
Significance of Mucosa-Associated Microbiota and Its Impacts on Intestinal Health of Pigs Challenged with F18+ E. coli


Abstract
:1. Introduction
2. Results
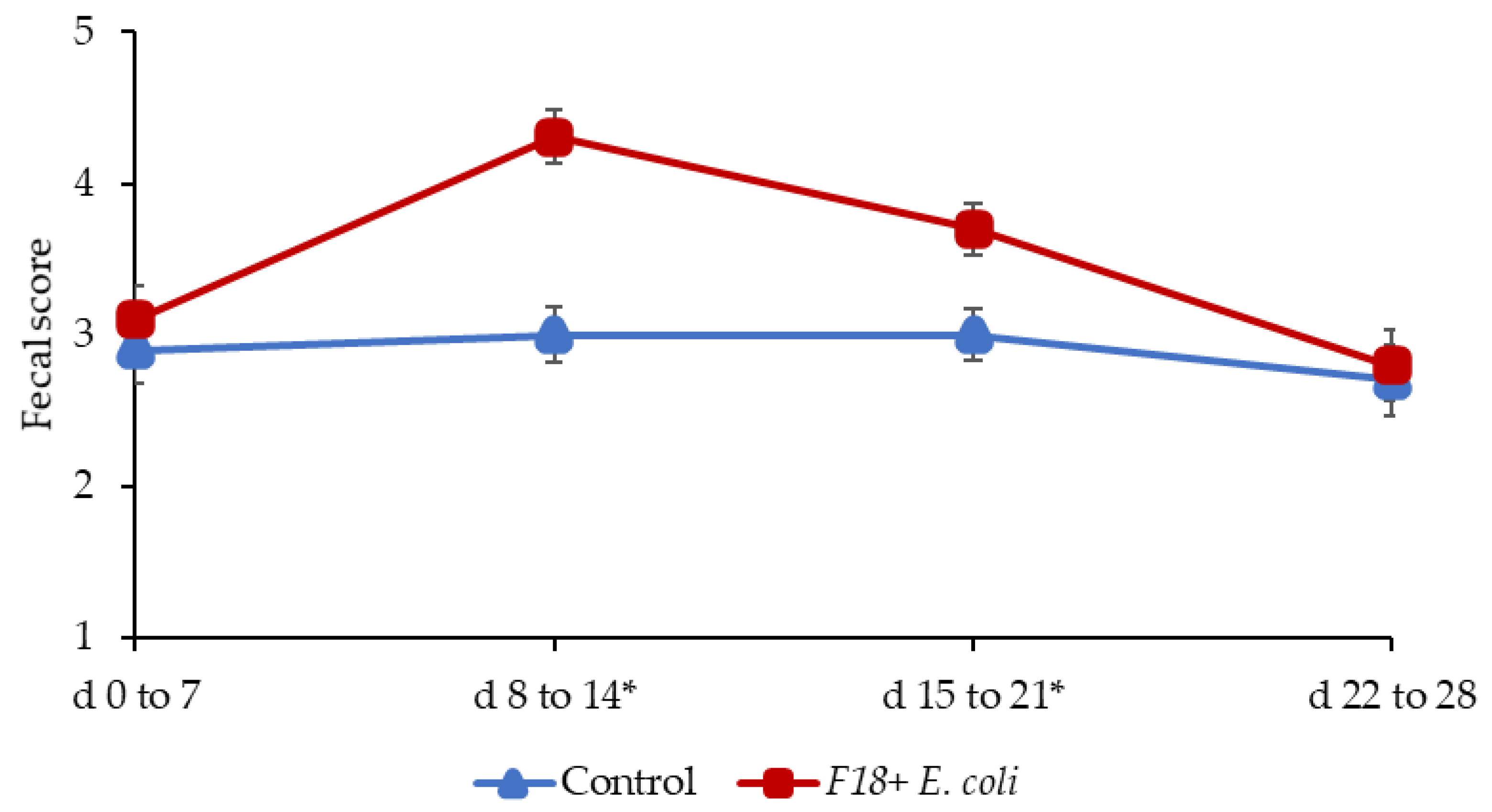
2.1. Growth Performance and Fecal Score
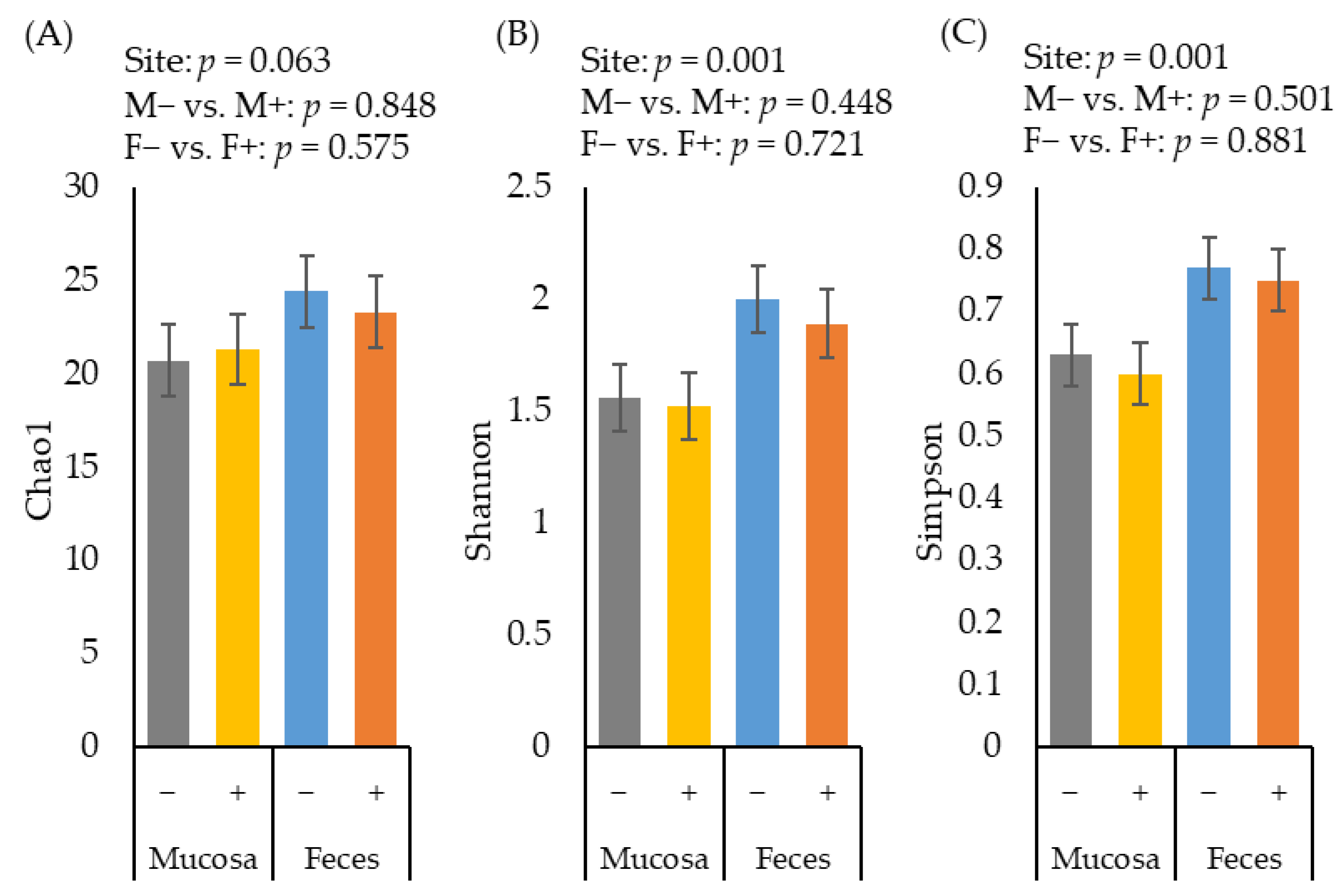
2.2. Alpha Diversity of Microbiota
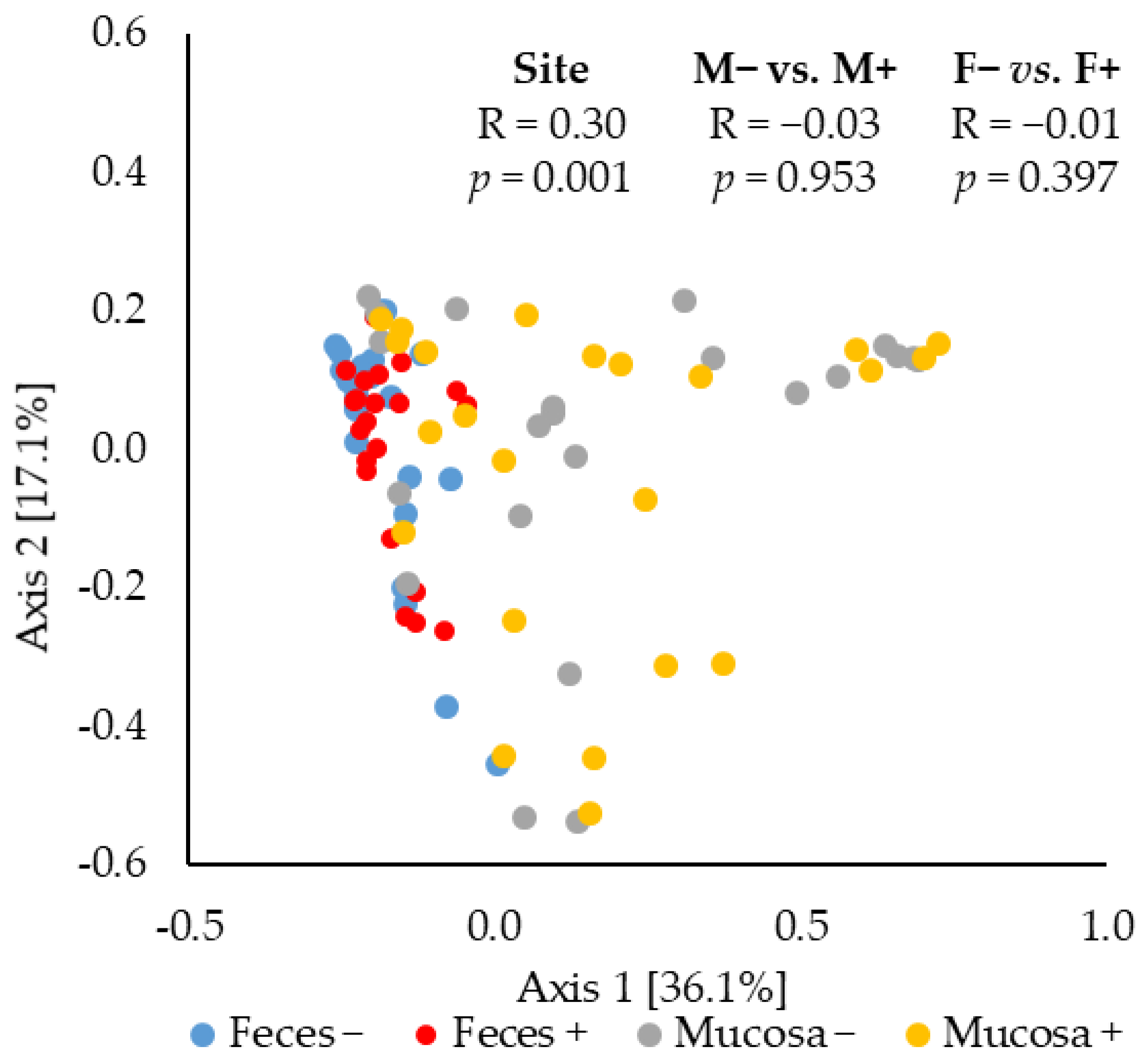
2.3. Beta Diversity of Microbiota
2.4. Relative Abundance of Microbiota
3. Discussion
4. Materials and Methods
4.1. Animals, Diets, Experimental Design, and Inoculum
4.2. Sample Collection and Processing
4.3. Inflammatory and Oxidative Stress Parameters
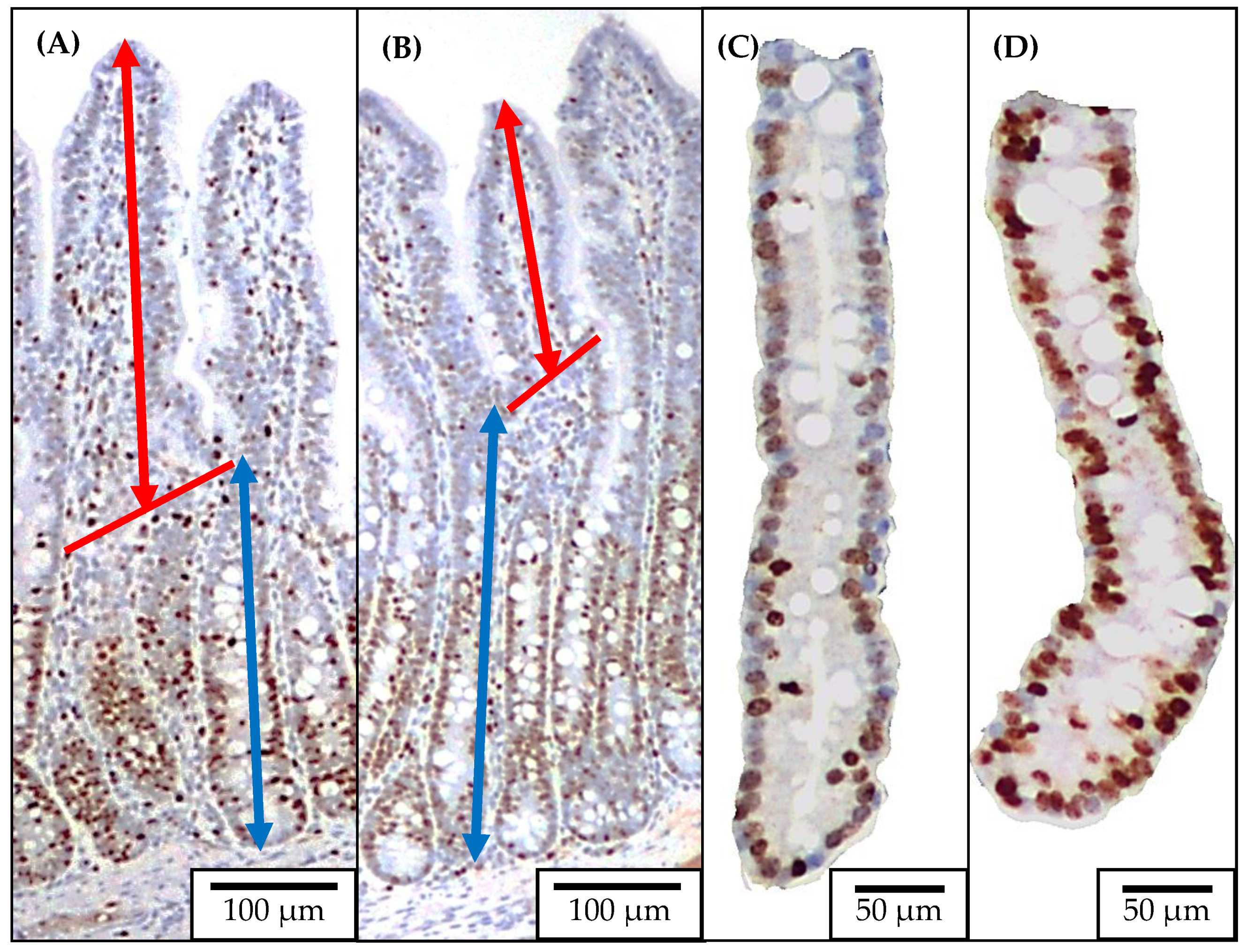
4.4. Jejunal Morphology and Crypt Cell Proliferation
4.5. Relative Abundance and Diversity of the Fecal and Mucosa-Associated Microbiota in Jejunum
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, Y.-C.; Duarte, M.E.; Kim, S.W. Nutritional and functional values of lysed Corynebacterium glutamicum cell mass for intestinal health and growth of nursery pigs. J. Anim. Sci. 2021, 99, skab331. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.E.; Kim, S.W. Intestinal microbiota and its interaction to intestinal health in nursery pigs. Anim. Nutr. 2022, 8, 169–184. [Google Scholar] [CrossRef]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H.; et al. Correlation between Intraluminal Oxygen Gradient and Radial Partitioning of Intestinal Microbiota. Gastroenterology 2014, 147, 1055–1063.e8. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, B.; Kim, S.; Kwon, Y. Characterization of microbiota associated with digesta and mucosa in different regions of gastrointestinal tract of nursery pigs. Int. J. Mol. Sci. 2019, 20, 1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rodas, B.; Youmans, B.P.; Danzeisen, J.L.; Tran, H.; Johnson, T.J. Microbiome profiling of commercial pigs from farrow to finish. J. Anim. Sci. 2018, 96, 1778–1794. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; DeRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral r egulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Friedman, E.S.; Bittinger, K.; Esipova, T.V.; Hou, L.; Chau, L.; Jiang, J.; Mesaros, C.; Lund, P.J.; Liang, X.; FitzGerald, G.A.; et al. Microbes vs. chemistry in the origin of the anaerobic gut lumen. Proc. Natl. Acad. Sci. USA 2018, 115, 4170–4175. [Google Scholar] [CrossRef] [Green Version]
- Etienne-Mesmin, L.; Chassaing, B.; Desvaux, M.; De Paepe, K.; Gresse, R.; Sauvaitre, T.; Forano, E.; van de Wiele, T.; Schüller, S.; Juge, N.; et al. Experimental models to study intestinal microbes–mucus interactions in health and disease. FEMS Microbiol. Rev. 2019, 43, 457–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Wang, Y.; Liu, S.; Huang, J.; Zhai, Z.; He, C.; Ding, J.; Wang, J.; Wang, H.; Fan, W.; et al. The dynamic distribution of porcine microbiota across different ages and gastrointestinal tract segments. PLoS ONE 2015, 10, e0117441. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Guo, P.; Zhang, J.; He, T.; Kim, S.W.; Zhang, G.; Ma, X. Nutrients mediate intestinal bacteria-mucosal immune crosstalk. Front. Immunol. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Shi, N.; Li, N.; Duan, X.; Niu, H. Interaction between the gut microbiome and mucosal immune system. Mil. Med. Res. 2017, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Luise, D.; Lauridsen, C.; Bosi, P.; Trevisi, P. Methodology and application of Escherichia coli F4 and F18 encoding infection models in post-weaning pigs. J. Anim. Sci. Biotechnol. 2019, 10, 53. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Kim, S.W. Intestinal challenge with enterotoxigenic Escherichia coli in pigs, and nutritional intervention to prevent postweaning diarrhea. Anim. Nutr. 2017, 3, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, U.V.; Coddens, A.; Melkebeek, V.; Devriendt, B.; Goetstouwers, T.; van Poucke, M.; Peelman, L.; Cox, E. High susceptibility prevalence for F4+ and F18+ Escherichia coli in Flemish pigs. Vet. Microbiol. 2017, 202, 52–57. [Google Scholar] [CrossRef]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut microbiota dysbiosis in postweaning piglets: Understanding thekeys to health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef]
