
Factors Affecting Arbovirus Midgut Escape in Mosquitoes


Abstract
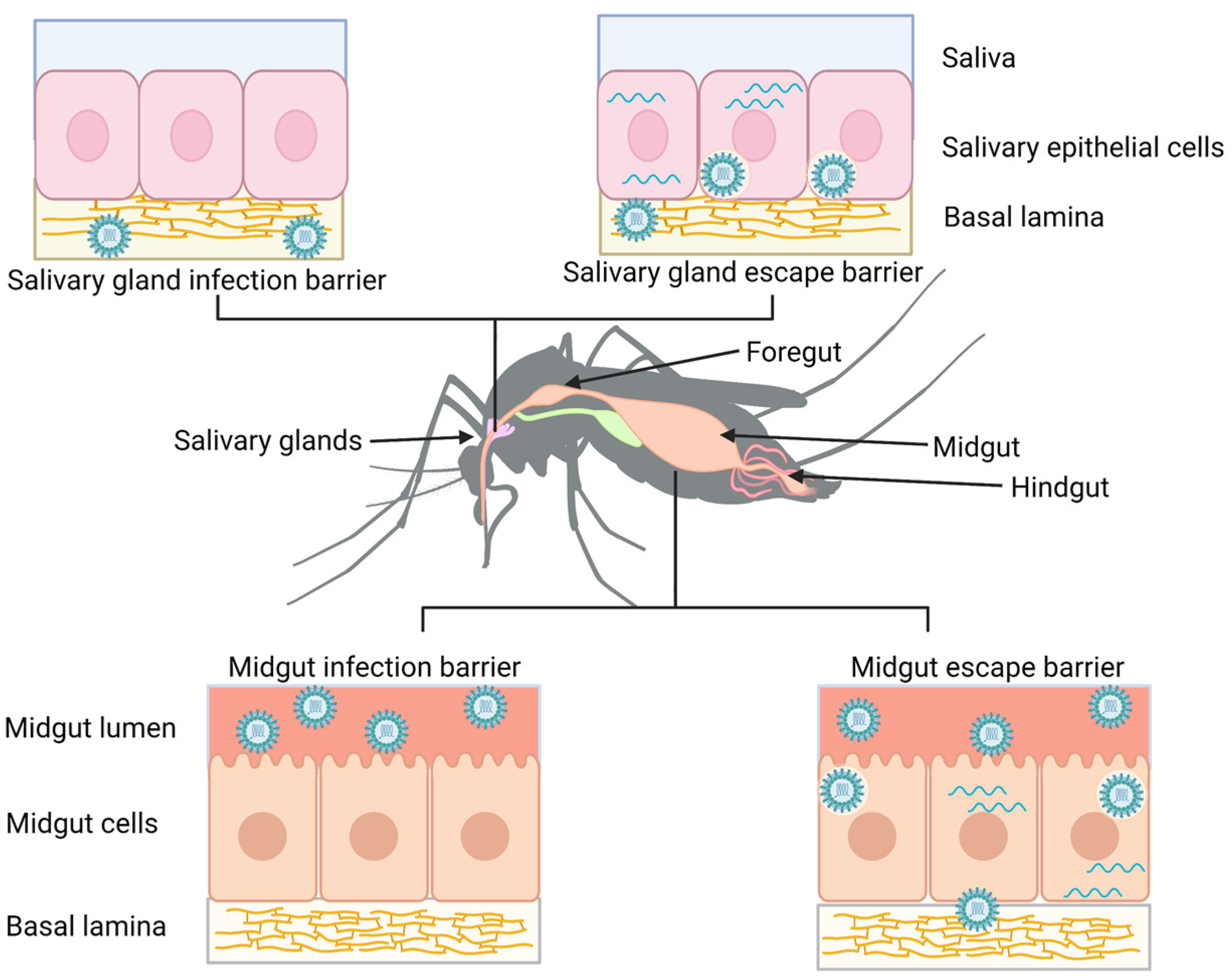
:1. Introduction
2. Routes of Midgut Escape
3. Factors Affecting Midgut Escape
3.1. External Factors
- Temperature
- Exposure to pesticides
- Larval density/competition
3.2. Mosquito Factors
- Physical characteristics—basal lamina thickness and structure
- Immune gene expression
- (i).
- RNAi pathway
- (ii).
- Jak/STAT, Toll and IMD pathways
- (iii).
- Apoptosis and cell turnover
- Behavior—feeding behavior
3.3. Viral Factors
- Midgut replication
- Viral diversity and replication error rate
- Co-infection
- Virus Dose
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Chikungunya and Dengue in the South-West Indian Ocean. WHO—Emergencies Preparedness, Response: Disease Outbreak News. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2006_03_17-en (accessed on 24 January 2023).
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The Global Distribution and Burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan American Health Organization/World Health Organization. Epidemiological Update Yellow Fever Situation Summary in the Americas; Pan Health Organization/World Health Organization: Washington, DC, USA, 2018. [Google Scholar]
- Fauci, A.S.; Morens, D.M. Zika Virus in the Americas—Yet Another Arbovirus Threat. N. Engl. J. Med. 2016, 374, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Liu-Helmersson, J.; Quam, M.; Wilder-Smith, A.; Stenlund, H.; Ebi, K.; Massad, E.; Rocklöv, J. Climate Change and Aedes Vectors: 21st Century Projections for Dengue Transmission in Europe. EBioMedicine 2016, 7, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global Expansion and Redistribution of Aedes-Borne Virus Transmission Risk with Climate Change. PLoS Negl. Trop. Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Mosquitoes in the United States|Mosquitoes|CDC. Available online: https://www.cdc.gov/mosquitoes/about/mosquitoes-in-the-us.html (accessed on 30 March 2022).
