
Polyphenols: Bioavailability, Microbiome Interactions and Cellular Effects on Health in Humans and Animals


Abstract
:1. Introduction
2. Metabolism of Dietary Polyphenols
2.1. Overview of Polyphenol Structure and Metabolism
2.2. The Role of the Microbiome in Polyphenol Digestion
2.3. Polyphenols in Circulation and Their Interactions with Organs, Tissues and Cellular Metabolism
2.4. Digestion and Absorption of Ellagitannins, Procyanidins and Flavonoids in Pigs
3. Health Effects of Dietary Polyphenols in Pigs and Other Animals
3.1. Oxidative Stress in Animals
3.2. The Anti-Oxidant Activity and Epigenetic Effects of Plant-Derived Polyphenols
3.3. Antibiotic Effects of Dietary Polyphenols
3.4. Polyphenols Can Reduce Inflammation in Pigs
4. Polyphenols in Bone
4.1. Dietary Polyphenols in Growing Bone
4.2. The Effect of Polyphenols on Bone Remodeling
4.3. Polyphenols in Bone, Potential for Forensic and Palaeodietary Studies
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Growth–Defense Tradeoffs in Plants: A Balancing Act to Optimize Fitness. Mol. Plant 2014, 7, 1267–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anton, D.; Bender, I.; Kaart, T.; Roasto, M.; Heinonen, M.; Luik, A.; Püssa, T. Changes in Polyphenols Contents and Antioxidant Capacities of Organically and Conventionally Cultivated Tomato (Solanum Lycopersicum L.) Fruits during Ripening. Int. J. Anal. Chem. 2017, 2017, e2367453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of Secondary Metabolites in Plant Defense against Pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Mokhtar, M.; Bouamar, S.; Di Lorenzo, A.; Temporini, C.; Daglia, M.; Riazi, A. The Influence of Ripeness on the Phenolic Content, Antioxidant and Antimicrobial Activities of Pumpkins (Cucurbita Moschata Duchesne). Molecules 2021, 26, 3623. [Google Scholar] [CrossRef]
- Boato, F.; Wortley, G.M.; Liu, R.H.; Glahn, R.P. Red Grape Juice Inhibits Iron Availability: Application of an in Vitro Digestion/Caco-2 Cell Model. J. Agric. Food Chem. 2002, 50, 6935–6938. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Noreen, H.; Farman, M.; McCullagh, J.S.O. Bioassay-Guided Isolation of Cytotoxic Flavonoids from Aerial Parts of Coronopus Didymus. J. Ethnopharmacol. 2016, 194, 971–980. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Rummun, N.; Rondeau, P.; Bourdon, E.; Pires, E.; McCullagh, J.; Claridge, T.D.W.; Bahorun, T.; Li, W.-W.; Neergheen, V.S. Terminalia Bentzoë, a Mascarene Endemic Plant, Inhibits Human Hepatocellular Carcinoma Cells Growth In Vitro via G0/G1 Phase Cell Cycle Arrest. Pharmaceuticals 2020, 13, 303. [Google Scholar] [CrossRef]
- Rummun, N.; Pires, E.; McCullagh, J.; Claridge, T.W.D.; Bahorun, T.; Li, W.-W.; Neergheen, V.S. Methyl Gallate—Rich Fraction of Syzygium Coriaceum Leaf Extract Induced Cancer Cell Cytotoxicity via Oxidative Stress. S. Afr. J. Bot. 2021, 137, 149–158. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of Polyphenols on Gut Microbiota and Implications in Human Health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [Green Version]
- Bieger, J.; Cermak, R.; Blank, R.; de Boer, V.C.J.; Hollman, P.C.H.; Kamphues, J.; Wolffram, S. Tissue Distribution of Quercetin in Pigs after Long-Term Dietary Supplementation. J. Nutr. 2008, 138, 1417–1420. [Google Scholar] [CrossRef]
- Ávila-Gálvez, M.A.; Giménez-Bastida, J.A.; González-Sarrías, A.; Espín, J.C. Tissue Deconjugation of Urolithin A Glucuronide to Free Urolithin A in Systemic Inflammation. Food Funct. 2019, 10, 3135–3141. [Google Scholar] [CrossRef] [Green Version]
- Alldritt, I.; Whitham-Agut, B.; Sipin, M.; Studholme, J.; Trentacoste, A.; Tripp, J.A.; Cappai, M.G.; Ditchfield, P.; Devièse, T.; Hedges, R.E.M.; et al. Metabolomics Reveals Diet-Derived Plant Polyphenols Accumulate in Physiological Bone. Sci. Rep. 2019, 9, 8047. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Benedetto, R.D.; Gargiulo, R.; Giovannini, C.; Masella, R. Polyphenols, Dietary Sources and Bioavailability. Ann. DellIstituto Super. Sanità 2007, 43, 348–361. [Google Scholar]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary Phenolics: Chemistry, Bioavailability and Effects on Health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Surai, P.F. Polyphenol Compounds in the Chicken/Animal Diet: From the Past to the Future. J. Anim. Physiol. Anim. Nutr. 2014, 98, 19–31. [Google Scholar] [CrossRef]
- Lipiński, K.; Mazur, M.; Antoszkiewicz, Z.; Purwin, C. Polyphenols in Monogastric Nutrition—A Review. Ann. Anim. Sci. 2017, 17, 41–58. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Afaq, F.; Saleem, M.; Ahmad, N.; Mukhtar, H. Targeting Multiple Signaling Pathways by Green Tea Polyphenol (-)-Epigallocatechin-3-Gallate. Cancer Res. 2006, 66, 2500–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giménez-Bastida, J.A.; González-Sarrías, A.; Larrosa, M.; Tomás-Barberán, F.; Espín, J.C.; García-Conesa, M.-T. Ellagitannin Metabolites, Urolithin A Glucuronide and Its Aglycone Urolithin A, Ameliorate TNF-α-Induced Inflammation and Associated Molecular Markers in Human Aortic Endothelial Cells. Mol. Nutr. Food Res. 2012, 56, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Pforte, H.; Hempel, J.; Jacobasch, G. Distribution Pattern of a Flavonoid Extract in the Gastrointestinal Lumen and Wall of Rats. Nahrung 1999, 43, 205–208. [Google Scholar] [CrossRef]
- Passamonti, S.; Vrhovsek, U.; Mattivi, F. The Interaction of Anthocyanins with Bilitranslocase. Biochem. Biophys. Res. Commun. 2002, 296, 631–636. [Google Scholar] [CrossRef]
- Doyle, B.; Griffiths, L.A. The Metabolism of Ellagic Acid in the Rat. Xenobiotica 1980, 10, 247–256. [Google Scholar] [CrossRef]
- Winter, J.; Moore, L.H.; Dowell, V.R.; Bokkenheuser, V.D. C-Ring Cleavage of Flavonoids by Human Intestinal Bacteria. Appl. Environ. Microbiol. 1989, 55, 1203–1208. [Google Scholar] [CrossRef] [Green Version]
- Scalbert, A.; Morand, C.; Manach, C.; Rémésy, C. Absorption and Metabolism of Polyphenols in the Gut and Impact on Health. Biomed. Pharmacother. 2002, 56, 276–282. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Yau, Y.F.; Leung, K.S.; El-Nezami, H.; Lee, J.C.-Y. Interaction of Polyphenols as Antioxidant and Anti-Inflammatory Compounds in Brain–Liver–Gut Axis. Antioxidants 2020, 9, 669. [Google Scholar] [CrossRef]
- Clifford, M.N. Diet-Derived Phenols in Plasma and Tissues and their Implications for Health. Planta Med. 2004, 70, 1103–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavefve, L.; Howard, L.R.; Carbonero, F. Berry Polyphenols Metabolism and Impact on Human Gut Microbiota and Health. Food Funct. 2020, 11, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Llorach, R.; Cerón, J.J.; Espín, J.C.; Tomás-Barberán, F.A. Evaluation of the Bioavailability and Metabolism in the Rat of Punicalagin, an Antioxidant Polyphenol from Pomegranate Juice. Eur. J. Nutr. 2003, 42, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Gasperotti, M.; Passamonti, S.; Tramer, F.; Masuero, D.; Guella, G.; Mattivi, F.; Vrhovsek, U. Fate of Microbial Metabolites of Dietary Polyphenols in Rats: Is the Brain Their Target Destination? ACS Chem. Neurosci. 2015, 6, 1341–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosme, F.; Gonçalves, B.; Bacelar, E.A.; Ines, A.; Jordão, A.M.; Vilela, A. Genotype, Environment and Management Practices on Red/ Dark-Colored Fruits Phenolic Composition and Its Impact on Sensory Attributes and Potential Health Benefits. In Phenolic Compounds-Natural Sources, Importance and Applications; Soto-Hernández, M., Palma-Tenango, M., Garcia-Mateos, M.R., Garcia-Mateos, Eds.; InTech Open: London, UK, 2017; pp. 261–282. ISBN 978-953-51-2957-8. [Google Scholar]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of Plant Polyphenols to Combat Oxidative Stress and Inflammatory Processes in Farm Animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef]
- Oesterle, I.; Braun, D.; Berry, D.; Wisgrill, L.; Rompel, A.; Warth, B. Polyphenol Exposure, Metabolism, and Analysis: A Global Exposomics Perspective. Annu. Rev. Food Sci. Technol. 2021, 12, 461–484. [Google Scholar] [CrossRef]
- Monteiro, M.; Farah, A.; Perrone, D.; Trugo, L.C.; Donangelo, C. Chlorogenic Acid Compounds from Coffee Are Differentially Absorbed and Metabolized in Humans. J. Nutr. 2007, 137, 2196–2201. [Google Scholar] [CrossRef] [Green Version]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the Polyphenols: Status and Controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef]
- Rafii, F.; Davis, C.; Park, M.; Heinze, T.M.; Beger, R.D. Variations in Metabolism of the Soy Isoflavonoid Daidzein by Human Intestinal Microfloras from Different Individuals. Arch. Microbiol. 2003, 180, 11–16. [Google Scholar] [CrossRef]
- Cerdá, B.; Espín, J.C.; Parra, S.; Martínez, P.; Tomás-Barberán, F.A. The Potent in Vitro Antioxidant Ellagitannins from Pomegranate Juice Are Metabolised into Bioavailable but Poor Antioxidant Hydroxy-6H-Dibenzopyran-6-One Derivatives by the Colonic Microflora of Healthy Humans. Eur. J. Nutr. 2004, 43, 205–220. [Google Scholar] [CrossRef]
- Cerdá, B.; Periago, P.; Espín, J.C.; Tomás-Barberán, F.A. Identification of Urolithin A as a Metabolite Produced by Human Colon Microflora from Ellagic Acid and Related Compounds. J. Agric. Food Chem. 2005, 53, 5571–5576. [Google Scholar] [CrossRef] [PubMed]
- Simons, A.L.; Renouf, M.; Hendrich, S.; Murphy, P.A. Human Gut Microbial Degradation of Flavonoids: Structure−function Relationships. J. Agric. Food Chem. 2005, 53, 4258–4263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastmanesh, R. High Polyphenol, Low Probiotic Diet for Weight Loss Because of Intestinal Microbiota Interaction. Chem. Biol. Interact. 2011, 189, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and Bioefficacy of Polyphenols in Humans. I. Review of 97 Bioavailability Studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chedea, V.; Palade, L.; Marin, D.; Pelmus, R.; Habeanu, M.; Rotar, M.; Gras, M.; Pistol, G.; Taranu, I. Intestinal Absorption and Antioxidant Activity of Grape Pomace Polyphenols. Nutrients 2018, 10, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Sarrías, A.; Espín, J.-C.; Tomás-Barberán, F.A.; García-Conesa, M.-T. Gene Expression, Cell Cycle Arrest and MAPK Signalling Regulation in Caco-2 Cells Exposed to Ellagic Acid and Its Metabolites, Urolithins. Mol. Nutr. Food Res. 2009, 53, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Cussotto, S.; Walsh, J.; Golubeva, A.V.; Zhdanov, A.V.; Strain, C.R.; Fouhy, F.; Stanton, C.; Dinan, T.G.; Hyland, N.P.; Clarke, G.; et al. The Gut Microbiome Influences the Bioavailability of Olanzapine in Rats. EBioMedicine 2021, 66, 103307. [Google Scholar] [CrossRef]
