
Patterns of ASFV Transmission in Domestic Pigs in Serbia
 , ,
, ,
Abstract
:1. Introduction
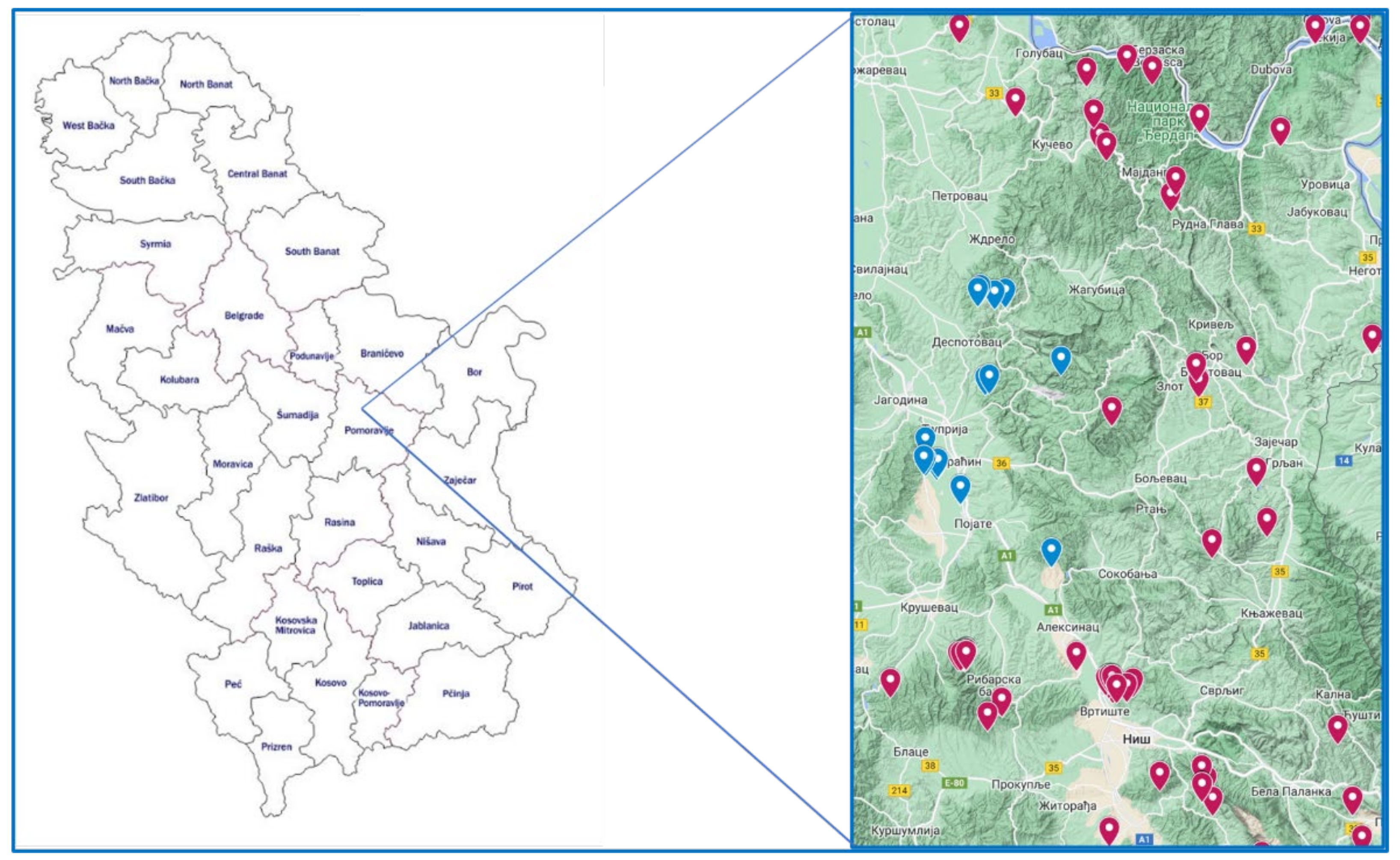
2. Materials and Methods
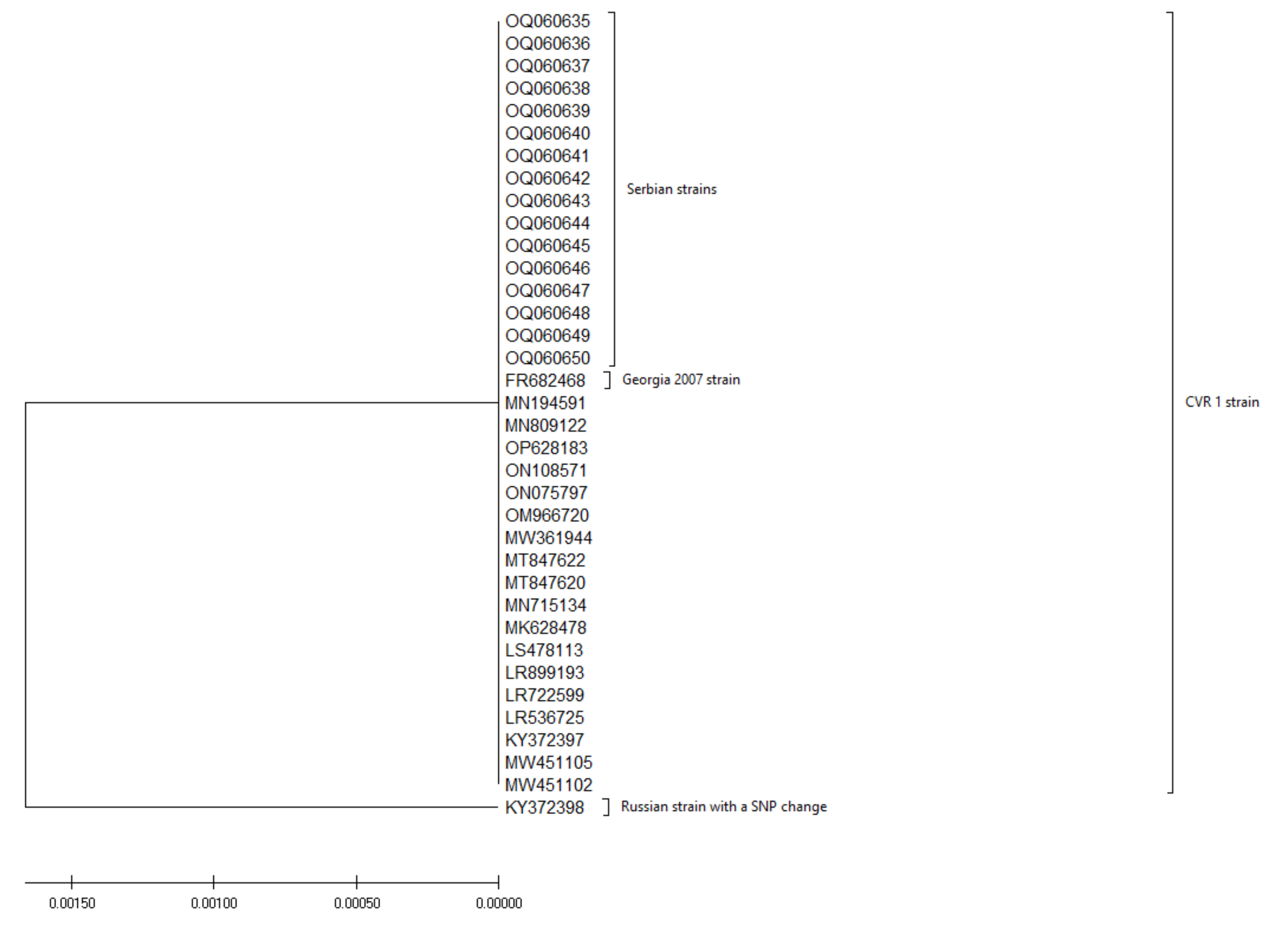
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, T.; Luo, R.; Sun, Y.; Qiu, H.J. Current Efforts towards Safe and Effective Live Attenuated Vaccines against African Swine Fever: Challenges and Prospects. Infect. Dis. Poverty 2021, 10, 137. [Google Scholar] [CrossRef]
- Depner, K.; Gortazar, C.; Guberti, V.; Masiulis, M.; More, S.; Olßsevskis, E.; Thulke, H.H.; Viltrop, A.; Wozniakowski, G.; Abrahantes, J.C.; et al. Epidemiological Analyses of African Swine Fever in the Baltic States and Poland (Update September 2016–September 2017). EFSA J. 2017, 15, e05068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M. ICTV Virus Taxonomy Profile: Asfarviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.R.; Harris, A.K. Cryo-EM Cools down Swine Fever. J. Biol. Chem. 2020, 295, 13–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur-Panasiuk, N.; Żmudzki, J.; Woźniakowski, G. African Swine Fever Virus—Persistence in Different Environmental Conditions and the Possibility of Its Indirect Transmission. J. Vet. Res. 2019, 63, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Carlson, J.; Fischer, M.; Zani, L.; Eschbaumer, M.; Fuchs, W.; Mettenleiter, T.; Beer, M.; Blome, S. Stability of African Swine Fever Virus in Soil and Options to Mitigate the Potential Transmission Risk. Pathogens 2020, 9, 977. [Google Scholar] [CrossRef]
- Prodelalova, J.; Kavanova, L.; Salat, J.; Moutelikova, R.; Kobzova, S.; Krasna, M.; Vasickova, P.; Simek, B.; Vaclavek, P. Experimental Evidence of the Long-Term Survival of Infective African Swine Fever Virus Strain Ba71V in Soil under Different Conditions. Pathogens 2022, 11, 648. [Google Scholar] [CrossRef]
- Gallardo, M.C.; Reoyo, A.D.L.T.; Fernández-Pinero, J.; Iglesias, I.; Muñoz, M.J.; Arias, M.L. African Swine Fever: A Global View of the Current Challenge. Porc. Health Manag. 2015, 1, 21. [Google Scholar] [CrossRef] [Green Version]