- Duarte, M.E.; Tyus, J.; Kim, S.W. Synbiotic effects of enzyme and probiotics on intestinal health and growth of newly weaned pigs challenged with enterotoxigenic F18+ Escherichia coli. Front. Vet. Sci. 2020, 7, 573. [Google Scholar] [CrossRef]
- Boeckman, J.X.; Sprayberry, S.; Korn, A.M.; Suchodolski, J.S.; Paulk, C.; Genovese, K.; Rech, R.R.; Giaretta, P.R.; Blick, A.K.; Callaway, T.; et al. Effect of chronic and acute enterotoxigenic E. coli challenge on growth performance, intestinal inflammation, microbiome, and metabolome of weaned piglets. Sci. Rep. 2022, 12, 5024. [Google Scholar] [CrossRef]
- Winter, S.E.; Winter, M.G.; Xavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D.; et al. Host-Derived Nitrate Boosts Growth of E. coli in the Inflamed Gut. Science 2013, 339, 708–711. [Google Scholar] [CrossRef] [Green Version]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017, 10, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Duarte, M.E.; Kim, S.W. Dietary inclusion of multispecies probiotics to reduce the severity of post-weaning diarrhea caused by Escherichia coli F18+ in pigs. Anim. Nutr. 2021, 7, 326–333. [Google Scholar] [CrossRef]
- McLamb, B.L.; Gibson, A.J.; Overman, E.L.; Stahl, C.; Moeser, A.J. Early Weaning Stress in Pigs Impairs Innate Mucosal Immune Responses to Enterotoxigenic E. coli Challenge and Exacerbates Intestinal Injury and Clinical Disease. PLoS ONE 2013, 8, e59838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutler, S.A.; Lonergan, S.M.; Cornick, N.; Johnson, A.K.; Stahl, C.H. Dietary inclusion of colicin E1 is effective in preventing postweaning diarrhea caused by F18-positive Escherichia coli in pigs. Antimicrob. Agents Chemother. 2007, 51, 3830–3835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; He, Y.; Xiong, X.; Ehrlich, A.; Li, X.; Raybould, H.; Atwill, E.R.; Maga, E.A.; Jørgensen, J.; Liu, Y. Dietary supplementation of Bacillus subtilis influenced intestinal health of weaned pigs experimentally infected with a pathogenic E. coli. J. Anim. Sci. Biotechnol. 2019, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Levesque, C.L.; Yu, H.; Gong, J.; de Lange, C.F.M. Ileal mucosa-associated—But not ileal digesta—Bacterial profiles in grower pigs are influenced by nutrition and use of antibiotics for weaner pigs1. J. Anim. Sci. 2012, 90, 448–450. [Google Scholar] [CrossRef]
- Liu, H.; Zeng, X.; Zhang, G.; Hou, C.; Li, N.; Yu, H.; Shang, L.; Zhang, X.; Trevisi, P.; Yang, F.; et al. Maternal milk and fecal microbes guide the spatiotemporal development of mucosa-associated microbiota and barrier function in the porcine neonatal gut. BMC Biol. 2019, 17, 106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wu, W.; Lee, Y.K.; Xie, J.; Zhang, H. Spatial heterogeneity and co-occurrence of mucosal and luminal microbiome across swine intestinal tract. Front. Microbiol. 2018, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Leser, T.D.; Mølbak, L. Better living through microbial action: The benefits of the mammalian gastrointestinal microbiota on the host. Environ. Microbiol. 2009, 11, 2194–2206. [Google Scholar] [CrossRef]
- Beeby, M. Motility in the epsilon-proteobacteria. Curr. Opin. Microbiol. 2015, 28, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Mace, O.J.; Marshall, F. Gut chemosensing and the regulation of nutrient absorption and energy supply. J. Anim. Sci. 2013, 91, 1932–1945. [Google Scholar] [CrossRef] [Green Version]