- Cortés-Martín, A.; García-Villalba, R.; González-Sarrías, A.; Romo-Vaquero, M.; Loria-Kohen, V.; Ramírez-de-Molina, A.; Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. The Gut Microbiota Urolithin Metabotypes Revisited: The Human Metabolism of Ellagic Acid Is Mainly Determined by Aging. Food Funct. 2018, 9, 4100–4106. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; García-Villalba, R.; González-Sarrías, A.; Selma, M.V.; Espín, J.C. Ellagic Acid Metabolism by Human Gut Microbiota: Consistent Observation of Three Urolithin Phenotypes in Intervention Trials, Independent of Food Source, Age, and Health Status. J. Agric. Food Chem. 2014, 62, 6535–6538. [Google Scholar] [CrossRef]
- Romo-Vaquero, M.; García-Villalba, R.; González-Sarrías, A.; Beltrán, D.; Tomás-Barberán, F.A.; Espín, J.C.; Selma, M.V. Interindividual Variability in the Human Metabolism of Ellagic Acid: Contribution of Gordonibacter to Urolithin Production. J. Funct. Foods 2015, 17, 785–791. [Google Scholar] [CrossRef]
- Crettol, S.; Petrovic, N.; Murray, M. Pharmacogenetics of Phase I and Phase II Drug Metabolism. Curr. Pharm. Des. 2010, 16, 204–219. [Google Scholar] [CrossRef] [PubMed]
- Galindo, P.; Rodriguez-Gómez, I.; González-Manzano, S.; Dueñas, M.; Jiménez, R.; Menéndez, C.; Vargas, F.; Tamargo, J.; Santos-Buelga, C.; Pérez-Vizcaíno, F.; et al. Glucuronidated Quercetin Lowers Blood Pressure in Spontaneously Hypertensive Rats via Deconjugation. PLoS ONE 2012, 7, e32673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sfakianos, J.; Coward, L.; Kirk, M.; Barnes, S. Intestinal Uptake and Biliary Excretion of the Isoflavone Genistein in Rats. J. Nutr. 1997, 127, 1260–1268. [Google Scholar] [CrossRef]
- Piskula, M.K.; Terao, J. Accumulation of (−)-Epicatechin Metabolites in Rat Plasma after Oral Administration and Distribution of Conjugation Enzymes in Rat Tissues. J. Nutr. 1998, 128, 1172–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donovan, J.L.; Lee, A.; Manach, C.; Rios, L.; Morand, C.; Scalbert, A.; Rémésy, C. Procyanidins Are Not Bioavailable in Rats Fed a Single Meal Containing a Grapeseed Extract or the Procyanidin Dimer B3. Br. J. Nutr. 2002, 87, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Misheva, M.; Ilott, N.E.; McCullagh, J.S.O. Recent Advances and Future Directions in Microbiome Metabolomics. Curr. Opin. Endocr. Metab. Res. 2021, 20, 100283. [Google Scholar] [CrossRef]
- Selma, M.; Tomás-Barberán, F.; Beltran, D.; Villalba, R.; Espín, J.C. Gordonibacter Urolithinfaciens Sp. Nov., an Urolithin-Producing Bacterium Isolated from Human Gut. Int. J. Syst. Evol. Microbiol. 2014, 64, 2346–2352. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; Beltrán, D.; García-Villalba, R.; Espín, J.C.; Tomás-Barberán, F.A. Description of Urolithin Production Capacity from Ellagic Acid of Two Human Intestinal Gordonibacter Species. Food Funct. 2014, 5, 1779–1784. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Zárate, P.; Cruz-Hernández, M.A.; Montañez, J.C.; Belmares-Cerda, R.E.; Aguilar, C.N. Bacterial Tannases: Production, Properties and Applications. Rev. Mex. Ing. Química 2014, 13, 63–74. [Google Scholar]
- López de Felipe, F.; de las Rivas, B.; Muñoz, R. Bioactive Compounds Produced by Gut Microbial Tannase: Implications for Colorectal Cancer Development. Front. Microbiol. 2014, 5, 684. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the Rescue of “Old” Metabolites to Understand a “New” Concept: Metabotypes as a Nexus among Phenolic Metabolism, Microbiota Dysbiosis, and Host Health Status. Mol. Nutr. Food Res. 2017, 61, 1500901. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; González-Sarrías, A.; Yáñez-Gascón, M.J.; Selma, M.V.; Azorín-Ortuño, M.; Toti, S.; Tomás-Barberán, F.; Dolara, P.; Espín, J.C. Anti-Inflammatory Properties of a Pomegranate Extract and Its Metabolite Urolithin-A in a Colitis Rat Model and the Effect of Colon Inflammation on Phenolic Metabolism. J. Nutr. Biochem. 2010, 21, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Anhê, F.F.; Pilon, G.; Roy, D.; Desjardins, Y.; Levy, E.; Marette, A. Triggering Akkermansia with Dietary Polyphenols: A New Weapon to Combat the Metabolic Syndrome? Gut Microbes 2016, 7, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Hur, H.-G.; Beger, R.; Heinze, T.; Lay, J.; Freeman, J.; Dore, J.; Rafii, F. Isolation of an Anaerobic Intestinal Bacterium Capable of Cleaving the C-Ring of the Isoflavonoid Daidzein. Arch. Microbiol. 2002, 178, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Braune, A.; Blaut, M. Intestinal Bacterium Eubacterium Cellulosolvens Deglycosylates Flavonoid C- and O- Glucosides. Appl. Environ. Microbiol. 2012, 78, 8151–8153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joannou, G.E.; Kelly, G.E.; Reeder, A.Y.; Waring, M.; Nelson, C. A Urinary Profile Study of Dietary Phytoestrogens. The Identification and Mode of Metabolism of New Isoflavonoids. J. Steroid Biochem. Mol. Biol. 1995, 54, 167–184. [Google Scholar] [CrossRef]
- Wang, X.-L.; Hur, H.-G.; Lee, J.H.; Kim, K.T.; Kim, S.-I. Enantioselective Synthesis of S-Equol from Dihydrodaidzein by a Newly Isolated Anaerobic Human Intestinal Bacterium. Appl. Environ. Microbiol. 2005, 71, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Matthies, A.; Blaut, M.; Braune, A. Isolation of a Human Intestinal Bacterium Capable of Daidzein and Genistein Conversion. Appl. Environ. Microbiol. 2009, 75, 1740–1744. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.-S.; Kitahara, M.; Sakamoto, M.; Hattori, M.; Benno, Y. Slackia Equolifaciens Sp. Nov., a Human Intestinal Bacterium Capable of Producing Equol. Int. J. Syst. Evol. Microbiol. 2010, 60, 1721–1724. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.Q.; Meselhy, M.R.; Li, Y.; Qin, G.W.; Hattori, M. Human Intestinal Bacteria Capable of Transforming Secoisolariciresinol Diglucoside to Mammalian Lignans, Enterodiol and Enterolactone. Chem. Pharm. Bull. 2000, 48, 1606–1610. [Google Scholar] [CrossRef] [Green Version]
- Clavel, T.; Henderson, G.; Alpert, C.-A.; Philippe, C.; Rigottier-Gois, L.; Doré, J.; Blaut, M. Intestinal Bacterial Communities That Produce Active Estrogen-Like Compounds Enterodiol and Enterolactone in Humans. Appl. Environ. Microbiol. 2005, 71, 6077–6085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás-Barberán, F.; García-Villalba, R.; Quartieri, A.; Raimondi, S.; Amaretti, A.; Leonardi, A.; Rossi, M. In Vitro Transformation of Chlorogenic Acid by Human Gut Microbiota. Mol. Nutr. Food Res. 2014, 58, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Axling, U.; Olsson, C.; Xu, J.; Fernandez, C.; Larsson, S.; Ström, K.; Ahrné, S.; Holm, C.; Molin, G.; Berger, K. Green Tea Powder and Lactobacillus Plantarum Affect Gut Microbiota, Lipid Metabolism and Inflammation in High-Fat Fed C57BL/6J Mice. Nutr. Metab. 2012, 9, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, H.; Orita, N.; Hatano, S.; Ichikawa, H.; Hara, Y.; Matsumoto, N.; Kimura, Y.; Terada, A.; Mitsuoka, T. Effect of Tea Polyphenols on Fecal Flora and Fecal Metabolic Products of Pigs. J. Vet. Med. Sci. 1995, 57, 45–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queipo-Ortuño, M.I.; Boto-Ordóñez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Cardona Diaz, F.; Andrés-Lacueva, C.; Tinahones, F.J. Influence of Red Wine Polyphenols and Ethanol on the Gut Microbiota Ecology and Biochemical Biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef]
- Etxeberria, U.; Fernández-Quintela, A.; Milagro, F.I.; Aguirre, L.; Martínez, J.A.; Portillo, M.P. Impact of Polyphenols and Polyphenol-Rich Dietary Sources on Gut Microbiota Composition. J. Agric. Food Chem. 2013, 61, 9517–9533. [Google Scholar] [CrossRef]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. A Survey of Modulation of Gut Microbiota by Dietary Polyphenols. BioMed Res. Int. 2015, 2015, 850902. [Google Scholar] [CrossRef]
- Burger-van Paassen, N.; Vincent, A.; Puiman, P.J.; van der Sluis, M.; Bouma, J.; Boehm, G.; van Goudoever, J.B.; van Seuningen, I.; Renes, I.B. The Regulation of Intestinal Mucin MUC2 Expression by Short-Chain Fatty Acids: Implications for Epithelial Protection. Biochem. J. 2009, 420, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Indias, I.; Sánchez-Alcoholado, L.; Pérez-Martínez, P.; Andrés-Lacueva, C.; Cardona, F.; Tinahones, F.; Queipo-Ortuño, M.I. Red Wine Polyphenols Modulate Fecal Microbiota and Reduce Markers of the Metabolic Syndrome in Obese Patients. Food Funct. 2016, 7, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Lewis, K.; Lutgendorff, F.; Phan, V.; Söderholm, J.D.; Sherman, P.M.; McKay, D.M. Enhanced Translocation of Bacteria across Metabolically Stressed Epithelia Is Reduced by Butyrate. Inflamm. Bowel Dis. 2010, 16, 1138–1148. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia Muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufour, C.; Dangles, O. Flavonoid–Serum Albumin Complexation: Determination of Binding Constants and Binding Sites by Fluorescence Spectroscopy. Biochim. Biophys. Acta BBA-Gen. Subj. 2005, 1721, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Yin, Y.-M.; Liu, H.-L.; Xie, M. Interactions of Flavonoids with Biomacromolecules. Curr. Org. Chem. 2011, 15, 2627–2640. [Google Scholar] [CrossRef]
- Poloni, D.M.; Dangles, O.; Vinson, J.A. Binding of Plant Polyphenols to Serum Albumin and LDL: Healthy Implications for Heart Disease. J. Agric. Food Chem. 2019, 67, 9139–9147. [Google Scholar] [CrossRef] [PubMed]
- Dangles, O.; Dufour, C.; Manach, C.; Morand, C.; Remesy, C. Binding of Flavonoids to Plasma Proteins. Methods Enzymol. 2001, 335, 319–333. [Google Scholar] [CrossRef]
- Tachibana, H.; Koga, K.; Fujimura, Y.; Yamada, K. A Receptor for Green Tea Polyphenol EGCG. Nat. Struct. Mol. Biol. 2004, 11, 380–381. [Google Scholar] [CrossRef]