- Endris, R.G.; Haslett, T.M.; Hess, W.R. African Swine Fever Virus Infection in the Soft Tick, Ornithodoros (Alectorobius) Puertoricensis (Acari: Argasidae). J. Med. Entomol. 1992, 29, 990–994. [Google Scholar] [CrossRef]
- Podgórski, T.; Borowik, T.; Łyjak, M.; Woźniakowski, G. Spatial Epidemiology of African Swine Fever: Host, Landscape and Anthropogenic Drivers of Disease Occurrence in Wild Boar. Prev. Vet. Med. 2020, 177, 104691. [Google Scholar] [CrossRef]
- Hu, J.H.; Pei, X.; Sun, G.Q.; Jin, Z. Risk Analysis of the Transmission Route for the African Swine Fever Virus in Mainland China. Front. Phys. 2021, 9, 785885. [Google Scholar] [CrossRef]
- Dione, M.; Ouma, E.; Opio, F.; Kawuma, B.; Pezo, D. Qualitative Analysis of the Risks and Practices Associated with the Spread of African Swine Fever within the Smallholder Pig Value Chains in Uganda. Prev. Vet. Med. 2016, 135, 102–112. [Google Scholar] [CrossRef]
- Gallardo, C.; Mwaengo, D.M.; MacHaria, J.M.; Arias, M.; Taracha, E.A.; Soler, A.; Okoth, E.; Martín, E.; Kasiti, J.; Bishop, R.P. Enhanced Discrimination of African Swine Fever Virus Isolates through Nucleotide Sequencing of the P54, P72, and PB602L (CVR) Genes. Virus Genes 2009, 38, 85–95. [Google Scholar] [CrossRef]
- Faburay, B. Genome Plasticity of African Swine Fever Virus: Implications for Diagnostics and Live-Attenuated Vaccines. Pathogens 2022, 11, 145. [Google Scholar] [CrossRef]
- Gallardo, C.; Fernández-Pinero, J.; Pelayo, V.; Gazaev, I.; Markowska-Daniel, I.; Pridotkas, G.; Nieto, R.; Fernández-Pacheco, P.; Bokhan, S.; Nevolko, O.; et al. Genetic Variation among African Swine Fever Genotype II Viruses, Eastern and Central Europe. Emerg. Infect. Dis. 2014, 20, 1544–1547. [Google Scholar] [CrossRef] [Green Version]
- Mazur-Panasiuk, N.; Walczak, M.; Juszkiewicz, M.; Woźniakowski, G. The Spillover of African Swine Fever in Western Poland Revealed Its Estimated Origin on the Basis of O174L, K145R, MGF 505-5R and IGR I73R/I329L Genomic Sequences. Viruses 2020, 12, 1094. [Google Scholar] [CrossRef]