- Espey, M.G. Role of oxygen gradients in shaping redox relationships between the human intestine and its microbiota. Free Radic. Biol. Med. 2013, 55, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Wiarda, J.E.; Trachsel, J.M.; Bond, Z.F.; Byrne, K.A.; Gabler, N.K.; Loving, C.L. Intraepithelial T cells diverge by intestinal location as pigs age. Front. Immunol. 2020, 11, 1139. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Jia, J.; Zhang, L.; Chen, Q.; Zhang, X.; Sun, W.; Ma, C.; Xu, F.; Zhan, S.; Ma, L.; et al. Jejunal inflammatory cytokines, barrier proteins and microbiome-metabolome responses to early supplementary feeding of Bamei suckling piglets. BMC Microbiol. 2020, 20, 169. [Google Scholar] [CrossRef]
- Mu, C.; Yang, Y.; Su, Y.; Zoetendal, E.G.; Zhu, W. Differences in microbiota membership along the gastrointestinal tract of piglets and their differential alterations following an early-life antibiotic intervention. Front. Microbiol. 2017, 8, 797. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.; Daly, K.; Moran, A.W.; Ryan, S.; Bravo, D.; Shirazi-Beechey, S.P. Composition and diversity of mucosa-associated microbiota along the entire length of the pig gastrointestinal tract; dietary influences. Environ. Microbiol. 2017, 19, 1425–1438. [Google Scholar] [CrossRef] [Green Version]
- Plancade, S.; Clark, A.; Philippe, C.; Helbling, J.-C.; Moisan, M.-P.; Esquerré, D.; Le Moyec, L.; Robert, C.; Barrey, E.; Mach, N. Unraveling the effects of the gut microbiota composition and function on horse endurance physiology. Sci. Rep. 2019, 9, 9620. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Peng, X.; Burrough, E.R.; Sahin, O.; Gould, S.A.; Gabler, N.K.; Loving, C.L.; Dorman, K.S.; Patience, J.F. Dietary Soluble and Insoluble Fiber with or without Enzymes Altered the Intestinal Microbiota in Weaned Pigs Challenged with Enterotoxigenic E. coli F18. Front. Microbiol. 2020, 11, 1110. [Google Scholar] [CrossRef]
- Guo, P.; Zhang, K.; Ma, X.; He, P. Clostridium species as probiotics: Potentials and challenges. J. Anim. Sci. Biotechnol. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Heeney, D.D.; Gareau, M.G.; Marco, M.L. Intestinal Lactobacillus in health and disease, a driver or just along for the ride? Curr. Opin. Biotechnol. 2018, 49, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Moita, V.H.C.; Duarte, M.E.; da Silva, S.N.; Kim, S.W. Supplemental Effects of Functional Oils on the Modulation of Mucosa-Associated Microbiota, Intestinal Health, and Growth Performance of Nursery Pigs. Animals 2021, 11, 1591. [Google Scholar] [CrossRef] [PubMed]
- Van Den Broeck, W.; Cox, E.; Oudega, B.; Goddeeris, B.M. The F4 fimbrial antigen of Escherichia coli and its receptors. Vet. Microbiol. 2000, 71, 223–244. [Google Scholar] [CrossRef]
- Devriendt, B.; Stuyven, E.; Verdonck, F.; Goddeeris, B.M.; Cox, E. Enterotoxigenic Escherichia coli (K88) induce proinflammatory responses in porcine intestinal epithelial cells. Dev. Comp. Immunol. 2010, 34, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhong, Z.; Luo, Y.; Cox, E.; Devriendt, B. Heat-stable enterotoxins of enterotoxigenic escherichia coli and their impact on host immunity. Toxins 2019, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Loos, M.; Geens, M.; Schauvliege, S.; Gasthuys, F.; van der Meulen, J.; Dubreuil, J.D.; Goddeeris, B.M.; Niewold, T.; Cox, E. Role of heat-stable enterotoxins in the induction of early immune responses in piglets after infection with enterotoxigenic Escherichia coli. PLoS ONE 2012, 7, e41041. [Google Scholar] [CrossRef] [Green Version]