- Murakami, A.; Ohnishi, K. Target Molecules of Food Phytochemicals: Food Science Bound for the next Dimension. Food Funct. 2012, 3, 462–476. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, A.I.; Real, R.; Pérez, M.; Mendoza, G.; Prieto, J.G.; Merino, G. Modulation of the Activity of ABC Transporters (P-Glycoprotein, MRP2, BCRP) by Flavonoids and Drug Response. J. Pharm. Sci. 2010, 99, 598–617. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Giménez-Bastida, J.A.; Núñez-Sánchez, M.Á.; Larrosa, M.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. Phase-II Metabolism Limits the Antiproliferative Activity of Urolithins in Human Colon Cancer Cells. Eur. J. Nutr. 2014, 53, 853–864. [Google Scholar] [CrossRef]
- Shen, S.-C.; Chen, Y.-C.; Hsu, F.-L.; Lee, W.-R. Differential Apoptosis-Inducing Effect of Quercetin and Its Glycosides in Human Promyeloleukemic HL-60 Cells by Alternative Activation of the Caspase 3 Cascade. J. Cell. Biochem. 2003, 89, 1044–1055. [Google Scholar] [CrossRef] [PubMed]
- Aires, V.; Limagne, E.; Cotte, A.K.; Latruffe, N.; Ghiringhelli, F.; Delmas, D. Resveratrol Metabolites Inhibit Human Metastatic Colon Cancer Cells Progression and Synergize with Chemotherapeutic Drugs to Induce Cell Death. Mol. Nutr. Food Res. 2013, 57, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Gálvez, M.Á.; Espín, J.C.; González-Sarrías, A. Physiological Relevance of the Antiproliferative and Estrogenic Effects of Dietary Polyphenol Aglycones versus Their Phase-II Metabolites on Breast Cancer Cells: A Call of Caution. J. Agric. Food Chem. 2018, 66, 8547–8555. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Barron, D.; Orfila, C.; Dionisi, F.; Krajcsi, P.; Williamson, G. Interaction of Hydroxycinnamic Acids and Their Conjugates with Organic Anion Transporters and ATP-Binding Cassette Transporters. Mol. Nutr. Food Res. 2011, 55, 979–988. [Google Scholar] [CrossRef]
- Shimoi, K.; Saka, N.; Nozawa, R.; Sato, M.; Amano, I.; Nakayama, T.; Kinae, N. Deglucuronidation of a Flavonoid, Luteolin Monoglucuronide, during Inflammation. Drug Metab. Dispos. Biol. Fate Chem. 2001, 29, 1521–1524. [Google Scholar]
- Shimoi, K.; Nakayama, T. Glucuronidase Deconjugation in Inflammation. Methods Enzymol. 2005, 400, 263–272. [Google Scholar] [CrossRef]
- Kunihiro, A.G.; Brickey, J.A.; Frye, J.B.; Luis, P.B.; Schneider, C.; Funk, J.L. Curcumin, but Not Curcumin-Glucuronide, Inhibits Smad Signaling in TGFβ-Dependent Bone Metastatic Breast Cancer Cells and Is Enriched in Bone Compared to Other Tissues. J. Nutr. Biochem. 2019, 63, 150–156. [Google Scholar] [CrossRef]
- Menendez, C.; Dueñas, M.; Galindo, P.; González-Manzano, S.; Jimenez, R.; Moreno, L.; Zarzuelo, M.J.; Rodríguez-Gómez, I.; Duarte, J.; Santos-Buelga, C.; et al. Vascular Deconjugation of Quercetin Glucuronide: The Flavonoid Paradox Revealed? Mol. Nutr. Food Res. 2011, 55, 1780–1790. [Google Scholar] [CrossRef]
- Ishisaka, A.; Kawabata, K.; Miki, S.; Shiba, Y.; Minekawa, S.; Nishikawa, T.; Mukai, R.; Terao, J.; Kawai, Y. Mitochondrial DysfunctionlLeads to Deconjugation of Quercetin Glucuronides in Inflammatory Macrophages. PLoS ONE 2013, 8, e80843. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.M. Regulation of Intracellular Ca2+ by Reactive Oxygen Species in Osteoblasts Treated with Antimycin A. J. Appl. Toxicol. 2012, 32, 118–125. [Google Scholar] [CrossRef]
- Kawai, Y. Understanding Metabolic Conversions and Molecular Actions of Flavonoids in Vivo: Toward New Strategies for Effective Utilization of Natural Polyphenols in Human Health. J. Med. Investig. 2018, 65, 162–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, E.R.; Ullrey, D.E. The Pig as a Model for Human Nutrition. Annu. Rev. Nutr. 1987, 7, 361–382. [Google Scholar] [CrossRef] [PubMed]
- Espín, J.C.; González-Barrio, R.; Cerdá, B.; López-Bote, C.; Rey, A.I.; Tomás-Barberán, F.A. Iberian Pig as a Model to Clarify Obscure Points in the Bioavailability and Metabolism of Ellagitannins in Humans. J. Agric. Food Chem. 2007, 55, 10476–10485. [Google Scholar] [CrossRef] [PubMed]
- French, K.E.; Harvey, J.; McCullagh, J.S.O. Targeted and Untargeted Metabolic Profiling of Wild Grassland Plants Identifies Antibiotic and Anthelmintic Compounds Targeting Pathogen Physiology, Metabolism and Reproduction. Sci. Rep. 2018, 8, 1695. [Google Scholar] [CrossRef] [Green Version]
- Larrosa, M.; Tomás-Barberán, F.A.; Espín, J.C. The Dietary Hydrolysable Tannin Punicalagin Releases Ellagic Acid That Induces Apoptosis in Human Colon Adenocarcinoma Caco-2 Cells by Using the Mitochondrial Pathway. J. Nutr. Biochem. 2006, 17, 611–625. [Google Scholar] [CrossRef]
- Andreux, P.A.; Blanco-Bose, W.; Ryu, D.; Burdet, F.; Ibberson, M.; Aebischer, P.; Auwerx, J.; Singh, A.; Rinsch, C. The Mitophagy Activator Urolithin A Is Safe and Induces a Molecular Signature of Improved Mitochondrial and Cellular Health in Humans. Nat. Metab. 2019, 1, 595–603. [Google Scholar] [CrossRef]
- Haddock, E.A.; Gupta, R.K.; Al-Shafi, S.M.K.; Haslam, E.; Magnolato, D. The Metabolism of Gallic Acid and Hexahydroxydiphenic Acid in Plants. Part 1. Introduction. Naturally Occurring Galloyl Esters. J. Chem. Soc. Perkin 1982, 1, 2515–2524. [Google Scholar] [CrossRef]
- Clifford, M.N.; Scalbert, A. Ellagitannins—Nature, Occurrence and Dietary Burden. J. Sci. Food Agric. 2000, 80, 1118–1125. [Google Scholar] [CrossRef]
- Cerdá, B.; Tomás-Barberán, F.A.; Espín, J.C. Metabolism of Antioxidant and Chemopreventive Ellagitannins from Strawberries, Raspberries, Walnuts, and Oak-Aged Wine in Humans: Identification of Biomarkers and Individual Variability. J. Agric. Food Chem. 2005, 53, 227–235. [Google Scholar] [CrossRef]
- Cappai, M.G.; Wolf, P.; Rust, P.; Pinna, W.; Kamphues, J. Raw Hulled Shredded Acorns from Downy Oak (Quercus pubescens) in the Diet of Pigs: Effects on Digestibility and Faeces Characteristics. J. Anim. Physiol. Anim. Nutr. 2013, 97, 1–5. [Google Scholar] [CrossRef]
- Dacrema, M.; Sommella, E.; Santarcangelo, C.; Bruno, B.; Marano, M.G.; Insolia, V.; Saviano, A.; Campiglia, P.; Stornaiuolo, M.; Daglia, M. Metabolic Profiling, in Vitro Bioaccessibility and in Vivo Bioavailability of a Commercial Bioactive Epilobium angustifolium L. Extract. Biomed. Pharmacother. 2020, 131, 110670. [Google Scholar] [CrossRef] [PubMed]
- García-Villalba, R.; Giménez-Bastida, J.A.; Cortés-Martín, A.; Ávila-Gálvez, M.Á.; Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C.; González-Sarrías, A. Urolithins: A Comprehensive Update on Their Metabolism, Bioactivity, and Associated Gut Microbiota. Mol. Nutr. Food Res. 2022, e2101019. [Google Scholar] [CrossRef] [PubMed]
- Pfundstein, B.; Haubner, R.; Würtele, G.; Gehres, N.; Ulrich, C.M.; Owen, R.W. Pilot Walnut Intervention Study of Urolithin Bioavailability in Human Volunteers. J. Agric. Food Chem. 2014, 62, 10264–10273. [Google Scholar] [CrossRef]
- Li, Z.; Summanen, P.H.; Komoriya, T.; Henning, S.M.; Lee, R.-P.; Carlson, E.; Heber, D.; Finegold, S.M. Pomegranate Ellagitannins Stimulate Growth of Gut Bacteria in Vitro: Implications for Prebiotic and Metabolic Effects. Anaerobe 2015, 34, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Aronson, W.J.; Zhang, Y.; Henning, S.M.; Moro, A.; Lee, R.; Sartippour, M.; Harris, D.M.; Rettig, M.; Suchard, M.A.; et al. Pomegranate Ellagitannin-Derived Metabolites Inhibit Prostate Cancer Growth and Localize to the Mouse Prostate Gland. J. Agric. Food Chem. 2007, 55, 7732–7737. [Google Scholar] [CrossRef] [PubMed]
- González-Sarrías, A.; Giménez-Bastida, J.A.; García-Conesa, M.T.; Gómez-Sánchez, M.B.; García-Talavera, N.V.; Gil-Izquierdo, A.; Sánchez-Álvarez, C.; Fontana-Compiano, L.O.; Morga-Egea, J.P.; Pastor-Quirante, F.A.; et al. Occurrence of Urolithins, Gut Microbiota Ellagic Acid Metabolites and Proliferation Markers Expression Response in the Human Prostate Gland upon Consumption of Walnuts and Pomegranate Juice. Mol. Nutr. Food Res. 2010, 54, 311–322. [Google Scholar] [CrossRef]
- De Boer, V.C.J.; Dihal, A.A.; van der Woude, H.; Arts, I.C.W.; Wolffram, S.; Alink, G.M.; Rietjens, I.M.C.M.; Keijer, J.; Hollman, P.C.H. Tissue Distribution of Quercetin in Rats and Pigs. J. Nutr. 2005, 135, 1718–1725. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.-H.; Lee, D.-Y.; Kim, S.; Chung, J.-O.; Choi, J.-K.; Joo, K.-M.; Woo Jeong, H.; Kee Kim, J.; Gi Kim, W.; Shim, S.-M. Influence of Flavonol-Rich Excipient Food (Onion Peel and Dendropanax morbifera) on the Bioavailability of Green Tea Epicatechins in vitro and in vivo. Food Funct. 2017, 8, 3664–3674. [Google Scholar] [CrossRef]
- Appeldoorn, M.M.; Vincken, J.-P.; Gruppen, H.; Hollman, P.C.H. Procyanidin Dimers A1, A2, and B2 Are Absorbed without Conjugation or Methylation from the Small Intestine of Rats. J. Nutr. 2009, 139, 1469–1473. [Google Scholar] [CrossRef] [Green Version]
- Sroka, Z.; Cisowski, W. Hydrogen Peroxide Scavenging, Antioxidant and Anti-Radical Activity of Some Phenolic Acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) Models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as Antitumor Agents Targeting Key Players in Cancer-Driving Signaling Pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef]
- Janisch, K.M.; Williamson, G.; Needs, P.; Plumb, G.W. Properties of Quercetin Conjugates: Modulation of LDL Oxidation and Binding to Human Serum Albumin. Free Radic. Res. 2004, 38, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Mohos, V.; Fliszár-Nyúl, E.; Schilli, G.; Hetényi, C.; Lemli, B.; Kunsági-Máté, S.; Bognár, B.; Poór, M. Interaction of Chrysin and Its Main Conjugated Metabolites Chrysin-7-Sulfate and Chrysin-7-Glucuronide with Serum Albumin. Int. J. Mol. Sci. 2018, 19, 4073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreyev, A.Y.; Kushnareva, Y.E.; Starkov, A.A. Mitochondrial Metabolism of Reactive Oxygen Species. Biochemistry 2005, 70, 200–214. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.-W.; Tsuji, Y. Reactive Oxygen Species (ROS) Homeostasis and Redox Regulation in Cellular Signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Vlahos, R.; Stambas, J.; Selemidis, S. Suppressing Production of Reactive Oxygen Species (ROS) for Influenza A Virus Therapy. Trends Pharmacol. Sci. 2012, 33, 3–8. [Google Scholar] [CrossRef]