- Myles, K.M.; Pierro, D.J.; Olson, K.E. Comparison of the Transmission Potential of Two Genetically Distinct Sindbis Viruses After Oral Infection of Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2004, 41, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Lambrechts, L.; Chevillon, C.; Albright, R.G.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Scott, T.W. Genetic Specificity and Potential for Local Adaptation between Dengue Viruses and Mosquito Vectors. BMC Evol. Biol. 2009, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A Single Mutation in Chikungunya Virus Affects Vector Specificity and Epidemic Potential. PLoS Pathog. 2007, 3, 1895–1906. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; McGee, C.E.; Higgs, S. Chikungunya Virus Adaptation to Aedes albopictus Mosquitoes Does Not Correlate with Acquisition of Cholesterol Dependence or Decreased PH Threshold for Fusion Reaction. Virol. J. 2011, 8, 376. [Google Scholar] [CrossRef] [Green Version]
- Thiberville, S.D.; Moyen, N.; Dupuis-Maguiraga, L.; Nougairede, A.; Gould, E.A.; Roques, P.; de Lamballerie, X. Chikungunya Fever: Epidemiology, Clinical Syndrome, Pathogenesis and Therapy. Antivir. Res. 2013, 99, 345–370. [Google Scholar] [CrossRef]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue Barriers to Arbovirus Infection in Mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef]
- Houk, E.J.; Hardy, J.L.; Chiles, R.E. Permeability of the Midgut Basal Lamina in the Mosquito, Culex Tarsalis Coquillett (Insecta, Diptera). Acta Trop. 1981, 38, 163–171. [Google Scholar] [PubMed]
- Bowers, D.F.; Abell, B.A.; Brown, D.T. Replication and Tissue Tropism of the Alphavirus Sindbis in the Mosquito Aedes albopictus. Virology 1995, 212, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, Y.A.; Klingler, K.A.; Higgs, S. West Nile Virus Dissemination and Tissue Tropisms in Orally Infected Culex pipiens Quinquefasciatus. Vector Borne Zoonotic Dis. 2004, 4, 109–122. [Google Scholar] [CrossRef]
- Miller, B.R.; Mitchell, C.J.; Ballinger, M.E. Replication, Tissue Tropisms and Transmission of Yellow Fever Virus in Aedes albopictus. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Salazar, M.I.; Richardson, J.H.; Sanchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue Virus Type 2: Replication and Tropisms in Orally Infected Aedes aegypti Mosquitoes. BMC Microbiol. 2007, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romoser, W.S.; Wasieloski, L.P.; Pushko, P.; Kondig, J.P.; Lerdthusnee, K.; Neira, M.; Ludwig, G.V. Evidence for Arbovirus Dissemination Conduits from the Mosquito (Diptera: Culicidae) Midgut. J. Med. Entomol. 2004, 41, 467–475. [Google Scholar] [CrossRef] [Green Version]
- Kantor, A.M.; Grant, D.G.; Balaraman, V.; White, T.A.; Franz, A.W.E. Ultrastructural Analysis of Chikungunya Virus Dissemination from the Midgut of the Yellow Fever Mosquito, Aedes aegypti. Viruses 2018, 10, 571. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Grant, D.G.; Lin, J.; Yu, X.; Franz, A.W.E. Zika Virus Dissemination from the Midgut of Aedes aegypti Is Facilitated by Bloodmeal-Mediated Structural Modification of the Midgut Basal Lamina. Viruses 2019, 11, 1056. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C.; Scott, T.W.; Lorenz, L.H.; Lerdthusnee, K.; Romoser, W.S. Togavirus-Associated Pathologic Changes in the Midgut of a Natural Mosquito Vector. J. Virol. 1988, 62, 2083–2090. [Google Scholar] [CrossRef] [Green Version]
- Lerdthusnee, K.; Romoser, W.S.; Faran, M.E.; Dohm, D.J. Rift Valley Fever Virus in the Cardia of Culex pipiens: An Immunocytochemical and Ultrastructural Study. Am. J. Trop. Med. Hyg. 1995, 53, 331–337. [Google Scholar] [CrossRef]
- Romoser, W.S.; Faran, M.E.; Bailey, C.L. Newly Recognized Route of Arbovirus Dissemination from the Mosquito (Diptera: Culicidae) Midgut. J. Med. Entomol. 1987, 24, 431–432. [Google Scholar] [CrossRef] [PubMed]