- Levesque, C.L.; Hooda, S.; Swanson, K.S.; De Lange, K. Alterations in ileal mucosa bacteria related to diet complexity and growth performance in young pigs. PLoS ONE 2014, 9, e108472. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhu, Y.-H.; Zhou, D.; Wu, Q.; Song, D.; Dicksved, J.; Wang, J.-F. Oral Administration of a Select Mixture of Bacillus Probiotics Affects the Gut Microbiota and Goblet Cell Function following Escherichia coli Challenge in Newly Weaned Pigs of Genotype MUC4 That Are Supposed to Be Enterotoxigenic E. coli F4ab/ac Receptor. Appl. Environ. Microbiol. 2017, 83, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Sido, A.; Radhakrishnan, S.; Kim, S.W.; Eriksson, E.; Shen, F.; Li, Q.; Bhat, V.; Reddivari, L.; Vanamala, J.K.P. A food-based approach that targets interleukin-6, a key regulator of chronic intestinal inflammation and colon carcinogenesis. J. Nutr. Biochem. 2017, 43, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Duarte, M.E.; Zhou, F.X.; Dutra, W.M.; Kim, S.W. Dietary supplementation of xylanase and protease on growth performance, digesta viscosity, nutrient digestibility, immune and oxidative stress status, and gut health of newly weaned pigs. Anim. Nutr. 2019, 5, 351–358. [Google Scholar] [CrossRef]
- Duarte, M.E.; Sparks, C.; Kim, S.W. Modulation of jejunal mucosa-associated microbiota in relation to intestinal health and nutrient digestibility in pigs by supplementation of β-glucanase to corn–soybean meal-based diets with xylanase. J. Anim. Sci. 2021, 99, skab190. [Google Scholar] [CrossRef]
- Kim, S.W.; Duarte, M.E. Understanding intestinal health in nursery pigs and the relevant nutritional strategies. Anim. Biosci. 2021, 34, 338–344. [Google Scholar] [CrossRef]
- Zheng, L.; Duarte, M.E.; Sevarolli Loftus, A.; Kim, S.W. Intestinal health of pigs upon weaning: Challenges and nutritional intervention. Front. Vet. Sci. 2021, 8, 628258. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012; ISBN 978-0-309-22423-9. [Google Scholar]
- Nagy, B.; Whipp, S.C.; Imberechts, H.; Bertschinger, H.U.; Dean-Nystrom, E.A.; Casey, T.A.; Salajka, E. Biological relationship between F18ab and F18ac fimbriae of enterotoxigenic and verotoxigenic Escherichia coli from weaned pigs with oedema disease or diarrhoea. Microb. Pathog. 1997, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.B.; Kim, S.W. Supplemental effects of dietary nucleotides on intestinal health and growth performance of newly weaned pigs. J. Anim. Sci. 2019, 97, 4875–4882. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site 1 | Mucosa | Feces | p Value 3 | |||||
|---|---|---|---|---|---|---|---|---|
| Challenge 2 | − | + | − | + | SEM | Site | M− vs. M+ | F− vs. F+ |
| Firmicutes | 36.11 | 35.07 | 48.61 | 46.73 | 5.40 | 0.014 | 0.878 | 0.781 |
| Proteobacteria | 35.93 | 43.47 | 12.27 | 15.10 | 5.60 | <0.001 | 0.287 | 0.689 |
| Bacteroidetes | 21.55 | 16.04 | 35.23 | 32.15 | 3.24 | <0.001 | 0.230 | 0.500 |
| Chlamydiae | 3.72 | 2.91 | <0.01 | 1.71 | 2.05 | 0.203 | 0.769 | 0.523 |
| Spirochaetes | 1.53 | 0.11 | 1.09 | 0.34 | 0.64 | 0.867 | 0.122 | 0.410 |
| Actinobacteria | 0.41 | 0.57 | 1.17 | 0.55 | 0.36 | 0.316 | 0.754 | 0.231 |
| Cyanobacteria | 0.33 | 1.51 | 0.03 | 0.03 | 0.74 | 0.223 | 0.254 | 0.996 |
| Others | 0.31 | 0.21 | 1.21 | 3.10 | 1.57 | 0.357 | 0.987 | 0.167 |
| Site 1 | Mucosa | Feces | p Value 3 | |||||
|---|---|---|---|---|---|---|---|---|
| Challenge 2 | − | + | − | + | SEM | Site | M− vs. M+ | F− vs. F+ |
| Helicobacteraceae | 14.31 | 29.89 | 0.62 | 0.77 | 5.21 | <0.001 | 0.017 | 0.982 |
| Prevotellaceae | 19.83 | 14.77 | 29.39 | 28.81 | 2.92 | <0.001 | 0.219 | 0.888 |
| Lactobacillaceae | 13.37 | 14.34 | 9.37 | 11.62 | 3.44 | 0.331 | 0.843 | 0.645 |
| Veillonellaceae | 8.27 | 7.16 | 18.95 | 17.14 | 2.96 | <0.001 | 0.713 | 0.550 |
| Campylobacteraceae | 8.30 | 2.04 | 1.29 | 0.41 | 1.98 | 0.039 | 0.027 | 0.757 |
| Pseudomonadaceae | 1.80 | 4.74 | <0.01 | <0.01 | 1.60 | <0.001 | 0.034 | 0.951 |
| Clostridiaceae | 4.47 | 2.27 | 1.60 | 1.67 | 1.87 | 0.235 | 0.288 | 0.975 |
| Enterobacteriaceae | 5.24 | 1.44 | 0.12 | 2.14 | 1.65 | 0.038 | 0.021 | 0.140 |
| Streptococcaceae | 2.96 | 4.31 | 1.31 | 1.36 | 2.16 | 0.138 | 0.535 | 0.982 |
| Chlamydiaceae | 3.56 | 2.27 | 0.02 | <0.01 | 1.90 | 0.065 | 0.587 | 0.979 |
| Lachnospiraceae | 1.31 | 1.94 | 4.81 | 4.73 | 0.84 | <0.001 | 0.541 | 0.942 |