- Panday, A.; Sahoo, M.K.; Osorio, D.; Batra, S. NADPH Oxidases: An Overview from Structure to Innate Immunity-Associated Pathologies. Cell. Mol. Immunol. 2015, 12, 5–23. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Carr, A.C.; McCall, M.R.; Frei, B. Oxidation of LDL by Myeloperoxidase and Reactive Nitrogen Species. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1716–1723. [Google Scholar] [CrossRef] [Green Version]
- Ozgocmen, S.; Kaya, H.; Fadillioglu, E.; Aydogan, R.; Yilmaz, Z. Role of Antioxidant Systems, Lipid Peroxidation, and Nitric Oxide in Postmenopausal Osteoporosis. Mol. Cell. Biochem. 2007, 295, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L. Modulation of Skeletal Muscle Antioxidant Defense by Exercise: Role of Redox Signaling. Free Radic. Biol. Med. 2008, 44, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Skrip, M.M.; McWilliams, S.R. Oxidative Balance in Birds: An Atoms-to-Organisms-to-Ecology Primer for Ornithologists. J. Field Ornithol. 2016, 87, 1–20. [Google Scholar] [CrossRef]
- Vogel, R.; Wiesinger, H.; Hamprecht, B.; Dringen, R. The Regeneration of Reduced Glutathione in Rat Forebrain Mitochondria Identifies Metabolic Pathways Providing the NADPH Required. Neurosci. Lett. 1999, 275, 97–100. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive Oxygen Species, Cellular Redox Systems, and Apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.I.; Griendling, K.K. Nox Proteins in Signal Transduction. Free Radic. Biol. Med. 2009, 47, 1239–1253. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, M.; Arora, S.; Li, J.; Rahmani, R.; Sun, L.; Steinlauf, A.F.; Mechanick, J.I.; Zaidi, M. Bone, Inflammation, and Inflammatory Bowel Disease. Curr. Osteoporos. Rep. 2011, 9, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Takeno, A.; Kanazawa, I.; Tanaka, K.; Notsu, M.; Yokomoto, M.; Yamaguchi, T.; Sugimoto, T. Activation of AMP-Activated Protein Kinase Protects against Homocysteine-Induced Apoptosis of Osteocytic MLO-Y4 Cells by Regulating the Expressions of NADPH Oxidase 1 (Nox1) and Nox2. Bone 2015, 77, 135–141. [Google Scholar] [CrossRef]
- Ashkenazi, A.; Dixit, V.M. Death Receptors: Signaling and Modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef] [Green Version]
- Green, D.R.; Reed, J.C. Mitochondria and Apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef]
- Whitehead, T.P.; Robinson, D.; Allaway, S.; Syms, J.; Hale, A. Effect of Red Wine Ingestion on the Antioxidant Capacity of Serum. Clin. Chem. 1995, 41, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Cheon, B.S.; Kim, Y.H.; Son, K.S.; Chang, H.W.; Kang, S.S.; Kim, H.P. Effects of Prenylated Flavonoids and Biflavonoids on Lipopolysaccharide-Induced Nitric Oxide Production from the Mouse Macrophage Cell Line RAW 264.7. Planta Med. 2000, 66, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A Review of Probable Mechanisms of Action and Potential Applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sharma, U.K.; Sharma, A.K.; Pandey, A.K. Protective Efficacy of Solanum Xanthocarpum Root Extracts against Free Radical Damage: Phytochemical Analysis and Antioxidant Effect. Cell Mol. Biol. 2012, 58, 174–181. [Google Scholar] [CrossRef]
- Hider, R.C.; Liu, Z.D.; Khodr, H.H. Metal Chelation of Polyphenols. Methods Enzymol. 2001, 335, 190–203. [Google Scholar] [CrossRef]
- Palierse, E.; Przybylski, C.; Brouri, D.; Jolivalt, C.; Coradin, T. Interactions of Calcium with Chlorogenic and Rosmarinic Acids: An Experimental and Theoretical Approach. Int. J. Mol. Sci. 2020, 21, 4948. [Google Scholar] [CrossRef]
- Perron, N.R.; Brumaghim, J.L. A Review of the Antioxidant Mechanisms of Polyphenol Compounds Related to Iron Binding. Cell Biochem. Biophys. 2009, 53, 75–100. [Google Scholar] [CrossRef]
- Cherrak, S.A.; Mokhtari-Soulimane, N.; Berroukeche, F.; Bensenane, B.; Cherbonnel, A.; Merzouk, H.; Elhabiri, M. In Vitro Antioxidant versus Metal Ion Chelating Properties of Flavonoids: A Structure-Activity Investigation. PLoS ONE 2016, 11, e0165575. [Google Scholar] [CrossRef]
- Lesjak, M.; Srai, S.K.S. Role of Dietary Flavonoids in Iron Homeostasis. Pharmaceuticals 2019, 12, 119. [Google Scholar] [CrossRef] [Green Version]
- Tanigawa, S.; Fujii, M.; Hou, D.-X. Action of Nrf2 and Keap1 in ARE-Mediated NQO1 Expression by Quercetin. Free Radic. Biol. Med. 2007, 42, 1690–1703. [Google Scholar] [CrossRef]
- Cardozo, L.F.M.F.; Pedruzzi, L.M.; Stenvinkel, P.; Stockler-Pinto, M.B.; Daleprane, J.B.; Leite, M.; Mafra, D. Nutritional Strategies to Modulate Inflammation and Oxidative Stress Pathways via Activation of the Master Antioxidant Switch Nrf2. Biochimie 2013, 95, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Ma, W.; Huang, C.; Yang, C.S. Inhibition of Tumor Promoter-Induced Activator Protein 1 Activation and Cell Transformation by Tea Polyphenols, (-)-Epigallocatechin Gallate, and Theaflavins. Cancer Res. 1997, 57, 4414–4419. [Google Scholar] [PubMed]
- Kim, H.P.; Mani, I.; Iversen, L.; Ziboh, V.A. Effects of Naturally-Occurring Flavonoids and Biflavonoids on Epidermal Cyclooxygenase and Iipoxygenase from Guinea-Pigs. Prostaglandins Leukot. Essent. Fat. Acids 1998, 58, 17–24. [Google Scholar] [CrossRef]
- Uche, F.I.; Abed, M.; Abdullah, M.I.; Drijfhout, F.P.; McCullagh, J.; Claridge, T.W.D.; Richardson, A.; Li, W.-W. O9 Isolation, Identification and Anti-Cancer Activity of Minor Alkaloids from Triclisia subcordata Oliv. Biochem. Pharmacol. 2017, 139, 112. [Google Scholar] [CrossRef]
- Uche, F.I.; McCullagh, J.; Claridge, T.W.D.; Richardson, A.; Li, W.-W. Synthesis of (Aminoalkyl)Cycleanine Analogues: Cytotoxicity, Cellular Uptake, and Apoptosis Induction in Ovarian Cancer Cells. Bioorg. Med. Chem. Lett. 2018, 28, 1652–1656. [Google Scholar] [CrossRef]
- Fadayomi, I.E.; Johnson-Ajinwo, O.R.; Pires, E.; McCullagh, J.; Claridge, T.D.W.; Forsyth, N.R.; Li, W.-W. Clerodane Diterpenoids from an Edible Plant Justicia insularis: Discovery, Cytotoxicity, and Apoptosis Induction in Human Ovarian Cancer Cells. Molecules 2021, 26, 5933. [Google Scholar] [CrossRef]
- Pan, M.-H.; Lai, C.-S.; Wu, J.-C.; Ho, C.-T. Epigenetic and Disease Targets by Polyphenols. Curr. Pharm. Des. 2013, 19, 6156–6185. [Google Scholar] [CrossRef]
- Arora, I.; Sharma, M.; Tollefsbol, T.O. Combinatorial Epigenetics Impact of Polyphenols and Phytochemicals in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2019, 20, 4567. [Google Scholar] [CrossRef] [Green Version]
- Arora, I.; Sharma, M.; Sun, L.Y.; Tollefsbol, T.O. The Epigenetic Link between Polyphenols, Aging and Age-Related Diseases. Genes 2020, 11, 1094. [Google Scholar] [CrossRef]
- Fang, M.; Chen, D.; Yang, C.S. Dietary Polyphenols May Affect DNA Methylation. J. Nutr. 2007, 137, 223S–228S. [Google Scholar] [CrossRef] [Green Version]
- Fang, M.Z.; Wang, Y.; Ai, N.; Hou, Z.; Sun, Y.; Lu, H.; Welsh, W.; Yang, C.S. Tea Polyphenol (−)-Epigallocatechin-3-Gallate Inhibits DNA Methyltransferase and Reactivates Methylation-Silenced Genes in Cancer Cell Lines. Cancer Res. 2003, 63, 7563–7570. [Google Scholar] [PubMed]
- Tili, E.; Michaille, J.-J.; Alder, H.; Volinia, S.; Delmas, D.; Latruffe, N.; Croce, C.M. Resveratrol Modulates the Levels of MicroRNAs Targeting Genes Encoding Tumor-Suppressors and Effectors of TGFβ Signaling Pathway in SW480 Cells. Biochem. Pharmacol. 2010, 80, 2057–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abtin, M.; Alivand, M.R.; Khaniani, M.S.; Bastami, M.; Zaeifizadeh, M.; Derakhshan, S.M. Simultaneous Downregulation of MiR-21 and MiR-155 through Oleuropein for Breast Cancer Prevention and Therapy. J. Cell. Biochem. 2018, 119, 7151–7165. [Google Scholar] [CrossRef] [PubMed]
- Fabiani, R.; Vella, N.; Rosignoli, P. Epigenetic Modifications Induced by Olive Oil and Its Phenolic Compounds: A Systematic Review. Molecules 2021, 26, 273. [Google Scholar] [CrossRef] [PubMed]
- Nanda, N.; Mahmood, S.; Bhatia, A.; Mahmood, A.; Dhawan, D.K. Chemopreventive Role of Olive Oil in Colon Carcinogenesis by Targeting Noncoding RNAs and Methylation Machinery. Int. J. Cancer 2019, 144, 1180–1194. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Falconi, A.; Di Germanio, C.; Micioni Di Bonaventura, M.V.; Costa, A.; Caramuta, S.; Del Carlo, M.; Compagnone, D.; Dainese, E.; Cifani, C.; et al. Extravirgin Olive Oil Up-Regulates CB1 Tumor Suppressor Gene in Human Colon Cancer Cells and in Rat Colon via Epigenetic Mechanisms. J. Nutr. Biochem. 2015, 26, 250–258. [Google Scholar] [CrossRef]
- Magrone, T.; Kumazawa, Y.; Jirillo, E. Polyphenol-Mediated Beneficial Effects in Healthy Status and Disease with Special Reference to Immune-Based Mechanisms. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 467–479. ISBN 978-0-12-398456-2. [Google Scholar]
- Ding, S.; Jiang, H.; Fang, J. Regulation of Immune Function by Polyphenols. J. Immunol. Res. 2018, 2018, 1264074. [Google Scholar] [CrossRef] [Green Version]
- Umeda, D.; Yano, S.; Yamada, K.; Tachibana, H. Involvement of 67-KDa Laminin Receptor-Mediated Myosin Phosphatase Activation in Antiproliferative Effect of Epigallocatechin-3-O-Gallate at a Physiological Concentration on Caco-2 Colon Cancer Cells. Biochem. Biophys. Res. Commun. 2008, 371, 172–176. [Google Scholar] [CrossRef]
- Shammas, M.A.; Neri, P.; Koley, H.; Batchu, R.B.; Bertheau, R.C.; Munshi, V.; Prabhala, R.; Fulciniti, M.; Tai, Y.T.; Treon, S.P.; et al. Specific Killing of Multiple Myeloma Cells by (-)-Epigallocatechin-3-Gallate Extracted from Green Tea: Biologic Activity and Therapeutic Implications. Blood 2006, 108, 2804–2810. [Google Scholar] [CrossRef]
- Fujimura, Y.; Umeda, D.; Yano, S.; Maeda-Yamamoto, M.; Yamada, K.; Tachibana, H. The 67kDa Laminin Receptor as a Primary Determinant of Anti-Allergic Effects of O-Methylated EGCG. Biochem. Biophys. Res. Commun. 2007, 364, 79–85. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Lee, D.-H.; Jeong, J.-H.; Guo, Z.S.; Lee, Y.J. Quercetin Augments TRAIL-Induced Apoptotic Death: Involvement of the ERK Signal Transduction Pathway. Biochem. Pharmacol. 2008, 75, 1946–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Shi, D.; Liu, L.; Wang, J.; Xie, X.; Kang, T.; Deng, W. Quercetin Suppresses Cyclooxygenase-2 Expression and Angiogenesis through Inactivation of P300 Signaling. PLoS ONE 2011, 6, e22934. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, P.A.; Braune, A.; Hölzlwimmer, G.; Quintanilla-Fend, L.; Haller, D. Quercetin Inhibits TNF-Induced NF-κB Transcription Factor Recruitment to Proinflammatory Gene Promoters in Murine Intestinal Epithelial Cells. J. Nutr. 2007, 137, 1208–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Shan, S.; Zhang, C.; Shi, J.; Li, H.; Li, Z. Inhibitory Effects of Bound Polyphenol from Foxtail Millet Bran on Colitis-Associated Carcinogenesis by the Restoration of Gut Microbiota in a Mice Model. J. Agric. Food Chem. 2020, 68, 3506–3517. [Google Scholar] [CrossRef] [PubMed]
