
Occurrence of Escherichia coli Pathotypes in Diarrheic Calves in a Low-Income Setting
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
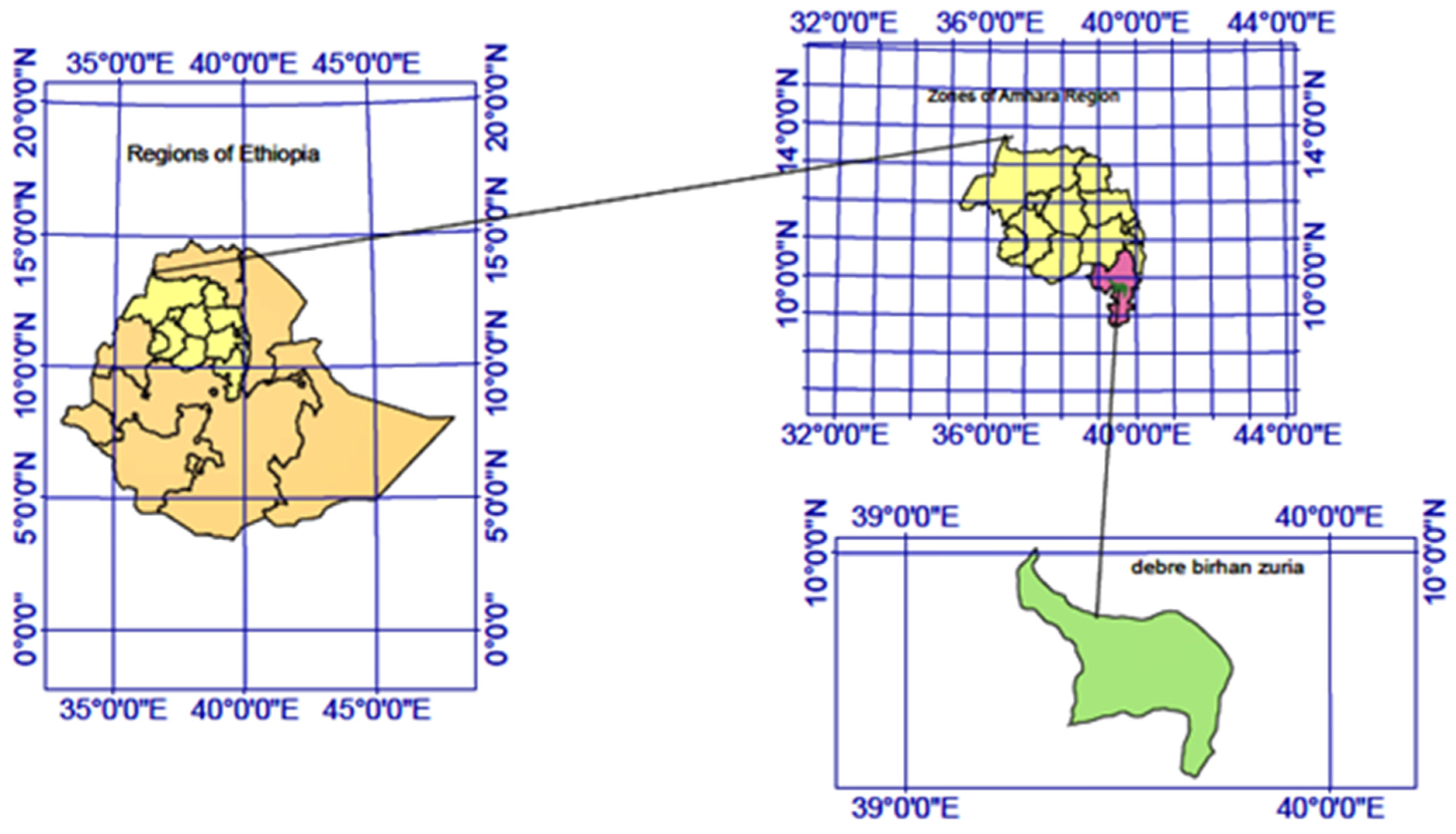
2.1. Study Area and Period
2.2. Study Design and Sampling Technique
2.3. Isolation and Characterization of E. coli
2.4. DNA Extraction
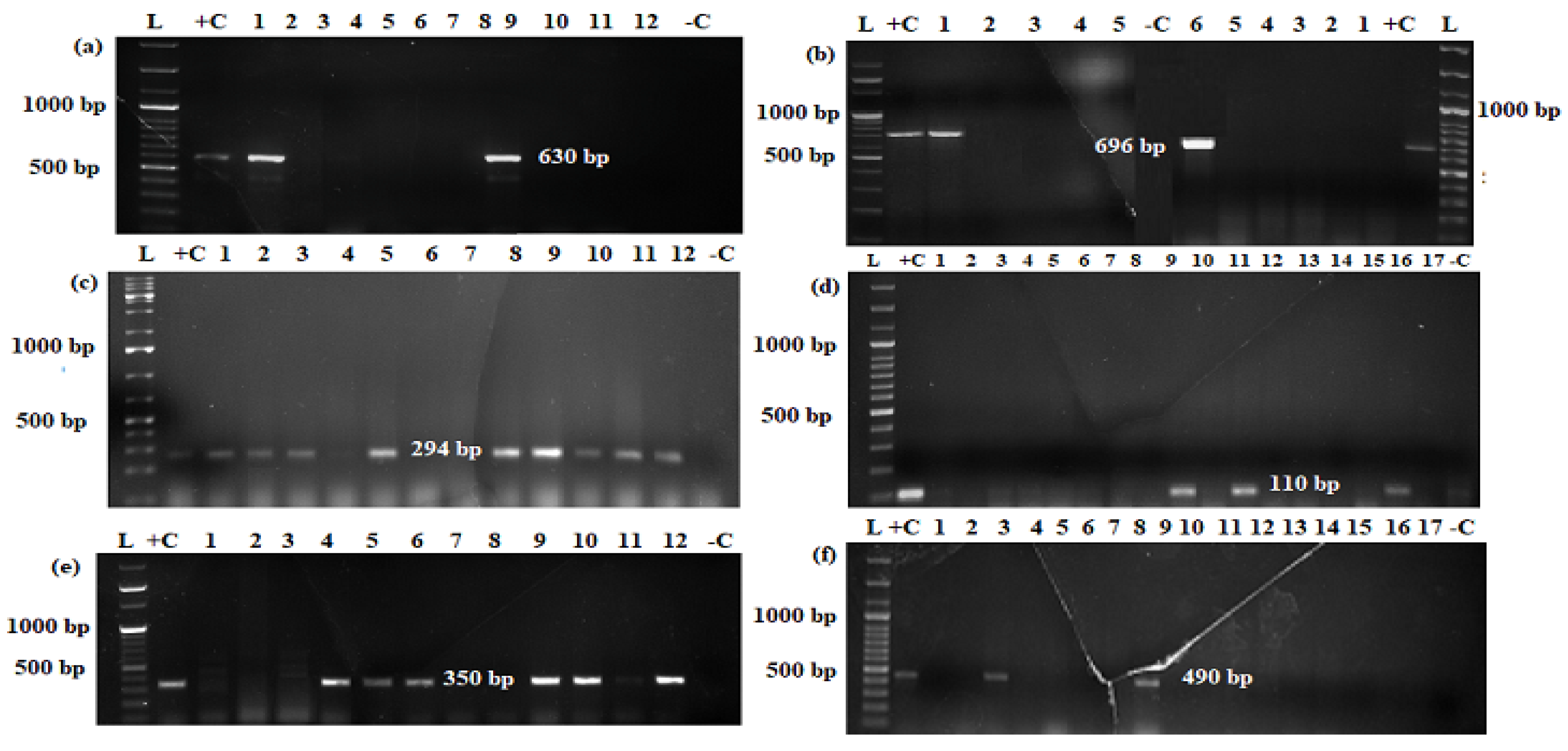
2.5. E. coli Pathotyping Using PCR
2.6. Farm Data
2.7. Data Processing and Analysis
3. Results
3.1. Farm Description and Calf Management
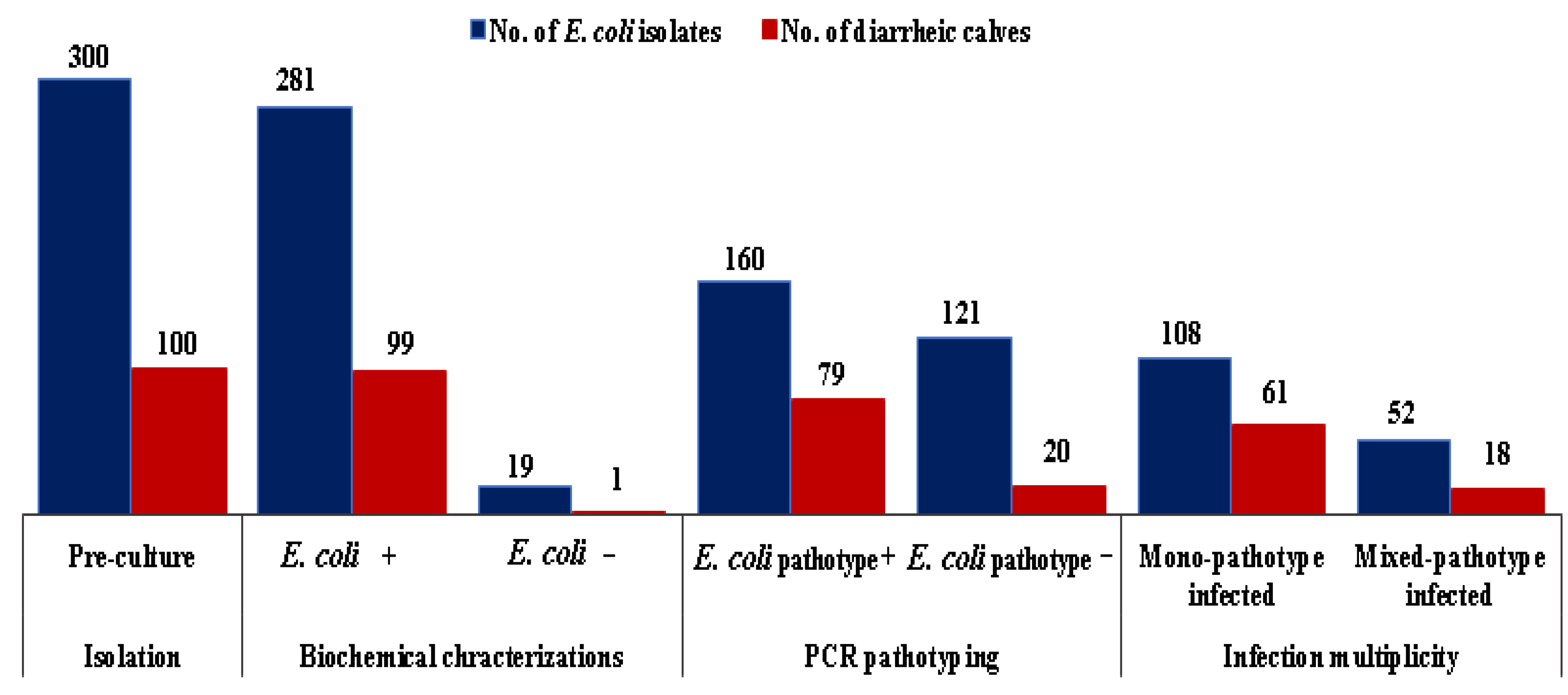
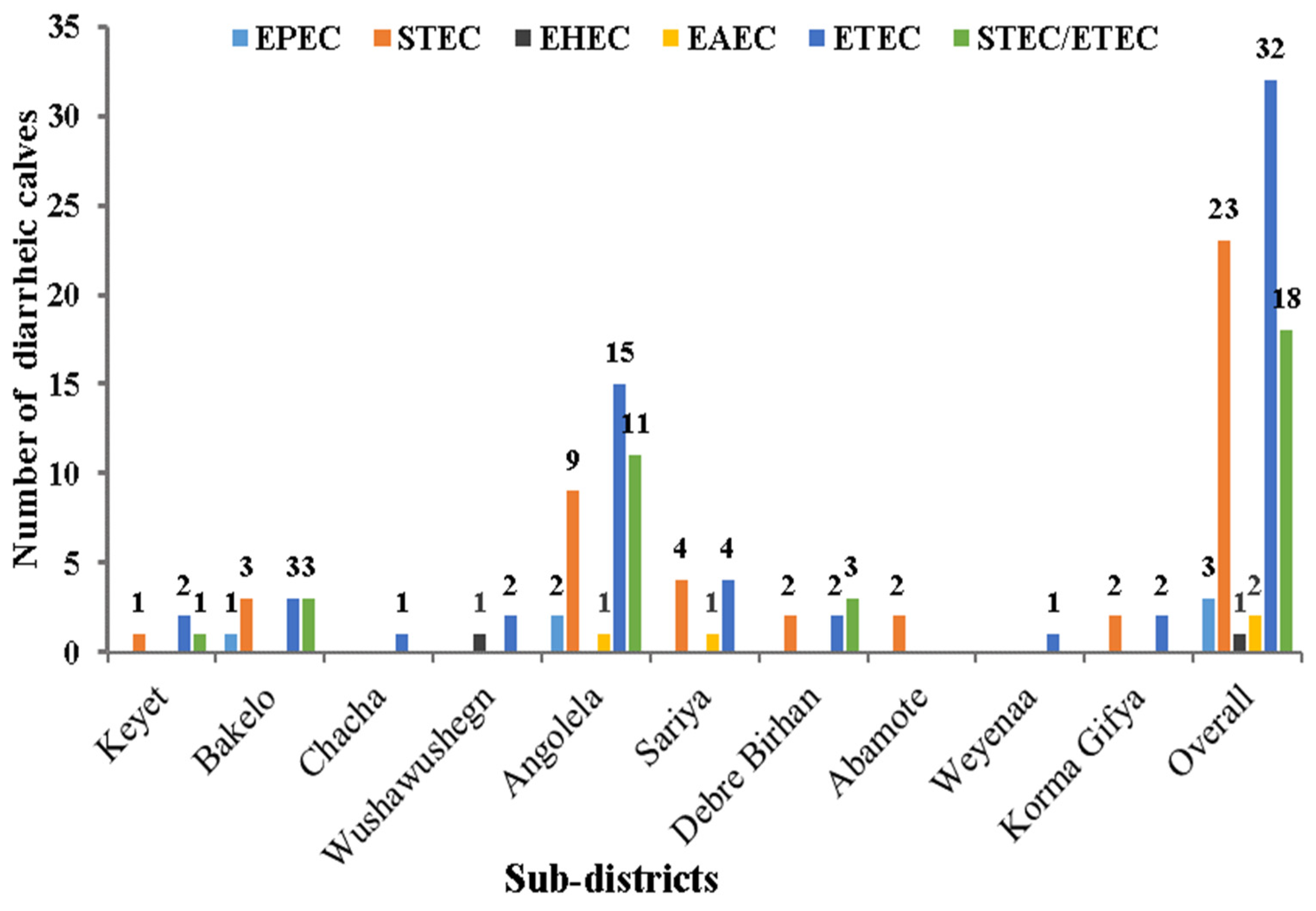
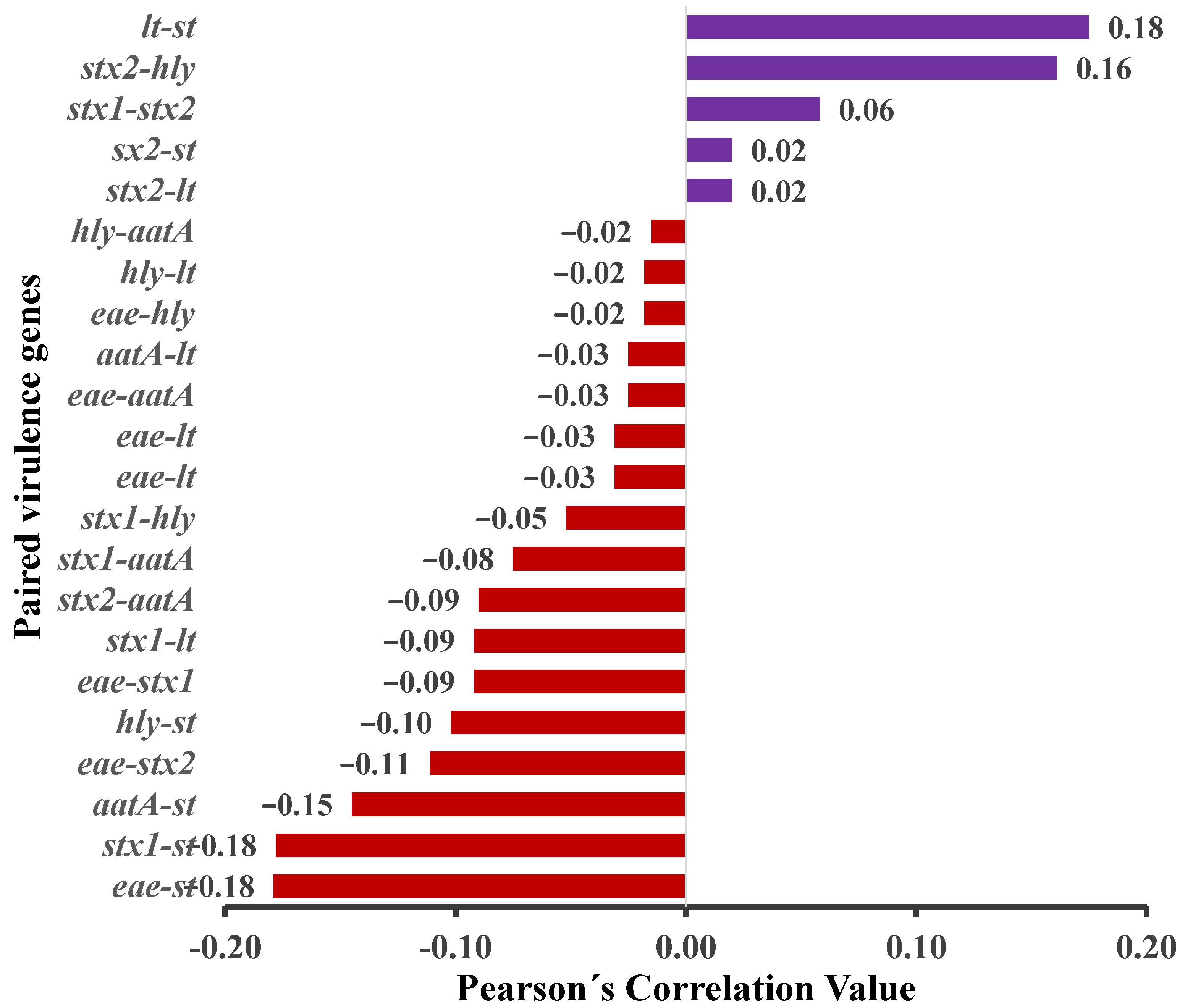
3.2. E. coli Isolation, Virulence Genes (VGs) Detection, and E. coli Pathotyping
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lugsomya, K.; Yindee, J.; Niyomtham, W.; Tribuddharat, C.; Tummaruk, P.; Hampson, D.J.; Prapasarakul, N. Antimicrobial Resistance in Commensal Escherichia Coli Isolated from Pigs and Pork Derived from Farms Either Routinely Using or Not Using In-Feed Antimicrobials. Microb. Drug Resist. 2018, 24, 1054–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angappan, M.; Ghatak, S.; Milton, A.A.P.; Verma, A.K.; Inbaraj, S.; Abhishek; Chaudhuri, P.; Agarwal, R.K.; Thomas, P. Detection of Novel Sequence Types and Zoonotic Transmission Potentiality among Strains of Shiga Toxigenic Escherichia Coli (STEC) from Dairy Calves, Animal Handlers and Associated Environments. Brazilian J. Microbiol. 2021, 52, 2541–2546. [Google Scholar] [CrossRef] [PubMed]
- Engelen, F.; Thiry, D.; Devleesschauwer, B.; Heyndrickx, M.; Mainil, J.; De Zutter, L.; Cox, E. Pathogenic Potential of Escherichia Coli O157 and O26 Isolated from Young Belgian Dairy Calves by Recto-Anal Mucosal Swab Culturing. J. Appl. Microbiol. 2021, 131, 964–972. [Google Scholar] [CrossRef]
- Pakbin, B.; Brück, W.M.; Rossen, J.W.A. Virulence Factors of Enteric Pathogenic Escherichia Coli: A Review. Int. J. Mol. Sci. 2021, 22, 9922. [Google Scholar] [CrossRef]
- World Health Organization. Who Estimates of the Global Burden of Foodborne Diseases 2007–2015. In FOODBORNE Disease Burden Epidemiology Reference Group 2007–2015; WHO: Geneva, Switzerland, 2015; p. 72. [Google Scholar]
- Abuelo, A. Investigation of an Outbreak of Neonatal Calf Diarrhoea in a Dairy Herd. Vet. Rec. Case Rep. 2016, 4, e000372. [Google Scholar] [CrossRef]
- Kayasaki, F.; Okagawa, T.; Konnai, S.; Kohara, J.; Sajiki, Y.; Watari, K.; Ganbaatar, O.; Goto, S.; Nakamura, H.; Shimakura, H.; et al. Direct Evidence of the Preventive Effect of Milk Replacer–Based Probiotic Feeding in Calves against Severe Diarrhea. Vet. Microbiol. 2021, 254, 108976. [Google Scholar] [CrossRef]
- Tora, E.; Abayneh, E.; Seyoum, W.; Shurbe, M. Longitudinal Study of Calf Morbidity and Mortality on Smallholder Farms in Southern Ethiopia. PLoS ONE 2021, 16, e0257139. [Google Scholar] [CrossRef]