| Succinivibrionaceae | 0.79 | 0.46 | 7.48 | 10.00 | 3.68 | <0.001 | 0.917 | 0.428 |
| Ruminococcaceae | 1.25 | 1.34 | 5.64 | 4.86 | 0.64 | <0.001 | 0.908 | 0.319 |
| Brachyspiraceae | 2.44 | 0.08 | 0.55 | 0.03 | 0.78 | 0.214 | 0.035 | 0.634 |
| Moraxellaceae | 1.13 | 0.84 | <0.01 | 0.03 | 0.52 | 0.029 | 0.667 | 0.980 |
| Oxalobacteraceae | 0.47 | 1.18 | 0.57 | 0.34 | 0.72 | 0.501 | 0.367 | 0.780 |
| Acidaminococcaceae | 1.35 | 1.10 | 2.65 | 3.55 | 0.73 | <0.001 | 0.643 | 0.088 |
| Nostocaceae | 0.29 | 1.25 | <0.01 | <0.01 | 0.60 | 0.197 | 0.254 | 0.999 |
| Xanthomonadaceae | 0.35 | 1.03 | <0.01 | <0.01 | 0.25 | <0.001 | 0.013 | 0.969 |
| Porphyromonadaceae | 0.96 | 0.74 | 2.54 | 2.62 | 0.54 | <0.001 | 0.717 | 0.893 |
| Pseudanabaenaceae | 0.60 | 0.87 | 0.07 | 0.08 | 0.45 | 0.011 | 0.458 | 0.970 |
| Erysipelotrichaceae | 0.45 | 0.76 | 1.23 | 1.42 | 0.33 | 0.033 | 0.514 | 0.694 |
| Bacillaceae | 1.19 | 0.03 | 0.08 | 0.03 | 0.48 | 0.244 | 0.089 | 0.944 |
| Bifidobacteriaceae | 0.34 | 0.21 | 0.99 | 0.29 | 0.36 | 0.289 | 0.786 | 0.146 |
| Eubacteriaceae | 0.64 | 0.54 | 2.42 | 2.58 | 0.37 | <0.001 | 0.824 | 0.707 |
| Hyphomicrobiaceae | 0.29 | 0.29 | 0.26 | 0.43 | 0.12 | 0.592 | 0.973 | 0.299 |
| Bacteroidaceae | 0.25 | 0.11 | 0.27 | 0.24 | 0.08 | 0.326 | 0.229 | 0.816 |
| Flavobacteriaceae | 0.27 | 0.09 | 0.27 | 0.36 | 0.10 | 0.173 | 0.214 | 0.547 |
| Cytophagaceae | 0.27 | 0.19 | 1.63 | 1.14 | 0.67 | 0.012 | 0.900 | 0.446 |
| Spirochaetaceae | 0.07 | 0.02 | 0.70 | 0.25 | 0.17 | 0.005 | 0.789 | 0.041 |
| Oscillospiraceae | 0.09 | 0.06 | 0.52 | 0.34 | 0.09 | <0.001 | 0.738 | 0.037 |
| Peptostreptococcaceae | 0.06 | 0.49 | 0.06 | 0.12 | 0.15 | 0.098 | 0.008 | 0.698 |
| Mycoplasmataceae | 0.13 | 0.07 | 0.35 | 0.15 | 0.13 | 0.132 | 0.623 | 0.139 |
| Spiroplasmataceae | 0.01 | 0.01 | 0.24 | 0.31 | 0.06 | <0.001 | 0.928 | 0.368 |
| Caulobacteraceae | 0.43 | 0.08 | <0.01 | <0.01 | 0.14 | 0.006 | 0.016 | 0.968 |
| Rikenellaceae | 0.05 | 0.05 | 0.28 | 0.28 | 0.13 | 0.011 | 0.995 | 0.980 |
| Others | 1.79 | 1.88 | 4.13 | 1.94 | 0.72 | 0.100 | 0.921 | 0.016 |
| Site 1 | Mucosa | Feces | p Value 3 | |||||
|---|---|---|---|---|---|---|---|---|
| Challenge 2 | − | + | − | + | SEM | Site | M− vs. M+ | F− vs. F+ |
| Prevotella | 22.15 | 15.01 | 33.78 | 30.10 | 3.60 | <0.001 | 0.165 | 0.472 |
| Lactobacillus | 16.04 | 18.44 | 12.61 | 18.44 | 4.31 | 0.688 | 0.691 | 0.335 |
| Helicobacter | 16.83 | 32.83 | 0.51 | 0.65 | 5.20 | <0.001 | 0.015 | 0.983 |
| Succinivibrio | 0.74 | 0.40 | 9.81 | 12.90 | 4.83 | <0.001 | 0.930 | 0.420 |
| Campylobacter | 10.07 | 2.55 | 2.25 | 0.70 | 2.55 | 0.071 | 0.040 | 0.669 |
| Clostridium | 4.74 | 2.62 | 1.25 | 1.17 | 2.51 | 0.152 | 0.381 | 0.972 |
| Megasphaera | 1.90 | 1.27 | 6.16 | 4.44 | 1.68 | 0.002 | 0.703 | 0.297 |
| Mitsuokella | 2.75 | 1.38 | 2.99 | 4.91 | 1.40 | 0.102 | 0.397 | 0.240 |
| Pseudomonas | 3.53 | 6.57 | <0.01 | <0.01 | 1.74 | <0.001 | 0.073 | 0.963 |
| Faecalibacterium | 1.58 | 1.34 | 4.70 | 4.08 | 0.97 | <0.001 | 0.809 | 0.532 |
| Streptococcus | 3.49 | 4.76 | 1.45 | 1.92 | 2.34 | 0.135 | 0.579 | 0.839 |
| Phascolarctobacterium | 1.78 | 1.12 | 3.49 | 2.91 | 0.82 | 0.003 | 0.412 | 0.466 |
| Selenomonas | 0.35 | 0.33 | 4.39 | 2.51 | 1.15 | <0.001 | 0.989 | 0.249 |
| Chlamydia | 4.12 | 2.44 | <0.01 | <0.01 | 2.07 | 0.056 | 0.515 | 0.967 |
| Dialister | 1.17 | 0.71 | 1.70 | 3.96 | 1.01 | <0.001 | 0.518 | 0.002 |
| Roseburia | 0.54 | 0.49 | 2.03 | 1.94 | 0.40 | <0.001 | 0.910 | 0.867 |
| Bifidobacterium | 0.24 | 0.42 | 2.23 | 0.46 | 0.85 | 0.220 | 0.883 | 0.133 |
| Acidaminococcus | 0.30 | 0.30 | 0.83 | 2.36 | 0.53 | 0.001 | 0.997 | 0.005 |
| Eubacterium | 0.33 | 0.38 | 1.34 | 0.94 | 0.44 | 0.041 | 0.922 | 0.454 |
| Oscillibacter | <0.01 | <0.01 | 1.68 | 0.54 | 0.48 | <0.001 | 0.880 | 0.004 |
| Acinetobacter | 1.43 | 1.05 | <0.01 | 0.02 | 0.59 | 0.024 | 0.619 | 0.922 |
| Gemmiger | 0.40 | 0.44 | 0.47 | 0.78 | 0.20 | 0.304 | 0.890 | 0.278 |