- Kubczak, M.; Szustka, A.; Rogalińska, M. Molecular Targets of Natural Compounds with Anti-Cancer Properties. Int. J. Mol. Sci. 2021, 22, 13659. [Google Scholar] [CrossRef]
- Uche, F.I.; Drijfhout, F.P.; McCullagh, J.; Richardson, A.; Li, W.-W. Cytotoxicity Effects and Apoptosis Induction by Bisbenzylisoquinoline Alkaloids from Triclisia subcordata. Phytother. Res. 2016, 30, 1533–1539. [Google Scholar] [CrossRef] [PubMed]
- Bernard, F.X.; Sablé, S.; Cameron, B.; Provost, J.; Desnottes, J.F.; Crouzet, J.; Blanche, F. Glycosylated Flavones as Selective Inhibitors of Topoisomerase IV. Antimicrob. Agents Chemother. 1997, 41, 992–998. [Google Scholar] [CrossRef] [Green Version]
- China, R.; Mukherjee, S.; Sen, S.; Bose, S.; Datta, S.; Koley, H.; Ghosh, S.; Dhar, P. Antimicrobial Activity of Sesbania Grandiflora Flower Polyphenol Extracts on Some Pathogenic Bacteria and Growth Stimulatory Effect on the Probiotic Organism Lactobacillus acidophilus. Microbiol. Res. 2012, 167, 500–506. [Google Scholar] [CrossRef]
- Muazzam, S.; Harvey, J.; Deviese, T.; Farman, M.; McCullagh, J. Targeted and Untargeted Metabolite Profiling of the Ethnobotanical Martynia annua L. Identifies Bioactive Compounds with Medicinal Properties. Planta Medica Int. Open 2018, 5, e68–e78. [Google Scholar] [CrossRef] [Green Version]
- Partouche, D.; Turbant, F.; El Hamoui, O.; Campidelli, C.; Bombled, M.; Trépout, S.; Wien, F.; Arluison, V. Epigallocatechin Gallate Remodelling of Hfq Amyloid-like Region Affects Escherichia coli Survival. Pathogens 2018, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Engels, C.; Schieber, A.; Gänzle, M.G. Inhibitory Spectra and Modes of Antimicrobial Action of Gallotannins from Mango Kernels (Mangifera indica L.). Appl. Environ. Microbiol. 2011, 77, 2215–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Din, W.M.; Jin, K.T.; Ramli, R.; Khaithir, T.M.N.; Wiart, C. Antibacterial Effects of Ellagitannins from Acalypha wilkesiana Var. macafeana Hort.: Surface Morphology Analysis with Environmental Scanning Electron Microcopy and Synergy with Antibiotics. Phytother. Res. 2013, 27, 1313–1320. [Google Scholar] [CrossRef] [PubMed]
- Yoda, Y.; Hu, Z.-Q.; Shimamura, T.; Zhao, W.-H. Different Susceptibilities of Staphylococcus and Gram-Negative Rods to Epigallocatechin Gallate. J. Infect. Chemother. 2004, 10, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.-I.; Apostolidis, E.; Labbe, R.G.; Shetty, K. Inhibition of Staphylococcus aureus by Phenolic Phytochemicals of Selected Clonal Herbs Species of Lamiaceae Family and Likely Mode of Action through Proline Oxidation. Food Biotechnol. 2007, 21, 71–89. [Google Scholar] [CrossRef]
- Caturla, N.; Vera-Samper, E.; Villalaín, J.; Mateo, C.R.; Micol, V. The Relationship between the Antioxidant and the Antibacterial Properties of Galloylated catechins and the Structure of Phospholipid Model Membranes. Free Radic. Biol. Med. 2003, 34, 648–662. [Google Scholar] [CrossRef]
- Laporta, O.; Funes, L.; Garzón, M.T.; Villalaín, J.; Micol, V. Role of Membranes on the Antibacterial and Anti-Inflammatory Activities of the Bioactive Compounds from Hypoxis rooperi Corm Extract. Arch. Biochem. Biophys. 2007, 467, 119–131. [Google Scholar] [CrossRef]
- Bernal, P.; Lemaire, S.; Pinho, M.G.; Mobashery, S.; Hinds, J.; Taylor, P.W. Insertion of Epicatechin Gallate into the Cytoplasmic Membrane of Methicillin-Resistant Staphylococcus aureus Disrupts Penicillin-Binding Protein (PBP) 2a-Mediated β-Lactam Resistance by Delocalizing PBP2. J. Biol. Chem. 2010, 285, 24055–24065. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-H.; Wang, M.-S.; Zeng, X.-A.; Xu, X.-M.; Brennan, C.S. Membrane and Genomic DNA Dual-Targeting of Citrus Flavonoid Naringenin against Staphylococcus aureus. Integr. Biol. 2017, 9, 820–829. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Truchado, P.; Larrosa, M.; Espín, J.C.; Tomás-Barberán, F.A.; Allende, A.; García-Conesa, M.T. Urolithins, Ellagitannin Metabolites Produced by Colon Microbiota, Inhibit Quorum Sensing in Yersinia enterocolitica: Phenotypic Response and Associated Molecular Changes. Food Chem. 2012, 132, 1465–1474. [Google Scholar] [CrossRef]
- Feldman, M.; Grenier, D. Cranberry Proanthocyanidins Act in Synergy with Licochalcone A to Reduce Porphyromonas gingivalis Growth and Virulence Properties, and to Suppress Cytokine Secretion by Macrophages. J. Appl. Microbiol. 2012, 113, 438–447. [Google Scholar] [CrossRef]
- Howell, A.B. Bioactive Compounds in Cranberries and Their Role in Prevention of Urinary Tract Infections. Mol. Nutr. Food Res. 2007, 51, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Uberos, J.; Iswaldi, I.; Belmonte, R.R.; Segura-Carretero, A.; Fernández-Puentes, V.; Molina-Carballo, A.; Muñoz-Hoyos, A. Cranberry (Vaccinium macrocarpon) Changes the Surface Hydrophobicity and Biofilm Formation of E. coli. Microbiol. Insights 2011, 4. [Google Scholar] [CrossRef]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Encinar, J.A.; Rodríguez-Díaz, J.C.; Micol, V. Antimicrobial Capacity of Plant Polyphenols against Gram-Positive Bacteria: A Comprehensive Review. Curr. Med. Chem. 2020, 27, 2576–2606. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.M.; Dias, R.O.; Franco, O.L. Phenolic Compounds in Antimicrobial Therapy. J. Med. Food 2017, 20, 1031–1038. [Google Scholar] [CrossRef]
- Lin, R.-D.; Chin, Y.-P.; Hou, W.-C.; Lee, M.-H. The Effects of Antibiotics Combined with Natural Polyphenols against Clinical Methicillin-Resistant Staphylococcus aureus (MRSA). Planta Med. 2008, 74, 840–846. [Google Scholar] [CrossRef]
- Swiergiel, A.H.; Dunn, A.J. The Roles of IL-1, IL-6, and TNFalpha in the Feeding Responses to Endotoxin and Influenza Virus Infection in Mice. Brain Behav. Immun. 1999, 13, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.J.; Jiang, X.R.; Mantovani, G.; Lumbreras, A.E.V.; Comi, M.; Alborali, G.; Savoini, G.; Dell’Orto, V.; Bontempo, V. Modulation of Plasma Antioxidant Activity in Weaned Piglets by Plant Polyphenols. Ital. J. Anim. Sci. 2014, 13, 3242. [Google Scholar] [CrossRef]
- Ahasan, A.S.M.L.; Invernizzi, G.; Farina, G.; Pilotto, A.; Barbé, F.; Bontempo, V.; Rossi, R.; Bellagamba, F.; Lecchi, C.; Savoini, G.; et al. The Effects of Superoxide Dismutase-Rich Melon Pulp Concentrate on Inflammation, Antioxidant Status and Growth Performance of Challenged Post-Weaning Piglets. Animal 2019, 13, 136–143. [Google Scholar] [CrossRef]
- Ryu, E. Prophylactic Effect of Tea on Pathogenic Micro-Organism Infection to Human and Animals. (1). Growth Inhibitive and Bacteriocidal Effect of Tea on Food Poisoning and Other Pathogenic Enterobacterium in Vitro. Int. J. Zoonoses 1980, 7, 164–170. [Google Scholar]
- Skaperda, Z.; Veskoukis, A.S.; Kouretas, D. Farm Animal Welfare, Productivity and Meat Quality: Interrelation with Redox Status Regulation and Antioxidant Supplementation as a Nutritional Intervention (Review). World Acad. Sci. J. 2019, 1, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and Antioxidants in Disease: Oxidative Stress in Farm Animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Eriksson, M. Vitamin E in Relation to Lipid Peroxidation in Experimental Septic Shock. Prostaglandins Leukot. Essent. Fat. Acids PLEFA 2000, 62, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Eriksson, M. Retinol Palmitate Counteracts Oxidative Injury during Experimental Septic Shock. Ann. Acad. Med. Singap. 2001, 30, 265–269. [Google Scholar] [PubMed]
- Basu, S.; Mutschler, D.K.; Larsson, A.O.; Kiiski, R.; Nordgren, A.; Eriksson, M.B. Propofol (Diprivan-EDTA) Counteracts Oxidative Injury and Deterioration of the Arterial Oxygen Tension during Experimental Septic Shock. Resuscitation 2001, 50, 341–348. [Google Scholar] [CrossRef]
- Bruins, M.J.; Luiking, Y.C.; Soeters, P.B.; Lamers, W.H.; Akkermans, L.M.A.; Deutz, N.E.P. Effects of Long-Term Intravenous and Intragastric l-Arginine Intervention on Jejunal Motility and Visceral Nitric Oxide Production in the Hyperdynamic Compensated Endotoxaemic Pig. Neurogastroenterol. Motil. 2004, 16, 819–828. [Google Scholar] [CrossRef]
- Matejovic, M.; Krouzecky, A.; Martinkova, V.; Rokyta, R.J.; Kralova, H.; Treska, V.; Radermacher, P.; Novak, I. Selective Inducible Nitric Oxide Synthase Inhibition during Long-Term Hyperdynamic Porcine Bacteremia. Shock 2004, 21, 458–465. [Google Scholar] [CrossRef]
- Miller, M.J.S.; Zhang, X.J.; Sadowska-Krowicka, H.; Chotinaruemol, S.; McIntyre, J.A.; Clark, D.A.; Bustamante, S.A. Nitric Oxide Release in Response to Gut Injury. Scand. J. Gastroenterol. 1993, 28, 149–154. [Google Scholar] [CrossRef]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.F.; Blisklager, A.T.; Moeser, A.J. Early Weaning Stress Impairs Development of Mucosal Barrier Function in the Porcine Intestine. Am. J. Physiol.-Gastrointest. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef] [Green Version]
- Moeser, A.J.; Ryan, K.A.; Nighot, P.K.; Blikslager, A.T. Gastrointestinal Dysfunction Induced by Early Weaning Is Attenuated by Delayed Weaning and Mast Cell Blockade in Pigs. Am. J. Physiol.-Gastrointest. Liver Physiol. 2007, 293, G413–G421. [Google Scholar] [CrossRef] [Green Version]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites Produced by Commensal Bacteria Promote Peripheral Regulatory T-Cell Generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Johnson, R.W. Immune and Endocrine Regulation of Food Intake in Sick Animals. Domest. Anim. Endocrinol. 1998, 15, 309–319. [Google Scholar] [CrossRef]
