.A.; Sanderson, G.C.; Rogers, J.P. Age Determination of Raccoons. J. Wildl. Manag. 1970, 34, 364–372. [Google Scholar] [CrossRef]
- Root, J.J. West Nile virus associations in wild mammals: A synthesis. Arch. Virol. 2013, 158, 735–752. [Google Scholar] [CrossRef]
- Eiden, M.; Vina-Rodriguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M.H. Two New Real-Time Quantitative Reverse Transcription Polymerase Chain Reaction Assays with Unique Target Sites for the Specific and Sensitive Detection of Lineages 1 and 2 West Nile Virus Strains. J. Vet. Diagn. Invest. 2010, 22, 748–753. [Google Scholar] [CrossRef]
- Pérez, R.; Bianchi, P.; Calleros, L.; Francia, L.; Hernández, M.; Maya, L.; Panzera, Y.; Sosa, K.; Zoller, S. Recent Spreading of a Divergent Canine Parvovirus Type 2a (CPV-2a) Strain in a CPV-2c Homogenous Population. Vet. Microbiol. 2012, 155, 214–219. [Google Scholar] [CrossRef]
- Nau, L.H.; Emirhar, D.; Obiegala, A.; Mylius, M.; Runge, M.; Jacob, J.; Bier, N.; Nöckler, K.; Imholt, C.; Below, D.; et al. Leptospirosis in Germany: Current Knowledge on Pathogen Species, Reservoir Hosts, and Disease in Humans and Animals. Bundesgesundheitsblatt Gesundh. Gesundh. 2019, 62, 1510–1521. [Google Scholar] [CrossRef]
- Guglielmini, J.; Bourhy, P.; Schiettekatte, O.; Zinini, F.; Brisse, S.; Picardeau, M. Genus-Wide Leptospira Core Genome Multilocus Sequence Typing for Strain Taxonomy and Global Surveillance. PLoS Negl. Trop. Dis. 2019, 13, e0008673. [Google Scholar] [CrossRef]
- Durski, K.N.; Jancloes, M.; Chowdhary, T.; Bertherat, E. A Global, Multi-Disciplinary, Multi-Sectorial Initiative to Combat Leptospirosis: Global Leptospirosis Environmental Action Network (GLEAN). Int. J. Environ. Res. Public Health 2014, 11, 6000–6008. [Google Scholar] [CrossRef]
- Hartskeerl, R.A.; Collares-Pereira, M.; Ellis, W.A. Emergence, Control and Re-emerging Leptospirosis: Dynamics of Infection in the Changing World. Clin. Microbiol. Infect. 2011, 17, 494–501. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| No. Pos (%; CI 95%) | ||||||
|---|---|---|---|---|---|---|
| Parameters | No. | Positives | CPPV-1 | CDV | pL | Ap |
| Total | 102 | 30 (28.3; 20.8–39.3) | 8 (7.8; 3.4–14.9) | 7 (6.9; 2.8–13.6) | 4 (3.9; 1.1–9.7) | 16 (15.7; 9.2–24.2) |
| Years | ||||||
| 2019 | 33 (32.4; 23.4–42.3) | 10 (30.3; 15.6–48.7) | 1 (3; 1.9–24.3) | 3 (9.1; 1.9–24.3) | 2 (6.1; 0.7–20.2) | 6 (18.2; 7–35.5) |
| 2020 | 69 (67.6; 57.7–76.6) | 20 (29; 18.7–41.2) | 7 (10.1; 4.2–19.8) | 4 (5.8; 1.6–14.2) | 2 (2.9; 0.4–10.1) | 10 (14.5; 7.2–25) |
| Area | ||||||
| rural | 34 (33.3; 24.3–43.4) | 14 (41.2; 24.6–59.3) | 0 (0; 0–10.3) | 4 (11.8; 3.3–27.5) | 3 (8.8; 1.9–23.7) | 10 (29.4; 15.1–47.5) § |
| urban | 68 (66.7; 56.6–75.7) | 16 (23.5; 14.1–35.4) | 8 (11.8; 5.2–21.9) | 3 (4.4; 0.9–12.4) | 1 (1.5; 0.03–7.9) | 6 (8.8; 3.3–18.2) § |
| Sex | ||||||