- Caffarena, R.D.; Casaux, M.L.; Schild, C.O.; Fraga, M.; Castells, M.; Colina, R.; Maya, L.; Corbellini, L.G.; Riet-Correa, F.; Giannitti, F. Causes of Neonatal Calf Diarrhea and Mortality in Pasture-Based Dairy Herds in Uruguay: A Farm-Matched Case-Control Study. Brazilian J. Microbiol. 2021, 52, 977–988. [Google Scholar] [CrossRef]
- Ferede, Y.; Mazengia, H.; Bimrew, T.; Bitew, A.; Nega, M.; Kebede, A. Pre-Weaning Morbidity and Mortality of Crossbred Calves in Bahir Dar Zuria and Gozamen Districts of Amhara Region, Northwest Ethiopia. OALib 2014, 1, e600. [Google Scholar] [CrossRef]
- Fentie, T.; Guta, S.; Mekonen, G.; Temesgen, W.; Melaku, A.; Asefa, G.; Tesfaye, S.; Niguse, A.; Abera, B.; Kflewahd, F.Z.; et al. Assessment of Major Causes of Calf Mortality in Urban and Periurban Dairy Production System of Ethiopia. Vet. Med. Int. 2020, 2020, 3075429. [Google Scholar] [CrossRef]
- United States Department of Agriculture. Dairy 2007 Part II: Changes in the U.S. Dairy Cattle Industry, 1991–2007. In Fort Collins: USDA-APHIS-VS, CEAH; USDA: Washington, DC, USA, 2008; pp. 57–61. [Google Scholar]
- Schinwald, M.; Creutzinger, K.; Keunen, A.; Winder, C.B.; Haley, D.; Renaud, D.L. Predictors of Diarrhea, Mortality, and Weight Gain in Male Dairy Calves. J. Dairy Sci. 2022, 105, 5296–5309. [Google Scholar] [CrossRef] [PubMed]
- Østerås, O.; Gjestvang, M.S.; Vatn, S.; Sølverød, L. Perinatal Death in Production Animals in the Nordic Countries—Incidence and Costs. Acta Vet. Scand. 2007, 49, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Fesseha, H.; Mathewos, M.; Aliye, S.; Mekonnen, E. Isolation and Antibiogram of Escherichia Coli O157: H7 from Diarrhoeic Calves in Urban and Peri-Urban Dairy Farms of Hawassa Town. Vet. Med. Sci. 2022, 8, 864–876. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Choi, C.Y.; Li, G.; Li, H.; Shi, Z. Pre-Weaned Dairy Calf Management Practices, Morbidity and Mortality of Bovine Respiratory Disease and Diarrhea in China. Livest. Sci. 2021, 251, 104608. [Google Scholar] [CrossRef]
- Endale, A.; Muktar, Y.; Tessema, T.S. Molecular Characterization of Diarrheagenic Escherichia Coli Strains Isolated from Dairy Calves in North Shoa Zone, Oromiya Region, Ethiopia. Res. Sq. 2020, 17, e0275229. [Google Scholar]
- Negeri, A.A.; Mamo, H.; Gurung, J.M.; Firoj Mahmud, A.K.M.; Fällman, M.; Seyoum, E.T.; Feleke Desta, A.; Francis, M.S. Antimicrobial Resistance Profiling and Molecular Epidemiological Analysis of Extended Spectrum β-Lactamases Produced by Extraintestinal Invasive Escherichia Coli Isolates from Ethiopia: The Presence of International High-Risk Clones ST131 and ST410 Reveal. Front. Microbiol. 2021, 12, 706846. [Google Scholar] [CrossRef]
- Ayenew, H.Y.; Mitiku, B.A.; Tesema, T.S. Occurrence of Virulence Genes and Antimicrobial Resistance of E. Coli O157:H7 Isolated from the Beef Carcass of Bahir Dar City, Ethiopia. Vet. Med. Int. 2021, 2021, 8046680. [Google Scholar] [CrossRef]