- King, D.P.; Reid, S.M.; Hutchings, G.H.; Grierson, S.S.; Wilkinson, P.J.; Dixon, L.K.; Bastos, A.D.S.; Drew, T.W. Development of a TaqMan PCR Assay with Internal Amplification Control for the Detection of African Swine Fever Virus. J. Virol. Methods 2003, 107, 53–61. [Google Scholar] [CrossRef]
- Bastos, A.D.S.; Penrith, M.-L.; Crucière, C.; Edrich, J.L.; Hutchings, G.; Roger, F.; Couacy-Hymann, E.; Thomson, G.R. Genotyping Field Strains of African Swine Fever Virus by Partial P72 Gene Characterisation. Arch. Virol. 2003, 148, 693–706. [Google Scholar] [CrossRef]
- Uzelac, J.; Đurić, B.; Ostojić, S.; Labus, T.; Nikolić, A.; Ćuk, J.; Živanović, D. Epizootiological Situation in Serbia in 2021. In Book of Abstracts, 33rd Conference of Serbian Veterinarians; Dimitrijević, V., Lazarević, M., Eds.; Srpsko Veterinarsko Drustvo: Zlatibor, Serbia, 2022; pp. 9–10. [Google Scholar]
- Sánchez-Cordón, P.J.; Nunez, A.; Neimanis, A.; Wikström-Lassa, E.; Montoya, M.; Crooke, H.; Gavier-Widén, D. African Swine Fever: Disease Dynamics in Wild Boar Experimentally Infected with ASFV Isolates Belonging to Genotype I and II. Viruses 2019, 11, 852. [Google Scholar] [CrossRef] [Green Version]
- Vergne, T.; Guinat, C.; Petkova, P.; Gogin, A.; Kolbasov, D.; Blome, S.; Molia, S.; Pinto Ferreira, J.; Wieland, B.; Nathues, H.; et al. Attitudes and Beliefs of Pig Farmers and Wild Boar Hunters Towards Reporting of African Swine Fever in Bulgaria, Germany and the Western Part of the Russian Federation. Transbound. Emerg. Dis. 2016, 63, e194–e204. [Google Scholar] [CrossRef]
- Mur, L.; Atzeni, M.; Martínez-López, B.; Feliziani, F.; Rolesu, S.; Sanchez-Vizcaino, J.M. Thirty-Five-Year Presence of African Swine Fever in Sardinia: History, Evolution and Risk Factors for Disease Maintenance. Transbound. Emerg. Dis. 2016, 63, e165–e177. [Google Scholar] [CrossRef] [PubMed]