| Herbaspirillum | <0.01 | <0.01 | 1.08 | 0.69 | 0.39 | <0.001 | 0.998 | 0.270 |
| Massilia | 0.52 | 1.05 | <0.01 | <0.01 | 0.50 | 0.051 | 0.850 | 0.984 |
| Bacillus | 1.44 | 0.02 | <0.01 | <0.01 | 0.57 | 0.219 | 0.084 | 0.999 |
| Coprococcus | 0.01 | 0.06 | 1.00 | 0.15 | 0.41 | 0.177 | 0.928 | 0.134 |
| Catenibacterium | 0.13 | 0.16 | 0.16 | 0.69 | 0.25 | 0.254 | 0.920 | 0.126 |
| Anaerovibrio | 0.09 | 0.22 | 0.44 | 0.27 | 0.15 | 0.115 | 0.434 | 0.339 |
| Dorea | 0.25 | 0.26 | 0.23 | 0.37 | 0.12 | 0.651 | 0.968 | 0.356 |
| Tepidimonas | <0.01 | <0.01 | 0.80 | 0.10 | 0.27 | 0.027 | 0.973 | 0.027 |
| Ruminococcus | 0.06 | 0.21 | 0.28 | 0.35 | 0.09 | 0.049 | 0.262 | 0.563 |
| Blautia | 0.16 | 0.50 | <0.01 | <0.01 | 0.15 | 0.004 | 0.039 | 0.879 |
| Brachyspira | 0.19 | 0.10 | 0.51 | 0.05 | 0.32 | 0.461 | 0.744 | 0.207 |
| Enterococcus | 0.60 | 0.01 | <0.01 | <0.01 | 0.29 | 0.293 | 0.147 | 0.999 |
| Treponema | 0.13 | 0.05 | 0.33 | 0.8 | 0.15 | 0.411 | 0.715 | 0.207 |
| Others | 1.74 | 1.77 | 1.12 | 1.54 | 0.39 | 0.225 | 0.812 | 0.445 |
| Site 1 | Mucosa | Feces | p Value 3 | |||||
|---|---|---|---|---|---|---|---|---|
| Challenge 2 | − | + | − | + | SEM | Site | M− vs. M+ | F− vs. F+ |
| Prevotella copri | 23.63 | 14.64 | 33.52 | 28.23 | 4.88 | 0.001 | 0.073 | 0.288 |
| Succinivibrio dextrinosolvens | 2.04 | 0.91 | 12.54 | 10.26 | 5.64 | 0.002 | 0.799 | 0.607 |
| Lactobacillus kitasatonis | 4.13 | 5.62 | 3.39 | 8.75 | 3.20 | 0.664 | 0.701 | 0.169 |
| Helicobacter mastomyrinus | 7.75 | 14.40 | <0.01 | <0.01 | 2.67 | <0.001 | 0.079 | 0.999 |
| Helicobacter rappini | 5.23 | 15.59 | 0.90 | 1.07 | 3.96 | 0.001 | 0.011 | 0.965 |
| Faecalibacterium prausnitzii | 3.54 | 2.67 | 5.24 | 5.54 | 1.10 | 0.027 | 0.546 | 0.834 |
| Prevotella stercorea | 2.91 | 2.24 | 3.90 | 3.91 | 1.15 | 0.068 | 0.511 | 0.990 |
| Lactobacillus mucosae | 4.22 | 4.42 | 0.70 | 2.20 | 1.58 | 0.052 | 0.923 | 0.469 |
| Campylobacter coli | 9.03 | 0.43 | 1.11 | 0.27 | 2.65 | 0.143 | 0.021 | 0.824 |
| Phascolarctobacterium succinatutens | 2.65 | 1.45 | 3.92 | 3.43 | 0.97 | 0.024 | 0.232 | 0.622 |
| Lactobacillus delbrueckii | 1.55 | 1.86 | 3.14 | 2.60 | 1.23 | 0.265 | 0.837 | 0.715 |
| Mitsuokella jalaludinii | 2.40 | 1.05 | 2.14 | 3.70 | 1.05 | 0.253 | 0.288 | 0.358 |
| Dialister succinatiphilus | 1.93 | 1.19 | 2.35 | 5.12 | 1.29 | 0.005 | 0.484 | 0.010 |
| Roseburia faecis | 0.74 | 1.08 | 3.48 | 2.72 | 0.56 | <0.001 | 0.674 | 0.346 |
| Streptococcus alactolyticus | 3.32 | 3.36 | 1.22 | 1.01 | 2.07 | 0.080 | 0.981 | 0.908 |
| Lactobacillus johnsonii | 1.03 | 1.37 | 1.64 | 1.54 | 0.90 | 0.516 | 0.693 | 0.908 |
| Chlamydia suis | 3.04 | 2.49 | <0.01 | <0.01 | 1.75 | 0.058 | 0.797 | 0.935 |
| Lactobacillus salivarius | 0.65 | 1.01 | 1.48 | 2.38 | 0.89 | 0.072 | 0.669 | 0.294 |
| Selenomonas lipolytica | 0.51 | <0.01 | 2.96 | 0.85 | 0.87 | 0.015 | 0.621 | 0.036 |
| Gemmiger formicilis | 0.92 | 1.09 | 0.63 | 1.66 | 0.51 | 0.978 | 0.763 | 0.195 |
| Mitsuokella multacida | 1.62 | 0.69 | 0.31 | 1.30 | 0.54 | 0.522 | 0.226 | 0.199 |
| Megasphaera hominis | 0.50 | 0.30 | 2.62 | 0.89 | 1.09 | 0.130 | 0.877 | 0.172 |
| Acidaminococcus fermentans | 0.20 | 0.33 | 0.83 | 2.30 | 0.45 | 0.003 | 0.826 | 0.015 |
| Helicobacter equorum | 0.05 | 3.42 | <0.01 | 0.04 | 1.21 | 0.163 | 0.054 | 0.980 |
| Selenomonas bovis | 0.25 | 0.08 | 2.17 | 0.90 | 0.60 | 0.004 | 0.827 | 0.235 |
| Prevotella sp. | 0.60 | 0.31 | 0.85 | 1.79 | 0.62 | 0.005 | 0.504 | 0.028 |
| Streptococcus hyointestinalis | 0.01 | 1.87 | 0.15 | 0.78 | 0.69 | 0.485 | 0.058 | 0.516 |
| Streptococcus infantarius | 1.11 | 1.72 | 0.30 | 0.27 | 0.89 | 0.062 | 0.478 | 0.974 |
| Acinetobacter radioresistens | 1.48 | 0.80 | <0.01 | <0.01 | 0.71 | 0.044 | 0.432 | 0.982 |
| Acinetobacter lwoffii | 1.99 | 0.42 | <0.01 | <0.01 | 0.54 | 0.013 | 0.027 | 0.991 |
| Campylobacter lanienae | 0.29 | 0.24 | 1.51 | 0.21 | 0.31 | 0.058 | 0.910 | 0.004 |