- Engelhard, E.K.; Kam-Morgan, L.N.W.; Washburn, J.O.; Volkman, L.E. The Insect Tracheal System: A Conduit for the Systemic Spread of Autographa Californica M Nuclear Polyhedrosis Virus. Proc. Natl. Acad. Sci. USA 1994, 91, 3224–3227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Means, J.C.; Passarelli, A.L. Viral Fibroblast Growth Factor, Matrix Metalloproteases, and Caspases Are Associated with Enhancing Systemic Infection by Baculoviruses. Proc. Natl. Acad. Sci. USA 2010, 107, 9825–9830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romoser, W.S.; Turell, M.J.; Lerdthusnee, K.; Neira, M.; Dohm, D.; Ludwig, G.; Wasieloski, L. Pathogenesis of Rift Valley Fever Virus in Mosquitoes--Tracheal Conduits & the Basal Lamina as an Extra-Cellular Barrier. Arch. Virol. Suppl. 2005, 19, 89–100. [Google Scholar] [CrossRef]
- Paulson, S.L.; Grimstad, P.R.; Craig, G.B. Midgut and Salivary Gland Barriers to La Crosse Virus Dissemination in Mosquitoes of the Aedes triseriatus Group. Med. Vet. Entomol. 1989, 3, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Hartman, D.A.; Bergren, N.A.; Kondash, T.; Schlatmann, W.; Webb, C.T.; Kading, R.C. Susceptibility and Barriers to Infection of Colorado Mosquitoes with Rift Valley Fever Virus. PLoS Negl. Trop. Dis. 2021, 15, e0009837. [Google Scholar] [CrossRef]
- Bennett, K.E.; Olson, K.E.; Muñoz, M.d.L.; Fernandez-Salas, I.; Farfan-Ale, J.A.; Higgs, S.; Black IV, W.C.; Beaty, B.J. Variation in Vector Competence for Dengue 2 Virus among 24 Collections of Aedes aegypti from Mexico and the United States. Am. J. Trop. Med. Hyg. 2002, 67, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Boromisa, R.D.; Grayson, M.A. Oral Transmission of Jamestown Canyon Virus by Aedes Provocans Mosquitoes from Northeastern New York. J. Am. Mosq. Control Assoc. 1991, 7, 42–47. [Google Scholar]
- Khoo, C.C.; Doty, J.B.; Held, N.L.; Olson, K.E.; Franz, A.W. Isolation of Midgut Escape Mutants of Two American Genotype Dengue 2 Viruses from Aedes aegypti. Virol. J. 2013, 10, 257. [Google Scholar] [CrossRef] [Green Version]
- Turell, M.J.; Linthicum, K.J.; Patrican, L.A.; Davies, F.G.; Kairo, A.; Bailey, C.L. Vector Competence of Selected African Mosquito (Diptera: Culicidae) Species for Rift Valley Fever Virus. J. Med. Entomol. 2008, 45, 102–108. [Google Scholar] [CrossRef]
- Kramer, L.D.; Hardy, J.L.; Presser, S.B.; Houk, E.J. Dissemination Barriers for Western Equine Encephalomyelitis Virus in Culex Tarsalis Infected after Ingestion of Low Viral Doses. Am. J. Trop. Med. Hyg. 1981, 30, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Dohm, D.J.; O’Guinn, M.L.; Turell, M.J. Effect of Environmental Temperature on the Ability of Culex pipiens (Diptera: Culicidae) to Transmit West Nile Virus. J. Med. Entomol. 2002, 39, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.L.; Mores, C.N.; Lord, C.C.; Tabachnick, W.J. Impact of Extrinsic Incubation Temperature and Virus Exposure on Vector Competence of Culex pipiens Quinquefasciatus Say (Diptera: Culicidae) for West Nile Virus. Vector-Borne Zoonotic Dis. 2007, 7, 629–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alto, B.W.; Bettinardi, D. Temperature and Dengue Virus Infection in Mosquitoes: Independent Effects on the Immature and Adult Stages. Am. J. Trop. Med. Hyg. 2013, 88, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Mbaika, S.; Lutomiah, J.; Chepkorir, E.; Mulwa, F.; Khayeka-Wandabwa, C.; Tigoi, C.; Oyoo-Okoth, E.; Mutisya, J.; Ng’Ang’A, Z.; Sang, R. Vector Competence of Aedes aegypti in Transmitting Chikungunya Virus: Effects and Implications of Extrinsic Incubation Temperature on Dissemination and Infection Rates. Virol. J. 2016, 13, 114. [Google Scholar] [CrossRef]
- Richards, S.L.; Lord, C.C.; Pesko, K.; Tabachnick, W.J. Environmental and Biological Factors Influencing Culex pipiens Quinquefasciatus Say (Diptera: Culicidae) Vector Competence for Saint Louis Encephalitis Virus. Am. J. Trop. Med. Hyg. 2009, 81, 264–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilpatrick, A.M.; Meola, M.A.; Moudy, R.M.; Kramer, L.D. Temperature, Viral Genetics, and the Transmission of West Nile Virus by Culex pipiens Mosquitoes. PLoS Pathog. 2008, 4, e1000092. [Google Scholar] [CrossRef] [Green Version]
- Westbrook, C.J.; Reiskind, M.H.; Pesko, K.N.; Greene, K.E.; Lounibos, L.P. Larval Environmental Temperature and the Susceptibility of Aedes albopictus Skuse (Diptera: Culicidae) to Chikungunya Virus. Vector-Borne Zoonotic Dis. 2010, 10, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Muturi, E.J.; Blackshear, M.; Montgomery, A. Temperature and Density-Dependent Effects of Larval Environment on Aedes aegypti Competence for an Alphavirus. J. Vector Ecol. 2012, 37, 154–161. [Google Scholar] [CrossRef]