- Sebre, S.; Erku Abegaz, W.; Seman, A.; Awoke, T.; Mihret, W.; Desalegn, Z.; Abebe, T.; Mihret, A. Molecular Characterization of Extended-Spectrum Beta-Lactamase-Producing Enterobacteriaceae Isolates Collected from Inanimate Hospital Environments in Addis Ababa, Ethiopia. In Advances in Microbiology, Infectious Disease and Public Health; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Central Statistical Agency Central Statistical Agency (2015) Agricultural Sample Survey 2014/15. Report on Livestock and Livestock Characteristics. Stat. Bull. 2015, 2, 578. [Google Scholar]
- Central Statistical Agency. Statistical Report for Amhara Region; CSA: Addis Ababa, Ethiopia, 2007. [Google Scholar]
- Cheesbrough, M. Medical Laboratory Manual for Tropical Countries; Tropical Health Technology: London, UK, 1985; Volume II, ISBN 0950743429. [Google Scholar]
- Moore, E.; Arnscheidt, A.; KrÜger, A.; StrÖmpl, C.; Mau, M. Section 1 Update: Simplified Protocols for the Preparation of Genomic DNA from Bacterial Cultures. In Molecular Microbial Ecology Manual; Kluwer Academic: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Khan, A.; Yamasaki, S.; Sato, T.; Ramamurthy, T.; Pal, A.; Datta, S.; Chowdhury, N.R.; Das, S.C.; Sikdar, A.; Tsukamoto, T.; et al. Prevalence and Genetic Profiling of Virulence Determinants of Non-O157 Shiga Toxin-Producing Escherichia Coli Isolated from Cattle, Beef, and Humans, Calcutta, India. Emerg. Infect. Dis. 2002, 8, 54–62. [Google Scholar] [CrossRef]
- Pal, A.; Ghosh, S.; Ramamurthy, T.; Yamasaki, S.; Tsukamoto, T.; Bhattacharya, S.K.; Takeda, Y. Shiga-Toxin Producing Escherichia Coli from Healthy Cattle in a Semi-Urban Community in Calcutta, India. Indian J. Med. Res. 1999, 110, 83. [Google Scholar]
- Selim, S.A.; Ahmed, S.F.; Abdel Aziz, M.H.; Zakaria, A.M.; Klena, J.D.; Pangallo, D. Prevalence and Characterization of Shiga-Toxin O157:H7 and Non-O157:H7 Enterohemorrhagic Escherichia Coli Isolated from Different Sources. Biotechnol. Biotechnol. Equip. 2013, 27, 3834–3842. [Google Scholar] [CrossRef] [Green Version]
- Havt, A.; Lima, I.F.; Medeiros, P.H.; Clementino, M.A.; Santos, A.K.; Amaral, M.S.; Veras, H.N.; Prata, M.M.; Lima, N.L.; Di Moura, A.; et al. Prevalence and Virulence Gene Profiling of Enteroaggregative Escherichia Coli in Malnourished and Nourished Brazilian Children. Diagn. Microbiol. Infect. Dis. 2017, 89, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Gunzburg, S.T.; Tornieporth, N.G.; Riley, L.W. Identification of Enteropathogenic Escherichia Coli by PCR-Based Detection of the Bundle-Forming Pilus Gene. J. Clin. Microbiol. 1995, 33, 1375–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nada, R.A.; Shaheen, H.I.; Touni, I.; Fahmy, D.; Armstrong, A.W.; Weiner, M.; Klena, J.D. Design and Validation of a Multiplex Polymerase Chain Reaction for the Identification of Enterotoxigenic Escherichia Coli and Associated Colonization Factor Antigens. Diagn. Microbiol. Infect. Dis. 2010, 67, 134–142. [Google Scholar] [CrossRef]
- Schultsz, C.; Pool, G.J.; Van Ketel, R.; De Wever, B.; Speelman, P.; Dankert, J. Detection of Enterotoxigenic Escherichia Coli in Stool Samples by Using Nonradioactively Labeled Oligonucleotide DNA Probes and PCR. J. Clin. Microbiol. 1994, 32, 2393–2397. [Google Scholar] [CrossRef] [Green Version]