| Treponema porcinum | 0.13 | 0.08 | 1.48 | 0.31 | 0.37 | 0.016 | 0.915 | 0.011 |
| Campylobacter upsaliensis | 0.91 | 1.08 | 0.07 | 0.09 | 0.74 | 0.127 | 0.736 | 0.996 |
| Dorea longicatena | 0.51 | 0.60 | 0.26 | 0.45 | 0.17 | 0.247 | 0.723 | 0.428 |
| Others | 9.44 | 11.56 | 5.66 | 6.41 | 2.61 | 0.055 | 0.516 | 0.816 |
| Item | No challenge (A) | Challenge (B) | SEM | p value |
|---|---|---|---|---|
| Serum | ||||
| TNF-α, pg/mL | 98.6 | 113.7 | 25.1 | 0.172 |
| Protein carbonyl, nmol/mg of protein | 1.74 | 2.10 | 0.45 | 0.396 |
| Jejunal mucosa | ||||
| TNF-α, pg/mg of protein | 1.15 | 1.31 | 0.27 | 0.551 |
| Protein carbonyl, nmol/mg of protein | 2.33 | 3.17 | 0.26 | 0.026 |
| Item | No Challenge (A) | Challenge (B) | SEM | p Value |
|---|---|---|---|---|
| Jejunal mucosa | ||||
| Villus Height, µm | 494 | 434 | 23 | 0.026 |
| Crypt Depth, µm | 209 | 224 | 30 | 0.169 |
| VH:CD 1 | 2.60 | 2.14 | 0.30 | 0.044 |
| Ki67+, % | 51 | 55 | 8 | 0.632 |
| Item 1 | Family (r, p Value) | Species (r, p Value) |
|---|---|---|
| TNF-α, serum | Lactobacillaceae (0.30, 0.048) | Succinivibrio dextrinosolvens (−0.33, 0.030) |
| Veillonellaceae (0.32, 0.034) | Faecalibacterium prausnitzii (−0.36, 0.017) | |
| Enterobacteriaceae (−0.32, 0.037) | Lactobacillus mucosae (0.51, <0.001) | |
| Acidaminococcaceae (0.36, 0.018) | Megasphaera hominis (0.49, 0.001) | |
| Pseudomonadaceae (−0.49, 0.001) | Campylobacter lanienae (0.49, 0.001) | |
| Porphyromonadaceae (0.43, 0.004) | ||
| Eubacteriaceae (0.33, 0.027) | ||
| Cytophagaceae (0.33, 0.027) | ||
| Xanthomonadaceae (−0.32, 0.036) | ||
| Pseudanabaenaceae (0.35, 0.018) | ||
| Oscillospiraceae (0.37, 0.015) | ||
| Spiroplasmataceae (0.33, 0.028) | ||
| Caulobacteraceae (−0.41, 0.005) | ||
| TNF-α, mucosa | Bifidobacteriaceae (−0.63, <0.001) | Mitsuokella jalaludinii (0.45, 0.009) |
| Dialister succinatiphilus (0.52, 0.002) | ||
| Mitsuokella multacida (0.42, 0.016) | ||
| Acidaminococcus fermentans (0.60, <0.001) | ||
| PC, serum | Oscillospiraceae (0.42, 0.005) | Lactobacillus johnsonii (0.49, 0.001) |
| Spiroplasmataceae (0.69, <0.001) | Megasphaera hominis (0.45, 0.024) | |
| Campylobacter lanienae (0.66, <0.001) | ||
| PC, mucosa | Erysipelotrichaceae (0.32, 0.034) | Acidaminococcus fermentans (0.30, 0.044) |
| Bifidobacteriaceae (−0.33, 0.031) | Acinetobacter lwoffii (0.33, 0.031) | |
| Spiroplasmataceae (0.44, 0.003) | ||
| Villus height | Veillonellaceae (0.33, 0.031) | Campylobacter coli (0.33, 0.027) |
| Brachyspiraceae (0.34, 0.023) | Megasphaera hominis (0.37, 0.013) | |
| Pseudomonadaceae (0.33, 0.026) | ||
| Oscillospiraceae (0.42, 0.041) | ||
| Crypt depth | Prevotellaceae (−0.36, 0.019) | Succinivibrio dextrinosolvens (−0.50, 0.001) |
| Lactobacillaceae (0.41, 0.006) | Helicobacter rappini (0.32, 0.039) | |
| Succinivibrionaceae (−0.39, 0.010) | Faecalibacterium prausnitzii (−0.55, <0.001) | |
| Lachnospiraceae (−0.40, 0.008) | Dialister succinatiphilus (0.30, 0.047) | |
| Ruminococcaceae (−0.36, 0.019) | Roseburia faecis (−0.38, 0.013) | |
| Clostridiaceae (−0.49, 0.001) | Gemmiger formicilis (−0.38, 0.013) | |
| Pseudomonadaceae (−0.30, 0.049) | Megasphaera hominis (0.41, 0.007) | |
| Xanthomonadaceae (−0.32, 0.034) | Selenomonas bovis (0.33, 0.035) | |
| Pseudanabaenaceae (0.36, 0.017) | Campylobacter lanienae (0.35, 0.020) | |
| Mycoplasmataceae (−0.34, 0.028) | Dorea longicatena (−0.40, 0.008) | |
| VH:CD | Lactobacillaceae (−0.32, 0.034) | Succinivibrio dextrinosolvens (0.35, 0.021) |
| Lachnospiraceae (0.31, 0.046) | Helicobacter rappini (−0.34, 0.024) | |
| Clostridiaceae (0.48, 0.001) | Faecalibacterium prausnitzii (0.43, 0.004) | |
| Xanthomonadaceae (0.32, 0.036) | Roseburia faecis (0.34, 0.024) | |
| Acinetobacter lwoffii (0.33, 0.032) | ||
| Dorea longicatena (0.35, 0.020) |
| Item 1 | Family (r, p Value) | Species (r, p Value) |
|---|---|---|
| TNF-α, serum | Veillonellaceae (0.37, 0.013) | Prevotella copri (0.49, <0.001) |