- Knecht, H.; Richards, S.; Balanay, J.; White, A. Impact of Mosquito Age and Insecticide Exposure on Susceptibility of Aedes albopictus (Diptera: Culicidae) to Infection with Zika Virus. Pathogens 2018, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.L.; White, A.V.; Balanay, J.A.G. Potential for Sublethal Insecticide Exposure to Impact Vector Competence of Aedes albopictus (Diptera: Culicidae) for Dengue and Zika Viruses. Res. Rep. Trop. Med. 2017, 8, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Muturi, E.J.; Kim, C.-H.; Alto, B.W.; Berenbaum, M.R.; Schuler, M.A. Larval Environmental Stress Alters Aedes aegypti Competence for Sindbis Virus. Trop. Med. Int. Health 2011, 16, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Muturi, E.J.; Alto, B.W. Larval Environmental Temperature and Insecticide Exposure Alter Aedes aegypti Competence for Arboviruses. Vector-Borne Zoonotic Dis. 2011, 11, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.M.; Tomé, H.V.V.; Miranda, F.R.; Gonçalves, W.G.; Pascini, T.V.; Serrão, J.E.; Martins, G.F. Aedes aegypti Larvae Treated with Spinosad Produce Adults with Damaged Midgut and Reduced Fecundity. Chemosphere 2019, 221, 464–470. [Google Scholar] [CrossRef]
- Moltini-Conclois, I.; Stalinski, R.; Tetreau, G.; Després, L.; Lambrechts, L. Larval Exposure to the Bacterial Insecticide Bti Enhances Dengue Virus Susceptibility of Adult Aedes aegypti Mosquitoes. Insects 2018, 9, 193. [Google Scholar] [CrossRef] [Green Version]
- Alto, B.W.; Lounibos, L.P.; Higgs, S.; Juliano, S.A. Larval Competition Differentially Affects Arbovirus Infection in Aedes Mosquitoes. Ecology 2005, 86, 3279–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bevins, S.N. Invasive Mosquitoes, Larval Competition, and Indirect Effects on the Vector Competence of Native Mosquito Species (Diptera: Culicidae). Biol. Invasions 2008, 10, 1109–1117. [Google Scholar] [CrossRef]
- Bennett, K.E.; Beaty, B.J.; Black, W.C. Selection of D2S3, an Aedes aegypti (Diptera: Culicidae) Strain with High Oral Susceptibility to Dengue 2 Virus and D2MEB, a Strain with a Midgut Barrier to Dengue 2 Escape. J. Med. Entomol. 2005, 42, 110–119. [Google Scholar] [CrossRef]
- Grimstad, P.R.; Walker, E.D. Aedes triseriatus (Diptera: Culicidae) and La Crosse Virus. IV. Nutritional Deprivation of Larvae Affects the Adult Barriers to Infection and Transmission. J. Med. Entomol. 1991, 28, 378–386. [Google Scholar] [CrossRef]
- Thomas, R.E.; Wu, W.K.; Verleye, D.; Rai, K.S. Midgut Basal Lamina Thickness and Dengue-1 Virus Dissemination Rates in Laboratory Strains of Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 1993, 30, 326–331. [Google Scholar] [CrossRef]
- Armstrong, P.M.; Ehrlich, H.Y.; Magalhaes, T.; Miller, M.R.; Conway, P.J.; Bransfield, A.; Misencik, M.J.; Gloria-Soria, A.; Warren, J.L.; Andreadis, T.G.; et al. Successive Blood Meals Enhance Virus Dissemination within Mosquitoes and Increase Transmission Potential. Nat. Microbiol. 2020, 5, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and Specific Genetic Interference by Double-Stranded RNA in Caenorhabditis Elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Blair, C.D. Mosquito RNAi Is the Major Innate Immune Pathway Controlling Arbovirus Infection and Transmission. Future Microbiol. 2011, 6, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, C.L.; Keene, K.M.; Brackney, D.E.; Olson, K.E.; Blair, C.D.; Wilusz, J.; Foy, B.D. Aedes aegypti Uses RNA Interference in Defense against Sindbis Virus Infection. BMC Microbiol. 2008, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Vargas, I.; Scott, J.C.; Poole-Smith, B.K.; Franz, A.W.E.; Rie Barbosa-Solomieu, V.; Wilusz, J.; Olson, K.E.; Blair, C.D. Dengue Virus Type 2 Infections of Aedes aegypti Are Modulated by the Mosquito’s RNA Interference Pathway. PLoS Pathog. 2009, 5, e1000299. [Google Scholar] [CrossRef] [Green Version]
- Khoo, C.C.; Piper, J.; Sanchez-Vargas, I.; Olson, K.E.; Franz, A.W. The RNA Interference Pathway Affects Midgut Infection- and Escape Barriers for Sindbis Virus in Aedes aegypti. BMC Microbiol. 2010, 10, 130. [Google Scholar] [CrossRef] [Green Version]