- Burfeind, K.G.; Michaelis, K.A.; Marks, D.L. The Central Role of Hypothalamic Inflammation in the Acute Illness Response and Cachexia. Semin. Cell Dev. Biol. 2016, 54, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors Influencing the Structure and Function of the Small Intestine in the Weaned Pig: A Review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- McCracken, B.A.; Spurlock, M.E.; Roos, M.A.; Zuckermann, F.A.; Gaskins, H.R. Weaning Anorexia May Contribute to Local Inflammation in the Piglet Small Intestine. J. Nutr. 1999, 129, 613–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pié, S.; Lallès, J.P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning Is Associated with an Upregulation of Expression of Inflammatory Cytokines in the Intestine of Piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [Green Version]
- McLamb, B.L.; Gibson, A.J.; Overman, E.L.; Stahl, C.; Moeser, A.J. Early Weaning Stress in Pigs Impairs Innate Mucosal Immune Responses to Enterotoxigenic E. coli Challenge and Exacerbates Intestinal Injury and Clinical Disease. PLoS ONE 2013, 8, e59838. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, S.; Ringseis, R.; Hillen, S.; Becker, S.; Erhardt, G.; Reiner, G.; Eder, K. Genome-Wide Transcript Profiling Indicates Induction of Energy-Generating Pathways and an Adaptive Immune Response in the Liver of Sows during Lactation. Comp. Biochem. Physiol. Part D Genom. Proteom. 2012, 7, 370–381. [Google Scholar] [CrossRef]
- Rosenbaum, S.; Ringseis, R.; Hillen, S.; Becker, S.; Erhardt, G.; Reiner, G.; Eder, K. The Stress Signalling Pathway Nuclear Factor E2-Related Factor 2 Is Activated in the Liver of Sows during Lactation. Acta Vet. Scand. 2012, 54, 59. [Google Scholar] [CrossRef] [Green Version]
- Gessner, D.K.; Gröne, B.; Couturier, A.; Rosenbaum, S.; Hillen, S.; Becker, S.; Erhardt, G.; Reiner, G.; Ringseis, R.; Eder, K. Dietary Fish Oil Inhibits Pro-Inflammatory and ER Stress Signalling Pathways in the Liver of Sows during Lactation. PLoS ONE 2015, 10, e0137684. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; García-Villanova, B.; Guerra-Hernández, E.; Verardo, V. Grape Seeds Proanthocyanidins: An Overview of In Vivo Bioactivity in Animal Models. Nutrients 2019, 11, 2435. [Google Scholar] [CrossRef] [Green Version]
- Kafantaris, I.; Stagos, D.; Kotsampasi, B.; Hatzis, A.; Kypriotakis, A.; Gerasopoulos, K.; Makri, S.; Goutzourelas, N.; Mitsagga, C.; Giavasis, I.; et al. Grape Pomace Improves Performance, Antioxidant Status, Fecal Microbiota and Meat Quality of Piglets. Animal 2018, 12, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Kim, I.H. Effect of Dietary Grape Pomace Fermented by Saccharomyces boulardii on the Growth Performance, Nutrient Digestibility and Meat Quality in Finishing Pigs. Asian-Australas. J. Anim. Sci. 2011, 24, 1763–1770. [Google Scholar] [CrossRef]
- Taranu, I.; Gras, M.A.; Habeanu, M.; Pistol, G.C.; Lefter, N.; Palade, M.L.; Ropota, M.; Chedea, V.S.; Marin, D.E. Active Ingredients from Oil By-Products Modulate Spleen Inflammatory and Antioxidant Response in Pigs. Arch. Zootech. 2020, 23, 81–97. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, G.; Kebreab, E.; Yu, Q.; Li, J.; Zhang, X.; He, H.; Fang, R.; Dai, Q. Effects of Dietary Grape Seed Polyphenols Supplementation during Late Gestation and Lactation on Antioxidant Status in Serum and Immunoglobulin Content in Colostrum of Multiparous Sows. J. Anim. Sci. 2019, 97, 2515–2523. [Google Scholar] [CrossRef] [PubMed]
- Hao, R.; Li, Q.; Zhao, J.; Li, H.; Wang, W.; Gao, J. Effects of Grape Seed Procyanidins on Growth Performance, Immune Function and Antioxidant Capacity in Weaned Piglets. Livest. Sci. 2015, 178, 237–242. [Google Scholar] [CrossRef]
- He, P.; Hua, H.; Tian, W.; Zhu, H.; Liu, Y.; Xu, X. Holly (Ilex latifolia Thunb.) Polyphenols Extracts Alleviate Hepatic Damage by Regulating Ferroptosis Following Diquat Challenge in a Piglet Model. Front. Nutr. 2020, 7, 604328. [Google Scholar] [CrossRef]
- Ahmed, S.T.; Hossain, M.E.; Kim, G.M.; Hwang, J.A.; Ji, H.; Yang, C.J. Effects of Resveratrol and Essential Oils on Growth Performance, Immunity, Digestibility and Fecal Microbial Shedding in Challenged Piglets. Asian-Australas. J. Anim. Sci. 2013, 26, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Diao, H.; Zheng, P.; Yu, B.; He, J.; Mao, X.; Yu, J.; Chen, D. Effects of Benzoic Acid and Thymol on Growth Performance and Gut Characteristics of Weaned Piglets. Asian-Australas. J. Anim. Sci. 2015, 28, 827–839. [Google Scholar] [CrossRef] [Green Version]
- Lallès, J.-P.; Lacan, D.; David, J.-C. A Melon Pulp Concentrate Rich in Superoxide Dismutase Reduces Stress Proteins along the Gastrointestinal Tract of Pigs. Nutrition 2011, 27, 358–363. [Google Scholar] [CrossRef]
- Bruno, D.G.; Martins, S.M.M.K.; Parazzi, L.J.; Afonso, E.R.; Del Santo, T.A.; de Teixeira, S.M.N.; Moreno, A.M.; de Moretti, A.S. Phytogenic Feed Additives in Piglets Challenged with Salmonella Typhimurium. Rev. Bras. Zootec. 2013, 42, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.B.; Luciano, F.B.; Miyada, V.S.; Gois, F.D. Herbal Extracts and Organic Acids as Natural Feed Additives in Pig Diets. S. Afr. J. Anim. Sci. 2013, 43, 181–193. [Google Scholar] [CrossRef]
- Kaschubek, T.; Mayer, E.; Rzesnik, S.; Grenier, B.; Bachinger, D.; Schieder, C.; König, J.; Teichmann, K. Effects of Phytogenic Feed Additives on Cellular Oxidative Stress and Inflammatory Reactions in Intestinal Porcine Epithelial Cells1. J. Anim. Sci. 2018, 96, 3657–3669. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, W.; Wang, L.; Chen, Y.; Zhang, H.; Wang, T.; Yang, X.; Xing, F.; Yan, J.; Fang, X. Protective Effects of Fucoidan against Hydrogen Peroxide-Induced Oxidative Damage in Porcine Intestinal Epithelial Cells. Animals 2019, 9, 1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buettner, G.R. The Pecking Order of Free Radicals and Antioxidants: Lipid Peroxidation, Alpha-Tocopherol, and Ascorbate. Arch. Biochem. Biophys. 1993, 300, 535–543. [Google Scholar] [CrossRef]
- Frank, J. Beyond Vitamin E Supplementation: An Alternative Strategy to Improve Vitamin E Status. J. Plant Physiol. 2005, 162, 834–843. [Google Scholar] [CrossRef]
- Valenzuela-Grijalva, N.V.; Pinelli-Saavedra, A.; Muhlia-Almazan, A.; Domínguez-Díaz, D.; González-Ríos, H. Dietary Inclusion Effects of Phytochemicals as Growth Promoters in Animal Production. J. Anim. Sci. Technol. 2017, 59, 8. [Google Scholar] [CrossRef] [Green Version]
- Beghelli, D.; de Cosmo, A.; Faeti, V.; Lupidi, G.; Bailetti, L.; Cavallucci, C.; Polidori, P. Origanum Vulgare L. and Rosmarinus Officinalis L. Aqueous Extracts in Growing-Finishing Pig Nutrition: Effects on Antioxidant Status, Immune Responses, Polyphenolic Content and Sensorial Properties. J. Food Res. 2019, 8, 90–99. [Google Scholar] [CrossRef] [Green Version]
- Serra, V.; Salvatori, G.; Pastorelli, G. Dietary Polyphenol Supplementation in Food Producing Animals: Effects on the Quality of Derived Products. Animals 2021, 11, 401. [Google Scholar] [CrossRef]
- Sjögren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Bäckhed, F.; Ohlsson, C. The Gut Microbiota Regulates Bone Mass in Mice. J. Bone Miner. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef] [Green Version]
- Novince, C.M.; Whittow, C.R.; Aartun, J.D.; Hathaway, J.D.; Poulides, N.; Chavez, M.B.; Steinkamp, H.M.; Kirkwood, K.A.; Huang, E.; Westwater, C.; et al. Commensal Gut Microbiota Immunomodulatory Actions in Bone Marrow and Liver Have Catabolic Effects on Skeletal Homeostasis in Health. Sci. Rep. 2017, 7, 5747. [Google Scholar] [CrossRef] [Green Version]
- Warner, A.J.; Hathaway-Schrader, J.D.; Lubker, R.; Davies, C.; Novince, C.M. Tetracyclines and Bone: Unclear Actions with Potentially Lasting Effects. Bone 2022, 159, 116377. [Google Scholar] [CrossRef] [PubMed]
- Frost, H.M. Tetracyclines and Fetal Bones. Henry Ford Hosp. Med. J. 1965, 13, 403–410. [Google Scholar] [PubMed]
- Steckoll, S.H.; Goffer, Z.; Haas, N.; Nathan, H. Red Stained Bones from Qumran. Nature 1971, 231, 469–470. [Google Scholar] [CrossRef]
- Bassett, E.; Keith, M.; Armelagos, G.; Martin, D.; Villanueva, A. Tetracycline-Labeled Human Bone from Ancient Sudanese Nubia (A.D. 350). Science 1980, 209, 1532–1534. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.; Molto, E.; Anderson, C. Fluorochrome Labelling in Roman Period Skeletons from Dakhleh Oasis, Egypt. Am. J. Phys. Anthropol. 1989, 80, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Bensimon-Brito, A.; Cardeira, J.; Dionísio, G.; Huysseune, A.; Cancela, M.L.; Witten, P.E. Revisiting in Vivo Staining with Alizarin Red S—A Valuable Approach to Analyse Zebrafish Skeletal Mineralization during Development and Regeneration. BMC Dev. Biol. 2016, 16, 2. [Google Scholar] [CrossRef] [Green Version]
- Frost, H.M. Relation between Bone Tissue and Cell Population Dynamics, Histology and Tetracycline Labeling. Clin. Orthop. Relat. Res. 1966, 49, 65–76. [Google Scholar] [CrossRef]
- Myers, H.M. Alizarin and Tetracycline Binding by Bone Mineral. Am. J. Phys. Anthropol. 1968, 29, 179–182. [Google Scholar] [CrossRef]
- Rigueur, D.; Lyons, K.M. Whole-Mount Skeletal Staining. Methods Mol. Biol. 2014, 1130, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Nuñez-Sánchez, M.A.; García-Villalba, R.; Monedero-Saiz, T.; García-Talavera, N.V.; Gómez-Sánchez, M.B.; Sánchez-Álvarez, C.; García-Albert, A.M.; Rodríguez-Gil, F.J.; Ruiz-Marín, M.; Pastor-Quirante, F.A.; et al. Targeted Metabolic Profiling of Pomegranate Polyphenols and Urolithins in Plasma, Urine and Colon Tissues from Colorectal Cancer Patients. Mol. Nutr. Food Res. 2014, 58, 1199–1211. [Google Scholar] [CrossRef]
- Hedges, R.E.M.; Clement, J.G.; Thomas, C.D.L.; O’Connell, T.C. Collagen Turnover in the Adult Femoral Mid-Shaft: Modeled from Anthropogenic Radiocarbon Tracer Measurements. Am. J. Phys. Anthropol. 2007, 133, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Clarke, E.D.; Rollo, M.E.; Collins, C.E.; Wood, L.; Callister, R.; Philo, M.; Kroon, P.A.; Haslam, R.L. The Relationship between Dietary Polyphenol Intakes and Urinary Polyphenol Concentrations in Adults Prescribed a High Vegetable and Fruit Diet. Nutrients 2020, 12, 3431. [Google Scholar] [CrossRef] [PubMed]