- Batista, C.; Carolina, M.; Souza, S.D.; Serra, P.T.; Santos, I.; Balieiro, A.; Pieri, F.A.; Nogueira, P.A.; Orlandi, P.P. Virulence Factors Associated with Pediatric Shigellosis in Brazilian Amazon. Biomed. Res. Int. 2014, 2014, 539697. [Google Scholar] [CrossRef] [Green Version]
- Vidal, M.; Kruger, E.; Durán, C.; Lagos, R.; Levine, M.; Prado, V.; Toro, C.; Vidal, R. Single Multiplex PCR Assay to Identify Simultaneously the Six Categories of Diarrheagenic Escherichia Coli Associated with Enteric Infections. J. Clin. Microbiol. 2005, 43, 5362–5365. [Google Scholar] [CrossRef] [Green Version]
- Umpiérrez, A.; Ernst, D.; Fernández, M.; Oliver, M.; Casaux, M.L.; Caffarena, R.D.; Schild, C.; Giannitti, F.; Fraga, M.; Zunino, P. Virulence Genes of Escherichia Coli in Diarrheic and Healthy Calves. Rev. Argent. Microbiol. 2021, 53, 34–38. [Google Scholar] [CrossRef]
- Ryu, J.H.; Kim, S.H.; Park, J.; Choi, K.S. Characterization of Virulence Genes in Escherichia Coli Strains Isolated from Pre-Weaned Calves in the Republic of Korea. Acta Vet. Scand. 2020, 62, 45. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, W.; Cao, M.; Zhu, Q.; Ma, T.; Zhang, Y.; Liu, G.; Zhou, X.; Li, B.; Shi, Y.; et al. Virulence Factors and Molecular Characteristics of Shigella Flexneri Isolated from Calves with Diarrhea. BMC Microbiol. 2021, 21, 214. [Google Scholar] [CrossRef]
- Dall Agnol, A.M.; Lorenzetti, E.; Leme, R.A.; Ladeia, W.A.; Mainardi, R.M.; Bernardi, A.; Headley, S.A.; Freire, R.L.; Pereira, U.P.; Alfieri, A.F.; et al. Severe Outbreak of Bovine Neonatal Diarrhea in a Dairy Calf Rearing Unit with Multifactorial Etiology. Brazilian J. Microbiol. 2021, 52, 2547–2553. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Luo, Q.; Davis, S.M.; Westra, C.; Vickers, T.J. Molecular Determinants of Enterotoxigenic Escherichia Coli Heat-Stable Toxin Secretion and Delivery. ASM J. 2018, 86, e00526-18. [Google Scholar] [CrossRef] [PubMed]
- Von Mentzer, A.; Connor, T.R.; Wieler, L.H.; Semmler, T.; Iguchi, A.; Thomson, N.R.; Rasko, D.A.; Joffre, E.; Corander, J.; Pickard, D.; et al. Identification of Enterotoxigenic Escherichia Coli (ETEC) Clades with Long-Term Global Distribution. Nat. Genet. 2014, 46, 1321–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepard, S.M.; Danzeisen, J.L.; Isaacson, R.E.; Seemann, T.; Achtman, M.; Johnson, T.J. Genome Sequences and Phylogenetic Analysis of K88- and F18- Positive Porcine Enterotoxigenic Escherichia Coli. J. Bacteriol. 2012, 194, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Awad, W.S.; El-Sayed, A.A.; Mohammed, F.F.; Bakry, N.M.; Abdou, N.E.M.I.; Kamel, M.S. Molecular Characterization of Pathogenic Escherichia Coli Isolated from Diarrheic and In-Contact Cattle and Buffalo Calves. Trop. Anim. Health Prod. 2020, 52, 3173–3185. [Google Scholar] [CrossRef]
- Shabana, I.I.; Zaraket, H.; Suzuki, H. Molecular Studies on Diarrhea-Associated Escherichia Coli Isolated from Humans and Animals in Egypt. Vet. Microbiol. 2013, 167, 532–539. [Google Scholar] [CrossRef]