- Franz, A.W.E.; Sanchez-Vargas, I.; Adelman, Z.N.; Blair, C.D.; Beaty, B.J.; James, A.A.; Olson, K.E. Engineering RNA Interference-Based Resistance to Dengue Virus Type 2 in Genetically Modified Aedes aegypti. Proc. Natl. Acad. Sci. USA 2006, 103, 4198–4203. [Google Scholar] [CrossRef] [Green Version]
- Bonizzoni, M.; Dunn, W.A.; Campbell, C.L.; Olson, K.E.; Marinotti, O.; James, A.A. Strain Variation in the Transcriptome of the Dengue Fever Vector, Aedes aegypti. G3 Genes|Genomes|Genet. 2012, 2, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Carvalho-Leandro, D.; Ayres, C.F.J.; Guedes, D.R.D.; Suesdek, L.; Melo-Santos, M.A.V.; Oliveira, C.F.; Cordeiro, M.T.; Regis, L.N.; Marques, E.T.; Gil, L.H.; et al. Immune Transcript Variations among Aedes aegypti Populations with Distinct Susceptibility to Dengue Virus Serotype 2. Acta Trop. 2012, 124, 113–119. [Google Scholar] [CrossRef]
- Dostert, C.; Jouanguy, E.; Irving, P.; Troxler, L.; Galiana-Arnoux, D.; Hetru, C.; Hoffmann, J.A.; Imler, J.L. The Jak-STAT Signaling Pathway Is Required but Not Sufficient for the Antiviral Response of Drosophila. Nat. Immunol. 2005, 6, 946–953. [Google Scholar] [CrossRef]
- Souza-Neto, J.A.; Sim, S.; Dimopoulos, G. An Evolutionary Conserved Function of the JAK-STAT Pathway in Anti-Dengue Defense. Proc. Natl. Acad. Sci. USA 2009, 106, 17841–17846. [Google Scholar] [CrossRef] [PubMed]
- Jupatanakul, N.; Sim, S.; Angleró-Rodríguez, Y.I.; Souza-Neto, J.; Das, S.; Poti, K.E.; Rossi, S.L.; Bergren, N.; Vasilakis, N.; Dimopoulos, G. Engineered Aedes aegypti JAK/STAT Pathway-Mediated Immunity to Dengue Virus. PLoS Negl. Trop. Dis. 2017, 11, e0005187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behura, S.K.; Gomez-Machorro, C.; Debruyn, B.; Lovin, D.D.; Harker, B.W.; Romero-Severson, J.; Mori, A.; Severson, D.W. Influence of Mosquito Genotype on Transcriptional Response to Dengue Virus Infection. Funct. Integr. Genom. 2014, 14, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, J.A. The Immune Response of Drosophila. Nature 2003, 426, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Dimopoulos, G. The Toll Immune Signaling Pathway Control Conserved Anti-Dengue Defenses across Diverse Ae. aegypti Strains and against Multiple Dengue Virus Serotypes. Dev. Comp. Immunol. 2010, 34, 625–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti Toll Pathway Controls Dengue Virus Infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef]
- Sanders, H.R.; Foy, B.D.; Evans, A.M.; Ross, L.S.; Beaty, B.J.; Olson, K.E.; Gill, S.S. Sindbis Virus Induces Transport Processes and Alters Expression of Innate Immunity Pathway Genes in the Midgut of the Disease Vector, Aedes aegypti. Insect Biochem. Mol. Biol. 2005, 35, 1293–1307. [Google Scholar] [CrossRef]
- Costa, A.; Jan, E.; Sarnow, P.; Schneider, D. The Imd Pathway Is Involved in Antiviral Immune Responses in Drosophila. PLoS ONE 2009, 4, e7436. [Google Scholar] [CrossRef] [Green Version]
- Oberst, A.; Bender, C.; Green, D.R. Living with Death: The Evolution of the Mitochondrial Pathway of Apoptosis in Animals. Cell Death Differ. 2008, 15, 1139–1146. [Google Scholar] [CrossRef] [Green Version]
- Vaux, D.L.; Korsmeyer, S.J. Cell Death in Development. Cell 1999, 96, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Rudin, C.M.; Thompson, C.B. APOPTOSIS AND DISEASE: Regulation and Clinical Relevance of Programmed Cell Death. Annu. Rev. Med. 1997, 48, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Clouston, W.M.; Kerr, J.F.R. Apoptosis, Lymphocytotoxicity and the Containment of Viral Infections. Med. Hypotheses 1985, 18, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Clem, R.J.; Miller, L.K. Apoptosis Reduces Both the in Vitro Replication and the in Vivo Infectivity of a Baculovirus. J. Virol. 1993, 67, 3730–3738. [Google Scholar] [CrossRef] [Green Version]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Programmed Cell Death (Apoptosis). In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Crook, N.E.; Clem, R.J.; Miller, L.K. An Apoptosis-Inhibiting Baculovirus Gene with a Zinc Finger-like Motif. J. Virol. 1993, 67, 2168–2174. [Google Scholar] [CrossRef] [Green Version]