| ♀ | 39 (38.2; 28.8–48.4) | 11 (28.2; 15–44.9) | 3 (7.7; 1.6–20.9) | 4 (10.3; 2.9–24.2) | 2 (5.1; 0.6–17.3) | 5 (12.8; 4.3–27.4) |
| ♂ | 63 (61.8; 51.6–71.2) | 19 (30.2; 19.2–43) | 5 (7.9; 2.6–17.6) | 3 (4.8; 1–13.3) | 2 (3.2;0.4–11) | 11 (17.5; 9.1–29.1) |
| Age | ||||||
| >1 a | 56 (54.9; 44.7–64.8) | 23 (41.1; 28.2–55.0) | 6 (10.7; 4–21.9) | 6 (10.7; 4–21.9) | 2 (3.6; 0.4–12.3) | 12 (21.4; 11.6–34.4) |
| <1 a | 46 (45.1; 35.2–55.3) | 7 (15.2; 6.3–28.9) | 2 (4.4; 0.5–14.8) | 1 (2.2; 0.1–11.5) | 2 (4.4; 0.5–14.8) | 4 (8.7; 2.4–20.8) |
| Age-Sex Class | ||||||
| >1 a, ♀ | 16 (15.7; 9.2–24.2) | 7 (43.8; 19.8–70.1) | 2 (12.5; 1.6–38.3) | 3 (18.8; 4.1–45.6) | 0 (0; 0–20.6) | 3 (18.8; 4.1–45.6) |
| >1 a, ♂ | 40 (39.2; 29.7–49.4) | 16 (40; 24.9–56.7) | 4 (10; 2.8–23.7) | 3 (7.5; 1.6–20.4) | 2 (5; 0.6–16.9) | 9 (22.5; 10.8–38.5) |
| <1 a, ♀ | 23 (22.5; 14.9–31.9) | 4 (17.4; 5–38.8) | 1 (4.4; 0.1–21.9) | 1 (4.4; 0.1–21.9) | 2 (8.7; 1.1–28) | 2 (8.7; 1.1–28) |
| <1 a, ♂ | 23 (22.5; 14.9–31.9) | 3 (13; 2.8–33.6) | 1 (4.4; 0.1–21.9) | 0 (0; 0–14.8) | 0 (0; 0–14.8) | 2 (8.7; 1.1–28) |
| Raccoon No. | District | Municipality | Area Type | Sex | Age | CDV | Ap | CPPV-1 | pL |
|---|---|---|---|---|---|---|---|---|---|
| WB 18 | SHA | Frankenhardt | rural | ♂ | >1 a | + | - | - | + |
| WB 19 | SHA | Frankenhardt | rural | ♀ | <1 a | + | - | - | + |
| WB 34 | OAK | Aalen | urban | ♀ | >1 a | + | - | + | - |
| WB 73 | SHA | Mainhardt | rural | ♀ | <1 a | - | + | - | + |
| WB 91 | OAK | Aalen | urban | ♂ | >1 a | - | + | + | - |
| Pathogen | Sample Material | qPCR |
|---|---|---|
| CDV | brain stem, frontal cortex [44,47] | Elia et al. [38] |
| CPPV-1 | small intestine [53] | Decaro et al. [37] |
| IAV | lung [26,27] | Virotype® Influenza A Kit (Indical) |
| WNV | brain stem, frontal cortex [77] | Eiden et al. [78] |
| pL | kidney, liver [39] | Adiavet Lepto Real Time Kit® (Bio-X, Adiagene), Ferreira et al. [39] |
| Ap | blood, spleen [40,48] | Courtney et al. [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reinhardt, N.P.; Köster, J.; Thomas, A.; Arnold, J.; Fux, R.; Straubinger, R.K. Bacterial and Viral Pathogens with One Health Relevance in Invasive Raccoons (Procyon lotor, Linné 1758) in Southwest Germany. Pathogens 2023, 12, 389. https://doi.org/10.3390/pathogens12030389
Reinhardt NP, Köster J, Thomas A, Arnold J, Fux R, Straubinger RK. Bacterial and Viral Pathogens with One Health Relevance in Invasive Raccoons (Procyon lotor, Linné 1758) in Southwest Germany. Pathogens. 2023; 12(3):389. https://doi.org/10.3390/pathogens12030389
Chicago/Turabian StyleReinhardt, Nico P., Judith Köster, Astrid Thomas, Janosch Arnold, Robert Fux, and Reinhard K. Straubinger. 2023. "Bacterial and Viral Pathogens with One Health Relevance in Invasive Raccoons (Procyon lotor, Linné 1758) in Southwest Germany" Pathogens 12, no. 3: 389. https://doi.org/10.3390/pathogens12030389