- Khawaskar, D.P.; Sinha, D.K.; Lalrinzuala, M.V.; Athira, V.; Kumar, M.; Chhakchhuak, L.; Mohanapriya, K.; Sophia, I.; Abhishek; Kumar, O.R.V.; et al. Pathotyping and Antimicrobial Susceptibility Testing of Escherichia Coli Isolates from Neonatal Calves. Vet. Res. Commun. 2021, 46, 353–362. [Google Scholar] [CrossRef]
- Leomil, L.; Aidar-Ugrinovich, L.; Guth, B.E.C.; Irino, K.; Vettorato, M.P.; Onuma, D.L.; De Castro, A.F.P. Frequency of Shiga Toxin-Producing Escherichia Coli (STEC) Isolates among Diarrheic and Non-Diarrheic Calves in Brazil. Vet. Microbiol. 2003, 97, 103–109. [Google Scholar] [CrossRef]
- Srivani, M.; Narasimha Reddy, Y.; Subramanyam, K.V.; Ramakoti Reddy, M.; Srinivasa Rao, T. Prevalence and Antimicrobial Resistance Pattern of Shiga Toxigenic Escherichia Coli in Diarrheic Buffalo Calves. Vet. World 2017, 10, 774–778. [Google Scholar] [CrossRef] [Green Version]
- Marashifard, M.; Aliabad, Z.K.; Ali, S.; Malek, A.; Darban-sarokhalil, D. Determination of Antibiotic Resistance Pattern and Virulence Genes in Escherichia Coli Isolated from Bovine with Subclinical Mastitis in Southwest of Iran. Trop. Anim. Health Prod. 2018, 51, 575–580. [Google Scholar] [CrossRef]
- Sobhy, N.M.; Yousef, S.G.A.; Aboubakr, H.A.; Nisar, M.; Nagaraja, K.V.; Mor, S.K.; Valeris-Chacin, R.J.; Goyal, S.M. Virulence Factors and Antibiograms of Escherichia Coli Isolated from Diarrheic Calves of Egyptian Cattle and Water Buffaloes. PLoS One 2020, 15, e0232890. [Google Scholar] [CrossRef] [PubMed]
- Coura, F.M.; Diniz, S.D.A.; Silva, M.X.; de Oliveira, C.H.S.; Mussi, J.M.S.; de Oliveira, C.S.F.; Lage, A.P.; Heinemann, M.B. Virulence Factors and Phylotyping of Escherichia Coli Isolated from Non-Diarrheic and Diarrheic Water Buffalo Calves. Cienc. Rural 2019, 49, 1–9. [Google Scholar] [CrossRef]
- Wani, S.A.; Bhat, M.A.; Samanta, I.; Nishikawa, Y.; Buchh, A.S. Isolation and Characterization of Shiga Toxin-Producing Escherichia Coli (STEC) and Enteropathogenic Escherichia Coli (EPEC) from Calves and Lambs with Diarrhoea in India. Lett. Appl. Microbiol. 2003, 37, 121–126. [Google Scholar] [CrossRef]
- Aref, N.E.M.; Abdel-Raheem, A.R.A.; Kamaly, H.F.; Hussien, S.Z. Clinical and Sero-Molecular Characterization of Escherichia Coli with an Emphasis on Hybrid Strain in Healthy and Diarrheic Neonatal Calves in Egypt. Open Vet. J. 2018, 8, 351–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Zhi, S.; Guo, D.; Jiang, Y.; Xu, X.; Zhao, L.; Lv, J. Prevalence, Antimicrobial Resistance, and Whole Genome Sequencing Analysis of Shiga Toxin-Producing Escherichia Coli (STEC) and Enteropathogenic Escherichia Coli (EPEC) from Imported Foods in China during 2015–2021. Toxins 2022, 14, 68. [Google Scholar] [CrossRef]
- Santos, A.C.D.M.; Santos, F.F.; Silva, R.M.; Gomes, T.A.T. Diversity of Hybrid- and Hetero-Pathogenic Escherichia Coli and Their Potential Implication in More Severe Diseases. Front. Cell. Infect. Microbiol. 2020, 10, 339. [Google Scholar] [CrossRef]