- Colin, S.D.; Nava, V.E.; Gedrich, R.W.; Clem, R.J.; Van Dongen, J.L.; Gilfillan, M.C.; Shiels, H.; Hardwick, J.M.; Thompson, C.B. A Conserved Family of Cellular Genes Related to the Baculovirus Iap Gene and Encoding Apoptosis Inhibitors. EMBO J. 1996, 15, 2685–2694. [Google Scholar] [CrossRef]
- Hay, B.A.; Wassarman, D.A.; Rubin, G.M. Drosophila Homologs of Baculovirus Inhibitor of Apoptosis Proteins Function to Block Cell Death. Cell 1995, 83, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Fraser, A.G.; James, C.; Evan, G.I.; Hengartner, M.O. Caenorhabditis Elegans Inhibitor of Apoptosis Protein (IAP) Homologue BIR-1 Plays a Conserved Role in Cytokinesis. Curr. Biol. 1999, 9, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Bryant, B.; Blair, C.D.; Olson, K.E.; Clem, R.J. Annotation and Expression Profiling of Apoptosis-Related Genes in the Yellow Fever Mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 2008, 38, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Steller, H. Regulation of Apoptosis in Drosophila. Cell Death Differ. 2008, 15, 1132–1138. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, R.M.; Kriventseva, E.V.; Meister, S.; Xi, Z.; Alvarez, K.S.; Bartholomay, L.C.; Barillas-Mury, C.; Bian, G.; Blandin, S.; Christensen, B.M.; et al. Evolutionary Dynamics of Immune-Related Genes and Pathways in Disease-Vector Mosquitoes. Science 2007, 316, 1738–1743. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Clem, R.J. Defining the Core Apoptosis Pathway in the Mosquito Disease Vector Aedes aegypti: The Roles of Iap1, Ark, Dronc, and Effector Caspases. Apoptosis 2011, 16, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Pridgeon, J.W.; Zhao, L.; Becnel, J.J.; Strickman, D.A.; Clark, G.G.; Linthicum, K.J. Topically Applied AaeIAP1 Double-Stranded RNA Kills Female Adults of Aedes aegypti. J. Med. Entomol. 2014, 45, 414–420. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Clem, R.J. The Role of IAP Antagonist Proteins in the Core Apoptosis Pathway of the Mosquito Disease Vector Aedes aegypti. Apoptosis 2011, 16, 235–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Jiang, G.; Chan, G.; Santos, C.P.; Severson, D.W.; Xiao, L. Michelob_x Is the Missing Inhibitor of Apoptosis Protein Antagonist in Mosquito Genomes. EMBO Rep. 2005, 6, 769–774. [Google Scholar] [CrossRef]
- Vaidyanathan, R.; Scott, T.W. Apoptosis in Mosquito Midgut Epithelia Associated with West Nile Virus Infection. Apoptosis 2006, 11, 1643–1651. [Google Scholar] [CrossRef]
- Ocampo, C.B.; Caicedo, P.A.; Jaramillo, G.; Ursic Bedoya, R.; Baron, O.; Serrato, I.M.; Cooper, D.M.; Lowenberger, C. Differential Expression of Apoptosis Related Genes in Selected Strains of Aedes aegypti with Different Susceptibilities to Dengue Virus. PLoS ONE 2013, 8, e61187. [Google Scholar] [CrossRef]
- Serrato, I.M.; Caicedo, P.A.; Orobio, Y.; Lowenberger, C.; Ocampo, C.B. Vector Competence and Innate Immune Responses to Dengue Virus Infection in Selected Laboratory and Field-Collected Stegomyia Aegypti (=Aedes aegypti). Med. Vet. Entomol. 2017, 31, 312–319. [Google Scholar] [CrossRef]
- O’Neill, K.; Huang, N.; Unis, D.; Clem, R.J. Rapid Selection against Arbovirus-Induced Apoptosis during Infection of a Mosquito Vector. Proc. Natl. Acad. Sci. USA 2015, 112, E1152–E1161. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, A.; Santos, S.R.; Clem, R.J. Expressing the Pro-Apoptotic Reaper Protein via Insertion into the Structural Open Reading Frame of Sindbis Virus Reduces the Ability to Infect Aedes aegypti Mosquitoes. Viruses 2022, 14, 2035. [Google Scholar] [CrossRef]
- Wang, H.; Gort, T.; Boyle, D.L.; Clem, R.J. Effects of Manipulating Apoptosis on Sindbis Virus Infection of Aedes aegypti Mosquitoes. J. Virol. 2012, 86, 6546–6554. [Google Scholar] [CrossRef] [Green Version]
- Eng, M.W.; van Zuylen, M.N.; Severson, D.W. Apoptosis-Related Genes Control Autophagy and Influence DENV-2 Infection in the Mosquito Vector, Aedes aegypti. Insect Biochem. Mol. Biol. 2016, 76, 70–83. [Google Scholar] [CrossRef]
- Taracena, M.L.; Bottino-Rojas, V.; Talyuli, O.A.C.; Walter-Nuno, A.B.; Oliveira, J.H.M.; Angleró-Rodriguez, Y.I.; Wells, M.B.; Dimopoulos, G.; Oliveira, P.L.; Paiva-Silva, G.O. Regulation of Midgut Cell Proliferation Impacts Aedes aegypti Susceptibility to Dengue Virus. PLoS Negl. Trop. Dis. 2018, 12, e0006498. [Google Scholar] [CrossRef] [Green Version]