- Milicévić, V.; Kureljušić, B.; Maksimović Zorić, J.; Savić, B.; Stanojević, S.; Milakara, E. First Occurence of African Swine Fever in Serbia. Acta Vet. Brno. 2019, 69, 443–449. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, I.; Martínez, M.; Montes, F.; de la Torre, A. Velocity of ASF Spread in Wild Boar in the European Union (2014–2017). Int. J. Infect. Dis. 2019, 79, 69. [Google Scholar] [CrossRef] [Green Version]
- Boklund, A.; Cay, B.; Depner, K.; Földi, Z.; Guberti, V.; Masiulis, M.; Miteva, A.; More, S.; Olsevskis, E.; Šatrán, P.; et al. Epidemiological Analyses of African Swine Fever in the European Union (November 2017 until November 2018). EFSA J. 2018, 16, e05494. [Google Scholar] [CrossRef] [PubMed]
- Nešković, M.; Ristić, B.; Došenović, R.; Grubač, S.; Petrović, T.; Prodanov-Radulović, J.; Polaček, V. African Swine Fever Outbreak Investigation on Large Commercial Pig Farm in Serbia. Acta Vet. Bel. 2021, 71, 219–229. [Google Scholar] [CrossRef]
- Schulz, K.; Sonnenburg, J.; Schauer, B.; Vergne, T.; Peyre, M.-I.; Staubach, C.; Conraths, F.J. Classical Swine Fever in Wild Boar: Surveillance Strategies under the Microscope. In International Congress for Veterinary Virology; Schulz, K., Sonnenburg, J., Schauer, B., Vergne, T., Peyre, M.-I., Staubach, C., Conraths, F.J., Eds.; CIRAD: Montpellier, France, 2015; pp. 101–103. [Google Scholar]
- Pol, F.; Rossi, S.; Mesplède, A.; Kuntz-Simon, G.; Le Potier, M.-F. Two Outbreaks of Classical Swine Fever in Wild Boar in France. Vet. Rec. 2008, 162, 811–816. [Google Scholar] [CrossRef]
- Šatrán, P.; Gruzdev, K. Expert Mission on African Swine Fever in Serbia; FAO: Rome, Italy; OIE: Paris, France, 2019. [Google Scholar]
- Costard, S.; Zagmutt, F.J.; Porphyre, T.; Pfeiffer, D.U. Small-Scale Pig Farmers’ Behavior, Silent Release of African Swine Fever Virus and Consequences for Disease Spread. Sci. Rep. 2015, 5, 17074. [Google Scholar] [CrossRef] [Green Version]
- Chantziaras, I.; Dewulf, J.; Van Limbergen, T.; Stadejek, T.; Niemi, J.; Kyriazakis, I.; Maes, D. Biosecurity Levels of Pig Fattening Farms from Four EU Countries and Links with the Farm Characteristics. Livest. Sci. 2020, 237, 104037. [Google Scholar] [CrossRef]