- Badouei, M.A.; Morabito, S.; Najafifar, A.; Mazandarani, E. Molecular Characterization of Enterohemorrhagic Escherichia Coli Hemolysin Gene (EHEC-HlyA)-Harboring Isolates from Cattle Reveals a Diverse Origin and Hybrid Diarrheagenic Strains. Infect. Genet. Evol. 2016, 39, 342–348. [Google Scholar] [CrossRef]
- Frank, C.; Werber, D.; Cramer, J.P.; Askar, M.; Faber, M.; an der Heiden, M.; Bernard, H.; Fruth, A.; Prager, R.; Spode, A.; et al. Epidemic Profile of Shiga-Toxin–Producing Escherichia Coli O104:H4 Outbreak in Germany. N. Engl. J. Med. 2011, 365, 1771–1780. [Google Scholar] [CrossRef] [Green Version]
- Osman, K.M.; Mustafa, A.M.; Elhariri, M.; Abdelhamed, G.S. The Distribution of Escherichia Coli Serovars, Virulence Genes, Gene Association and Combinations and Virulence Genes Encoding Serotypes in Pathogenic E. Coli Recovered from Diarrhoeic Calves, Sheep and Goat. Transbound. Emerg. Dis. 2013, 60, 69–78. [Google Scholar] [CrossRef]
- Algammal, A.M.; El-Kholy, A.W.; Riad, E.M.; Mohamed, H.E.; Elhaig, M.M.; Al Yousef, S.A.; Hozzein, W.N.; Ghobashy, M.O.I.I. Genes Encoding the Virulence and the Antimicrobial Resistance in Enterotoxigenic and Shiga-Toxigenic E. Coli Isolated from Diarrheic Calves. Toxins 2020, 12, 383. [Google Scholar] [CrossRef]
- Liang, G.; Malmuthuge, N.; Bao, H.; Stothard, P.; Griebel, P.J.; Guan, L.L. Transcriptome Analysis Reveals Regional and Temporal Differences in Mucosal Immune System Development in the Small Intestine of Neonatal Calves. BMC Genomics 2016, 17, 602. [Google Scholar] [CrossRef] [Green Version]
- Nissen, A.; Andersen, P.H.; Bendixen, E.; Ingvartsen, K.L.; Røntved, C.M. Colostrum and Milk Protein Rankings and Ratios of Importance to Neonatal Calf Health Using a Proteomics Approach. J. Dairy Sci. 2017, 100, 2711–2728. [Google Scholar] [CrossRef] [PubMed]
- DuBourdieu, D. Colostrum Antibodies, Egg Antibodies and Monoclonal Antibodies Providing Passive Immunity for Animals. In Nutraceuticals in Veterinary Medicine; Springer: Cham, Switzerland, 2019; pp. 245–257. ISBN 9783030046248. [Google Scholar]
- Ma, T.; O’Hara, E.; Song, Y.; Fischer, A.J.; He, Z.; Steele, M.A.; Guan, L.L. Altered Mucosa-Associated Microbiota in the Ileum and Colon of Neonatal Calves in Response to Delayed First Colostrum Feeding. J. Dairy Sci. 2019, 102, 7073–7086. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, M.H.; Sadri, H.; Steinhoff-Wagner, J.; Hammon, H.M.; Sauerwein, H. Effects of Colostrum Feeding on the MRNA Abundance of Genes Related to Toll-like Receptors, Key Antimicrobial Defense Molecules, and Tight Junctions in the Small Intestine of Neonatal Dairy Calves. J. Dairy Sci. 2021, 104, 10363–10373. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Oligonucleotide Sequence 5′ to 3′ | Gene | Pathotype | Product Size (bp) | Reference |
|---|---|---|---|---|---|
| EAE1 EAE2 | F:AAACAGGTGAAACTGTTGCC R:CTCTGCAGATTAACCTCTGC | eae | EPEC/EHEC | 490 | [25] |
| EVS1 EVC2 | F:ATCAGTCGTCACTCACTGGT R:CTGCTGTCACAGTGACAAA | stx1 | STEC/EHEC | 110 | [26] |