- Scott, T.W.; Clark, G.G.; Lorenz, L.H.; Amerasinghe, P.H.; Reiter, P.; Edman, J.D. Detection of Multiple Blood Feeding in Aedes aegypti (Diptera: Culicidae) During a Single Gonotrophic Cycle Using a Histologic Technique. J. Med. Entomol. 1993, 30, 94–99. [Google Scholar] [CrossRef]
- Boorman, J. Observations on the Amount of Virus Present in the Haemolymph of Aedes aegypti Infected with Uganda S, Yellow Fever and Semliki Forest Viruses. Trans. R. Soc. Trop. Med. Hyg. 1960, 54, 362–365. [Google Scholar] [CrossRef]
- Miles, J.A.R.; Pillai, J.S.; Maguire, T. Multiplication of Whataroa Virus in Mosquitoes. J. Med. Entomol. 1973, 10, 176–185. [Google Scholar] [CrossRef]
- Weaver, S.C.; Scott, T.W.; Lorenz, L.H.; Repik, P.M. Detection of Eastern Equine Encephalomyelitis Virus Deposition in Culiseta Melanura Following Ingestion of Radiolabeled Virus in Blood Meals. Am. J. Trop. Med. Hyg. 1991, 44, 250–259. [Google Scholar] [CrossRef]
- Paquette, C.C.H.; Phanse, Y.; Perry, J.L.; Sanchez-Vargas, I.; Airs, P.M.; Dunphy, B.M.; Xu, J.; Carlson, J.O.; Luft, J.C.; DeSimone, J.M.; et al. Biodistribution and Trafficking of Hydrogel Nanoparticles in Adult Mosquitoes. PLoS Negl. Trop. Dis. 2015, 9, e0003745. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, A.; Bryant, W.B.; Santos, S.R.; Clem, R.J. Infection of Aedes aegypti Mosquitoes with Midgut-Attenuated Sindbis Virus Reduces, but Does Not Eliminate, Disseminated Infection. J. Virol. 2021, 95, 136–157. [Google Scholar] [CrossRef]
- Vazeille, M.; Madec, Y.; Mousson, L.; Bellone, R.; Barré-Cardi, H.; Sousa, C.A.; Jiolle, D.; Yébakima, A.; de Lamballerie, X.; Failloux, A.B. Zika Virus Threshold Determines Transmission by European Aedes albopictus Mosquitoes. Emerg. Microbes Infect. 2019, 8, 1668–1678. [Google Scholar] [CrossRef] [Green Version]
- Bosio, C.F.; Beaty, B.J.; Black IV, W.C. Quantitative Genetics of Vector Competence for Dengue-2 Virus in Aedes aegypti. Am. J. Trop. Med. Hyg. 1998, 59, 965–970. [Google Scholar] [CrossRef] [Green Version]
- Dickson, L.B.; Sanchez-Vargas, I.; Sylla, M.; Fleming, K.; Black, W.C. Vector Competence in West African Aedes aegypti Is Flavivirus Species and Genotype Dependent. PLoS Negl. Trop. Dis. 2014, 8, e3153. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Adams, A.P.; Kenney, J.L.; Wang, E.; Weaver, S.C. Venezuelan Equine Encephalitis Virus in the Mosquito Vector Aedes Taeniorhynchus: Infection Initiated by a Small Number of Susceptible Epithelial Cells and a Population Bottleneck. Virology 2008, 372, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Forrester, N.L.; Guerbois, M.; Seymour, R.L.; Spratt, H.; Weaver, S.C. Vector-Borne Transmission Imposes a Severe Bottleneck on an RNA Virus Population. PLoS Pathog. 2012, 8, e1002897. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Ehrbar, D.J.; Van Slyke, G.A.; Payne, A.F.; Willsey, G.G.; Viscio, R.E.; Kramer, L.D. Quantification of Intrahost Bottlenecks of West Nile Virus in Culex pipiens Mosquitoes Using an Artificial Mutant Swarm. Infect. Genet. Evol. 2012, 12, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Drake, J.W.; Holland, J.J. Mutation Rates among RNA Viruses. Proc. Natl. Acad. Sci. USA 1999, 96, 13910–13913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffey, L.L.; Beeharry, Y.; Bordería, A.V.; Blanc, H.; Vignuzzi, M. Arbovirus High Fidelity Variant Loses Fitness in Mosquitoes and Mice. Proc. Natl. Acad. Sci. USA 2011, 108, 16038–16043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warmbrod, K.L.; Patterson, E.I.; Kautz, T.F.; Stanton, A.; Rockx-Brouwer, D.; Kalveram, B.K.; Khanipov, K.; Thangamani, S.; Fofanov, Y.; Forrester, N.L. Viral RNA-Dependent RNA Polymerase Mutants Display an Altered Mutation Spectrum Resulting in Attenuation in Both Mosquito and Vertebrate Hosts. PLoS Pathog. 2019, 15, e1007610. [Google Scholar] [CrossRef]
- Ciota, A.T.; Lovelace, A.O.; Jones, S.A.; Payne, A.; Kramer, L.D. Adaptation of Two Flaviviruses Results in Differences in Genetic Heterogeneity and Virus Adaptability. J. Gen. Virol. 2007, 88 Pt 9, 2398. [Google Scholar] [CrossRef]
- Simonsen, P.E.; Fischer, P.U.; Hoerauf, A.; Weil, G.J. The Filariases. In Manson’s Tropical Diseases, 23rd ed.; W.B. Saunders: Philadelphia, PA, USA, 2013; pp. 737–765. [Google Scholar] [CrossRef]