- Stenhouse, M.J.; Baxter, M.S. The Uptake of Bomb 14C in Humans. In Radiocarbon Dating: Proceedings of the Ninth International Conference Los Angeles and La Jolla, 1979; University of California Press: Berkeley, CA, USA, 1979; pp. 324–341. [Google Scholar]
- Geyh, M.A. Bomb Radiocarbon Dating of Animal Tissues and Hair. Radiocarbon 2001, 43, 723–730. [Google Scholar] [CrossRef] [Green Version]
- Caplan, A.I. Mesenchymal Stem Cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Spencer, J.A.; Koh, B.I.; Kobayashi, T.; Fujisaki, J.; Clemens, T.L.; Lin, C.P.; Kronenberg, H.M.; Scadden, D.T. Endogenous Bone Marrow MSCs Are Dynamic, Fate-Restricted Participants in Bone Maintenance and Regeneration. Cell Stem Cell 2012, 10, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Blair, H.C.; Larrouture, Q.C.; Li, Y.; Lin, H.; Beer-Stoltz, D.; Liu, L.; Tuan, R.S.; Robinson, L.J.; Schlesinger, P.H.; Nelson, D.J. Osteoblast Differentiation and Bone Matrix Formation In Vivo and In Vitro. Tissue Eng. Part B Rev. 2017, 23, 268–280. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yang, Y.; Liu, L.; Blair, H.C.; Friedman, P.A. NHERF1 Regulation of PTH-Dependent Bimodal Pi Transport in Osteoblasts. Bone 2013, 52, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Gomes, C.M.F.; van Paassen, H.; Romeo, S.; Welling, M.M.; Feitsma, R.I.J.; Abrunhosa, A.J.; Botelho, M.F.; Hogendoorn, P.C.W.; Pauwels, E.; Cleton-Jansen, A.M. Multidrug Resistance Mediated by ABC Transporters in Osteosarcoma Cell Lines: MRNA Analysis and Functional Radiotracer Studies. Nucl. Med. Biol. 2006, 33, 831–840. [Google Scholar] [CrossRef]
- Monjo, M.; Rubert, M.; Ellingsen, J.E.; Lyngstadaas, S.P. Rosuvastatin Promotes Osteoblast Differentiation and Regulates SLCO1A1 Transporter Gene Expression in MC3T3-E1 Cells. Cell. Physiol. Biochem. 2010, 26, 647–656. [Google Scholar] [CrossRef]
- Verstraeten, S.V.; Keen, C.L.; Schmitz, H.H.; Fraga, C.G.; Oteiza, P.I. Flavan-3-Ols and Procyanidins Protect Liposomes against Lipid Oxidation and Disruption of the Bilayer Structure. Free Radic. Biol. Med. 2003, 34, 84–92. [Google Scholar] [CrossRef]
- Sirk, T.W.; Brown, E.F.; Sum, A.K.; Friedman, M. Molecular Dynamics Study on the Biophysical Interactions of Seven Green Tea Catechins with Lipid Bilayers of Cell Membranes. J. Agric. Food Chem. 2008, 56, 7750–7758. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.T.T.; Yoda, T.; Chahal, B.; Morita, M.; Takagi, M.; Vestergaard, M.C. Structure-Dependent Interactions of Polyphenols with a Biomimetic Membrane System. Biochim. Biophys. Acta-Biomembr. 2014, 1838, 2670–2677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, H.C.; Garimella, R.; Tague, S.E. The Role of Matrix Vesicles in Growth Plate Development and Biomineralization. Front. Biosci. J. Virtual Libr. 2005, 10, 822–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golub, E.E. Role of Matrix Vesicles in Biomineralization. Biochim. Biophys. Acta 2009, 1790, 1592–1598. [Google Scholar] [CrossRef] [Green Version]
- Plonska-Brzezinska, M.E.; Bobrowska, D.M.; Sharma, A.; Rodziewicz, P.; Tomczyk, M.; Czyrko, J.; Brzezinski, K. Triple Helical Collagen-like Peptide Interactions with Selected Polyphenolic Compounds. RSC Adv. 2015, 5, 95443–95453. [Google Scholar] [CrossRef]
- Wu, L.; Shao, H.; Fang, Z.; Zhao, Y.; Cao, C.Y.; Li, Q. Mechanism and Effects of Polyphenol Derivatives for Modifying Collagen. ACS Biomater. Sci. Eng. 2019, 5, 4272–4284. [Google Scholar] [CrossRef]
- Choi, Y.; Kim, H.-J.; Min, K.-S. Effects of Proanthocyanidin, a Crosslinking Agent, on Physical and Biological Properties of Collagen Hydrogel Scaffold. Restor. Dent. Endod. 2016, 41, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Russo, N.; Cassinelli, C.; Torre, E.; Morra, M.; Iviglia, G. Improvement of the Physical Properties of Guided Bone Regeneration Membrane from Porcine Pericardium by Polyphenols-Rich Pomace Extract. Materials 2019, 12, 2564. [Google Scholar] [CrossRef] [Green Version]
- Reddy, R.R.; Phani Kumar, B.V.N.; Shanmugam, G.; Madhan, B.; Mandal, A.B. Molecular Level Insights on Collagen–Polyphenols Interaction Using Spin–Relaxation and Saturation Transfer Difference NMR. J. Phys. Chem. B 2015, 119, 14076–14085. [Google Scholar] [CrossRef]
- Viguet-Carrin, S.; Garnero, P.; Delmas, P.D. The Role of Collagen in Bone Strength. Osteoporos. Int. 2006, 17, 319–336. [Google Scholar] [CrossRef]
- Hagerman, A.E.; Butler, L.G. The Specificity of Proanthocyanidin-Protein Interactions. J. Biol. Chem. 1981, 256, 4494–4497. [Google Scholar] [CrossRef]
- Velmurugan, P.; Singam, E.R.A.; Jonnalagadda, R.R.; Subramanian, V. Investigation on Interaction of Tannic Acid with Type I Collagen and Its Effect on Thermal, Enzymatic, and Conformational Stability for Tissue Engineering Applications. Biopolymers 2014, 101, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Shavandi, A.; Bekhit, A.E.-D.A.; Saeedi, P.; Izadifar, Z.; Bekhit, A.A.; Khademhosseini, A. Polyphenol Uses in Biomaterials Engineering. Biomaterials 2018, 167, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Madhan, B.; Subramanian, V.; Rao, J.R.; Nair, B.U.; Ramasami, T. Stabilization of Collagen Using Plant Polyphenol: Role of Catechin. Int. J. Biol. Macromol. 2005, 37, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.; Zhao, J.R.; Wong, W.; Burt, H. The Inhibition of Collagenase Induced Degradation of Collagen by the Galloyl-Containing Polyphenols Tannic Acid, Epigallocatechin Gallate and Epicatechin Gallate. J. Mater. Sci. Mater. Med. 2010, 21, 1435–1443. [Google Scholar] [CrossRef]
- Olde Damink, L.H.H.; Dijkstra, P.J.; Van Luyn, M.J.A.; Van Wachem, P.B.; Nieuwenhuis, P.; Feijen, J. Glutaraldehyde as a Crosslinking Agent for Collagen-Based Biomaterials. J. Mater. Sci. Mater. Med. 1995, 6, 460–472. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.R.; Covington, A.D.; Hancock, R.A. Structure–Activity Relationships in the Hydrophobic Interactions of Polyphenols with Cellulose and Collagen. Biopolymers 2003, 70, 403–413. [Google Scholar] [CrossRef]
- He, L.; Mu, C.; Shi, J.; Zhang, Q.; Shi, B.; Lin, W. Modification of Collagen with a Natural Cross-Linker, Procyanidin. Int. J. Biol. Macromol. 2011, 48, 354–359. [Google Scholar] [CrossRef]
- Sinder, B.P.; Pettit, A.R.; McCauley, L.K. Macrophages: Their Emerging Roles in Bone. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2015, 30, 2140–2149. [Google Scholar] [CrossRef] [Green Version]
- Brandao-Burch, A.; Utting, J.C.; Orriss, I.R.; Arnett, T.R. Acidosis Inhibits Bone Formation by Osteoblasts In Vitro by Preventing Mineralization. Calcif. Tissue Int. 2005, 77, 167–174. [Google Scholar] [CrossRef]
- Muzylak, M.; Arnett, T.R.; Price, J.S.; Horton, M.A. The in Vitro Effect of PH on Osteoclasts and Bone Resorption in the Cat: Implications for the Pathogenesis of FORL. J. Cell. Physiol. 2007, 213, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Arnett, T.R. Extracellular PH Regulates Bone Cell Function. J. Nutr. 2008, 138, 415S–418S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parfitt, A.M. The Coupling of Bone Formation to Bone Resorption: A Critical Analysis of the Concept and of Its Relevance to the Pathogenesis of Osteoporosis. Metab. Bone Dis. Relat. Res. 1982, 4, 1–6. [Google Scholar] [CrossRef]
- Eriksen, E.F. Normal and Pathological Remodeling of Human Trabecular Bone: Three Dimensional Reconstruction of the Remodeling Sequence in Normals and in Metabolic Bone Disease. Endocr. Rev. 1986, 7, 379–408. [Google Scholar] [CrossRef] [PubMed]
- Đudarić, L.; Fužinac-Smojver, A.; Muhvić, D.; Giacometti, J. The Role of Polyphenols on Bone Metabolism in Osteoporosis. Food Res. Int. 2015, 77, 290–298. [Google Scholar] [CrossRef]
- Lin, S.-Y.; Kang, L.; Wang, C.-Z.; Huang, H.H.; Cheng, T.-L.; Huang, H.-T.; Lee, M.-J.; Lin, Y.-S.; Ho, M.-L.; Wang, G.-J.; et al. (-)-Epigallocatechin-3-Gallate (EGCG) Enhances Osteogenic Differentiation of Human Bone Marrow Mesenchymal Stem Cells. Molecules 2018, 23, 3221. [Google Scholar] [CrossRef] [Green Version]
- Delaisse, J.-M.; Andersen, T.L.; Kristensen, H.B.; Jensen, P.R.; Andreasen, C.M.; Søe, K. Re-Thinking the Bone Remodeling Cycle Mechanism and the Origin of Bone Loss. Bone 2020, 141, 115628. [Google Scholar] [CrossRef]
- Huang, H.-T.; Cheng, T.-L.; Lin, S.-Y.; Ho, C.-J.; Chyu, J.Y.; Yang, R.-S.; Chen, C.-H.; Shen, C.-L. Osteoprotective Roles of Green Tea Catechins. Antioxidants 2020, 9, 1136. [Google Scholar] [CrossRef]
- Almeida, M.; O’Brien, C.A. Basic Biology of Skeletal Aging: Role of Stress Response Pathways. J. Gerontol.-Ser. Biol. Sci. Med. Sci. 2013, 68, 1197–1208. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Han, L.; Martin-Millan, M.; Plotkin, L.I.; Stewart, S.A.; Roberson, P.K.; Kousteni, S.; O’Brien, C.A.; Bellido, T.; Parfitt, A.M.; et al. Skeletal Involution by Age-Associated Oxidative Stress and Its Acceleration by Loss of Sex Steroids. J. Biol. Chem. 2007, 282, 27285–27297. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Ma, W.; Dong, H.; Yong, Z.; Su, R. Establishing a Rapid Animal Model of Osteoporosis with Ovariectomy plus Low Calcium Diet in Rats. Int. J. Clin. Exp. Pathol. 2014, 7, 5123–5128. [Google Scholar] [PubMed]
- Li, J.; Karim, M.A.; Che, H.; Geng, Q.; Miao, D. Deletion of P16 Prevents Estrogen Deficiency-Induced Osteoporosis by Inhibiting Oxidative Stress and Osteocyte Senescence. Am. J. Transl. Res. 2020, 12, 672–683. [Google Scholar] [PubMed]
- Basu, S.; Michaëlsson, K.; Olofsson, H.; Johansson, S.; Melhus, H. Association between Oxidative Stress and Bone Mineral Density. Biochem. Biophys. Res. Commun. 2001, 288, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Bonaccorsi, G.; Cremonini, E.; Romani, A.; Fila, E.; Castaldini, M.C.; Ferrazzini, S.; Giganti, M.; Massari, L. Oxidative Stress and Bone Resorption Interplay as a Possible Trigger for Postmenopausal Osteoporosis. BioMed Res. Int. 2014, 2014, 569563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Key, L.L.; Wolf, W.C.; Gundberg, C.M.; Ries, W.L. Superoxide and Bone Resorption. Bone 1994, 15, 431–436. [Google Scholar] [CrossRef]
- Darden, A.G.; Ries, W.L.; Wolf, W.C.; Rodriguiz, R.M.; Key, L.L. Osteoclastic Superoxide Production and Bone Resorption: Stimulation and Inhibition by Modulators of NADPH Oxidase. J. Bone Miner. Res. 1996, 11, 671–675. [Google Scholar] [CrossRef]