| EVT1 EVT2 | F:CAACACTGGATGATCTCAGC R:CCCCCTCAACTGCTAATA | stx2 | STEC/EHEC | 350 | |
| EHEC EHEC | F:ACGATGTGGTTTATTCTGGA R:CTTCACGTCACCATACATAT | hly | EHEC | 167 | [27] |
| EAEC EAEC | F: CTGGCGAAAGACTGTATCAT R:CAATGTATAGAAATCCGCTGTT | aatA | EAEC | 630 | [28] |
| BFP BFP | F:AATGGTGCTTGCGCTTGCTGC R:GCCGCTTTATCCAACCTGGTA | bfpA | Typical EPEC | 324 | [29] |
| ST1 ST2 | F: TTT ATT TCT GTA TTG TCT T R:GCAGGATTACAACACAATTC | St | ETEC | 294 | [30] |
| LT1 LT2 | F: GGCGACAGATTATACCGTGC R: CCGAATTCTGTTATATATGTC | lt | ETEC | 696 | [31] |
| IAL F IAL R | F: CTGGATGGTATGGTGAGG R:GGAGGCCAACAACATTATTTCC | ial | EIEC | 320 | [32] |
| daaE 1 daaE 2 | F:GAACGTTGGTTAATGTGGGGT R:TATTCACCGGTCGGTTATCAG | daaE | DAEC | 542 | [33] |
| Pathotype | Virulence Genes | Diarrheic Calves | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| eae | stx1 | stx2 | hly | aatA | lt | st | n (%) | n (%) | |
| EPEC | + | - | - | - | - | - | 3 (3.0) | 3 (3.0) | |
| STEC | - | + | - | - | - | - | - | 8 (8.1) | 23 (23.2) |
| - | + | + | - | - | - | - | 6 (6.1) | ||
| - | - | + | - | - | - | - | 9 (9.1) | ||
| EHEC | - | - | + | + | - | - | - | 1 (1.0) | 1 (1.0) |
| EAEC | - | - | - | - | + | - | - | 2 (2.0) | 2 (2.0) |
| ETEC | - | - | - | - | - | + | + | 2 (2.0) | 32 (32.3) |
| - | - | - | - | - | - | + | 30 (30.3) | ||
| STEC/ETEC | - | + | - | - | - | - | + | 6 (6.1) | 18 (18.2) |
| - | - | + | - | - | - | + | 10 (10.1) | ||
| - | + | + | - | - | - | + | 1 (1.0) | ||
| - | - | + | - | + | + | 1 (1.0) | |||
| TPD | - | - | - | - | - | - | - | - | 79 (79.8) |
| NPD | - | - | - | - | - | - | - | - | 20 (20.2) |
| Overall | - | - | - | - | - | - | - | - | 99 (100.0) |
| Pathotype | Diarrheic Calves—n (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Farm Size | Sex | Age (Weeks) | Breed | |||||||
| SF | MF | F | M | ≤2 | (2–4] | (4–7] | (7–10] | Indigenous | Crossbreed | |
| EPEC | 1 (2.0) | 2 (4.2) | 0 (0.0 b) | 3 (6.5 b) | 0 (0.0) | 0 (0.0) | 1 (3.7) | 2 (5.1) | 0 (0.0) | 3 (3.6) |
| STEC | 13 (25.5) | 10 (20.8) | 13 (24.5) | 10 (21.7) | 3 (100.0 a) | 4 (13.3 a) | 5 (18.5 a) | 11 (28.2 a) | 3 (20.0) | 20 (23.8) |
| EHEC | 1 (2.0) | 0 (0.0) | 0 (0.0) | 1 (2.2) | 0 (0.0) | 1 (3.3) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (1.2) |
| EAEC | 1 (2.0) | 1 (2.1) | 1 (1.9) | 1 (2.2) | 0 (0.0) | 0 (0.0) | 1 (3.7) | 1 (2.6) | 0 (0.0) | 2 (2.4) |
| ETEC | 16 (31.4) | 16 (33.3) | 16 (30.2) | 16 (34.8) | 0 (0.0) | 12 (40.0) | 8 (29.6) | 12 (30.8) | 4 (26.7) | 28 (33.3) |
| STEC/ETEC | 8 (15.7) | 10 (20.8) | 11 (20.8) | 7 (15.2) | 0 (0.0) | 7 (23.3) | 5 (18.5) | 6 (15.4) | 5 (33.3) | 13 (15.5) |
| NPD= | 11 (21.6) | 9 (18.8) | 12 (22.6) | 8 (17.4) | 0 (0.0) | 6 (20.0) | 7 (25.9) | 7 (17.9) | 3 (20.0) | 17 (20.2) |
| Total (n) | 51 | 48 | 53 | 46 | 3 | 30 | 27 | 39 | 15 | 84 |
| Pathotype | Diarrheic Calves—n (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Colostrum Feeding | Supplementary Feed | ||||||||||
| Feeding | Feeding Time (Hours) | Feeding Method | |||||||||