- Turell, M.J.; Rossignol, P.A.; Spielman, A.; Rossi, C.A.; Bailey, C.L. Enhanced Arboviral Transmission by Mosquitoes That Concurrently Ingested Microfilariae. Science 1984, 225, 1039–1041. [Google Scholar] [CrossRef]
- Zytoon, E.M.; El-Belbasi, H.I.; Matsumura, T. Mechanism of Increased Dissemination of Chikungunya Virus in Aedes albopictus Mosquitoes Concurrently Ingesting Microfilariae of Dirofilaria Immitis. Am. J. Trop. Med. Hyg. 1993, 49, 201–207. [Google Scholar] [CrossRef]
- Turell, M.J.; Mather, T.N.; Spielman, A.; Bailey, C.L. Increased Dissemination of Dengue 2 Virus in Aedes aegypti Associated with Concurrent Ingestion of Microfilariae of Brugia Malayi. Am. J. Trop. Med. Hyg. 1987, 37, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.A.; Turell, M.J. Brugia Malayi Microfilariae Transport Alphaviruses across the Mosquito Midgut. PLoS ONE 2017, 12, e0172309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of Simultaneous Exposure to Arboviruses on Infection and Transmission by Aedes aegypti Mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göertz, G.P.; Vogels, C.B.F.; Geertsema, C.; Koenraadt, C.J.M.; Pijlman, G.P. Mosquito Co-Infection with Zika and Chikungunya Virus Allows Simultaneous Transmission without Affecting Vector Competence of Aedes aegypti. PLoS Negl. Trop. Dis. 2017, 11, e0005654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhaes, T.; Robison, A.; Young, M.C.; Black, W.C.; Foy, B.D.; Ebel, G.D.; Rückert, C. Sequential Infection of Aedes aegypti Mosquitoes with Chikungunya Virus and Zika Virus Enhances Early Zika Virus Transmission. Insects 2018, 9, 177. [Google Scholar] [CrossRef] [Green Version]
- Muturi, E.J.; Bara, J. Sindbis Virus Interferes with Dengue 4 Virus Replication and Its Potential Transmission by Aedes albopictus. Parasites Vectors 2015, 8, 65. [Google Scholar] [CrossRef] [Green Version]
- Baidaliuk, A.; Miot, E.F.; Lequime, S.; Moltini-Conclois, I.; Delaigue, F.; Dabo, S.; Dickson, L.B.; Aubry, F.; Merkling, S.H.; Cao-Lormeau, V.-M.; et al. Cell-Fusing Agent Virus Reduces Arbovirus Dissemination in Aedes aegypti Mosquitoes In Vivo. J. Virol. 2019, 93, e00705-19. [Google Scholar] [CrossRef] [Green Version]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission Dynamics of an Insect-Specific Flavivirus in a Naturally Infected Culex pipiens Laboratory Colony and Effects of Co-Infection on Vector Competence for West Nile Virus. Virology 2012, 427, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, F.; Chiles, R.E.; Fang, Y.; Green, E.N.; Reisen, W.K. Effects of Time after Infection, Mosquito Genotype, and Infectious Viral Dose on the Dynamics of Culex Tarsalis Vector Competence for Western Equine Encephalomyelitis Virus. J. Am. Mosq. Control Assoc. 2006, 22, 272–281. [Google Scholar] [CrossRef]
- Tesla, B.; Demakovsky, L.R.; Packiam, H.S.; Mordecai, E.A.; Rodríguez, A.D.; Bonds, M.H.; Brindley, M.A.; Murdock, C.C. Estimating the Effects of Variation in Viremia on Mosquito Susceptibility, Infectiousness, and R0 of Zika in Aedes aegypti. PLoS Negl. Trop. Dis. 2018, 12, e0006733. [Google Scholar] [CrossRef]
- Merwaiss, F.; Filomatori, C.V.; Susuki, Y.; Bardossy, E.S.; Alvarez, D.E.; Saleh, M.-C. Chikungunya Virus Replication Rate Determines the Capacity of Crossing Tissue Barriers in Mosquitoes. J. Virol. 2021, 95, e01956-20. [Google Scholar] [CrossRef]


{kind=link}
| Virus | Vector | Reference |
|---|---|---|
| La Crosse virus | Ae. triseriatus | [28] |
| Jamestown Canyon virus | Ae. provocans | [31] |
| Rift Valley fever virus | Ae. vexans Ae. increpitus Ae. melanimon Cx. Antennatus Cx. Pipiens Cx. quinquefasciatus | [29,33] |
| dengue virus | Ae. aegypti | [30,32] |
| Western equine encephalitis virus | Culex tarsalis | [34] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carpenter, A.; Clem, R.J. Factors Affecting Arbovirus Midgut Escape in Mosquitoes. Pathogens 2023, 12, 220. https://doi.org/10.3390/pathogens12020220
Carpenter A, Clem RJ. Factors Affecting Arbovirus Midgut Escape in Mosquitoes. Pathogens. 2023; 12(2):220. https://doi.org/10.3390/pathogens12020220
Chicago/Turabian StyleCarpenter, Alexis, and Rollie J. Clem. 2023. "Factors Affecting Arbovirus Midgut Escape in Mosquitoes" Pathogens 12, no. 2: 220. https://doi.org/10.3390/pathogens12020220