- Lean, J.M.; Jagger, C.J.; Kirstein, B.; Fuller, K.; Chambers, T.J. Hydrogen Peroxide Is Essential for Estrogen-Deficiency Bone Loss and Osteoclast Formation. Endocrinology 2005, 146, 728–735. [Google Scholar] [CrossRef]
- Ru, J.; Wang, Y. Osteocyte Apoptosis: The Roles and Key Molecular Mechanisms in Resorption-Related Bone Diseases. Cell Death Dis. 2020, 11, 846. [Google Scholar] [CrossRef]
- Weitzmann, M.N.; Pacifici, R. Estrogen Deficiency and Bone Loss: An Inflammatory Tale. J. Clin. Investig. 2006, 116, 1186–1194. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Takahashi, N.; Jimi, E.; Udagawa, N.; Takami, M.; Kotake, S.; Nakagawa, N.; Kinosaki, M.; Yamaguchi, K.; Shima, N.; et al. Tumor Necrosis Factor Alpha Stimulates Osteoclast Differentiation by a Mechanism Independent of the ODF/RANKL-RANK Interaction. J. Exp. Med. 2000, 191, 275–286. [Google Scholar] [CrossRef]
- Gilbert, L.; He, X.; Farmer, P.; Rubin, J.; Drissi, H.; van Wijnen, A.J.; Lian, J.B.; Stein, G.S.; Nanes, M.S. Expression of the Osteoblast Differentiation Factor RUNX2 (Cbfa1/AML3/Pebp2alpha A) Is Inhibited by Tumor Necrosis Factor-Alpha. J. Biol. Chem. 2002, 277, 2695–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cenci, S.; Weitzmann, M.N.; Roggia, C.; Namba, N.; Novack, D.; Woodring, J.; Pacifici, R. Estrogen Deficiency Induces Bone Loss by Enhancing T-Cell Production of TNF-α. J. Clin. Investig. 2000, 106, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Heino, T.J.; Hentunen, T.A.; Väänänen, H.K. Osteocytes Inhibit Osteoclastic Bone Resorption through Transforming Growth Factor-β: Enhancement by Estrogen. J. Cell. Biochem. 2002, 85, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Léotoing, L.; Wauquier, F.; Guicheux, J.; Miot-Noirault, E.; Wittrant, Y.; Coxam, V. The Polyphenol Fisetin Protects Bone by Repressing NF-κB and MKP-1-Dependent Signaling Pathways in Osteoclasts. PLoS ONE 2013, 8, e68388. [Google Scholar] [CrossRef] [Green Version]
- Austermann, K.; Baecker, N.; Stehle, P.; Heer, M. Putative Effects of Nutritive Polyphenols on Bone Metabolism in Vivo—Evidence from Human Studies. Nutrients 2019, 11, 871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.-L.; Yeh, J.K.; Cao, J.J.; Tatum, O.L.; Dagda, R.Y.; Wang, J.-S. Green Tea Polyphenols Mitigate Bone Loss of Female Rats in a Chronic Inflammation-Induced Bone Loss Model. J. Nutr. Biochem. 2010, 21, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lazarenko, O.P.; Kang, J.; Blackburn, M.L.; Ronis, M.J.J.; Badger, T.M.; Chen, J.-R. Feeding Blueberry Diets to Young Rats Dose-Dependently Inhibits Bone Resorption through Suppression of RANKL in Stromal Cells. PLoS ONE 2013, 8, e70438. [Google Scholar] [CrossRef]
- Byun, M.R.; Sung, M.K.; Kim, A.R.; Lee, C.H.; Jang, E.J.; Jeong, M.G.; Noh, M.; Hwang, E.S.; Hong, J.-H. (-)-Epicatechin Gallate (ECG) Stimulates Osteoblast Differentiation via Runt-Related Transcription Factor 2 (RUNX2) and Transcriptional Coactivator with PDZ-Binding Motif (TAZ)-Mediated Transcriptional Activation. J. Biol. Chem. 2014, 289, 9926–9935. [Google Scholar] [CrossRef] [Green Version]
- Wattel, A.; Kamel, S.; Prouillet, C.; Petit, J.-P.; Lorget, F.; Offord, E.; Brazier, M. Flavonoid Quercetin Decreases Osteoclastic Differentiation Induced by RANKL via a Mechanism Involving NFκB and AP-1. J. Cell. Biochem. 2004, 92, 285–295. [Google Scholar] [CrossRef]
- Van der Woude, H.; Ter Veld, M.G.R.; Jacobs, N.; van der Saag, P.T.; Murk, A.J.; Rietjens, I.M.C.M. The Stimulation of Cell Proliferation by Quercetin Is Mediated by the Estrogen Receptor. Mol. Nutr. Food Res. 2005, 49, 763–771. [Google Scholar] [CrossRef]
- Weaver, C.M.; Alekel, D.L.; Ward, W.E.; Ronis, M.J. Flavonoid Intake and Bone Health. J. Nutr. Gerontol. Geriatr. 2012, 31, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Figueira, M.-E.; Oliveira, M.; Direito, R.; Rocha, J.; Alves, P.; Serra, A.-T.; Duarte, C.; Bronze, R.; Fernandes, A.; Brites, D.; et al. Protective Effects of a Blueberry Extract in Acute Inflammation and Collagen-Induced Arthritis in the Rat. Biomed. Pharmacother. 2016, 83, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Bu, S.Y.; Hunt, T.S.; Smith, B.J. Dried Plum Polyphenols Attenuate the Detrimental Effects of TNF-α on Osteoblast Function Coincident with up-Regulation of Runx2, Osterix and IGF-I. J. Nutr. Biochem. 2009, 20, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Mora, R.; Casado-Díaz, A.; De Castro, M.D.; Quesada-Gómez, J.M. Oleuropein Enhances Osteoblastogenesis and Inhibits Adipogenesis: The Effect on Differentiation in Stem Cells Derived from Bone Marrow. Osteoporos. Int. 2011, 22, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-R.; Lazarenko, O.P.; Wu, X.; Kang, J.; Blackburn, M.L.; Shankar, K.; Badger, T.M.; Ronis, M.J. Dietary-Induced Serum Phenolic Acids Promote Bone Growth via P38 MAPK/β-Catenin Canonical Wnt Signaling. J. Bone Miner. Res. 2010, 25, 2399–2411. [Google Scholar] [CrossRef]
- Li, X.; Udagawa, N.; Itoh, K.; Suda, K.; Murase, Y.; Nishihara, T.; Suda, T.; Takahashi, N. P38 MAPK-Mediated Signals Are Required for Inducing Osteoclast Differentiation but Not for Osteoclast Function. Endocrinology 2002, 143, 3105–3113. [Google Scholar] [CrossRef]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. P38 MAPK Signaling in Osteoblast Differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Blair, H.C.; Kalyvioti, E.; Papachristou, N.I.; Tourkova, I.L.; Syggelos, S.A.; Deligianni, D.; Orkoula, M.G.; Kontoyannis, C.G.; Karavia, E.A.; Kypreos, K.E.; et al. Apolipoprotein A-1 Regulates Osteoblast and Lipoblast Precursor Cells in Mice. Lab. Investig. 2016, 96, 763–772. [Google Scholar] [CrossRef]
- Papachristou, N.I.; Blair, H.C.; Kypreos, K.E.; Papachristou, D.J. High-Density Lipoprotein (HDL) Metabolism and Bone Mass. J. Endocrinol. 2017, 233, R95–R107. [Google Scholar] [CrossRef]
- Romero, G.; Sneddon, W.B.; Yang, Y.; Wheeler, D.; Blair, H.C.; Friedman, P.A. Parathyroid Hormone Receptor Directly Interacts with Dishevelled to Regulate Beta-Catenin Signaling and Osteoclastogenesis. J. Biol. Chem. 2010, 285, 14756–14763. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.H.; Park, S.-Y.; Baek, J.-E.; Lee, S.-Y.; Baek, W.-Y.; Lee, S.-Y.; Lee, Y.-S.; Yoo, H.J.; Kim, H.; Lee, S.H.; et al. Free Fatty Acid Receptor 4 (GPR120) Stimulates Bone Formation and Suppresses Bone Resorption in the Presence of Elevated n-3 Fatty Acid Levels. Endocrinology 2016, 157, 2621–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCullagh, J.S.O. Isotopic Analysis at the Molecular Level: A Frontier in Palaeodietary Reconstruction. Oxf. Univ. Sch. Archaeol. Monogr. 2007, 66, 73. [Google Scholar]
- Pollard, A.M.; Ditchfield, P.; McCullagh, J.S.O.; Allen, T.G.; Gibson, M.; Boston, C.; Clough, S.; Marquez-Grant, N.; Nicholson, R.A. “These Boots Were Made for Walking”: The Isotopic Analysis of a C4 Roman Inhumation from Gravesend, Kent, UK. Am. J. Phys. Anthropol. 2011, 146, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Nalawade-Chavan, S.; McCullagh, J.; Hedges, R.; Bonsall, C.; Boroneanţ, A.; Ramsey, C.B.; Higham, T. Compound-Specific Radiocarbon Dating of Essential and Non-Essential Amino Acids: Towards Determination of Dietary Reservoir Effects in Humans. Radiocarbon 2013, 55, 709–719. [Google Scholar] [CrossRef] [Green Version]
- Lee-Thorp, J.A. On Isotopes and Old Bones. Archaeometry 2008, 50, 925–950. [Google Scholar] [CrossRef]
- Wibowo, M.C.; Yang, Z.; Borry, M.; Hübner, A.; Huang, K.D.; Tierney, B.T.; Zimmerman, S.; Barajas-Olmos, F.; Contreras-Cubas, C.; García-Ortiz, H.; et al. Reconstruction of Ancient Microbial Genomes from the Human Gut. Nature 2021, 594, 234–239. [Google Scholar] [CrossRef] [PubMed]


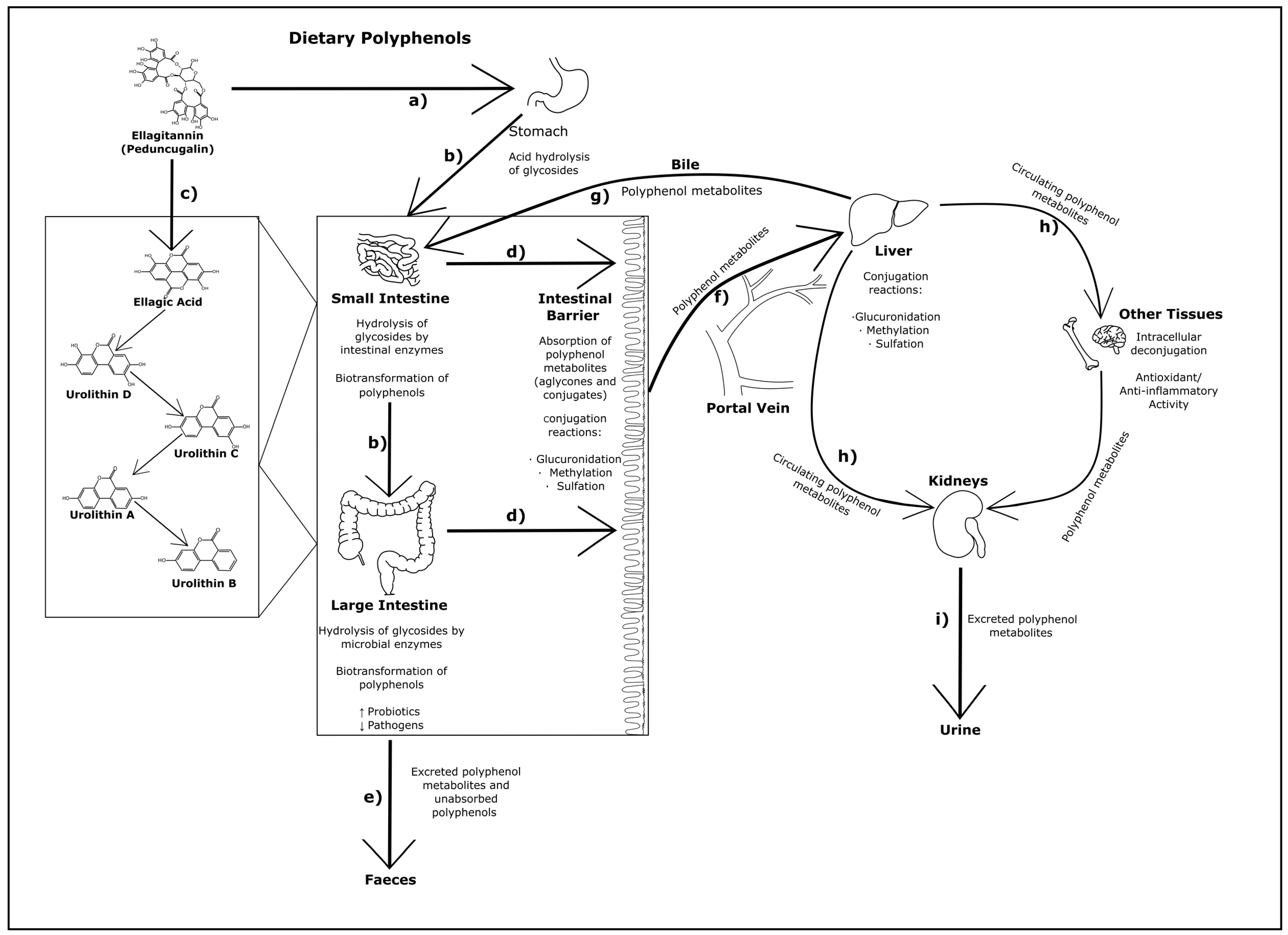{kind=link}
| Class | Example Compounds | Example Structure | Example Food Sources |
|---|---|---|---|
| Flavonoids | |||
| Anthocyanidins | Cyanidin, Delphinidin, Malvidin, Pelargonidin, Peonidin |  (Cyanidin) | Aubergine, Blackcurrant, Blueberry, Black grape, Cherry, Elderberry, Grape, Orange, Plum, Red wine, Rhubarb, Strawberry |
| Flavanols | Catechins (Epicatechin, Epigallocatechin, Epigallocatechin gallate, Gallocatechin), Procyanidins |  (Epigallocatechin) | Apricot, Black tea, Blackberry, Cherry, Chocolate, Grape, Green tea Legumes, Peach, Teas |
| Flavones | Apigenin, Luteolin |  (Apigenin) | Capsicum pepper, Celery, Oregano, Parsley, Rosemary |
| Flavonols | Kaempferol, Myricetin, Quercetin, |  (Quercetin) | Apple, Apricot, Black tea, Broccoli, Green bean, Green tea, Kale, Leek, Onion, Red wine, Tomato |
| Flavonones | Hesperetin, Hesperidin, Naringenin |  (Hesperetin) | Citrus fruits, Grapefruit, Orange, Peppermint, Tangerine, Tomato |
| Isoflavones | Genistein, Daidzein, Glycitein |  (Daidzein) | Lentils, Soy |
| Non-flavonoids | |||
| Phenolic Acids | Hydroxycinnamic acids (Chlorogenic acid, Caffeic acid, Ferulic acid, Sinapic acid) Hydroxybenzoic acids (Gallic acid, Protocatechuic acid) |  (Chlorogenic Acid) | Artichoke Aubergine, Apple, Blackcurrant, Black tea, Blackberry, Blueberry, Cereals, Cherry, Coffee (Chlorogenic Acid), Kiwi, Pear, Plum, Potato, Raspberry, Red wine, Strawberry |
| Lignans | Secoisolariciresinol, Matairesinol |  (Secoisolariciresinol) | Linseed, Sesame seed |
| Hydrolysable Tannins | Ellagitannins (Pedunculagin, Punicalagin) Gallotannins |  (Pedunculagin) | Walnut, Acorn, Oak, Pomegranate |
| Stilbenes | Resveratrol |  (Resveratrol) | Blueberry, Red grapes, Red wine, Raspberry |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scott, M.B.; Styring, A.K.; McCullagh, J.S.O. Polyphenols: Bioavailability, Microbiome Interactions and Cellular Effects on Health in Humans and Animals. Pathogens 2022, 11, 770. https://doi.org/10.3390/pathogens11070770
Scott MB, Styring AK, McCullagh JSO. Polyphenols: Bioavailability, Microbiome Interactions and Cellular Effects on Health in Humans and Animals. Pathogens. 2022; 11(7):770. https://doi.org/10.3390/pathogens11070770
Chicago/Turabian StyleScott, Michael B., Amy K. Styring, and James S. O. McCullagh. 2022. "Polyphenols: Bioavailability, Microbiome Interactions and Cellular Effects on Health in Humans and Animals" Pathogens 11, no. 7: 770. https://doi.org/10.3390/pathogens11070770