- Stoffel, C.; Buholzer, P.; Fanelli, A.; De Nardi, M. Analysis of the Drivers of ASF Introduction into the Officially Approved Pig Compartments in South Africa and Implications for the Revision of Biosecurity Standards. Porc. Health Manag. 2022, 8, 43. [Google Scholar] [CrossRef]
- Fasina, F.O.; Kissinga, H.; Mlowe, F.; Mshang’a, S.; Matogo, B.; Mrema, A.; Mhagama, A.; Makungu, S.; Mtui-Malamsha, N.; Sallu, R.; et al. Drivers, Risk Factors and Dynamics of African Swine Fever Outbreaks, Southern Highlands, Tanzania. Pathogens 2020, 9, 155. [Google Scholar] [CrossRef]
- O’Hara, K.C.; Beltrán-Alcrudo, D.; Hovari, M.; Tabakovski, B.; Martínez-López, B. Descriptive and Multivariate Analysis of the Pig Sector in North Macedonia and Its Implications for African Swine Fever Transmission. Front. Vet. Sci. 2021, 8, 733157. [Google Scholar] [CrossRef]
- Manuel Sánchez-Vizcaíno, J.; Martínez-López, B.; Martínez-Avilés, M.; Martins, C.; Boinas, F.; Vial, L.; Michaud, V.; Jori, F.; Etter, E.; Albina, E.; et al. Scientific Review on African Swine Fever. EFSA Support. Publ. 2009, 6, 5E. [Google Scholar] [CrossRef]
- Acevedo, P.; Aleksovski, V.; Apollonio, M.; Berdión, O.; Blanco-Aguiar, J.; del Rio, L.; Ertürk, A.; Fajdiga, L.; Escribano, F.; Ferroglio, E.; et al. Wild Boar Density Data Generated by Camera Trapping in Nineteen European Areas. EFSA Support. Publ. 2022, 19, 7214E. [Google Scholar] [CrossRef]
- More, S.; Miranda, M.A.; Bicout, D.; Bøtner, A.; Butterworth, A.; Calistri, P.; Edwards, S.; Garin-Bastuji, B.; Good, M.; Michel, V.; et al. African Swine Fever in Wild Boar. EFSA J. 2018, 16, e05344. [Google Scholar] [CrossRef] [Green Version]
- Forth, J.H.; Forth, L.F.; Blome, S.; Höper, D.; Beer, M. African Swine Fever Whole-Genome Sequencing—Quantity Wanted but Quality Needed. PLoS Pathog. 2020, 16, e1008779. [Google Scholar] [CrossRef]