| No | Yes | (0–6] | (6–24] | Suckle | Hand | NS | Gz | Con | Hay | Com | |
| EPEC | 0 (0.0) | 3 (3.4) | 1 (2.3) | 2 (4.4) | 3 (4.2) | 0 (0.0) | 0 (0.0) | 2 (7.7) | 0 (0.0) | 0 (0.0) | 1 (4.0) |
| STEC | 3 (27.3) | 20 (22.7) | 8 (18.6) | 12 (26.7) | 12 (16.9 b) | 8 (47.1 b) | 3 (18.8) | 9 (34.6) | 0 (0.0) | 5 (21.7) | 6 (24.0) |
| EHEC | 0 (0.0) | 1 (1.1) | 0 (0.0) | 1 (2.2) | 1 (1.4) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (4.0) |
| EAEC | 0 (0.0) | 2 (2.3) | 0 (0.0) | 2 (4.4) | 0 (0.0) | 2 (11.8 a) | 0 (0.0) | 1 (4.0) | 0 (0.0) | 0 (0.0) | 1 (4.0) |
| ETEC | 2 (18.2) | 30 (34.1) | 16 (37.2) | 14 (31.1) | 27 (38.0) | 3 (17.6) | 7 (43.8) | 5 (19.2) | 4 (44.4) | 9 (39.1) | 7 (28.0) |
| STEC/ETEC | 4 (36.4) | 14 (15.9) | 5 (11.6) | 9 (20.0) | 11 (15.5) | 3 (17.6) | 4 (25.0) | 4 (15.4) | 2 (22.2) | 4 (17.4) | 4 (16.0) |
| NPD | 2 (18.2) | 18 (20.5) | 13 (30.2) | 5 (11.1) | 17 (23.9) | 1 (5.9) | 2 (12.5) | 5 (19.2) | 3 (33.3) | 5 (21.7) | 5 (20.0) |
| Total (n) | 11 | 88 | 43 | 45 | 71 | 17 | 16 | 26 | 9 | 23 | 25 |
| Pathotype | Diarrheic Calves—n (%) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Housing Type | Housing Floor | Housing Hygiene | ||||||||||||||||
| Pen | Barn | Soil | Concrete | Other | Very Poor | Poor | Good | Very Good | ||||||||||
| n | % | n | % | n | % | n | % | n | % | n | % | n | % | n | % | n | % | |
| EPEC | 1 | 1.4 | 2 | 6.7 | 2 | 2.8 | 1 | 11.1 | 0 | 0.0 | 0.0 | 2 | 3.3 | 0 | 0.0 | 1 | 14.3 | |
| STEC | 15 | 21.7 | 8 | 26.7 | 15 | 20.8 | 3 | 33.3 | 5 | 27.8 | 5 | 29.4 | 13 | 21.7 | 2 | 13.3 | 3 | 42.9 |
| EHEC | 1 | 1.4 | 0 | 0.0 | 1 | 1.4 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 1.7 | 0 | 0.0 | 0 | 0.0 |
| EAEC | 2 | 2.9 | 0 | 0.0 | 1 | 1.4 | 0 | 0.0 | 1 | 5.6 | 0 | 0.0 | 2 | 3.3 | 0 | 0.0 | 0 | 0.0 |
| ETEC | 24 | 34.8 | 8 | 26.7 | 25 | 34.7 | 1 | 11.1 | 6 | 33.3 | 2 | 11.8 | 22 | 36.7 | 7 | 46.7 | 1 | 14.3 |
| STEC/ETEC | 11 | 15.9 | 7 | 23.3 | 14 | 19.4 | 2 | 22.2 | 2 | 11.1 | 6 | 35.3 | 8 | 13.3 | 2 | 13.3 | 2 | 28.6 |
| NPD | 15 | 21.7 | 5 | 16.7 | 14 | 19.4 | 2 | 22.2 | 4 | 44.4 | 4 | 23.5 | 12 | 20.0 | 4 | 26.7 | 0 | 0.0 |
| Total (n) | 69 | 100.0 | 30 | 100.0 | 72 | 100.0 | 9 | 100.0 | 18 | 100.0 | 17 | 100.0 | 60 | 100.0 | 15 | 100.0 | 7 | 100.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chekole, W.S.; Adamu, H.; Sternberg-Lewrein, S.; Magnusson, U.; Tessema, T.S. Occurrence of Escherichia coli Pathotypes in Diarrheic Calves in a Low-Income Setting. Pathogens 2023, 12, 42. https://doi.org/10.3390/pathogens12010042
Chekole WS, Adamu H, Sternberg-Lewrein S, Magnusson U, Tessema TS. Occurrence of Escherichia coli Pathotypes in Diarrheic Calves in a Low-Income Setting. Pathogens. 2023; 12(1):42. https://doi.org/10.3390/pathogens12010042
Chicago/Turabian StyleChekole, Wagaw Sendeku, Haileeyesus Adamu, Susanna Sternberg-Lewrein, Ulf Magnusson, and Tesfaye Sisay Tessema. 2023. "Occurrence of Escherichia coli Pathotypes in Diarrheic Calves in a Low-Income Setting" Pathogens 12, no. 1: 42. https://doi.org/10.3390/pathogens12010042