| Succinivibrionaceae (−0.33, 0.031) | Succinivibrio dextrinosolvens (−0.49, <0.001) | |
| Streptococcaceae (0.47, 0.001) | Phascolarctobacterium succinatutens (0.36, 0.016) | |
| Acidaminococcaceae (0.31, 0.041) | Streptococcus alactolyticus (0.36, 0.015) | |
| Eubacteriaceae (0.34, 0.025) | Selenomonas lipolytica (−0.30, 0.046) | |
| Oxalobacteraceae (−0.31, 0.044) | Megasphaera hominis (0.46, 0.002) | |
| Cytophagaceae (0.41, 0.006) | Streptococcus hyointestinalis (0.34, 0.024) | |
| Flavobacteriaceae (0.38, 0.010) | Streptococcus infantarius (0.37, 0.013) | |
| Treponema porcinum (−0.32, 0.037) | ||
| Dorea longicatena (0.30, 0.049) | ||
| PC, serum | Acidaminococcaceae (0.31, 0.043) | Prevotella copri (0.35, 0.022) |
| Villus height | Flavobacteriaceae (0.33, 0.030) | Dialister succinatiphilus (0.41, 0.005) |
| Lactobacillus salivarius (0.34, 0.025) | ||
| Mitsuokella multacida (0.34, 0.022) | ||
| Treponema porcinum (−0.30, 0.049) | ||
| Crypt depth | Prevotellaceae (0.45, 0.003) | Prevotella copri (0.47, 0.002) |
| Lactobacillaceae (−0.34, 0.026) | Succinivibrio dextrinosolvens (−0.42, 0.004) | |
| Succinivibrionaceae (−0.42, 0.005) | Lactobacillus kitasatonis (−0.33, 0.030) | |
| Ruminococcaceae (0.35, 0.023) | Faecalibacterium prausnitzii (0.34, 0.023) | |
| Acidaminococcaceae (0.48, 0.001) | Prevotella stercorea (0.31, 0.040) | |
| Oxalobacteraceae (−0.33, 0.028) | Dialister succinatiphilus (0.42, 0.005) | |
| Cytophagaceae (0.34, 0.026) | Lactobacillus johnsonii (−0.37, 0.016) | |
| Bifidobacteriaceae (−0.32, 0.034) | Selenomonas lipolytica (−0.45, 0.003) | |
| Flavobacteriaceae (0.35, 0.020) | Selenomonas bovis (−0.41, 0.006) | |
| Oscillospiraceae (0.32, 0.034) | Prevotella sp. (0.45, 0.003) | |
| VH:CD | Prevotellaceae (−032, 0.034) | Selenomonas lipolytica (0.39, 0.009) |
| Lactobacillaceae (0.32, 0.036) | Selenomonas bovis (0.46, 0.002) | |
| Succinivibrionaceae (0.32, 0.038) | Prevotella sp. (−0.42, 0.005) | |
| Ruminococcaceae (−0.33, 0.028) | ||
| Acidaminococcaceae (−0.30, 0.049) | ||
| Oscillospiraceae (−0.31, 0.045) | ||
| Ki67+ | Lactobacillaceae (−0.42, 0.042) | Lactobacillus mucosae (−0.60, 0.002) |
| Ruminococcaceae (0.48, 0.018) | Lactobacillus delbrueckii (−0.51, 0.011) | |
| Oxalobacteraceae (−0.52, 0.009) | Phascolarctobacterium succinatutens (0.41, 0.044) |
| Item | Phase 1 | Phase 2 |
|---|---|---|
| Ingredient, % | ||
| Corn, yellow dent | 40.71 | 54.80 |
| Soybean meal, 48% CP | 22.00 | 23.50 |
| Whey permeate | 20.00 | 10.00 |
| Blood plasma | 6.00 | 3.00 |
| Poultry meal | 5.00 | 4.00 |
| Poultry fat | 3.50 | 1.80 |
| L-Lys HCl | 0.48 | 0.46 |
| DL-Met | 0.22 | 0.18 |
| L-Thr | 0.16 | 0.14 |
| Dicalcium phosphate | 0.25 | 0.68 |
| Limestone | 1.28 | 1.04 |
| Vitamin premix 1 | 0.03 | 0.03 |
| Mineral premix 2 | 0.15 | 0.15 |
| Salt | 0.22 | 0.22 |
| Calculated composition: | ||
| Dry matter, % | 91.5 | 90.4 |
| ME, kcal/kg | 3488 | 3398 |
| Crude protein, % | 23.2 | 21.7 |
| SID 3 Lys, % | 1.50 | 1.35 |
| SID Met + Cys, % | 0.82 | 0.74 |
| SID Trp, % | 0.25 | 0.22 |
| SID Thr, % | 0.88 | 0.79 |
| Ca, % | 0.85 | 0.80 |
| STTD 4 P, % | 0.45 | 0.40 |
| Total P, % | 0.65 | 0.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte, M.E.; Kim, S.W. Significance of Mucosa-Associated Microbiota and Its Impacts on Intestinal Health of Pigs Challenged with F18+ E. coli. Pathogens 2022, 11, 589. https://doi.org/10.3390/pathogens11050589
Duarte ME, Kim SW. Significance of Mucosa-Associated Microbiota and Its Impacts on Intestinal Health of Pigs Challenged with F18+ E. coli. Pathogens. 2022; 11(5):589. https://doi.org/10.3390/pathogens11050589
Chicago/Turabian StyleDuarte, Marcos Elias, and Sung Woo Kim. 2022. "Significance of Mucosa-Associated Microbiota and Its Impacts on Intestinal Health of Pigs Challenged with F18+ E. coli" Pathogens 11, no. 5: 589. https://doi.org/10.3390/pathogens11050589