- Bao, J.; Zhang, Y.; Shi, C.; Wang, Q.; Wang, S.; Wu, X.; Cao, S.; Xu, F.; Wang, Z. Genome-Wide Diversity Analysis of African Swine Fever Virus Based on a Curated Dataset. Animals 2022, 12, 2446. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mean | SD | Median | Min | Max | |
|---|---|---|---|---|---|
| External biosecurity score | 20.2 | 2.39 | 20 | 16 | 24 |
| Purchase of animals and semen | 29.7 | 4.52 | 30 | 22 | 38 |
| Transport of animals, removal of manure and dead animals | 8.1 | 1.45 | 8 | 6 | 10 |
| Feed, water, and equipment supply | 24 | 3.27 | 24 | 20 | 30 |
| Personnel and visitors | 8.8 | 2.44 | 8.5 | 6 | 14 |
| Vermin and bird control | 17.6 | 2.27 | 17.5 | 15 | 22 |
| Location of the farm | 28.4 | 3.20 | 28.5 | 24 | 35 |
| Internal biosecurity score | 30.2 | 1.32 | 30 | 28 | 32 |
| Disease management | 18.8 | 2.04 | 19 | 16 | 22 |
| Farrowing and suckling period management | 63 | 2.58 | 62.5 | 60 | 68 |
| Nursery unit management | 51.1 | 3.54 | 51 | 45 | 56 |
| Fattening unit management | 50.4 | 3.20 | 50.5 | 45 | 55 |
| Measures between compartments and use of equipment | 21.7 | 2.50 | 21.5 | 18 | 25 |
| Cleaning and disinfection | 0 | 0.00 | 0 | 0 | 0 |
| Overall biosecurity score | 27.6 | 2.07 | 28 | 24 | 30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glišić, D.; Milićević, V.; Veljović, L.; Milovanović, B.; Kureljušić, B.; Đorđević, I.; Anđelković, K.; Petković, J.; Dačić, M. Patterns of ASFV Transmission in Domestic Pigs in Serbia. Pathogens 2023, 12, 149. https://doi.org/10.3390/pathogens12010149
Glišić D, Milićević V, Veljović L, Milovanović B, Kureljušić B, Đorđević I, Anđelković K, Petković J, Dačić M. Patterns of ASFV Transmission in Domestic Pigs in Serbia. Pathogens. 2023; 12(1):149. https://doi.org/10.3390/pathogens12010149
Chicago/Turabian StyleGlišić, Dimitrije, Vesna Milićević, Ljubiša Veljović, Bojan Milovanović, Branislav Kureljušić, Igor Đorđević, Katarina Anđelković, Jelena Petković, and Miroljub Dačić. 2023. "Patterns of ASFV Transmission in Domestic Pigs in Serbia" Pathogens 12, no. 1: 149. https://doi.org/10.3390/pathogens12010149